

Abstract
Aging is associated with an increased production of free radicals and alterations in the mechanisms of adaptation to stress. A number of studies have shown a causal connection between age-related oxidant/antioxidant imbalance and the diminution of an organism’s melatonin levels in old age. Restoration of this rhythm may contribute to the re-stabilization of cellular homeostasis. The present work was aimed at examining the effect of the administration of melatonin or its precursor, the amino acid tryptophan, on heterophil lipid peroxidation (LPO) levels and superoxide dismutase (SOD) activity in heterophils from young (4–5 year) and old (12–14 year) ringdoves (Streptopelia risoria) under both basal and antigen-induced (stressful) conditions. Young animals were treated for 3 consecutive days with a single daily oral dose (0.25 mg) of melatonin at 1900 hours while old animals were treated for 3 consecutive days with either an oral dose (2.5 mg) of melatonin at 1900 hours or an oral capsule of 300 mg/kg body weight of l-tryptophan at 0900 hours. Non-treated old animals presented lower circulating levels of melatonin at night and higher levels of malonaldehyde (MDA) both day and night when compared to young birds. In both age groups, LPO levels were lower at night than during the day. Melatonin or tryptophan treatments significantly increased serum melatonin levels, reinforced SOD activity, and reduced MDA levels induced by the antigen. Melatonin and tryptophan may be useful agents for the treatment of disease states and processes in which an excessive production of oxidative damage occurs.
Keywords: Melatonin, Tryptophan, Lipid peroxidation, Superoxide dismutase, Heterophil, Aging
Introduction
The free radical theory of aging states that the organismal deterioration that occurs as a result of increasing longevity is specifically a consequence of the persistent accumulation of free radical-mediated damage to essential molecules, which gradually compromises the function of cells, tissues and, eventually, of the organism itself (Reiter et al. 2008a). Consequently, aging may be viewed as a process of irreversible injuries associated with accumulated oxidative debris.
To minimize the deleterious action of reactive species, cells are equipped with a series of antioxidative enzymes that metabolize toxic reactants to less reactive or totally innocuous molecules. The superoxide dismutases (SODs) are among these antioxidative enzymes. However, this protective machinery seems to be impaired with aging. In particular, SOD activity has been shown to decrease in aged individuals (Bolzán et al. 1997; Inal et al. 2001; Kedziora-Kornatowska et al. 2007), although this finding remains disputed (Kasapoglu and Özben 2001). On the other hand, it has been widely demonstrated that there is an age-associated increase in the steady-state concentrations of lipid peroxidation (LPO) products (Praticò 2002), probably due to the failure of antioxidative mechanisms or an imbalance between these and oxidation processes in advanced age.
During phagocytosis, a series of oxygen-dependent biochemical events take place, leading ultimately to the production of highly reactive oxidants, which play a key role in the microbicidal activity of phagocytes. Oxidative stress generated during phagocytosis provides protection against micro-organisms, but may cause tissue damage if it occurs to an excessive degree and/or the antioxidant mechanisms do not function properly, as has been described for aged individuals (Terrón et al. 2001).
Melatonin and its metabolites have direct scavenging actions against free radicals and related products (Peyrot and Ducrocq 2008; Tan et al. 2007) as well as indirect antioxidative action via its ability to stimulate antioxidant enzymes, including SOD (Kedziora-Kornatowska et al. 2007; Reiter et al. 2008b). Additionally, melatonin neutralizes superoxide anion radicals derived from phagocytic function (Paredes et al. 2007a, b; Terrón et al. 2004, 2005a). The amplitude of the night time melatonin peak decreases dramatically with age (Karasek and Reiter 2002; Touitou 2001). Given melatonin’s substantial antioxidant activity, this reduction may be possibly linked to the exacerbated free radical damage accumulated during aging (Reiter et al. 1999). Thus, the exogenous administration of the pineal indole or related molecules including its precursor, the amino acid tryptophan, may have positive effects in delaying free radical damage and oxidative stress due to age.
In the ringdove (Streptopelia risoria), aging provokes a significant decrease in the amplitude and circulating levels of the melatonin rhythm (Paredes et al. 2006). This is accompanied by significantly lowered circulating serotonin levels, impairments in the activity/rest and core and peripheral temperature rhythms (Paredes et al. 2006, 2007c, d; 2009a), as well as alterations in heterophil function (Paredes et al. 2007a, b) and elevated cytokine serum levels (Paredes et al. 2009a).
Since exogenous administration of both melatonin as well as its precursor, the amino acid tryptophan, reportedly restores the afore-mentioned rhythms in the aged ringdove (Paredes et al. 2007e, 2009b), and considering that aging may also be a consequence of the loss of resistance or adaptability to stress, the aim of the present work was to examine the effect of administration of melatonin or tryptophan on LPO and SOD activity in heterophils from young and old ringdoves in both basal and antigen-induced (stressful) conditions.
Materials and methods
Animals
Male and female ringdoves (Streptopelia risoria) of 4–5 years of age (young) and 12–14 years of age (old; average life span of 15 years) weighing 150 ± 20 g were used in the study (n = 10, per age group). The animals were bred in our department and individually housed in cages under controlled environmental conditions (22°C; 70% humidity), kept under a 12/12 h light/dark photoperiod (darkness from 2000 to 0800 hours), and fed ad libitum (food and water).
The study was approved by the Ethical Committee of the University of Extremadura (Badajoz, Spain) in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals.
Animal treatment
Various doses and patterns of administration of melatonin and tryptophan were tested in previously published reports (Paredes et al. 2007c, d), which were based on even earlier studies of administration of these molecules (Herichova et al. 1998; Huether et al. 1992; Terron et al. 2002). The results of these latter studies indicated that doses within this range induce significant changes in circulating melatonin levels. Thus, it was observed that a single daily oral dose of 0.25 or 2.5 mg melatonin (Sigma, St. Louis, MO) kg body weight (b.w.)−1 0.1 mL phosphate buffered saline−1 (PBS) per animal per day to young or old animals, respectively, 1 h before lights off (1900 hours), or a single oral capsule of 300 mg/kg b.w. l-tryptophan (Sigma) given to old animals at 0900 hours (1 h after the onset of the light period) restored the amplitude of the serum melatonin rhythm in old doves to that of young birds and improved the nocturnal rest in both age groups (Paredes et al. 2007c, d). The treatments also enhanced phagocytosis and free-radical scavenging activity in both young and old animals as well as correcting the age-related altered secretion of cytokines and thermoregulatory responses (Paredes et al. 2007a, b; 2009b). Birds were therefore treated in the present research with the afore-mentioned doses and patterns of administration for 3 consecutive days. Vehicle-treated birds received only 0.1 mL PBS or capsules containing the excipient—methylcellulose (Sigma)—with the same schedule as the melatonin- or tryptophan-treated animals, respectively. Non-treated values were obtained before treatment from animals that had not been given PBS, methylcellulose, melatonin, or tryptophan. As no significant variations were observed in these values with respect to the vehicle, the latter are not reported in the Results.
Serum collection
Blood samples were drawn from all ten birds at acrophases (times at which the variable reached its maximum) and nadirs (times at which the variable reached its minimum) of the melatonin rhythm, allowing at least 1 week between consecutive extractions. Based on previous research, the acrophases of the melatonin rhythm in the basal groups were established at 0200 hours and 0100 hours, and the nadirs at 1400 and 1300 hours, in young and old ringdoves, respectively (Paredes et al. 2006). In the melatonin-treated birds, the acrophases and nadirs were established at 0230 hours and 1430 hours, and 0300 hours and 1500 hours in young and old animals, respectively (Paredes et al. 2007d). In the tryptophan-treated animals, the acrophase and nadir were established at 0330 and 1530 hours, respectively (Paredes et al. 2007c). The collections (1 mL per animal per week) were taken from the brachial vein with a 25-gauge needle and a syringe, and then transferred unheparinized to a pre-prepared tube containing serum-separating gel. The samples were centrifuged at room temperature for 15 min at 300 g. The serum was then divided into aliquots in Eppendorf vials, and kept frozen at −30°C until the time of assay.
Measurement of melatonin in serum
Melatonin was determined by means of a commercial radioimmunoassay kit (IBL, Hamburg, Germany) that consisted of 125I-melatonin (140 kBq), assay buffer, enzyme, enzyme buffer, melatonin standards, rabbit anti-melatonin antiserum, precipitating agent and controls (lyophilized serum samples), according to the manufacturers instructions. Determinations were carried out in duplicate. Results are expressed in picograms per milliliter.
Isolation of heterophil leukocytes
Heterophil leukocytes were obtained immediately after the extraction of 1 mL blood (at the acrophase and nadir of the respective melatonin rhythm) from the brachial vein, to which 0.5 mL PBS and 0.5 mL lithium heparin were added, followed by centrifugation at 600 g for 15 min in a gradient using Histopaque (1 mL 1119, 1 mL 1077; Sigma). The heterophils were then washed in PBS and adjusted to 5 × 106 cells/mL medium. The cell precipitate was separated into aliquots of 300 μL per tube. Samples were divided into three groups: basal (containing only cells), control (cells incubated in a thermal bath at 37°C for 45 min), and control + latex [cells incubated for 45 min in a thermal bath at 37°C with 1.091 μm diameter latex beads (Sigma) at 1% in PBS].
Measurement of SOD activity
SOD levels were estimated using the Bioxytech® SOD-525™ assay kit (OxisInternational, Foster City, CA), following standard procedures (Rodríguez et al. 1998). The technique is based on the SOD-mediated increase in the rate of autoxidation of 5,6,6a,11b-tetrahydro-3,9,10-trihydroxybenzo[c]fluorene R1 in aqueous alkaline solution to yield a chromophore with maximum absorbance at 525 nm. SOD activity is determined from the ratio of the autoxidation rates in the presence (Vs) and in the absence (Vc) of SOD. The relationship between the Vs/Vc ratio and SOD activity is described by the equation 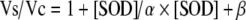 , where Vs = rate of sample containing SOD; Vc = average rate of blank (SOD = 0) samples; SOD = the SOD activity of the sample in SOD-525 units; α = dimensionless coefficient; β = coefficient in SOD-525 units. The resulting value is multiplied by the dilution factor and expressed as units of activity per milliliter.
, where Vs = rate of sample containing SOD; Vc = average rate of blank (SOD = 0) samples; SOD = the SOD activity of the sample in SOD-525 units; α = dimensionless coefficient; β = coefficient in SOD-525 units. The resulting value is multiplied by the dilution factor and expressed as units of activity per milliliter.
Measurement of LPO
LPO was measured using the LPO-586™ kit by Bioxytech® (OxisInternational), according to a previously described technique (Rodriguez et al. 1999; Terrón et al. 2005b). This assay quantifies the malondialdehyde (MDA) levels present in a sample. MDA is an end product of membrane LPO caused by free radicals and is one of the most widely used markers for in vivo free radical oxidation processes. The equation 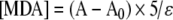 gives the concentration of MDA in a sample, where A is the absorbance in the presence of sample, A0 is the absorbance in the absence of sample, 5 is the sample dilution factor in the cuvette (200 μL sample in a total volume of 1 mL), and ε is the apparent molar extraction coefficient obtained from the standard curve using a standard (solution of 10 mM 1,1,3,3-tetramethoxypropane in 20 mM Tris-HCl buffer, pH 7.4, 0–20 μM) diluted 100 times with PBS. Since results are expressed as nmol MDA/mg protein, the levels of cell lysate total protein were measured using the Bradford protein assay (Sigma). The concentration of total protein was calculated by means of a standard curve with bovine albumin (Sigma) and the results expressed as milligrams of protein per milliliter.
gives the concentration of MDA in a sample, where A is the absorbance in the presence of sample, A0 is the absorbance in the absence of sample, 5 is the sample dilution factor in the cuvette (200 μL sample in a total volume of 1 mL), and ε is the apparent molar extraction coefficient obtained from the standard curve using a standard (solution of 10 mM 1,1,3,3-tetramethoxypropane in 20 mM Tris-HCl buffer, pH 7.4, 0–20 μM) diluted 100 times with PBS. Since results are expressed as nmol MDA/mg protein, the levels of cell lysate total protein were measured using the Bradford protein assay (Sigma). The concentration of total protein was calculated by means of a standard curve with bovine albumin (Sigma) and the results expressed as milligrams of protein per milliliter.
Statistical analysis
Data are expressed as mean ± SD of the number of determinations. The results were analysed by using Friedman ranges for paired samples, followed by Kruskal-Wallis multiple contrasts. Only values with P < 0.05 were accepted as significant.
Results
In all groups (non-treated, melatonin-treated, and tryptophan-treated), the melatonin values obtained at the nadir were significantly lower than those of the acrophase (P < 0.05; Fig. 1). Serum melatonin levels at the acrophase in non-treated young animals were significantly higher compared with values measured in old birds (P < 0.05). The administration of melatonin caused a significant rise (P < 0.05) in the circulating levels of the indoleamine in the nocturnal period in both age groups, causing the age difference in the melatonin values found between non-treated young and old groups to disappear. Tryptophan treatment also significantly elevated (P < 0.05) melatonin levels at both the diurnal and nocturnal periods as compared to their corresponding values in non-treated animals.
Fig. 1.

Serum levels of melatonin at the corresponding hours of acrophase and nadir in young and old ringdoves, either non-treated or after 3 days of treatment 1 h before lights out with 0.25 mg/kg b.w. and 2.5 mg/kg body weight (b.w.) melatonin, or after 3 days of treatment 1 h after lights on with 300 mg/kg b.w. tryptophan in old ringdoves. Each value represents the mean ± standard deviation of ten determinations performed in duplicate. Lower case letters indicate P < 0.05: a vs values obtained in the diurnal period, b vs values obtained in the corresponding non-treated groups, c vs values obtained in young animals
In non-treated young and old animals, there was a significant decrease (P < 0.05) in nocturnal LPO levels with respect to the values obtained during the day, under both basal and control conditions (Fig. 2a). This was also the case in the melatonin-treated young and old basal and control groups (Fig. 2b). In tryptophan-treated individuals, a similar pattern was observed, i.e., higher diurnal levels of MDA compared to those measured at night although the difference was non-significant (Fig. 2c). The LPO levels reached in old animals were significantly higher (P < 0.05) than those obtained in the basal and control young groups. Interestingly, in melatonin-treated and tryptophan-treated animals, this rise in MDA values in old animals compared to young was not found.
Fig. 2.

Levels of lipid peroxidase [LPO; nmol malondialdehyde (MDA)/mg protein] in heterophils from young and old ringdoves in basal, control and antigen-induced conditions in non-treated animals (a), and after 3 days of treatment 1 h before lights out with 0.25 mg/kg b.w. and 2.5 mg/kg b.w. melatonin in young and old ringdoves, respectively (b); or after 3 days of treatment 1 h after lights on with 300 mg/kg b.w. tryptophan in old ringdoves (c). Each value represents the mean ± standard deviation of six determinations performed in duplicate. Lower case letters indicate P < 0.05: a vs values obtained in the diurnal period, b vs values obtained in their respective basal groups, c vs values obtained in the corresponding control groups, d vs values obtained in young animals, e vs values obtained in the corresponding non-treated groups
Heterophil incubation with latex beads provoked a significant elevation (P < 0.05) in the LPO of both non-treated young and old animals with respect to the values obtained in the basal and control groups, with the elevation being higher (P < 0.05) in old birds. The antigen-induced LPO rise was observed during the day as well as at night. However, nocturnal MDA levels were significantly lower (P < 0.05) when compared with diurnal values. Similarly to non-treated animals, antigen-induced LPO was greater (P < 0.05) during the day than in the nocturnal period in the melatonin- and tryptophan-treated young and old groups. Nevertheless, the values obtained in both groups of old birds were significantly reduced (P < 0.05) when compared to the corresponding values in the non-treated animals.
SOD activity in non-treated young individuals (Fig. 3a) exhibited a significant increase (P < 0.05) at night under both basal and control conditions. The incubation with latex beads augmented the enzymatic function significantly (P < 0.05) with respect to the basal and control groups. In non-treated old animals, the afore-mentioned difference between diurnal and nocturnal values was not observed. In addition, in latex-challenged old cells obtained at night, SOD activity was reduced significantly (P < 0.05) with respect to the levels reached in young antigen-induced cells.
Fig. 3.

Levels of superoxide dismutase (SOD) activity in heterophils from young and old ringdoves in basal, control and antigen-induced conditions in non-treated animals (a), and after 3 days of treatment 1 h before lights off with 0.25 mg/kg b.w. and 2.5 mg/kg b.w. of melatonin in young and old ringdoves, respectively (b), or after 3 days of treatment 1 h after lights on with 300 mg/kg b.w. tryptophan in old ringdoves (c). Each value represents the mean ± standard deviation of six determinations performed in duplicate. Lower case letters indicate P < 0.05: a vs values obtained in the diurnal period, b vs values obtained in their respective basal groups, c vs values obtained in the corresponding control groups, d vs values obtained in young animals, e vs values obtained in the corresponding non-treated groups
Treatment with either melatonin or tryptophan caused a significant augmentation (P < 0.05) of SOD activity in both age groups at most basal and control conditions with respect to the values obtained in the non-treated groups (Fig. 3b,c). In most latex-induced melatonin and tryptophan-treated groups, SOD activity increased significantly (P < 0.05) with respect to values reached under basal conditions. However, SOD activity measured in young melatonin-treated animals was significantly reduced (P < 0.05) with respect to the corresponding non-treated values in both the diurnal and nocturnal periods.
Discussion
The age-related decline in both circulating levels and the amplitude of the rhythm of melatonin are well-documented phenomena in a variety of species (Lahiri et al. 2004; Pang et al. 1990), including the ringdove (Paredes et al. 2006, 2007c, d). The loss of this potent anti-oxidant during aging may be, in part, the cause of the onset of the age-related imbalance between prooxidants and antioxidants. In fact, there are reports that confirm that exogenous administration of this indole or its precursor, the amino acid tryptophan, reduces the inflammatory and oxidative processes associated with age (Paredes et al. 2009a; Rodriguez et al. 2007).
In the present study, we found a significant reduction in circulating melatonin levels during the nocturnal period compared to values observed in young birds. The diminution of the melatonin rhythm found in old birds could have serious implications in terms of antioxidant defense, taking into account the protective action of this molecule against reactive oxygen and nitrogen species (Reiter et al. 2008b). In this regard, we found that LPO values were higher during the day than at night, and that they were significantly raised when the cells were incubated in the presence of latex beads. The administration of melatonin significantly elevated the nocturnal circulating melatonin in young and old animals and resulted in a general decline of the LPO level, as observed after incubation with latex beads in both groups of age. Similar results were obtained in old animals treated with tryptophan, but in this case both the diurnal and nocturnal serum levels of the indole increased. This is in agreement with previous studies on ring dove heterophils, where it was observed that in vitro incubation with melatonin suppressed both basal and antigen-induced LPO (Rodriguez et al. 1999). Also, incubating heterophils from old animals with physiological concentrations of melatonin in young animals provoked a general reduction of MDA levels (Terrón et al. 2005b). In this context, in melatonin-treated old animals, where the circulating levels of the indolamine were significantly elevated, the LPO levels found under basal and control conditions were similar to those obtained in young doves, contrary to the case in non-treated animals, where old birds had higher MDA values than the young. Other studies have also shown geroprotective effects of this pineal indole on LPO (Kireev et al. 2008; Rodriguez et al. 2007; Somova et al. 2001), probably due to the pronounced antioxidant activity of this molecule and its metabolites (Peyrot and Ducrocq 2008; Reiter et al. 2008b; Tan et al. 2007). Melatonin, administered directly or via conversion from its precursor (the amino acid tryptophan), may thus act as an effective protector against age-related and/or latex-bead induced LPO toxicity.
It has been shown that elevated free radical production increases the expression and/or activity of antioxidative enzymes, including SOD (Warner et al. 1996). Melatonin has also been described as a regulator of this antioxidative enzyme (Rodriguez et al. 2004). In fact, it has been suggested that the circadian rhythm of melatonin may influence the endogenous rhythm that SOD exhibits under normal light/dark conditions, with generally higher levels of both enzyme activity and mRNA gene expression during the nocturnal period (Albarrán et al. 2001; Martín et al. 2003). In our study, an elevation of SOD levels was found in cells from non-treated young animals incubated in the presence of latex beads. This elevation was higher at night, which may reflect an additive effect of both free radical production and melatonin on the activity of the enzyme. Additionally, in the non-treated young basal and control groups, SOD levels were higher at night than during the day. However, in non-treated old animals, this day/night pattern was not found. Also, the elevation of SOD activity after latex bead exposure was not as pronounced as that obtained in young birds. One reason for this could be the SOD reduction observed in aged individuals (Bolzán et al. 1997; Inal et al. 2001; Kedziora-Kornatowska et al. 2007), although other causes should not be ruled out. For example, cytokines such as IL-1α, IL-1β, IL-6, or TNF-α are also implicated in the SOD regulation: higher than average levels of Mn-SOD expression were found after administration of cytokines, while Cu-Zn SOD levels remained unchanged or even decreased (Isoherranen et al. 1997; Kifle et al. 1996; Martín et al. 2003; Sugino et al. 1998). Since elevated production of IL-1β, IL-6, or TNF-α has been reported in old ringdoves (Paredes et al. 2009a), probably due to the fact that these cytokines are pro-inflammatory and age is associated with increased inflammation processes (Sergio 2008), further investigation is required to elucidate the role of these or other regulating molecules on the impaired enzymatic response to ageing and the antigen-induced activation found in our animal model. Interestingly, SOD activity was augmented in both basal and control tryptophan and melatonin-treated animals, but this elevation was significantly less than that obtained in non-treated birds when heterophils from these groups were incubated with latex-beads. The reason may lie in the powerful antioxidant and free-radical scavenging actions that both the indole and its precursor exert in phagocytic cells from birds and mammals, including the neutralization of the superoxide anion, the substrate of SOD (Paredes et al. 2007a, b; Sánchez et al. 2008; Terrón et al. 2004).
In summary, we found an age-related decline in the melatonin rhythm that was accompanied by increased levels of LPO and altered responses of SOD to antigen-induced activation. The replacement of the melatonin rhythm via the administration of exogenous melatonin or the amino acid tryptophan increased circulating levels of the indoleamine in old birds as well as reducing LPO generation and upregulating SOD activity. Similar results were obtained when young animals were treated with melatonin. Melatonin and tryptophan may thus be useful agents for the treatment of disease states and processes where elevated lipid oxidation and free radical damage have been reported.
Acknowledgments
This investigation was supported by a research grant from Consejería de Infraestructuras y Desarrollo Tecnológico (Junta de Extremadura, 3PR05A053). S.D.P., I.B. and M.P.T. were beneficiaries of grants from Consejería de Economía, Comercio e Innovación – Fondo Social Europeo (Junta de Extremadura, POS07012, PRE06070, and REI07004, respectively). The authors would like to express their thanks to Elena Circujano Vadillo, for technical assistance, and to Professors Manuel Molina and Manuel Mota, members of the Department of Mathematics (Section of Statistics and Operations Research) of the Faculty of Science, for their advice and collaboration in the statistical analysis.
References
- Albarrán MT, López-Burillo S, Pablos MI, Reiter RJ, Agapito MT (2001) Endogenous rhythms of melatonin, total antioxidant status and superoxide dismutase activity in several tissues of chick and their inhibition by light. J Pineal Res 30:227–233. doi:10.1034/j.1600-079X.2001.300406.x [DOI] [PubMed]
- Bolzán AD, Bianchi MS, Bianchi NO (1997) Superoxide dismutase, catalase and glutathione peroxidase activities in human blood: influence of sex, age and cigarette smoking. Clin Biochem 30:449–454. doi:10.1016/S0009-9120(97)00047-7 [DOI] [PubMed]
- Herichova I, Zeman M, Veselovsky J (1998) Effect of tryptophan administration on melatonin concentrations in the pineal gland, plasma and gastrointestinal tract of chickens. Acta Vet (Brno) 67:89–95
- Huether G, Poeggeler B, Reimer A, George A (1992) Effect of tryptophan administration on circulating melatonin levels in chicks and rats: evidence for stimulation of melatonin synthesis and release in the gastrointestinal tract. Life Sci 51:945–953. doi:10.1016/0024-3205(92)90402-B [DOI] [PubMed]
- Inal ME, Kaubak G, Sunal E (2001) Antioxidant enzyme activities and malondialdehyde levels related to aging. Clin Chim Acta 305:75–80. doi:10.1016/S0009-8981(00)00422-8 [DOI] [PubMed]
- Isoherranen K, Peltola V, Laurikainen L, Punnonen J, Laihia J, Ahotupa M, Punnonen K (1997) Regulation of copper/zinc and manganese dismutase by UVB irridation, oxidative stress and cytokines. J Photochem Photobiol B 40:288 –293 [DOI] [PubMed]
- Karasek M, Reiter RJ (2002) Melatonin and aging. Neuro Endocrinol Lett 23:14–16 [PubMed]
- Kasapoglu M, Özben T (2001) Alterations of antioxidant enzymes and oxidative stress markers in aging. Exp Gerontol 36:209–220. doi:10.1016/S0531-5565(00)00198-4 [DOI] [PubMed]
- Kedziora-Kornatowska K, Szewczyk-Golec K, Czuczejko J, van Marke de Lumen K, Pawluk H, Motyl J, Karasek M, Kedziora J (2007) Effect of melatonin on the oxidative stress in erythrocytes of healthy young and elderly subjects. J Pineal Res 42:153–158. doi:10.1111/j.1600-079X.2006.00394.x [DOI] [PubMed]
- Kifle Y, Monnier J, Chesrown SE, Raizada MK, Nick HS (1996) Regulation of the manganese superoxide dismutase and inducible nitric oxide synthase gene in rat neuronal and glial cells. J Neurochem 66:2128–2135 [DOI] [PubMed]
- Kireev RA, Tresguerres AC, Garcia C, Ariznavarreta C, Vara E, Tresguerres JA (2008) Melatonin is able to prevent the liver of old castrated female rats from oxidative and pro-inflammatory damage. J Pineal Res 45:394–402. doi:10.1111/j.1600-079X.2008.00606.x [DOI] [PubMed]
- Lahiri DK, Ge YW, Sharman EH, Bondy SC (2004) Age-related changes in serum melatonin in mice: higher levels of combined melatonin and 6-hydroxymelatonin sulfate in the cerebral cortex than serum, heart, liver and kidney tissues. J Pineal Res 36:217–223. doi:10.1111/j.1600-079X.2004.00120.x [DOI] [PubMed]
- Martín V, Sainz RM, Mayo JC, Antolín I, Herrera F, Rodríguez C (2003) Daily rhythm of gene expression in rat superoxide dismutases. Endocr Res 29:83–95. doi:10.1081/ERC-120018679 [DOI] [PubMed]
- Pang SF, Tsang CW, Hong GX, Yip PC, Tang PL, Brown GM (1990) Fluctuation of blood melatonin concentrations with age: result of changes in pineal melatonin secretion, body growth, and aging. J Pineal Res 8:179–192. doi:10.1111/j.1600-079X.1990.tb00678.x [DOI] [PubMed]
- Paredes SD, Terrón MP, Cubero J, Valero V, Barriga C, Reiter RJ, Rodríguez AB (2006) Comparative study of the activity/rest rhythms in young and old ringdove (Streptopelia risoria): correlation with serum levels of melatonin and serotonin. Chronobiol Int 23:779–793. doi:10.1080/07420520600827145 [DOI] [PubMed]
- Paredes SD, Terrón MP, Marchena AM, Barriga C, Pariente JA, Reiter RJ, Rodríguez AB (2007a) Effect of exogenous melatonin on viability, ingestion capacity, and free-radical scavenging in heterophils from young and old ringdoves (Streptopelia risoria). Mol Cell Biochem 304:305–314. doi:10.1007/s11010-007-9513-7 [DOI] [PubMed]
- Paredes SD, Terrón MP, Marchena AM, Barriga C, Pariente JA, Reiter RJ, Rodríguez AB (2007b) Tryptophan modulates cell viability, phagocytosis and oxidative metabolism in old ringdoves. Basic Clin Pharmacol Toxicol 101:56–62. doi:10.1111/j.1742-7843.2007.00076.x [DOI] [PubMed]
- Paredes SD, Terrón MP, Cubero J, Valero V, Barriga C, Reiter RJ, Rodríguez AB (2007c) Tryptophan increases nocturnal rest and affects melatonin and serotonin serum levels in old ringdove. Physiol Behav 90:576–582. doi:10.1016/j.physbeh.2006.11.007 [DOI] [PubMed]
- Paredes SD, Terrón MP, Valero V, Barriga C, Reiter RJ, Rodríguez AB (2007d) Orally administered melatonin improves nocturnal rest in young and old ringdoves (Streptopelia risoria). Basic Clin Pharmacol Toxicol 100:258–268. doi:10.1111/j.1742-7843.2006.00032.x [DOI] [PubMed]
- Paredes SD, Barriga C, Rodríguez AB (2007e) Altered circadian rhythms of corticosterone, melatonin, and phagocytic activity in response to stress in rats. Neuro Endocrinol Lett 28:757–760 [PubMed]
- Paredes SD, Marchena AM, Bejarano I, Espino J, Barriga C, Rial RV, Reiter RJ (2009a) Melatonin and tryptophan affect the activity-rest rhythm, core and peripheral temperatures, and interleukin levels in the ringdove: Changes with age. J Gerontol A Biol Sci Med Sci 63:340–350. doi:10.1093/gerona/gln054 [DOI] [PMC free article] [PubMed]
- Paredes SD, Barriga C, Reiter RJ, Rodríguez AB (2009b) Assessment of the potential role of tryptophan as the precursor of serotonin and melatonin for the aged sleep-wake cycle and immune function: Streptopelia risoria as a model. Int J Tryptophan Res 2:23–36 [DOI] [PMC free article] [PubMed]
- Peyrot F, Ducrocq C (2008) Potential role of tryptophan derivatives in stress responses characterized by the generation of reactive oxygen and nitrogen species. J Pineal Res 45:235–246. doi:10.1111/j.1600-079X.2008.00580.x [DOI] [PubMed]
- Praticò D (2002) Lipid peroxidation and the aging process. Sci Aging Knowl Environ 50:re5. doi:10.1126/sageke.2002.50.re5 [DOI] [PubMed]
- Reiter RJ, Tan D, Kim SJ, Manchester LC, Qi W, Garcia JJ, Cabrera JC, El-Sokkary G, Rouvier-Garay V (1999) Augmentation of indices of oxidative damage in life-long melatonin-deficient rats. Mech Ageing Dev 110:157–173. doi:10.1016/S0047-6374(99)00058-5 [DOI] [PubMed]
- Reiter RJ, Paredes SD, Korkmaz A, Manchester LC, Tan DX (2008a) Melatonin in relation to the "strong" and "weak" versions of the free radical theory of aging. Adv Med Sci 53:119–129. doi:10.2478/v10039-008-0032-x [DOI] [PubMed]
- Reiter RJ, Tan DX, Jou MJ, Korkmaz A, Manchester LC, Paredes SD (2008b) Biogenic amines in the reduction of oxidative stress: melatonin and its metabolites. Neuro Endocrinol Lett 29:391–398 [PubMed]
- Rodríguez AB, Nogales G, Ortega E, Barriga C (1998) Melatonin controls superoxide anion level: modulation of superoxide dismutase activity in ring dove heterophils. J Pineal Res 24:9–14. doi:10.1111/j.1600-079X.1998.tb00360.x [DOI] [PubMed]
- Rodriguez AB, Nogales G, Marchena JM, Ortega E, Barriga C (1999) Suppression of both basal and antigen-induced lipid peroxidation in ring dove heterophils by melatonin. Biochem Pharmacol 58:1301–1306. doi:10.1016/S0006-2952(99)00207-5 [DOI] [PubMed]
- Rodríguez C, Mayo JC, Sainz RM, Antolín I, Herrera F, Martín V, Reiter RJ (2004) Regulation of antioxidant enzymes: a significant role for melatonin. J Pineal Res 36:1–9. doi:10.1046/j.1600-079X.2003.00092.x [DOI] [PubMed]
- Rodriguez MI, Escames G, López LC, García JA, Ortiz F, López A, Acuña-Castroviejo D (2007) Melatonin administration prevents cardiac and diaphragmatic mitochondrial oxidative damage in senescence-accelerated mice. J Endocrinol 194:637–643. doi:10.1677/JOE-07-0260 [DOI] [PubMed]
- Sánchez S, Paredes SD, Sánchez CL, Barriga C, Reiter RJ, Rodríguez AB (2008) Tryptophan administration in rats enhances phagocytic function and reduces oxidative metabolism. Neuro Endocrinol Lett 29:1026–1032 [PubMed]
- Sergio G (2008) Exploring the complex relations between inflammation and aging (inflamm-aging): anti-inflamm-aging remodelling of inflamm-aging, from robustness to frailty. Inflamm Res 57:558–563. doi:10.1007/s00011-008-7243-2 [DOI] [PubMed]
- Somova EV, Kolodub FA, Bondarenko LA (2001) Chronobiological aspects of melatonin-produced antioxidant effects in senescent rats. Bull Exp Biol Med 132:887–889. doi:10.1023/A:1013187205570 [DOI] [PubMed]
- Sugino N, Telleria CM, Gibori G (1998) Differential regulation of copper-zinc superoxide dismutase and manganese superoxide dismutase in the rat corpus luteum: induction of manganese superoxide dismutase messenger ribonucleic acid by inflammatory cytokines. Biol Reprod 59:208–215 [DOI] [PubMed]
- Tan DX, Manchester LC, Terron MP, Flores LJ, Reiter RJ (2007) One molecule, many derivatives: a never-ending interaction of melatonin with reactive oxygen and nitrogen species? J Pineal Res 42:28–42. doi:10.1111/j.1600-079X.2006.00407.x [DOI] [PubMed]
- Terrón MP, Marchena JM, Shadi F, Harvey S, Lea RW, Rodríguez AB (2001) Melatonin: an antioxidant at physiological concentrations. J Pineal Res 31:95–96. doi:10.1034/j.1600-079X.2001.310114.x [DOI] [PubMed]
- Terron MP, Cubero J, Marchena JM, Barriga C, Rodriguez AB (2002) Melatonin and aging: in vitro effect of young and mature ring dove physiological concentrations of melatonin on the phagocytic function of heterophils from old ring dove. Exp Gerontol 37:421–426. doi:10.1016/S0531-5565(01)00209-1 [DOI] [PubMed]
- Terrón MP, Paredes SD, Barriga C, Ortega E, Rodríguez AB (2004) Comparative study of the heterophil phagocytic function in young and old ring doves (Streptopelia risoria) and its relationship with melatonin levels. J Comp Physiol [B] 174:421–427. doi:10.1007/s00360-004-0429-1 [DOI] [PubMed]
- Terrón del MP, Paredes SD, Barriga C, Ortega E, Reiter RJ, Rodríguez AB (2005a) Oral administration of melatonin to old ring doves (Streptopelia risoria) increases plasma levels of melatonin and heterophil phagocytic activity. J Gerontol A Biol Sci Med Sci 60:44–50 [DOI] [PubMed]
- Terrón MP, Paredes SD, Barriga C, Ortega E, Reiter RJ, Rodríguez AB (2005b) Melatonin, lipid peroxidation, and age in heterophils from the ring dove (Streptopelia risoria). Free Radic Res 39:613–619. doi:10.1080/10715760500097831 [DOI] [PubMed]
- Touitou Y (2001) Human aging and melatonin. Clinical relevance. Exp Gerontol 36:1083–1100. doi:10.1016/S0531-5565(01)00120-6 [DOI] [PubMed]
- Warner BB, Stuart L, Gebb S, Wispe JR (1996) Redox regulation of manganese superoxide dismutase. Am J Physiol 271:L150–L158 [DOI] [PubMed]