

Abstract
Objective
Major depressive disorder (MDD) is characterized by impaired reward processing, possibly due to dysfunction in the basal ganglia. However, few neuroimaging studies of depression have distinguished between anticipatory and consummatory phases of reward processing. Using functional magnetic resonance imaging (fMRI) and a task that dissociates anticipatory and consummatory phases of reward processing, the authors tested the hypothesis that MDD participants would show reduced reward-related responses in basal ganglia structures.
Method
A monetary incentive delay task was presented to 30 unmedicated MDD subjects and 31 healthy comparison subjects during fMRI scanning. Whole-brain analyses focused on neural responses to reward-predicting cues and rewarding outcomes (i.e., monetary gains). Secondary analyses focused on the relationship between anhedonic symptoms and basal ganglia volumes.
Results
Relative to comparison subjects, MDD participants showed significantly weaker responses to gains in the left nucleus accumbens and bilateral caudate. Group differences in these regions were specific to rewarding outcomes and did not generalize to neutral or negative outcomes, although relatively reduced responses to monetary penalties in MDD emerged in other caudate regions. By contrast, evidence for group differences during reward anticipation was weaker, although MDD subjects showed reduced activation to reward cues in a small sector of the left posterior putamen. Among MDD subjects, anhedonic symptoms and depression severity were associated with reduced bilateral caudate volume.
Conclusions
These results indicate that basal ganglia dysfunction in MDD may affect the consummatory phase of reward processing. Additionally, morphometric results suggest that anhedonia in MDD is related to caudate volume.
Introduction
Anhedonia-lack of reactivity to pleasurable stimuli-is a core symptom of major depressive disorder (MDD) (1-2). Relative to healthy controls, depressed individuals display reduced positive attentional biases (3), weaker positive affect in response to pleasant stimuli (4), and reduced reward responsiveness (5). Neuroimaging indicates that these deficits may reflect dysfunction in the basal ganglia, including the striatum (nucleus accumbens, caudate, putamen) and globus pallidus (6-11). However, the functional significance of basal ganglia dysfunction in MDD remains poorly understood. Specifically, whether dysfunction is more closely associated with deficits in the anticipatory or consummatory phase of reward processing is unclear.
Dissociating these phases is important for two reasons (12). First, they reflect different psychological states: anticipation is characterized by goal-directed behavior, whereas consummation involves pleasure experience (13). Second, they make separable contributions to goal-directed behavior (14). In non-human primates, unexpected rewards elicit phasic bursts in dopamine neurons projecting from the midbrain to basal ganglia (14). However, the bursts eventually shift from the rewards to reward-predicting cues. Because the basal ganglia are critical for motor control (15), this constitutes a mechanism by which reward-predicting cues can elicit motivated behavior. Given dopamine abnormalities in MDD (16), depression may involve impairments in the anticipatory and/or consummatory components of this mechanism.
To address this issue, a recent study used a monetary incentive delay task to investigate anticipatory versus consummatory phases of reward processing in 14 MDD participants and 12 controls (17). Surprisingly, there were no group differences in basal ganglia responses to reward cues. Furthermore, although MDD subjects showed reduced bilateral putamen responses to gains, no outcome-related differences emerged in the accumbens or the caudate, regions implicated in processing reward feedback (18, 19), particularly when reward delivery is unpredictable (20). However, there were also no group differences in behavior. Thus, these null results may have reflected intact reward processing in that particular MDD sample and/or limited statistical power.
In the present study, we used a similar task to probe anticipatory and consummatory phases of reward processing in a larger group of unmedicated depressed individuals (N=30) and healthy controls (N=31). To permit a balanced design, the task was modified such that 50% of reward and loss trials ended in monetary gains and penalties, respectively (21). Given the role of dopamine and the basal ganglia in reward anticipation (22), we predicted that depressed individuals would show blunted responses to reward cues, particularly in the ventral striatum. However, based on prior findings (17), and because gains were only delivered on 50% of reward trials (20), we hypothesized that MDD subjects might primarily show impaired striatal responses to rewarding outcomes. Finally, in light of recent work (23), we predicted that greater anhedonic symptoms would be associated with smaller caudate volume.
Methods
Participants
Depressed subjects were recruited from a treatment study comparing the effectiveness of the dietary supplement S-adenosyl l-methionine to escitalopram. Comparison subjects were recruited from the community. MDD participants had a DSM-IV diagnosis of MDD (24) and a score ≥16 on the 21-item Hamilton Depression Rating Scale (HRSD; 25). Exclusion criteria included psychotropic medication in the last 2 weeks (fluoxetine: 6 weeks; dopaminergic drugs or neuroleptics: 6 months), current or past history of MDD with psychotic features, and presence of other Axis I diagnosis (including lifetime substance dependence and substance use disorders in the last year), with the exception of anxiety disorders. Comparison subjects reported no medical or neurological illness, no current or past psychopathology (24), and no psychotropic medications. All subjects were right-handed.
The final sample included 30 MDD and 31 demographically matched comparison subjects (Table 1). MDD subjects were moderately depressed, as assessed by Beck Depression Inventory-II (BDI-II; 26) (27.48±10.60) and 17-item HRSD (17.97±4.19) scores. Eleven MDD subjects had a current anxiety disorder, and 3 had subthreshold anxiety symptoms. Among the MDD subjects, 11 (37%) had never received antidepressants and 16 (53%) reported prior antidepressant use; information about prior antidepressant treatment was unavailable for 3 individuals. Only three patients reported resistance to a prior antidepressant. All participants provided written informed consent to a protocol approved by the local IRBs.
TABLE 1.
Sociodemographic and Clinical Data in MDD (N=30) and Comparison (N=31) Subjects
| Comparison subjects | MDD subjects | |||||
|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Statistics | P value | |
| Age | 38.80 | 14.48 | 43.17 | 12.98 | t(59)=-1.36 | >0.18 |
| % Female | 42% | N/A | 50% | N/A | χ2(1)=0.39 | >0.53 |
| Education | 15.19 | 1.96 | 14.87 | 2.37 | t(59)=0.59 | >0.55 |
| Ethnicity (% Caucasian) | 77% | N/A | 67% | N/A | χ2(1)=0.73 | >0.39 |
| Marital status (% married) | 22.6% | N/A | 23.3% | N/A | χ2(1)=0.001 | >0.50 |
| Employment (% employed) | 58.1% | N/A | 40.0% | N/A | χ2(1)=1.99 | >0.15 |
| Age of MDD onset (years) | N/A | N/A | 29.39 | 15.98 | N/A | N/A |
| Length of current MDE (months) | N/A | N/A | 37.13 | 78.24 | N/A | N/A |
| Number of prior MDEs | N/A | N/A | 3.69 | 2.64 | N/A | N/A |
| BDI-II* | 2.20 | 2.41 | 27.48 | 10.60 | t(55)=12.12 | <0.001 |
| HRSD (17-item) | N/A | N/A | 17.97 | 4.9 | N/A | N/A |
Monetary Incentive Delay Task
The task has been described previously (21). Trials began with a visual cue (1.5 s) indicating the potential outcome (reward: +$; loss: -$; no-incentive: 0$). After a variable interstimulus interval (3-7.5 s), a red target square was briefly presented, to which subjects responded by pressing a button. After a second delay (4.4-8.9 s), visual feedback (1.5 s) indicated trial outcome (gain, penalty, no-change). A variable interval (3-12 s) separated the trials. The task involved five blocks with 24 trials (8/cue), yielding 40 and 20 trials for cue- and outcome-related analyses, respectively.
Participants were instructed that rapid responses maximized their chances of obtaining gains and avoiding penalties. However, gains and penalties were actually delivered in a predetermined pattern to allow a balanced design. For each block, half the reward trials yielded a monetary gain ($1.96-2.34; mean: $2.15) and half ended with no-change feedback. Similarly, half the loss trials yielded a monetary penalty (range: $1.81-2.19; mean: $2.00), and half resulted in no-change. No-incentive trials always ended with no-change feedback. To maximize feedback believability, target duration was longer for trials scheduled to be successful (e.g., gains on reward trials) than for trials scheduled to be unsuccessful (e.g., no-change on reward trials). Furthermore, target durations were individually titrated based on reaction time data collected during a practice session (Supplemental Material).
Procedure
Data collection occurred prior to treatment onset. After blocks two and four, participants rated their affective response to cues and outcomes for valence (1=most negative, 5=most positive) and arousal (1=low intensity, 5=high intensity). Participants were compensated ($80) for their time and “earned” $20-22 from the task.
Data Acquisition
Data were collected on a 1.5T Symphony/Sonata scanner (Siemens Medical Systems; Iselin, NJ) and consisted of a T1-weighted MPRAGE acquisition (TR/TE: 2730/3.39 ms; FOV: 256 mm; voxel dimensions: 1 × 1 × 1.33 mm; 128 slices) and gradient echo T2*-weighted echoplanar images, which were acquired using an optimized pulse sequence (21) (TR/TE: 2500/35ms; FOV: 200 mm; voxel: 3.125 × 3.125 × 3 mm; 35 interleaved slices).
Data Reduction and Statistics
Reaction Time and Affective Ratings
After removing outliers (responses exceeding mean±3SD), reaction time data were entered into a Group x Cue x Block ANOVA. For brevity, only effects involving Group or Cue are reported. Affective ratings were averaged across the two assessments and entered into Group x Cue or Group x Outcome ANOVAs.
Functional and Structural MRI
Analyses were conducted using FS-FAST (http://surfer.nmr.mgh.harvard.edu) and FreeSurfer (27). Pre-processing included slice-time and motion correction, removal of slow linear trends, intensity normalization, and spatial smoothing (6mm FWHM); a temporal whitening filter was used to correct for autocorrelation in the noise. Data for four MDD subjects were lost due to excessive motion (>5 mm), leaving 31 comparison and 26 MDD subjects for fMRI analysis. Prior to group analyses, data were re-sampled into MNI305 space (2 mm3 voxels).
Functional data were analyzed using the general linear model. The hemodynamic response was modeled as a gamma function and convolved with stimulus onsets; motion parameters were included as nuisance regressors. Between-group whole-brain random effects comparisons were computed for Reward Anticipation (reward cue vs. no-incentive cue) and Reward Outcome (gain vs. no-change feedback on no-incentive trials) contrasts. Note that, due to the double subtraction, clusters exceeding the statistical threshold show a significant Group x Condition interaction. Secondary analyses of loss-related contrasts are reported in the Supplemental Material. Due to a priori hypotheses about the basal ganglia, activation maps were thresholded using a peak voxel criterion of p<0.005 with a minimum cluster extent of 12 voxels; Monte Carlo simulations were performed to confirm that the primary findings held following correction for multiple comparisons (Supplemental Material). Findings emerging outside the basal ganglia should be considered preliminary. To assess whether findings in a priori regions were specific to rewards, follow-up Group x Condition ANOVAs were conducted on averaged beta weights (including for penalties) extracted from clusters showing group differences.
Structural MRI
Morphometric analyses used FreeSurfer’s automated parcellation approach (27, 28; Supplemental Material, Table S1) and focused on basal ganglia. To account for differences in cranial size, volumes were divided by the intracranial volume, and entered into a Group x Hemisphere x Region (nucleus accumbens, caudate, putamen, globus pallidus) ANOVA. Significant effects were followed-up with post-hoc t-tests. For MDD participants, Pearson correlations and hierarchical regressions (controlling for age and gender) were conducted to examine relationships between volumes and anhedonic symptoms or depression severity. As in prior work (29), anhedonia was assessed by computing an “anhedonic” BDI-II subscore (loss of pleasure, interest, energy, and libido; reliability coefficient: α=0.85).
Results
Reaction Time (RT)
A main effect of Cue emerged (F=30.15, df=2,118, p<0.0001), reflecting motivated responding (shorter RT) on reward and loss trials versus no-incentive trials. The main effect of Group was not significant (F=0.17, df=1,59, p>0.68), indicating that comparison (350.38±68.91) and MDD subjects (357.01±75.60) showed similar overall RT (Supplemental Material). These effects were qualified by a significant Group x Cue interaction (F=3.98, df=2,118, p<0.045). As evident from Figure 1A, the interaction reflected smaller RT differences on incentive versus no-incentive trials in MDD subjects. Relative to comparison subjects, the MDD group showed weaker reward-related RT modulation (RT no-incentive - RT reward; t=-2.09, df=59, p<0.047), with a similar trend for loss-related RT modulation (t=-1.97, df=59, p=0.053) (Figure 1B). However, no group differences in RT emerged for reward, loss, or no-incentive trials (ps>0.21). Moreover, both groups showed the shortest RT to reward cues, followed by loss and no-incentive cues (ps<0.002).
FIGURE 1.
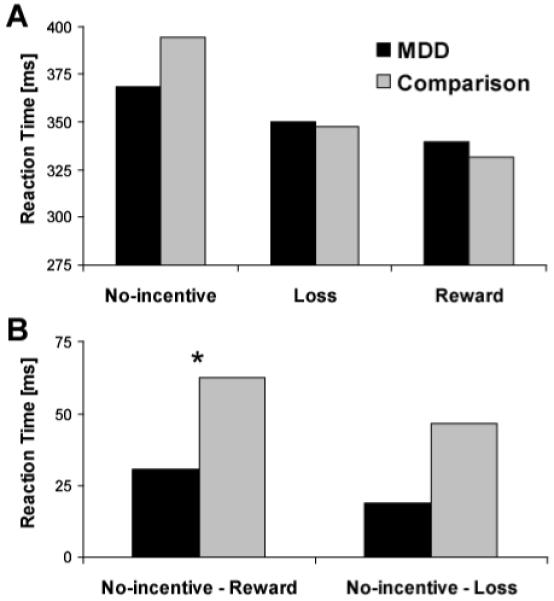
Behavioral findings during the monetary incentive delay task in MDD (N=30) and comparison (N=31) subjects.
(A) Reaction time (in ms) in response to the target as a function of reward, loss, or no-incentive cue. (B) Reaction Time difference scores (no-incentive - reward cue; no-incentive - loss cue) reveal significantly reduced relative reaction time speeding in MDD subjects for reward trials (p<0.047) and a similar trend for loss trials (p=0.053).
Mirroring the lack of Group effect in RTs collected during scanning, groups did not differ in target durations linked to successful or unsuccessful outcomes, which were selected based on RT during practice (Supplemental Material). There were also no group differences in the percentage of reward trials ending in gains or loss trials ending in penalties, or in total money earned (Supplemental Material, Table S2). Thus, fMRI findings were not confounded by group differences in task difficulty.
Affective Ratings
Ratings data indicated that the cues and outcomes elicited the intended responses (Supplemental Material, Figure S1). Critically, relative to comparison subjects, the MDD group reported overall reduced positive affect in response to both cue (Group: F=5.62, df=1,58, p<0.021) and feedback (Group: F=12.26, df=1,59, p<0.001) stimuli, as well as reduced arousal in response to gains (p<0.045) but not penalties or no-change feedback (ps>0.42), Group x Outcome interaction, F=3.20, df=2,118, p<0.045.
Functional MRI Data
Reward Anticipation (Reward cue-No-incentive cue)
A complete list of regions showing group differences is provided in the Supplemental Material (Table S3). Surprisingly, both groups showed robust basal ganglia responses to reward cues (Figure 2A). However, the MDD group showed relatively weaker activation in the left posterior putamen (Figure 2B/C).
FIGURE 2.

Reward-related anticipatory activation in MDD (N=26) and comparison (N=31) subjects.
Coronal (A) and axial (B) slices showing anticipatory reward activity [Reward cue - No-incentive cue] in basal ganglia regions are shown for both comparison and MDD subjects, as well as for the random effect analyses comparing the two groups. (A) Robust activation of ventral striatal regions, including the nucleus accumbens, is seen in both groups, leading to a lack of group differences. (B) Relative to comparison subjects, the MDD group shows significantly reduced activation during reward anticipation in the left putamen (x=-28, y=-13, z=-2). All contrasts are thresholded at p<0.005. Pt = Putamen, L = Left
Reward Outcome (Gain-No-change feedback)
Relative to comparison subjects, the MDD group showed significantly weaker responses to gain vs. no-change feedback in the left nucleus accumbens and bilateral dorsal caudate, including two sub-regions in right caudate and two in left caudate (Figure 3A/B). Both clusters in the right caudate and one in the left caudate remained significant following correction for multiple comparisons (Supplemental Material, Table S4); accordingly, differences in the nucleus accumbens should be considered preliminary. To test whether group differences were specific to reward outcomes, mean beta weights were extracted from each cluster and entered into Group x Condition (gains, penalties, no-change feedback) ANOVAs; for the caudate ROIs, the factor Subregion was added. For brevity, only effects involving Group are reported.
FIGURE 3.

Reward-related consummatory activation in MDD (N=26) and comparison (N=31) subjects.
Coronal slices showing consummatory reward activity [Gain feedback - No-change feedback] in basal ganglia regions are shown for both comparison and MDD subjects, as well as for the random effect analyses comparing the two groups. Relative to comparison subjects, the MDD group shows significantly reduced activation in response to gain feedback in the (A) left nucleus accumbens and (B) bilateral caudate. Follow-up analyses on beta weights extracted from the (C) nucleus accumbens and (D) bilateral caudate regions (averaged across three clusters) indicated that group differences were specific to reward outcome. All contrasts are thresholded at p<0.005. NAcc = Nucleus Accumbens, Cd = Caudate, L = Left
In the accumbens (Figure 3C), a main effect of Condition (F=3.46, df=2,110, p<0.040) was qualified by a trend for a Group x Condition interaction (F=2.94, df=2,110 p=0.063); the main effect of Group was not significant (p>0.085). Due to a priori hypotheses regarding the accumbens, and given the significant Group x Condition interaction in the whole-brain analysis, follow-up tests were performed to clarify the source of the interaction. Relative to comparison subjects, MDD subjects showed significantly weaker responses to gains (p<0.005) but not penalties or no-change feedback (ps>0.57). Furthermore, within-groups tests showed that while comparison subjects responded more strongly to gains versus both penalties (p<0.004) and no-change (p<0.001) feedback, in MDD subjects left accumbens activation was not modulated by condition (ps>0.39).
In the caudate (Figure 3D), the ANOVA revealed significant main effects of Subregion, Condition, and Group (ps<0.013), a significant Condition x Subregion interaction, and, most importantly, a significant Group x Condition interaction (F=7.89, df=2,110, p<0.002). This interaction was due to significantly greater activation for comparison versus MDD subjects in response to gains (p<0.0002), but not penalties (p>0.11) or no-incentive (p>0.45) feedback. Moreover, whereas comparison subjects showed increased bilateral caudate activation in response to both gains and losses (ps<0.0002) relative to no-change feedback, MDD subjects failed to show any feedback-dependent caudate modulation (ps>0.17). No correlations emerged between left putamen, left accumbens, or caudate activation and anhedonic symptoms in either group.
Morphometric Data
The Group x Hemisphere x Region ANOVA revealed no group differences (ps>0.18; Supplemental Material, Table S5). Among MDD participants, correlations were run between (i) proportional left accumbens and bilateral caudate volumes, and (ii) anhedonic symptoms and depression severity. For the left accumbens, no significant effects emerged. For the left and right caudate, volume was inversely related to total BDI (left: r=-0.489, p<0.015; right: r=-0.579, p<0.002) and anhedonic BDI (left: r=-0.553, p<0.004; right: r=-0.635, p<0.0001) subscores (Figure 4). Critically, both left and right caudate volumes predicted total BDI scores and anhedonic BDI subscores after adjusting for age and gender (total BDI score: left caudate ΔR2=0.203; right caudate ΔR2=0.309; anhedonic BDI subscore: left caudate ΔR2=0.281; right caudate ΔR2=0.387; all ΔF>6.09, ps<0.025).
FIGURE 4.
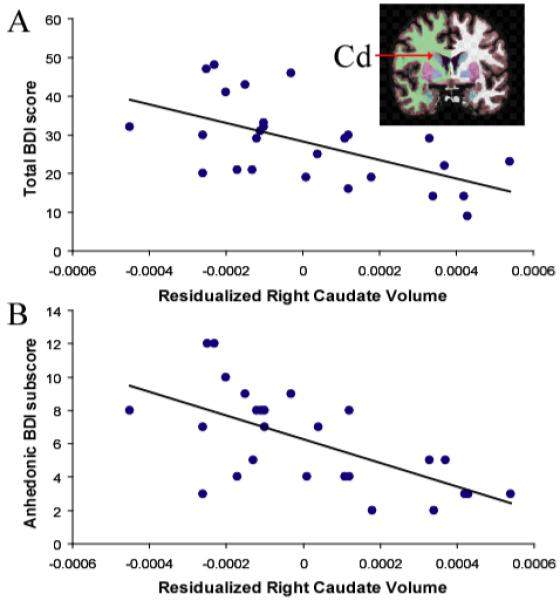
Relationship between clinical symptoms and caudate volume among MDD subjects (N=26).
Scatterplot and Pearson correlation between residualized right caudate volume (adjusted for age and gender) and (A) total BDI score (r=-0.579, p<0.002); and (B) anhedonic BDI subscore (r=-0.635, p<0.0001) among MDD subjects. Similar correlations emerged for the left caudate (total BDI: r=-0.489, p<0.015; anhedonic BDI: r=-0.553, p<0.004). The anhedonic BDI subscore was computed by summing item #4 (loss of pleasure), #12 (loss of interest), #15 (loss of energy), and #21 (loss of interest in sex). Cd = Caudate.
Control analyses (Supplemental Material)
In light of group differences in valence ratings for reward cues, and valence and arousal ratings for gains, control analyses evaluated whether group differences in left putamen reward cue responses and left accumbens and bilateral caudate gain responses remained after controlling for affective ratings. Regression analyses confirmed that this was the case. Moreover, group differences in accumbens and caudate gain responses remained after controlling for the volumes of these structures and group differences in reward-related RT modulation. In addition, no significant correlations between reward-related accumbens and caudate activation and volume of these regions emerged. Finally, there were no differences in basal ganglia activation for MDD subjects with (N=14) vs. without (N=16) comorbid anxiety.
Discussion
This study investigated anticipatory and consummatory phases of reward processing in depression. Behaviorally, the MDD group showed evidence of anhedonia, reporting generally reduced positive affect to reward stimuli and less arousal following gains. These findings were mirrored by group differences in basal ganglia responses to rewarding outcomes, as MDD participants showed weaker responses to gains in bilateral caudate and left nucleus accumbens. By contrast, there was less evidence of differences during reward anticipation. Both groups showed robust basal ganglia responses to reward cues, and although comparison subjects activated the left posterior putamen more strongly than MDD subjects, the size of the cluster was relatively small. Also, groups did not differ in reaction time as a function of cue, although relatively weaker modulation by reward was seen in MDD subjects (see difference scores). Finally, negative correlations between anhedonic symptoms (and depression severity) and caudate volume emerged in MDD subjects. These findings extend prior reports of basal ganglia dysfunction in MDD (6-11, 30), suggest that this dysfunction is more closely associated with consummatory rather than anticipatory deficits, and emphasize a role for reduced caudate volume in anhedonia.
Reduced Basal Ganglia Response to Rewarding Outcomes in MDD
The strong caudate response to gains in comparison subjects fits human (18, 20, 31) and animal (32) studies demonstrating this structure’s sensitivity to reward-related information. Importantly, the caudate responds maximally when rewards are unpredictable (e.g., when delivered on 50% of reward trials, as done here) and subjects believe that outcomes are contingent on their actions (31). Accordingly, the between-group caudate difference suggests weaker perceived action-outcome relationship and/or weaker responses to unpredictable rewards in depression.
Evidence for the first interpretation is mixed. Although groups differed in reward-related reaction time modulation (reaction time difference scores), there was no group difference in reactions on reward trials and both groups responded faster on reward trials than on loss or no-incentive trials. Thus, both groups behaved as though their responses influenced the chance of receiving gains. Alternatively, the impact of the gains may have been weaker in MDD subjects. This is consistent with the fact that MDD subjects reported overall blunted affective responses and decreased arousal to gains. In addition, group differences were also observed in the left nucleus accumbens, a region that responds strongly to rewarding stimuli (33). Importantly, activity in the accumbens appears to track the hedonic value of outcomes (31, 34). Thus, while the group difference in caudate responses suggests a depression-related deficit in expressing goal-directed behaviors, the finding in the accumbens indicates a more primary deficit in hedonic coding. These results are consistent with evidence indicating that deep brain stimulation to the accumbens (35) and ventral capsule/ventral striatum (36) significantly reduced symptom severity and anhedonia in treatment-resistant MDD patients. Collectively, these findings indicate that dysfunction in regions mediating hedonic impact (accumbens) and reinforcement of actions (caudate) play an important role in the pathophysiology of MDD.
The group differences in gain responses are intriguing in light of reports of reduced ability to modulate behavior as a function of intermittent rewards in MDD (5). Using a probabilistic reward task, we found that depressed subjects, particularly those reporting anhedonic symptoms, showed a reduced response bias toward a more frequently rewarded stimulus relative to controls. Furthermore, healthy controls with blunted response bias in the probabilistic task also generated weak basal ganglia responses to gains in the fMRI task used here (37). These considerations suggest that weak basal ganglia responses to unpredictable rewards may contribute to poor learning of action-reward contingencies in MDD.
Intact Basal Ganglia Responses to Reward Cues in MDD
Surprisingly, both groups showed robust basal ganglia responses to reward cues. However, in contrast to a prior study (17), the current MDD group showed weaker reward-related reaction time modulation and affective responses to reward-related stimuli relative to comparison subjects. Thus, behavioral evidence of reward processing deficits can coexist with significant basal ganglia responses to reward-predicting cues.
The nature of the intact basal ganglia response to reward cues in MDD subjects is unclear. In incentive delay tasks, anticipatory ventral striatal activity is typically regarded as related to the dopamine signal seen in response to reward cues in electrophysiological studies (38). In non-human primates, this signal is first elicited by unpredicted rewards and travels back to cues only when a cue-outcome contingency is learned (14). In our study, comparison subjects showed a significantly stronger basal ganglia response to gains than MDD subjects, yet the two groups showed few differences in response to reward cues. This suggests two possibilities: (i) the unlikely possibility that the dopamine signal traveled from the gains (consummatory phase) to the cues (anticipatory phase) more rapidly in MDD subjects, or (ii) the more likely possibility that the reward cues elicited a ventral striatal response on their own that was similar across groups and possibly independent of transmission of the dopamine signal elicited by gains. This possibility is rarely considered in studies using incentive delay tasks, but because participants know that reward cues can lead to gains, it is possible that the cues may elicit ventral striatal activation from the outset. However, even if this is the case, a group difference in ventral striatal response to reward cues might still be expected (8). Future studies in which participants learn cue-reward associations over time are necessary to investigate this issue.
Reduced Caudate Volume and Anhedonia
Replicating findings with non-clinical subjects (23), MDD subjects with elevated anhedonic symptoms showed reduced bilateral caudate volume. This relationship provides impetus for continued investigation of depressive endophenotypes (1, 2), because it is unclear whether reduced caudate volume predisposes individuals to anhedonic or more severe depression, or instead represents a state-related correlate of these symptoms.
Limitations
Several limitations should be emphasized. First, in spite of clear a priori hypotheses about the nucleus accumbens (8, 10, 11), the Group x Condition interaction in this region emerged at p<0.005, and this difference was not significant after correction for multiple comparisons due to the small cluster size (Supplemental Material). Moreover, no correlations between striatal activation and anhedonic symptoms emerged. Consequently, future studies are needed to confirm the role of the nucleus accumbens in reward dysfunction in MDD. Given mounting interest in the role of accumbens in the pathophysiology of MDD, as exemplified by recent deep brain stimulation studies targeting this region (35, 36), the current finding of reduced reward-related accumbal responses is nevertheless intriguing. Second, correlations between caudate volume and depression severity emerged for the BDI, but not the HRSD. Although the reason for this discrepancy is unclear, it is possible that several BDI items tapping anhedonia may have contributed to this finding. In spite of these limitations, this study indicates that anhedonia—a core component of MDD—may reflect weak reward consummatory responses in the basal ganglia, particularly the nucleus accumbens and caudate, and is related to reduced caudate size.
Supplementary Material
Acknowledgments
Disclosures and acknowledgments. Dr. Pizzagalli has received research support from GlaxoSmithKline and Merck & Co., Inc. Dr. Dougherty has received research support from Forest, Eli Lilly, Medtronic, Cyberonics, Northstar Neuroscience, Cephalon, and McNeil. He has received honoraria from Cyberonics, Medtronic, Northstar Neuroscience, and McNeil and has served as a consultant to Jazz Pharmaceuticals and Transcept Pharmaceuticals. Dr. Iosifescu has received research support from Aspect Medical Systems, Forest Laboratories, Janssen Pharmaceutica, and honoraria from Aspect Medical Systems, Cephalon, Gerson Lehrman Group, Eli Lilly & Co., Forest Laboratories and Pfizer, Inc. Dr. Rauch has received research support from Medtronics, Cyberonics, and Cephalon, and honoraria from Novartis, Neurogen, Sepracor, Primedia, and Medtronics, Inc. Dr. Fava has received research support from Abbott Laboratories, Alkermes, Aspect Medical Systems, Astra-Zeneca, Bristol-Myers Squibb Company, Cephalon, Eli Lilly & Company, Forest Pharmaceuticals Inc., GlaxoSmithKline, J & J Pharmaceuticals, Lichtwer Pharma GmbH, Lorex Pharmaceuticals, Novartis, Organon Inc., PamLab, LLC, Pfizer Inc, Pharmavite, Roche, Sanofi-Aventis, Solvay Pharmaceuticals, Inc., Synthelabo, and Wyeth-Ayerst Laboratories. He has received advisory/consulting fees from Abbott Laboratories, Amarin, Aspect Medical Systems, Astra-Zeneca, Auspex Pharmaceuticals, Bayer AG, Best Practice Project Management, Inc., Biovail Pharmaceuticals, Inc., BrainCells, Inc. Bristol-Myers Squibb Company, Cephalon, CNS Response, Compellis, Cypress Pharmaceuticals, Dov Pharmaceuticals, Eli Lilly & Company, EPIX Pharmaceuticals, Fabre-Kramer Pharmaceuticals, Inc., Forest Pharmaceuticals Inc., GlaxoSmithKline, Grunenthal GmBH, Janssen Pharmaceutica, Jazz Pharmaceuticals, J & J Pharmaceuticals, Knoll Pharmaceutical Company, Lorex Pharmaceuticals, Lundbeck, MedAvante, Inc., Merck, Neuronetics, Novartis, Nutrition 21, Organon Inc., PamLab, LLC, Pfizer Inc, PharmaStar, Pharmavite, Precision Human Biolaboratory, Roche, Sanofi-Aventis, Sepracor, Solvay Pharmaceuticals, Inc., Somaxon, Somerset Pharmaceuticals, Synthelabo, Takeda, Tetragenex, Transcept Pharmaceuticals, Vanda Pharmaceuticals Inc, and Wyeth-Ayerst Laboratories. In addition, Dr. Fava has received speaking fees from Astra-Zeneca, Boehringer-Ingelheim, Bristol-Myers Squibb Company, Cephalon, Eli Lilly & Company, Forest Pharmaceuticals Inc., GlaxoSmithkline, Novartis, Organon Inc., Pfizer Inc, PharmaStar, Primedia, Reed-Elsevier, and Wyeth-Ayerst Laboratories. Finally, Dr. Fava has equity holdings in Compellis and MedAvante, and holds patent applications for SPCD and for a combination of azapirones and bupropion in MDD, and receives copyright royalties for the MGH CPFQ, DESS, and SAFER. Mr. Holmes, Dr. Dillon, Ms. Goetz, Mr. Birk, and Mr. Bogdan report no competing interests.
This project was supported by Grant Number R01 MH68376 (DAP) from the National Institute of Mental Health (NIMH) and by Grant Numbers R21 AT002974 (DAP) and R01 AT1638 (MF) from the National Center for Complementary and Alternative Medicine (NCCAM). Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the NIMH, NCCAM, or the National Institutes of Health. The authors are grateful to Allison Jahn and Kyle Ratner for their assistance at early phases of this project, to James O’Shea and Decklin Foster for skilled technical assistance, and to Nancy Brooks, Christen Deveney, Deborah Shear, Judith Katz, Adrienne Van Nieuwenhuizen, Carrie Brintz, Sunny Dutra, and Mariko Jameson for assistance with subject recruitment.
ClinicalTrials.gov number: NCT00183755
Footnotes
Previous presentation.
The data in this paper were presented in preliminary form at the 22nd Annual Meeting of the Society for Research in Psychopathology, Pittsburgh, Pennsylvania, USA, September 25-28, 2008.
References
- 1.Hasler G, Drevets WC, Manji HK, Charney DS. Discovering endophenotypes for major depression. Neuropsychopharmacology. 2004;29:1765–1781. doi: 10.1038/sj.npp.1300506. [DOI] [PubMed] [Google Scholar]
- 2.Pizzagalli DA, Jahn AL, O’Shea JP. Toward an objective characterization of an anhedonic phenotype: A Signal-detection approach. Biol Psychiatry. 2005;57:319–327. doi: 10.1016/j.biopsych.2004.11.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Joormann J, Gotlib IH. Selective attention to emotional faces following recovery from depression. J Abnorm Psychol. 2007;116:80–85. doi: 10.1037/0021-843X.116.1.80. [DOI] [PubMed] [Google Scholar]
- 4.Berenbaum H, Oltmanns TF. Emotional experience and expression in schizophrenia and depression. J Abnorm Psychol. 1992;101:37–44. doi: 10.1037//0021-843x.101.1.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pizzagalli DA, Iosifescu D, Hallett LA, Ratner KG, Fava M. Reduced hedonic capacity in Major Depressive Disorder: Evidence from a probabilistic reward task. J Psychiatr Res. 2009;43:76–87. doi: 10.1016/j.jpsychires.2008.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Drevets WC, Videen TO, Price JL, Preskorn SH, Carmichael ST, Raichle ME. A functional anatomical study of unipolar depression. J Neurosci. 1992;12:3628–3641. doi: 10.1523/JNEUROSCI.12-09-03628.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Elliott R, Sahakian BJ, Michael A, Paykel ES, Dolan RJ. Abnormal neural response to feedback on planning and guessing tasks in patients with unipolar depression. Psychol Med. 1998;28:559–571. doi: 10.1017/s0033291798006709. [DOI] [PubMed] [Google Scholar]
- 8.Epstein J, Pan H, Kocsis JH, Yang Y, Butler T, Chusid, Hochberg H, Murrough J, Strohmayer E, Stern E, Silbersweig DA. Lack of ventral striatal response to positive stimuli in depressed versus normal subjects. Am J Psychiatry. 2006;163:1784–1790. doi: 10.1176/ajp.2006.163.10.1784. [DOI] [PubMed] [Google Scholar]
- 9.Keedwell PA, Andrew C, Williams SCR, Brammer MJ, Phillips ML. The neural correlates of anhedonia in Major Depressive Disorder. Biol Psychiatry. 2005;58:843–853. doi: 10.1016/j.biopsych.2005.05.019. [DOI] [PubMed] [Google Scholar]
- 10.Kumar P, Waiter G, Ahearn T, Milders M, Reid I, Steele JD. Abnormal temporal difference reward-learning signals in major depression. Brain. doi: 10.1093/brain/awn136. in press. [DOI] [PubMed] [Google Scholar]
- 11.Steele JD, Kumar P, Ebmeier KP. Blunted response to feedback information in depressive illness. Brain. 2007;130:2367–2374. doi: 10.1093/brain/awm150. [DOI] [PubMed] [Google Scholar]
- 12.Berridge KC, Robinson TE. What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? Brain Res Brain Res Rev. 1998;28:309–369. doi: 10.1016/s0165-0173(98)00019-8. [DOI] [PubMed] [Google Scholar]
- 13.Gard DE, Germans Gard M, Kring AM, John OP. Anticipatory and consummatory components of the experience of pleasure: a scale development study. J Res Person. 2006;40:1086–1102. [Google Scholar]
- 14.Schultz W. Multiple reward signals in the brain. Nat Rev Neurosci. 2000;1:199–207. doi: 10.1038/35044563. [DOI] [PubMed] [Google Scholar]
- 15.Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci. 1986;9:357–381. doi: 10.1146/annurev.ne.09.030186.002041. [DOI] [PubMed] [Google Scholar]
- 16.Dunlop BW, Nemeroff CB. The role of dopamine in the pathophysiology of depression. Arch Gen Psychiatry. 2007;64:327–337. doi: 10.1001/archpsyc.64.3.327. [DOI] [PubMed] [Google Scholar]
- 17.Knutson B, Bhanji JP, Cooney RE, Atlas LY, Gotlib IH. Neural responses to monetary incentives in major depression. Biol Psychiatry. 2008;63:686–692. doi: 10.1016/j.biopsych.2007.07.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Delgado MR, Nystrom LE, Fissell C, Noll DC, Fiez JA. Tracking the hemodynamic responses to reward and punishment in the striatum. J Neurophysiol. 2000;84:3072–3077. doi: 10.1152/jn.2000.84.6.3072. [DOI] [PubMed] [Google Scholar]
- 19.Delgado MR, Locke HM, Stenger VA, Fiez JA. Dorsal striatum responses to reward and punishment: effects of valence and magnitude manipulations. Cognit Affect Behav Neurosci. 2003;3:27–38. doi: 10.3758/cabn.3.1.27. [DOI] [PubMed] [Google Scholar]
- 20.Delgado MR, Miller MM, Inati S, Phelps EA. An fMRI study of reward-related probability learning. NeuroImage. 2005;24:862–873. doi: 10.1016/j.neuroimage.2004.10.002. [DOI] [PubMed] [Google Scholar]
- 21.Dillon DG, Holmes AJ, Jahn AL, Bogdan R, Wald LL, Pizzagalli DA. Dissociation of neural regions associated with anticipatory versus consummatory phases of incentive processing. Psychophysiology. 2008;45:36–49. doi: 10.1111/j.1469-8986.2007.00594.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Knutson B, Cooper JC. Functional magnetic resonance imaging of reward prediction. Curr Opin Neurol. 2005;18:411–417. doi: 10.1097/01.wco.0000173463.24758.f6. [DOI] [PubMed] [Google Scholar]
- 23.Harvey PO, Pruessner J, Czechowska Y, Lepage M. Individual differences in trait anhedonia: a structural and functional magnetic resonance imaging study in non-clinical subjects. Mol Psychiatry. 2007;12:767–775. doi: 10.1038/sj.mp.4002021. [DOI] [PubMed] [Google Scholar]
- 24.First MB, Spitzer RL, Gibbon M, Williams JBW. Structured Clinical Interview for DSM-IV-TR Axis I Disorders, Research Version, Patient Edition. (SCID-I/P) Biometrics Research, New York State Psychiatric Institute; New York, NY: 2002. [Google Scholar]
- 25.Hamilton M. A rating scale for depression. J Neurol Neurosurg Psychiatry. 1960;23:56–62. doi: 10.1136/jnnp.23.1.56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Beck AT, Steer RA, Brown GK. Beck Depression Inventory Manual. 2nd ed. The Psychological Corporation; San Antonio, TX: 1996. [Google Scholar]
- 27.Fischl B, Salat DH, Busa E, Albert M, Dieterich M, Haselgrove C, van der Kouwe A, Killiany R, Kennedy D, Klaveness S, Montillo A, Makris N, Rosen B, Dale AM. Whole brain segmentation: automated labeling of neuroanatomical structures in the human brain. Neuron. 2002;33:341–355. doi: 10.1016/s0896-6273(02)00569-x. [DOI] [PubMed] [Google Scholar]
- 28.Tae WS, Kim SS, Lee KU, Nam EC, Kim KW. Validation of hippocampal volumes measured using a manual method and two automated methods (FreeSurfer and IBASPM) in chronic major depressive disorder. Neuroradiology. 2008;50:569–581. doi: 10.1007/s00234-008-0383-9. [DOI] [PubMed] [Google Scholar]
- 29.Pizzagalli DA, Goetz E, Ostacher M, Iosifescu D, Perlis RH. Euthymic patients with Bipolar Disorder show decreased reward learning in a probabilistic reward task. Biol Psychiatry. 2008;64:162–168. doi: 10.1016/j.biopsych.2007.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tremblay LK, Naranjo CA, Graham SJ, Herrmann N, Mayberg HS, Hevenor S, Busto UE. Functional neuroanatomical substrates of altered reward processing in major depressive disorder revealed by a dopaminergic probe. Arch Gen Psychiatry. 2005;62:1228–1236. doi: 10.1001/archpsyc.62.11.1228. [DOI] [PubMed] [Google Scholar]
- 31.Tricomi EM, Delgado MR, Fiez JA. Modulation of caudate activity by action contingency. Neuron. 2004;41:281–292. doi: 10.1016/s0896-6273(03)00848-1. [DOI] [PubMed] [Google Scholar]
- 32.Kawagoe R, Takikawa Y, Hikosaka O. Expectation of reward modulates cognitive signals in the basal ganglia. Nat Neurosci. 1998;1:411–416. doi: 10.1038/1625. [DOI] [PubMed] [Google Scholar]
- 33.Berns GS, McClure SM, Pagnoni G, Montague PR. Predictability modulates human brain response to reward. J Neurosci. 2001;21:2793–2798. doi: 10.1523/JNEUROSCI.21-08-02793.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.O’Doherty J, Dayan P, Schultz J, Deichmann R, Friston K, Dolan RJ. Dissociable roles of ventral and dorsal striatum in instrumental conditioning. Science. 2004;304:452–454. doi: 10.1126/science.1094285. [DOI] [PubMed] [Google Scholar]
- 35.Schlaepfer TE, Cohen MX, Frick C, Kosel M, Brodesser D, Axmacher N, Joe AY, Kreft M, Lenartz D, Sturm V. Deep brain stimulation to reward circuitry alleviates anhedonia in refractory major depression. Neuropsychopharmacology. 2008;33:368–377. doi: 10.1038/sj.npp.1301408. [DOI] [PubMed] [Google Scholar]
- 36.Malone DA, Jr, Dougherty DD, Rezai AR, Carpenter LL, Friehs GM, Eskandar EN, Rauch SL, Rasmussen SA, Machado AG, Kubu CS, Tyrka AR, Price LH, Stypulkowski PH, Giftakis JE, Rise MT, Malloy PF, Salloway SP, Greenberg BD. Deep brain stimulation of the ventral capsule/ventral striatum for treatment-resistant depression. Biol Psychiatry. 2008 Oct 6; doi: 10.1016/j.biopsych.2008.08.029. Epub ahead of print. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Santesso DL, Dillon DG, Birk JL, Holmes AJ, Goetz E, Bogdan R, Pizzagalli DA. Individual differences in reinforcement learning: Behavioral, electrophysiological, and neuroimaging correlates. NeuroImage. 2008;42:807–816. doi: 10.1016/j.neuroimage.2008.05.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Knutson B, Gibbs SE. Linking nucleus accumbens dopamine and blood oxygenation. Psychopharmacology. 2007;191:813–822. doi: 10.1007/s00213-006-0686-7. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.