

Abstract
Leaf isoprene emission scales positively with light intensity, is inhibited by high carbon dioxide (CO2) concentrations, and may be enhanced or inhibited by low oxygen (O2) concentrations, but the mechanisms of environmental regulation of isoprene emission are still not fully understood. Emission controls by isoprene synthase, availability of carbon intermediates, or energetic cofactors have been suggested previously. In this study, we asked whether the short-term (tens of minutes) environmental control of isoprene synthesis results from alterations in the immediate isoprene precursor dimethylallyldiphosphate (DMADP) pool size, and to what extent DMADP concentrations are affected by the supply of carbon and energetic metabolites. A novel in vivo method based on postillumination isoprene release was employed to measure the pool size of DMADP simultaneously with the rates of isoprene emission and net assimilation at different light intensities and CO2 and O2 concentrations. Both net assimilation and isoprene emission rates increased hyperbolically with light intensity. The photosynthetic response to CO2 concentration was also hyperbolic, while the CO2 response curve of isoprene emission exhibited a maximum at close to CO2 compensation point. Low O2 positively affected both net assimilation and isoprene emission. In all cases, the variation in isoprene emission was matched with changes in DMADP pool size. The results of these experiments suggest that DMADP pool size controls the response of isoprene emission to light intensity and to CO2 and O2 concentrations and that the pool size is determined by the level of energetic metabolites generated in photosynthesis.
Plants interact with the atmosphere mainly through carbon dioxide (CO2), oxygen (O2), and water fluxes but also through various trace gas fluxes. Plant-generated volatile organic compounds are significant players in tropospheric photochemistry (Trainer et al., 1987; Monson et al., 1991), catalyzing the synthesis of ozone and other oxidants as well as prolonging the lifetime of the greenhouse gas methane (Monson et al., 2007). Plant emissions of isoprene are estimated worldwide to be the largest source of biogenic volatile organic compound emissions (Guenther et al., 2006; Arneth et al., 2008). Therefore, predicting plant isoprene fluxes is of key relevance in simulating atmospheric reactivity.
Apart from the atmospheric significance, isoprene plays an important role in protecting plants from heat and oxidative stresses (Fang et al., 1996; Litvak et al., 1996; Sharkey, 1996; Sharkey et al., 1996; Loreto et al., 2001; Loreto and Velikova, 2001). As only some specific plant species are able to produce isoprene, the capacity of plants to make isoprene can alter plant competitive relations under heat and ozone stresses (Lerdau, 2007; Darbah et al., 2008). It has further been suggested that the isoprenoid synthesis pathway can act as a metabolic protection valve for dissipation of excess assimilatory and reductive power (Fall, 1999; Rosenstiel et al., 2004; Sanadze, 2004; Magel et al., 2006). This information collectively emphasizes the need to gain mechanistic insight into the determinants of isoprene emission.
Despite the rich experimental evidence accumulated over several decades, there is still no general agreement about the mechanisms controlling the rate of isoprene synthesis under different environmental conditions. At present, the models of isoprene emission include a great deal of empiricism (Guenther et al., 1993; Monson et al., 2007; Grote and Niinemets, 2008; Wilkinson et al., 2009), and although they provide numerically good fits to the data, the capacity of these models to predict isoprene fluxes in future conditions and under stress is uncertain (Monson et al., 2007; Arneth et al., 2008). Isoprene emission rate in the emitting species depends on temperature, light intensity, and the concentrations of CO2 and O2 (Sanadze, 1969, 1990; Sanadze and Tarhnishvili, 1986; Monson and Fall, 1989; Loreto and Sharkey, 1990; Monson et al., 1991), but there is no consensus on what drives these dependencies.
Early discoveries of the light dependence of isoprene emission (Sanadze, 1969), the rapid appearance of 13C label from 13C-labeled CO2 in the isoprene signal (Sanadze et al., 1972; Mgalobilishvili et al., 1978), and the observation that isolated chloroplasts are capable of isoprene emission (Mgalobilishvili et al., 1978) have provided conclusive evidence of functional interdependence between photosynthesis and isoprene emission. Further studies have shown strong positive relationships between the photosynthetic carbon assimilation and isoprene synthesis mediated by photosynthetic metabolites that provide carbon skeletons for and photosynthetic ATP and NADPH energetically supporting the chloroplastic 2-C-methyl-erythritol 4-phosphate (MEP) pathway (Loreto and Sharkey, 1990, 1993; Eisenreich et al., 2001; Sharkey and Yeh, 2001). However, after the discovery of isoprene synthase (IspS) catalyzing the formation of isoprene from its immediate precursor dimethylallyldiphosphate (DMADP), light regulation of isoprene synthase activity has been hypothesized to be responsible for the light dependence of isoprene emission (Silver and Fall, 1991; Fall and Wildermuth, 1998; Wildermuth and Fall, 1998; Logan et al., 2000). At present, there is no consensus on whether the light response of isoprene emission is driven by changes in the activity of isoprene synthase (Wiberley et al., 2008) or by the availability of energetic cofactors and carbon intermediates that determine the availability of DMADP for IspS (Monson and Fall, 1989; Rosenstiel et al., 2002).
Another key uncertainty is what drives the CO2 response of isoprene emission. While net assimilation rate scales positively with the CO2 concentration, high CO2 concentrations inhibit isoprene release (Monson and Fall, 1989; Loreto and Sharkey, 1990). This has been hypothesized to reflect inhibition of the activity of IspS or another enzyme of the MEP pathway (Potosnak, 2002). However, DMADP pool size varies throughout the CO2 response of isoprene emission (Rosenstiel et al., 2003). Based on this variation, the alternative hypothesis suggests that transport of the intermediate of the MEP pathway, phosphoenolpyruvate (PEP), from cytosol becomes increasingly limited at elevated CO2 due to enhanced cytosolic PEP carboxylation at higher CO2 concentrations (Rosenstiel et al., 2003, 2004, 2006; Wilkinson et al., 2009). In fact, a negative correlation between PEP carboxylase activity and isoprene emission rate has been observed across a variety of environmental treatments (Loreto et al., 2007). Finally, it has been suggested that high CO2 concentrations inhibit isoprene emission due to limited availability of ATP for isoprene synthesis (Loreto and Sharkey, 1990, 1993; Sharkey et al., 1991b). Provided that the effective Km for ATP of the isoprenoid synthesis pathway is large, isoprenoid synthesis at high CO2 may become limited as a result of a drawdown of leaf ATP level. Such a drawdown commonly occurs due to increased consumption of ATP for carbon reduction that is typically limited by electron transport activity at higher CO2 (Farquhar et al., 1980) and because of sequestration of chloroplastic phosphate into sugar phosphates, thereby feedback inhibiting photosynthetic electron transport (Loreto and Sharkey, 1990, 1993; Sharkey et al., 1991b).
The least studied and most controversial is the influence of O2 on isoprene emission. Although mechanistically important, there has been little interest in O2 effects because of only moderate variations in its atmospheric concentration during and between seasons (Keeling and Shertz, 1992) and in the recent geological past (Kump, 2008). Among the available studies on O2 effects, some have reported the increase of isoprene emission under low O2 of 1.5% to 2% relative to the ambient level of 21% (Sanadze, 1966; Sanadze and Tarhnishvili, 1986; Hewitt et al., 1990). In other studies, isoprene emission was either enhanced or reduced by O2 concentration in dependence on ambient CO2 concentration (Loreto and Sharkey, 1990, 1993). Finally, short-term stimulation and long-term (>40–60 min) inhibition of isoprene emission by low O2 have been reported (Monson and Fall, 1989). As O2 importantly alters the share of the photosynthetic limitations between electron transport and carbon input, an analysis of O2 dependence can provide fundamental information on the key limitations of isoprene emission.
The controversies in the literature partly arise from the circumstance that it has been difficult to estimate the intermediates of the isoprene emission pathway simultaneously with foliage physiological characteristics. We have previously developed a novel in vivo method for the measurement of the immediate isoprene precursor DMADP pool responsible for isoprene emission (presumably the chloroplastic pool) by integrating the postillumination isoprene emission (Rasulov et al., 2009) that overcomes this difficulty. In this study, we combine the measurements of foliage photosynthetic traits, isoprene emission, and DMADP pool size at different light intensities and CO2 and O2 concentrations in hybrid aspen (Populus tremula × P. tremuloides) to ask (1) whether the alterations in isoprene synthase activity or in DMADP pool size are responsible for the short-term controls by these driving variables on isoprene emission, and (2) to what extent substrate concentrations are affected by the emission rate and the supply of carbon and energetic metabolites? The results of these experiments suggest that the variation in the DMADP pool size controls the response of isoprene emission to light intensity and CO2 and O2 concentrations and that the pool size is determined by the level of energetic metabolites (ATP) generated in photosynthesis.
RESULTS
Light Responses of Net Assimilation, Isoprene Emission, and DMADP Pool Size
Both the rate of isoprene emission and net assimilation scaled curvilinearly with quantum flux density (Q), but net assimilation rate saturated at lower Q than isoprene emission (Fig. 1). Fitting the data by Equation 1 suggested that the isoprene emission rate at the highest Q achieved by the system (550 μmol m−2 s−1) was approximately 70% of the light-saturated value and the net assimilation rate was 85% of the light-saturated value, while the isoprene emission rate was 80% and the net assimilation rate was 95% of the rates predicted at Q = 1,000 μmol m−2 s−1. The pool size of DMADP measured kinetically as an integral of the postillumination isoprene emission at each light intensity (Rasulov et al., 2009) increased in parallel with the rate of isoprene emission (Fig. 2). This relationship had an initial slope (DMADP pool range of 150–500 nmol m−2) of 0.034 s−1 (the maximum turnover rate of the DMADP pool) and the slope was 0.021 s−1 for higher DMADP pool sizes (600–1,000 nmol m−2), demonstrating that this dependence was nearly linear.
Figure 1.

Dependencies of the rates of isoprene emission and net assimilation, and the chloroplastic DMADP pool size, on incident photosynthetic quantum flux density in hybrid aspen (clone 200) leaves. The measurements were carried out at an ambient CO2 concentration of 390 μmol mol−1, O2 concentration of 210 mmol mol−1, and leaf temperature of 28°C to 30°C. An in vivo method based on the postillumination isoprene release was used to determine the chloroplastic DMADP pool size concomitant with leaf physiological measurements (Rasulov et al., 2009). Error bars show sd (n = 5). Data were fitted by Equation 1 (r2 > 0.98, P < 0.001 for all).
Figure 2.

Correlation between the isoprene emission rate and the chloroplastic DMADP pool size (the same data as in Fig. 1). Data were fitted by linear regression. Arrows with the values denote the quantum flux density (μmol m−2 s−1) corresponding to each measurement. The pool size of DMADP was determined according to an in vivo method based on postillumination isoprene release (Rasulov et al., 2009).
Temporal kinetics of dark/light activation of foliage physiological characteristics were studied after a 10-h dark period (Fig. 3A). Fitting the induction data by a single-exponential model (Eq. 2) indicated that net assimilation was induced with the fastest rate (k1 = 0.077 min−1), followed by stomata (k1 = 0.042 min−1 for the transpiration rate) and isoprene emission (k1 = 0.036 min−1; Fig. 3B). However, the induction of isoprene emission was delayed for about 5 min. In addition, the induction of isoprene emission continued for more than 80 min, while the rates of transpiration and net assimilation reached a saturation in approximately 30 min after switching on the light. In fact, a biphasic, double-exponential model (Eq. 3) better fitted the isoprene emission data (Fig. 3B; r2 = 0.99) than the monophasic, single-exponential model (Fig. 3B; r2 = 0.96).
Figure 3.

Time courses of the light activation of leaf photosynthesis, isoprene emission, and transpiration rates in hybrid aspen plants dark conditioned for 10 h (A), and normalized induction responses with single (Eq. 2) and double (Eq. 3) exponential data fits (B). After the dark period, light (550 μmol m−2 s−1) was switched on and leaf physiological characteristics were continuously monitored until full induction. To directly compare the shapes of the induction curves in B, all data were normalized such that the normalized rate was 0 at the beginning of the transient and 1 at the steady state. Leaf temperature was 24°C in the darkness and increased to 28°C to 30°C in the light.
Effects of Ambient CO2 Concentration on Net Assimilation, Isoprene Emission, and DMADP Pool Size
Net assimilation rate responded to ambient CO2 concentration according to a classical hyperbola, exhibiting photorespiratory CO2 evolution in the absence of external CO2, with CO2 compensation point at about 75 μmol mol−1 and approaching CO2 saturation at concentrations above 1,000 μmol mol−1 (Fig. 4A). In contrast, the CO2 response of isoprene emission was a curve with a maximum that occurred close to the CO2 compensation point of photosynthesis. The reduction of isoprene emission toward lower CO2 concentrations (minimum ambient CO2 concentration achieved was 8–12 μmol mol−1, corresponding to a CO2 concentration in substomatal cavities of 20–30 μmol mol−1) was about 20%. The reduction of isoprene emission toward higher CO2 concentrations was much larger, with the emission rate at the highest CO2 concentration being only approximately 10% of its maximum rate.
Figure 4.

Dependencies of the rates of net assimilation, isoprene emission, and transpiration (A) and the size of the chloroplastic DMADP pool (B) on ambient CO2 concentration (Ca) at saturating light (550 μmol m−2 s−1) and ambient O2 concentration (210 mmol mol−1) in hybrid aspen leaves. Leaf temperature was 28°C to 30°C. CO2 concentration in the substomatal cavities was 26 μmol mol−1 at the lowest Ca and 1,230 μmol mol−1 at the highest Ca (calculated from stomatal conductance, assimilation rate, and Ca). The inset in B demonstrates the dependence of isoprene emission rate on DMADP pool size. Chloroplastic DMADP pool size was estimated on the basis of an in vivo method using postillumination isoprene release (Rasulov et al., 2009). Error bars show sd (n = 5).
DMADP pool size responded to CO2 concentration analogously with the isoprene emission rate (Fig. 4B), and DMADP pool size and isoprene emission rate were strongly correlated throughout the entire CO2 response curve (Fig. 4B, inset). The initial slope of the isoprene emission versus DMADP pool size (DMADP pool turnover rate) was 0.029 s−1.
O2 Responses of Net Assimilation, Isoprene Emission, and DMADP Pool Size
O2 concentration was altered between 2% and 21% at different CO2 concentrations and light intensities. At close to ambient CO2 concentration of 350 μmol mol−1 and light intensity of 550 μmol m−2 s−1, a typical Warburg effect was observed (i.e. an increase of net assimilation rate by about 30% to 40% at 2% O2 compared with 21% O2; Fig. 5A). An even more pronounced positive response was observed in isoprene emission, which increased 1.5- to 1.7-fold in 2% O2 relative to 21% O2. The enhancement of isoprene emission at the low O2 concentration was persistent for 20 to 40 min and completely reversible (Fig. 5, A and D).
Figure 5.

Influence of O2 concentration on isoprene emission (white bars) and net assimilation (gray bars) rates at different quantum flux densities and ambient CO2 concentrations in the hybrid aspen leaves. High light corresponds to 550 μmol m−2 s−1 and low light to 200 μmol m−2 s−1, while normal CO2 concentration corresponds to 350 μmol mol−1 and high CO2 to 1,150 μmol mol−1. Leaf temperature was 28°C to 30°C in all experiments. Error bars denote se (n = 5). Lowercase letters indicate significant (P < 0.05) differences among the isoprene emission rates within a given panel (paired samples t test), while uppercase letters denote the differences among the net assimilation rates. The order of bars denotes the sequence of environmental changes for each individual leaf. Each leaf was kept at each set of environmental conditions until the steady-state rates of isoprene emission and net assimilation rate were observed. The conditions for A and D were the same to test whether the physiological status of the leaf changed during the experiment. In none of the pairwise comparisons were the average isoprene emission and net assimilation rates different between the corresponding treatments among A and D (P > 0.6).
At the low light intensity of 200 μmol m−2 s−1, the enhancement of isoprene emission by low O2 concentration (1.07-fold increase in 2% O2; Fig. 5B) was statistically nonsignificant. Under these conditions, net assimilation rate also did not significantly respond to low O2 (Fig. 5B). The low O2 effect was also small, but statistically significant, at the saturating CO2 concentration of 1,150 μmol mol−1 (1.13-fold increase in 2% O2; Fig. 5C), but net assimilation rate was not significantly affected by low O2 (Fig. 5C).
The strong positive effect of low O2 concentration on isoprene emission at high light/normal ambient CO2 concentration was paralleled by a significant increase in DMADP pool size under low O2 (Fig. 6). The enhancement in DMADP pool size was small, on the order of 10% in low O2 treatments, resulting in minor enhancement of isoprene emission (low light, high CO2; Fig. 5, B and C).
Figure 6.

Effect of O2 concentration on the chloroplastic DMADP pool size in the hybrid aspen leaves at a quantum flux density of 550 μmol m−2 s−1, ambient CO2 concentration of 390 μmol mol−1, and leaf temperature of 28°C to 30°C. DMADP pool size was estimated according to the in vivo method based on postillumination isoprene release (Rasulov et al., 2009).
Transient responses of isoprene emission to changing O2 concentration demonstrated that the response was not immediate but took approximately 2 to 3 min for full induction, consistent with the time required to alter the DMADP pool size (Fig. 7). Under low O2, isoprene emission could be further increased by reducing CO2 in agreement with the steady-state patterns (compare with Figs. 4A and 7), but again there was a certain delay of 2 to 3 min before reaching the steady state.
Figure 7.

Typical time courses of isoprene emission rate after changes in ambient O2 and CO2 concentrations in hybrid aspen leaves. The times of alteration of ambient air composition are indicated by arrows.
DISCUSSION
Is the Variation in Isoprene Emission Rate Driven by Changes in Substrate Availability after Light Changes?
To gain insight into the role of variations in DMADP concentrations in altering isoprene emission rate, we measured isoprene emission in parallel with the chloroplastic DMADP pool at different light intensities and CO2 and O2 concentrations. For the measurement of the in vivo DMADP pool, we used a nondestructive kinetic method based on integration of the postillumination isoprene emission (Rasulov et al., 2009). The dependence of isoprene emission on light intensity (Fig. 1) was reported already 40 years ago (Sanadze, 1969), but the mechanism of light-dependent regulation of isoprene emission is still a matter of discussion. After the discovery of the chloroplastic pathway of isoprene synthesis and the stromal and thylakoid-bound forms of IspS catalyzing the formation of isoprene from DMADP (Silver and Fall, 1991; Kuzma and Fall, 1993; Wildermuth and Fall, 1998), two major possibilities leading to light dependence have been suggested: light-induced activation of the rate-limiting IspS enzyme via changes in chloroplastic Mg2+ concentrations (Wildermuth and Fall, 1996; Fall and Wildermuth, 1998; Logan et al., 2000; Sasaki et al., 2005), and light-dependent changes in the availability of the substrate DMADP (Loreto and Sharkey, 1993). In the case of entirely enzymatic control, variation in isoprene emission rate would result from changes in enzyme activity at the saturating level of the DMADP pool. In the case of full substrate control, isoprene emission rate would vary in dependence on the changes in DMADP pool size at a constant level of enzyme activity. Although the positive correlations between isoprene synthase activity and isoprene emission rates in leaves of different ages and from different growth conditions have been found (Kuzma and Fall, 1993; Wildermuth and Fall, 1998; Lehning et al., 1999; Scholefield et al., 2004), to our knowledge, there is currently no experimental information on the activation of isoprene synthase in response to rapid light/dark changes. In fact, in vitro-measured IspS activity was similar between light- and dark-adapted leaves in Salix discolor (Wildermuth and Fall, 1998) and did not vary during the day in Quercus robur (Lehning et al., 1999). Nevertheless, transcription of the isoprene synthase gene is light dependent (Sasaki et al., 2005), and the transcription of the isoprene synthase gene and the amount of protein can vary during the day and between days (Loivamäki et al., 2007) and depending on long-term light availability (Schnitzler et al., 1997; Lehning et al., 1999), thereby affecting isoprene emission rate over longer time scales (Lehning et al., 1999; Loivamäki et al., 2007). For instance, it has been demonstrated that decreasing the light intensity by 50% by shading individual branches for 3 d decreased IspS activity by 60% (Lehning et al., 1999).
Different from the enzyme-level activation theory, the substrate-level control theory assumes that the variation in the synthesis of DMADP is responsible for the light dependence. In fact, the pool size of DMADP does vary and is commonly higher at higher quantum flux density (Brüggemann and Schnitzler, 2002; Rosenstiel et al., 2002; Mayrhofer et al., 2005; Kaiser et al., 2007; Rasulov et al., 2009). The level of chloroplastic DMADP is generally well below the level needed for the saturation of isoprene synthase (Nogués et al., 2006; Behnke et al., 2007; Rasulov et al., 2009). These observations of light-dependent variation in and overall low DMADP pool sizes demonstrate that substrate-level control of isoprene emission is principally possible. In our study, we observed a strong positive scaling of isoprene emission rate with DMADP pool size (Fig. 2). This relationship fitted close to a straight line, with only a small offset for residual isoprene emission and minor curvilinearity. Given the large Km value of approximately 3.8 μmol m−2 determined in vivo for Populus (Rasulov et al., 2009), substrate concentration was not saturating for isoprene synthesis in any part of this relationship. For mixed enzyme activity/substrate concentration control of isoprene emission, the emission rate would also less strongly respond to changes in substrate concentration at low light availability than at high light availability, because of low synthase activity under such conditions. In fact, the initial slope of isoprene emission rate versus DMADP pool size (Fig. 2) was actually somewhat larger at low DMADP than at high DMADP, as expected for typical Michaelis-Menten kinetics. Thus, this evidence suggests that the enzyme activity did not change over the range of light intensities applied. These results call into question the possibility that the activation state of isoprene synthase is regulated in a light-dependent manner in the time scale of tens of minutes. We conclude that changes in enzyme activity cannot be responsible for the light dependence of isoprene emission in the steady state.
Slow Turnover of DMADP Pool Size after Light Alteration
The numeric value of the slope of the isoprene emission rate versus DMADP pool size (Fig. 2) provides the rate constant (reciprocal of lifetime) for the conversion of the DMADP molecule to isoprene. We obtained a value of 0.034 s−1, corresponding to the average lifetime of DMADP of 30 to 40 s. Such a lifetime is rather long for a typical metabolic intermediate. However, the slow turnover of the DMADP pool observed in our study is in agreement with values recalculated from other reports: 0.01 s−1 (Brüggemann and Schnitzler, 2002; Rosenstiel et al., 2003) and 0.026 s−1 (Wildermuth and Fall, 1998). The lifetime of the DMADP pool is expected to depend reciprocally on the amount of the active enzyme, likely explaining the differences found in the literature.
The in vitro kinetic constant (turnover rate) of IspS is also rather small, 0.007 to 0.22 s−1 (Mayrhofer et al., 2005) or 0.08 s−1 (Sharkey et al., 2005). Thus, the kinetic constant of isoprene synthase and the average lifetime of the substrate DMADP are of the same order of magnitude. This suggests that the number of substrate molecules is of the same magnitude as the number of active sites of the fully activated enzyme. This correspondence between substrate and fully activated enzyme is akin to the situation with primary photosynthetic enzyme, Rubisco, and its substrate ribulose-1,5-bisphosphate (RuBP), where the number of active sites binding RuBP, up to 60 μmol m−2, forms a significant fraction of the total RuBP pool of 100 to 200 μmol m−2 (Eichelmann and Laisk, 1999; von Caemmerer, 2000). Provided that the affinity of the enzyme to the substrate is high, the comparable concentrations of enzyme sites and substrate molecules define a linear dependence of the reaction rate on the substrate concentration (Farquhar, 1979), as is visible in Figure 2.
The presence of a large DMADP pool size with slow turnover explains the significant release of isoprene for 3 to 5 min after switching off the light (Rasulov et al., 2009). Filling up this pool after a long-term dark period also likely explains the delay of approximately 3 to 5 min in isoprene emission after switching on the light (Fig. 3). Alternatively, light-dependent activation of isoprene synthase can be responsible for the delay in emission induction, but current analytical techniques cannot monitor isoprene synthase activity under transient conditions. Even if the early light activation of isoprene synthase activity is partly responsible for the delayed isoprene emission rate after switching on the light, current data suggest that changes in isoprene synthase activity were not responsible for differences in isoprene emission in the steady state, as the isoprene emission rate did not essentially vary at a given DMADP pool size. However, our observation of the absence of regulation of IspS activity during the steady-state light responses does not rule out the possibility of longer term regulation (e.g. changes resulting from the synthesis/degradation of IspS protein as stated above).
CO2 and O2 Responses of Isoprene Emission in Relation to Substrate Availability
Reduction in isoprene emission rate at higher CO2 concentrations has been reported in several studies (Loreto and Sharkey, 1990, 1993; Rosenstiel et al., 2003, 2006; Wilkinson et al., 2009), and this reduction is accompanied by decreased DMADP pool size (Rosenstiel et al., 2003, 2006). In our study, we further observed that the isoprene emission rate declines at lower CO2 concentrations (Fig. 4A). Despite the nonmonotonic CO2 response, the isoprene emission rate and DMADP pool size were strongly correlated throughout the entire CO2 response (Fig. 4B). Again, the relationship between isoprene emission rate and DMADP pool size observed throughout the entire CO2 response curve was almost linear (Fig. 4B), indicating that isoprene synthase activity did not change over the CO2 concentration range. The slope of isoprene emission versus DMADP pool size obtained, 0.029 s−1 (Fig. 4B), was only slightly lower than the slope obtained from the light responses (Fig. 2), demonstrating similar turnover rates of the DMADP pool during both light and CO2 responses. In addition, analogous positive scaling of isoprene emission with DMADP pool size was observed for different O2 concentrations (Figs. 5 and 6). Similar correspondence between isoprene emission rate and DMADP pool size has been observed even throughout leaf development from expanding to senescing leaves (Rasulov et al., 2009).
These data collectively suggest that changes in DMADP pool size rather than in the activation state of isoprene synthase were responsible for the observed CO2 and O2 effects on isoprene emission. Lack of CO2-related regulation of isoprene synthase activity has been reported by Kaiser et al. (2007) and Calfapietra et al. (2007), further confirming the importance of substrate effects in CO2 responses of isoprene emission.
Is the Control of Isoprene Synthesis by Substrate Due to Limited Availability of Carbon Skeletons or Energetic Cofactors?
The chloroplastic isoprene synthesis pathway (MEP pathway) begins with a condensation of glyceraldehyde 3-phosphate and pyruvate to form 1-deoxy-d-xylulose 5-phosphate. Pyruvate needed in this reaction is presumed to be formed by dephosphorylation of PEP. The availability of carbon skeletons for the pathway is determined by the availability of PEP and glyceraldehyde 3-phosphate. Farther along the pathway, NADPH and ATP are involved in the formation of a series of phosphorylated and reduced intermediates, such as MEP, 4-diphosphocytidyl-2-C-methyl-erythritol, and 2-C-methyl-erythritol 2,4-cyclophosphate, suggesting that the pathway can be controlled by the energetic cofactors NADPH and ATP, pending the effective Km values for energetic cofactors of the pathway. The energetic cofactors, as well as the carbon skeletons, are produced by leaf photosynthesis. Under nonstressed conditions, about 90% of the carbon in isoprene originates from the early products of photosynthesis (Delwiche and Sharkey, 1993; Karl et al., 2002; Funk et al., 2004).
The key question in understanding the environmental controls on isoprene emission is whether the rate of synthesis of DMADP is controlled by the availability of carbon skeletons or by energetic cofactors. Relying on the positive correlation between the pool size of ATP and isoprene emission rate at different external conditions, some authors have concluded that the synthesis of DMADP is limited by the availability of energetic cofactors (Sanadze and Baazov, 1985; Loreto and Sharkey, 1990, 1993; Niinemets et al., 1999), while others suggest that the supply of carbon skeletons is limiting (Brüggemann and Schnitzler, 2002; Rosenstiel et al., 2002; Mayrhofer et al., 2005). Especially in explaining the CO2 responses of isoprene emission, the supply of carbon skeletons has been considered to be the dominant limitation (Rosenstiel et al., 2003, 2006; Monson et al., 2007; Wilkinson et al., 2009).
Already the early measurements of isoprene emission suggested that only very low photosynthetic rates are needed to support a relatively fast isoprene emission rate (Tingey et al., 1981). Isoprene emission is significantly suppressed during the complete absence of CO2 and O2, but about 50 to 100 μmol CO2 mol−1 in the ambient air is sufficient to support a high rate of isoprene emission (Loreto and Sharkey, 1993). Our measurements demonstrating that the maximum isoprene emission rate occurred around the CO2 compensation point and decreased only to a minor extent when CO2 was completely removed from the ambient air (Fig. 4A), confirming the previous observations. This evidence indicates that carbon skeletons are sufficiently abundant for isoprene synthesis already when the net carbon supply is slightly positive. During the absence of photosynthesis in the dark, the isoprene precursor pools were depleted to the extent that a delay of about 5 min was needed to refill the MEP pathway intermediate pools, despite the fact that photosynthesis was already running fast (Fig. 3). These observations, along with the direct measurements of the carbon intermediate pools in the leaves (Badger et al., 1984), encourage us to focus on the regulation of the MEP pathway mainly by energetic cofactors.
An observation of an Emerson-type enhancement effect of isoprene emission when far-red and blue light were provided simultaneously (Sanadze and Baazov, 1985) has led to the suggestion that isoprene emission is controlled by energy supply. From the measurements of the ATP pool size, a direct relationship between ATP level and isoprene emission has been established (Loreto and Sharkey, 1993). The light-induced activation of isoprene emission observed in this work is consistent with the hypothesis of ATP-controlled DMADP pool size. In addition, saturation of isoprene emission rate at higher light than is needed to saturate photosynthesis (Fig. 1; Loreto and Sharkey, 1990, 1993; Harley et al., 1996, 1997) also supports the dominant role of energy supply. No further increase of the carbon pools is expected when photosynthetic CO2 uptake becomes light saturated, but the transthylakoid proton gradient, directly related to the ATP/ADP ratio, can still increase (Stitt, 1986; Maxwell et al., 1998).
CO2-Dependent Limitation of DMADP Synthesis: Energetic Cofactors or PEP?
How can the CO2 responses of isoprene emission and DMADP synthesis be explained on the basis of energetic cofactors? All known mechanisms explaining CO2 saturation of photosynthesis are based on limitation by ATP and/or NADPH. The theory of electron transport limitation of photosynthesis (Farquhar and von Caemmerer, 1982) assumed that the turnover of cytochrome b6f was the rate-limiting step of linear electron transport at high CO2 concentration, inducing a deficit in both ATP and NADPH. Experimental estimations of ATP level do demonstrate that its pool size is lower under high than under ambient CO2 (Cardon and Berry, 1992; Delwiche and Sharkey, 1993). The third player, phosphate (end-product synthesis) limitation, may sometimes occur at high CO2 concentrations (Sharkey, 1985). However, during phosphate limitation, the level of free inorganic phosphate, Pi, becomes limiting for ATP synthesis in the chloroplast, leading to the drop of ATP level, although the proton gradient may remain high (Harley and Sharkey, 1991). Thus, the phosphate limitation is actually also a situation with limited supply of energetic cofactors. As experimental evidence demonstrates, such reductions of ATP level due to limited Pi are associated with reduced isoprene emissions: addition of Man, which binds Pi, dramatically decreased both photosynthesis and isoprene emission (Monson and Fall, 1989).
On the other hand, at lower CO2, below the compensation point, photosynthetic electron transport rate becomes inhibited due to a lack of electron acceptors, resulting in reduced supply of ATP and NADPH (Laisk and Sumberg, 1994; Miyake et al., 2005). Such reduction in energy status can explain the suppression of DMADP synthesis under low CO2. Thus, we conclude that the available pieces of evidence collectively are consistent with the hypothesis that the level of energetic cofactors matches the rate of DMADP synthesis throughout the CO2 response curve (Fig. 4).
An alternative hypothesis to explain the inhibitory effect of high CO2 concentration on isoprene emission is the regulation of the isoprenoid synthesis pathway by the availability of cytosolic PEP that is presumed to be transported to chloroplasts by PEP/Pi translocator (Streatfield et al., 1999), where it is converted to the MEP pathway substrate pyruvate. As PEP is used in multiple processes, and the distribution of PEP between different pathways can depend on CO2 concentration, the competition for PEP can be responsible for the CO2 dependence of DMADP formation. In particular, it has been postulated that PEP carboxylation in cytosol, which likely strongly scales with CO2 concentration, draws increasingly more PEP away from the MEP pathway at higher CO2 concentrations (Rosenstiel et al., 2002, 2003, 2004; Affek and Yakir, 2003; Loreto et al., 2007). So far, the information regarding the regulation of PEP carboxylation in C3 plants is very limited, but there are several lines of evidence against the PEP control of DMADP synthesis under different CO2 levels. First, the rate of PEP carboxylation in C3 plants is overall very much slower than the rate of the photosynthetic metabolism, being at most 10% of the rate of photosynthesis (Roeske et al., 1989; Laisk and Sumberg, 1994). Second, cytosolic PEP that can be carboxylated by CO2 is a derivative from 3-phosphoglycerate (3-PGA) via 2-PGA. During photosynthesis at saturating CO2 concentrations, 3-PGA is available in great amounts (von Caemmerer and Edmondson, 1986) and the chloroplastic and cytosolic PGA pools are in equilibrium (for review, see Flügge and Heldt, 1991). Third, Ala, an amino acid formed mainly in chloroplasts from PEP via dephosphorylation and transamination (Schulze-Siebert et al., 1984), is synthesized at increased rates when photosynthesis becomes CO2 saturated (Platt et al., 1977; Keerberg et al., 1983; Murray et al., 1987), suggesting that PEP concentration in chloroplasts is high under CO2 saturation. In fact, the PEP pool is larger at higher CO2 concentrations (Platt et al., 1977). Fourth, given that PEP carboxylation scales positively with CO2 concentration, limited PEP availability would be unable to explain the decrease of isoprene emission and DMADP pool size at low CO2 concentrations (Fig. 4). Fifth, the PEP carboxylase competition hypothesis assumes that PEP and finally pyruvate for isoprene synthesis must come from cytosol. However, chloroplasts possess all enzymes of the glycolytic pathway that lead to the synthesis of pyruvate (Liedvogel and Bäuerle, 1986; Hoppe et al., 1993; Givan, 1999; Eastmond and Rawsthorne, 2000; Baud et al., 2007), although for some species, there is evidence that the chloroplasts of mesophyll cells may be deficient in terminal enzymes of glycolysis, such as enolase and phosphoglyceromutase (Plaxton, 1996). Alternatively, pyruvate may be formed by β-elimination of phosphate from the carbanion intermediate in the Rubisco reaction (Andrews and Kane, 1991) in sufficient amounts needed for isoprene synthesis (Sharkey et al., 1991a). In fact, the early experiments of Sanadze and colleagues with isolated chloroplasts have demonstrated that cytosolic PEP is not needed for high isoprene fluxes (Sanadze and Dzhaiani, 1972; Mgalobilishvili et al., 1978). High isotopic labeling observed in 13CO2 addition experiments (see above) is also not consistent with the extrachloroplastic origin of a large fraction of PEP used in isoprene synthesis. Depending on the contributions of chloroplastic and cytosolic PEP pools, PEP carboxylase should not necessarily interfere with DMADP synthesis at all. Finally, we note that PEP carboxylation is activated in parallel with light activation of photosynthesis (Samaras and Manetas, 1988; Pfeffer and Peisker, 1998). If PEP carboxylase would efficiently compete for PEP, isoprene synthesis rate would not increase with light intensity, contrary to the results obtained by us (Fig. 1) and others.
On the basis of this evidence, we conclude that the regulation of DMADP synthesis by energetic cofactors is the most likely explanation for the CO2 dependence of isoprene synthesis. However, the conclusion that isoprene emission is controlled at the level of energetic cofactors is valid for short-term (tens of minutes) regulations. During long exposures of plants to unfavorable conditions like low temperature (Sharkey et al., 2008), during the development of young leaves (Mayrhofer et al., 2005; Wiberley et al., 2008), during adjustments to changed light intensity (Lehning et al., 1999; Sasaki et al., 2005), and during other stresses (Brilli et al., 2007), the control of isoprene emission may shift over to the IspS transcription and to carbon sources (Fortunati et al., 2008).
How Does O2 Affect Isoprene Emission?
Our study (Figs. 5–7) confirms the previous observations of the enhancement of isoprene emission at low concentrations of O2 (Sanadze and Tarhnishvili, 1986; Loreto and Sharkey, 1990). Our results here show that the enhancement of isoprene emission at low O2 levels is the result of an increase in the DMADP pool size by about 35% (Fig. 6). The larger DMADP pool was not the result of increased carbon supply from the carbon reduction-oxidation cycles (CROC; CO2 assimilation and photorespiration), since isoprene emission increased after CO2 was removed from the ambient air in the presence of 2% O2. Under such conditions, CROC activity could only decrease. Although the enhanced isoprene emission in CO2-free atmosphere may not be persistent, as the depletion of CROC carbon pools would finally neutralize the positive effect (Loreto and Sharkey, 1990), the immediate positive response of isoprene emission on the removal of CO2 (Fig. 7) cannot be explained on the basis of carbon limitation.
The lack of carbon limitation under low O2 is also confirmed by 13C labeling experiments (Karl et al., 2002). In these experiments, the labeling of isoprene decreased but isoprene emission rate increased under low O2 (Karl et al., 2002). Thus, PEP labeling decreased due to decreased PGA level in the absence of RuBP oxygenation, but an unlabeled cytosolic PEP source immediately replaced the PEP of photosynthetic origin. Thus, there was no evidence of limitation of the MEP pathway by carbon sources. The lack of control of isoprene emission by carbon pools was demonstrated also in experiments where the addition of endogenous deoxyxylulose did not enhance isoprene emission under low O2 concentration (Wolfertz et al., 2003).
Taken together, the simplest explanation of our results is consistent with the ATP-related regulation of isoprene synthesis. The total turnover rate of CROC significantly decreased after RuBP oxidation was suppressed at the low O2 concentration. This resulted in increased pressure of the energetic cofactors ATP and NADPH, as reflected in enhanced DMADP synthesis and isoprene emission. The effect was smaller under lower light (Fig. 5B), where the increase of electron pressure was less due to an overall lower electron transport rate. Analogously, high CO2 can inhibit photosynthetic electron transport due to phosphate sequestration (Sharkey et al., 1986; Harley and Sharkey, 1991), resulting in a small effect of low O2 on isoprene emission (Fig. 5C). Even under ambient CO2, but at longer time scales (more than 30 min), photosynthetic electron transport rate can become inhibited under low O2 as the result of feedback inhibition due to sequestered Pi, and the isoprene emission rate can decline after the initial 20 to 30 min of increase under low O2 (Monson and Fall, 1989; Loreto and Sharkey, 1990).
CONCLUSION
We have shown that foliage isoprene emission is tightly related to photosynthesis via the supply of energetic cofactors. These data do not support short-term enzymatic limitation of the MEP pathway. Isoprene synthase usually operates with a nonsaturating pool of its substrate DMADP (Figs. 2 and 4B; Brüggemann and Schnitzler, 2002), whose concentration is even less than the concentration of the enzyme active sites (Rasulov et al., 2009). Even very low photosynthesis is sufficient to saturate the need for carbon skeletons for isoprene emission (Tingey et al., 1981; Wolfertz et al., 2003). Therefore, the variation in isoprene emission rates due to changes in ambient air composition occurs differently from carbon uptake rates. In contrast, the substrate DMADP for IspS is almost proportionally dependent on energetic and reductive equivalents, suggesting that the effective Km values for ATP and NADPH are large for the isoprenoid synthesis pathway. For developing process-based models of isoprene emission, the effective Km values for ATP and NADPH should be determined for the entire series of reactions leading to DMADP.
MATERIALS AND METHODS
Plants and Growth Conditions
One-year-old seedlings of hybrid aspen (Populus tremula × P. tremuloides) clone 200 (for a description of the genotype, see Vahala et al., 2003) were grown in a Percival AR-95 HIL growth chamber (CLF PlantClimatics) under photosynthetic quantum flux density of 500 μmol m−2 s−1 (14-h d) and relative humidity of 60%. Air temperature in the growth chamber was maintained at 26°C during the day and 20°C during the night. The plants were grown in plastic pots filled with 4 kg of a sand and peat mixture (1:1) and watered daily to field capacity with distilled water. Once a week, macroelements according to Knopp's and microelements according to Hoagland's standard solutions were added to the irrigation water.
Gas-Exchange System
The measurements were accomplished with a custom-made open gas-exchange system described in detail by Rasulov et al. (2009). In short, the measurement chamber of 3 dm3 was made of glass and was equipped with a fan for efficient air mixing in the chamber. Other parts of the system were made of Teflon (DuPont) or stainless steel. Four halogen lamps positioned at different sides of the chamber provided essentially uniform illumination of all leaves in the chamber. Light intensity could be dynamically regulated between 0 and 550 μmol m−2 s−1 by changing the lamp voltage. The air flow rate through the chamber was 3 dm3 min−1, resulting in the response half-time of approximately 40 s. Either the outside air low in pollutants (O3 concentrations < 20 nmol mol−1, NO < 5 nmol mol−1, isoprene concentration below the detection limit) taken through a 50-dm3 buffer volume (experiments with constant ambient O2 and CO2 concentrations) or air mixed from pure N2, O2, and CO2 using capillary mixers (Laisk and Oja, 1998) was used. In all cases, a constant air humidity of 60% was achieved using a custom-made thermostatted humidifier. The temperature inside the chamber was continuously measured with a negative temperature coefficient thermistor (model -001; RTI Electronics). Leaf temperature determined from leaf energy balance was within ±1°C of the air temperature in the chamber. In all experiments, leaf temperature was maintained between 28°C and 30°C. Experiments with widely varying temperature demonstrated that fluctuations of leaf temperature over this limited range did not qualitatively affect any of the relationships reported (data not shown). The gas-exchange system was computer operated using an A/D converter board ADIO 1600 (Kontron) and custom-made computer software.
Determination of Net Assimilation, Transpiration, and Isoprene Emission Rates
All measurements were conducted with fully mature leaves (25–35 d after bud burst) with stable photosynthetic capacity and isoprene emission rate (for age-dependent modifications in isoprene emission rate, see Monson et al., 1994). In our previous study, we have observed isoprene emission rates of up to approximately 35 nmol m−2 s−1 immediately after the cessation of leaf expansion (Rasulov et al., 2009). As these peak rates were supported only for a limited period of time, 2 to 3 d (data not shown), we used the fully mature nonsenescent leaves with stable (at least for 10 d) assimilation rates and isoprene emission potentials of approximately 25 nmol m−2 s−1.
Incoming and outgoing air CO2 and water vapor concentrations were measured with an infrared gas analyzer (LI-6262; Li-Cor). Isoprene concentration was measured with the Fast Isoprene Analyzer (Hills Scientific) specifically modified to optimize the sensitivity and stability of isoprene concentration measurements. This device is based on counting the photons emitted as the result of reactions between ozone and isoprene, resulting in chemiluminescence as the isoprene degradation product, electronically excited formaldehyde, reaches the ground state (Hills and Zimmerman, 1990). For this purpose, ozone generated from molecular O2 is continuously added to the gas stream entering the measurement cell. However, a significant background of photons is generated by direct ozone-ozone recombination reaction. The latter reaction is sensitively inhibited by water vapor, making the background count rate dependent on gas humidity. Since the ozone-ozone reaction is proportional to the square of ozone concentration, but the ozone-isoprene reaction follows the first-order kinetics, the humidity-dependent background was optimized by decreasing ozone concentration in the cell. For this purpose, we adjusted the O2 flow rate to 0.4 dm3 min−1, while the measurement gas flow was 0.8 dm3 min−1. For optimization and maximum stabilization of the background count level, the dew point of the gas entering the isoprene analyzer was set at 23°C using a custom-made humidifier. The isoprene analyzer was regularly calibrated with a standard gas (5.74 μmol mol−1 isoprene in N2).
Light response curves were measured starting with the highest light and decreasing stepwise the light intensity. Preliminary experiments demonstrated that the isoprene emission rate achieved at each light level did not depend on the order of measurements (i.e. there was no significant difference whether the measurements were started from the lowest or from the highest light). At each light level, the measurements were taken after steady-state values of isoprene emission and net assimilation rates had been observed, usually 15 to 20 min after changing the conditions. The light (Q) responses of net assimilation and isoprene emission were fitted by a hyperbolic relationship previously used to simulate photosynthetic characteristics and isoprene emission as a function of light (Harley and Tenhunen, 1991; Guenther et al., 1993):
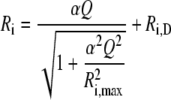 |
(1) |
where Ri is either the isoprene emission or the net assimilation rate, Ri,max is the light-saturated value, Ri,D is the rate in the darkness, and α is the initial quantum yield. As isoprene emission potentials are commonly assessed at Q = 1,000 μmol m−2 s−1 (Guenther et al., 1993), Equation 1 was also used to predict the emission rates and net assimilation rates at this Q. This allowed us to determine how far the measurements were from full light saturation. In addition to the response curve measurements, dark/light activation of isoprene emission was studied after a 10-h dark period. The time-dependent activations of isoprene emission, photosynthesis, and transpiration rates were fitted by single-exponential and double-exponential functions to gain information on the rate constants (k). For the single-exponential function, the rate, Ri, varies in time (t) as:
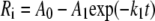 |
(2) |
while for the double exponential function:
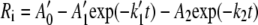 |
(3) |
where A0, A1, A0′, A1′, and A2 are the scaling constants and k1, k1′, and k2 are the rate constants.
To measure the CO2 responses, the leaf was stabilized at an ambient CO2 concentration of 390 μmol mol−1, and after the steady-state net assimilation and isoprene emission rates had been reached under the ambient CO2 concentrations, CO2 concentration was changed to the target value. Again, enough time was allowed to attain the new steady-state conditions, and the rates of isoprene emission and net assimilation were recorded. After the measurements at given CO2 concentrations, the CO2 concentration was switched back again to 390 μmol mol−1 and the leaf was stabilized until previous steady-state values were reached before taking the next point of the CO2 response curve. An analogous stabilization procedure was followed for O2 responses.
Each measurement at different light, CO2, and O2 levels was combined with postillumination isoprene release measurements. After reaching the steady-state value under given environmental conditions, light was switched off and postillumination isoprene release was measured for 5 min to estimate the in vivo DMADP pool size as detailed below.
In all cases, at least five replicate experiments with different plants were conducted and averages were calculated. After each experiment, leaves were scanned and leaf area was calculated. All gas-exchange characteristics were calculated per unit of leaf area according to von Caemmerer and Farquhar (1981).
In Vivo Estimation of the DMADP Pool Responsible for Isoprene Formation
The DMADP pool responsible for isoprene emission (chloroplastic isoprene pool) was measured kinetically in intact plants as described by Rasulov et al. (2009). These measurements are based on the observation that after darkening, isoprene emission continues for 3 to 5 min (Monson and Fall, 1989; Rasulov et al., 2009). Assuming that the synthesis of DMADP stops rapidly (within seconds) in the dark due to the lack of ATP and NADPH, and the isoprene emission can continue in the darkness only at the expense of the existing DMADP pool, the integral of the isoprene emission during the 3 to 5 min after the darkening gives the size of the DMADP pool responsible for isoprene emission. The pool size of DMADP estimated this way is in good correspondence with the chemical estimation of chloroplastic DMADP pool size (difference in DMADP pools in light- and dark-adapted leaf samples), although the chemical estimations provide approximately 10% larger estimates, possibly reflecting the DMADP that is used in other chloroplastic reactions occurring simultaneously with isoprene formation such as carotenoid biosynthesis (Rasulov et al., 2009). The advantage of the in vivo method is that the DMADP pool size responsible for isoprene emission can be repeatedly estimated for the same leaves, making it possible to investigate the influence of environmental drivers on and linking the emission measurements with the DMADP pool size for the same leaves. In practice, in vivo DMADP pool size estimations also require consideration of the response time of the gas-exchange system, as explained in detail by Rasulov et al. (2009) and done in this study.
Simultaneous measurements of isoprene emission rate and DMADP pool size were further used to determine the turnover rate of DMADP pool size (s−1) as the initial slope of isoprene emission rate versus the DMADP pool size.
This study was supported by the Human Frontiers of Science Program, the Estonian Science Foundation (grant nos. 6607, 7272, and 7645), the Estonian Academy of Sciences, and the Estonian Ministry of Education and Science (grant nos. SF1090065s07 and SF010045s08).
The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy described in the Instructions for Authors (www.plantphysiol.org) is: Ülo Niinemets (ylo.niinemets@emu.ee).
References
- Affek HP, Yakir D (2003) Natural abundance carbon isotope composition of isoprene reflects incomplete coupling between isoprene synthesis and photosynthetic carbon flow. Plant Physiol 131 1727–1736 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrews TJ, Kane HJ (1991) Pyruvate is a by-product of catalysis of ribulosebisphosphate carboxylase/oxygenase. J Biol Chem 266 9447–9452 [PubMed] [Google Scholar]
- Arneth A, Monson RK, Schurgers G, Niinemets Ü, Palmer PI (2008) Why are estimates of global isoprene emissions so similar (and why is this not so for monoterpenes)? Atmos Chem Phys 8 4605–4620 [Google Scholar]
- Badger MR, Sharkey TD, von Caemmerer S (1984) The relationship between steady-state gas exchange of bean leaves and the levels of carbon-reduction-cycle intermediates. Planta 160 305–313 [DOI] [PubMed] [Google Scholar]
- Baud S, Wuillème S, Dubreucq B, de Almeida A, Vuagnat C, Lepiniec L, Miquel M, Rochat C (2007) Function of plastidial pyruvate kinases in seeds of Arabidopsis thaliana. Plant J 52 405–419 [DOI] [PubMed] [Google Scholar]
- Behnke K, Ehlting B, Teuber M, Bauerfeind M, Louis S, Hänsch R, Polle A, Bohlmann J, Schnitzler JP (2007) Transgenic, non-isoprene emitting poplars don't like it hot. Plant J 51 485–499 [DOI] [PubMed] [Google Scholar]
- Brilli F, Barta C, Fortunati A, Lerdau M, Loreto F, Centritto M (2007) Response of isoprene emission and carbon metabolism to drought in white poplar (Populus alba) saplings. New Phytol 175 244–254 [DOI] [PubMed] [Google Scholar]
- Brüggemann N, Schnitzler JP (2002) Diurnal variation of dimethylallyl diphosphate concentrations in oak (Quercus robur) leaves. Physiol Plant 115 190–196 [DOI] [PubMed] [Google Scholar]
- Calfapietra C, Wiberley AE, Falbel TG, Linskey AR, Mugnozza GS, Karnosky DF, Loreto F, Sharkey TD (2007) Isoprene synthase expression and protein levels are reduced under elevated O3 but not under elevated CO2 (FACE) in field-grown aspen trees. Plant Cell Environ 30 654–661 [DOI] [PubMed] [Google Scholar]
- Cardon ZG, Berry J (1992) Effects of O2 and CO2 concentration on the steady-state fluorescence yield of single guard cell pairs in intact leaf discs of Tradescantia albiflora. Plant Physiol 99 1238–1244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darbah J, Sharkey T, Karnosky D (2008) Isoprene emission and carbon dioxide protect aspen leaves from heat stress. Nat Proc http://precedings.nature.com/documents/1533/version/1 (July 1, 2009)
- Delwiche CF, Sharkey TD (1993) Rapid appearance of 13C in biogenic isoprene when 13CO2 is fed to intact leaves. Plant Cell Environ 16 587–591 [Google Scholar]
- Eastmond PJ, Rawsthorne S (2000) Coordinate changes in carbon partitioning and plastidial metabolism during the development of oilseed rape embryos. Plant Physiol 122 767–774 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eichelmann H, Laisk A (1999) Ribulose-1,5-bisphosphate carboxylase/oxygenase content, assimilatory charge, and mesophyll conductance in leaves. Plant Physiol 119 179–189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisenreich W, Rohdich F, Bacher A (2001) Deoxyxylulose phosphate pathway to terpenoids. Trends Plant Sci 6 78–84 [DOI] [PubMed] [Google Scholar]
- Fall R (1999) Biogenic emissions of volatile organic compounds from higher plants. In CN Hewitt, ed, Reactive Hydrocarbons in the Atmosphere. Academic Press, San Diego, pp 41–95
- Fall R, Wildermuth MC (1998) Isoprene synthase: from biochemical mechanism to emission algorithm. J Geophys Res 103 25599–25609 [Google Scholar]
- Fang C, Monson RK, Cowling EB (1996) Isoprene emission, photosynthesis, and growth in sweetgum (Liquidambar styraciflua) seedlings exposed to short- and long-term drying cycles. Tree Physiol 16 441–446 [DOI] [PubMed] [Google Scholar]
- Farquhar GD (1979) Models describing the kinetics of ribulose bisphosphate carboxylase-oxygenase. Arch Biochem Biophys 193 456–468 [DOI] [PubMed] [Google Scholar]
- Farquhar GD, von Caemmerer S (1982) Modeling of photosynthetic response to environmental conditions. In OL Lange, PS Nobel, CB Osmond, H Ziegler, eds, Physiological Plant Ecology, Vol II. Springer-Verlag, Berlin, pp 549–588
- Farquhar GD, von Caemmerer S, Berry JA (1980) A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 149 78–90 [DOI] [PubMed] [Google Scholar]
- Flügge UI, Heldt HW (1991) Metabolite translocators of the chloroplast envelope. Annu Rev Plant Physiol Plant Mol Biol 42 129–144 [Google Scholar]
- Funk JL, Mak JE, Lerdau MT (2004) Stress-induced changes in carbon sources for isoprene production in Populus deltoides. Plant Cell Environ 27 747–755 [Google Scholar]
- Fortunati A, Barta C, Brilli F, Centritto M, Zimmer I, Schnitzler JP, Loreto F (2008) Isoprene emission is not temperature-dependent during and after severe drought stress: a physiological and biochemical analysis. Plant J 55 687–697 [DOI] [PubMed] [Google Scholar]
- Givan CV (1999) Evolving concepts in plant glycolysis: two centuries of progress. Biol Rev Camb Philos Soc 74 277–309 [Google Scholar]
- Grote R, Niinemets Ü (2008) Modeling volatile isoprenoid emission: a story with split ends. Plant Biol 10 8–28 [DOI] [PubMed] [Google Scholar]
- Guenther A, Karl T, Harley P, Wiedinmyer C, Palmer PI, Geron C (2006) Estimates of global terrestrial isoprene emissions using MEGAN (Model of Emissions of Gases and Aerosols from Nature). Atmos Chem Phys 6 3181–3210 [Google Scholar]
- Guenther AB, Zimmerman PR, Harley PC, Monson RK, Fall R (1993) Isoprene and monoterpene emission rate variability: model evaluations and sensitivity analyses. J Geophys Res 98 12609–12617 [Google Scholar]
- Harley P, Guenther A, Zimmerman P (1996) Effects of light, temperature and canopy position on net photosynthesis and isoprene emission from sweetgum (Liquidambar styraciflua) leaves. Tree Physiol 16 25–32 [DOI] [PubMed] [Google Scholar]
- Harley P, Guenther A, Zimmerman P (1997) Environmental controls over isoprene emission in deciduous oak canopies. Tree Physiol 17 705–714 [DOI] [PubMed] [Google Scholar]
- Harley PC, Sharkey TD (1991) An improved model of C3 photosynthesis at high CO2: reversed O2 sensitivity explained by lack of glycerate reentry into the chloroplast. Photosynth Res 27 169–178 [DOI] [PubMed] [Google Scholar]
- Harley PC, Tenhunen JD (1991) Modeling the photosynthetic response of C3 leaves to environmental factors. In KJ Boote, ed, Modeling Crop Photosynthesis: From Biochemistry to Canopy. Agronomy and Crop Science Society of America, Madison, WI, pp 17–39
- Hewitt CN, Monson RK, Fall R (1990) Isoprene emissions from the grass Arundo donax L. are not linked to photorespiration. Plant Sci 66 139–144 [Google Scholar]
- Hills AJ, Zimmerman PR (1990) Isoprene measurement by ozone-induced chemiluminescence. Anal Chem 62 1055–1060 [Google Scholar]
- Hoppe P, Heintze A, Riedel A, Creuzer C, Schultz G (1993) The plastidic 3-phosphoglycerate → acetyl-CoA pathway in barley leaves and its involvement in the synthesis of amino acids, plastidic isoprenoids and fatty acids during chloroplast development. Planta 190 253–262 [Google Scholar]
- Kaiser AJ, Cinege G, Louis S, Schnitzler JP (2007) Effects of varying ambient CO2-concentrations on isoprene biosynthesis in shoot cultures of Populus x canescens. In M Staudt, ed, VOCBAS Science Conference (October 2–5) Montpellier, France. CNRS, Montpellier, France, p 52
- Karl T, Fall R, Rosenstiel TN, Prazeller P, Larsen B, Seufert G, Lindinger W (2002) On-line analysis of the 13CO2 labeling of leaf isoprene suggests multiple subcellular origins of isoprene precursors. Planta 215 894–905 [DOI] [PubMed] [Google Scholar]
- Keeling R, Shertz S (1992) Seasonal and interannual variations in atmospheric oxygen and implications for the global carbon cycle. Nature 358 723–727 [Google Scholar]
- Keerberg O, Keerberg H, Pärnik T, Viil J, Vark E (1983) The metabolism of photosynthetically assimilated 14CO2 under different concentrations of carbon dioxide. Int J Appl Radiat Isot 34 861–864 [Google Scholar]
- Kump LR (2008) The rise of atmospheric oxygen. Nature 451 277–278 [DOI] [PubMed] [Google Scholar]
- Kuzma J, Fall R (1993) Leaf isoprene emission rate is dependent on leaf development and the level of isoprene synthase. Plant Physiol 101 435–440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laisk A, Oja V (1998) Dynamics of Leaf Photosynthesis: Rapid-Response Measurements and Their Interpretations. CSIRO Publishing, Canberra, Australia
- Laisk A, Sumberg A (1994) Partitioning of the leaf CO2 exchange into components using CO2 exchange and fluorescence measurements. Plant Physiol 106 689–695 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehning A, Zimmer I, Steinbrecher R, Brüggemann N, Schnitzler JP (1999) Isoprene synthase activity and its relation to isoprene emission in Quercus robur L. leaves. Plant Cell Environ 22 495–504 [Google Scholar]
- Lerdau M (2007) A positive feedback with negative consequences. Science 316 212–213 [DOI] [PubMed] [Google Scholar]
- Liedvogel B, Bäuerle R (1986) Fatty-acid synthesis in chloroplasts from mustard (Sinapis alba L.) cotyledons: formation of acetyl coenzyme A by intraplastid glycolytic enzymes and a pyruvate dehydrogenase complex. Planta 169 481–489 [DOI] [PubMed] [Google Scholar]
- Litvak ME, Loreto F, Harley PC, Sharkey TD, Monson RK (1996) The response of isoprene emission rate and photosynthetic rate to photon flux and nitrogen supply in aspen and white oak trees. Plant Cell Environ 19 549–559 [Google Scholar]
- Logan BA, Monson RK, Potosnak MJ (2000) Biochemistry and physiology of foliar isoprene production. Trends Plant Sci 5 477–481 [DOI] [PubMed] [Google Scholar]
- Loivamäki M, Louis S, Cinege G, Zimmer I, Fischbach RJ, Schnitzler JP (2007) Circadian rhythms of isoprene biosynthesis in grey poplar leaves. Plant Physiol 143 540–551 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loreto F, Centritto M, Barta C, Calfapietra C, Fares S, Monson RK (2007) The relationship between isoprene emission rate and dark respiration rate in white poplar (Populus alba L.) leaves. Plant Cell Environ 30 662–669 [DOI] [PubMed] [Google Scholar]
- Loreto F, Mannozzi M, Maris C, Nascetti P, Ferranti F, Pasqualini S (2001) Ozone quenching properties of isoprene and its antioxidant role in leaves. Plant Physiol 126 993–1000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loreto F, Sharkey TD (1990) A gas-exchange study of photosynthesis and isoprene emission in Quercus rubra L. Planta 182 523–531 [DOI] [PubMed] [Google Scholar]
- Loreto F, Sharkey TD (1993) On the relationship between isoprene emission and photosynthetic metabolites under different environmental conditions. Planta 189 420–424 [DOI] [PubMed] [Google Scholar]
- Loreto F, Velikova V (2001) Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol 127 1781–1787 [PMC free article] [PubMed] [Google Scholar]
- Magel E, Mayrhofer S, Müller A, Zimmer I, Hampp R, Schnitzler JP (2006) Photosynthesis and substrate supply for isoprene biosynthesis in poplar leaves. Atmos Environ 40 S138–S151 [Google Scholar]
- Maxwell K, Badger MR, Osmond CB (1998) A comparison of CO2 and O2 exchange patterns and the relationship with chlorophyll fluorescence during photosynthesis in C3 and CAM plants. Aust J Plant Physiol 25 45–52 [Google Scholar]
- Mayrhofer S, Teuber M, Zimmer I, Louis S, Fischbach RJ, Schnitzler JP (2005) Diurnal and seasonal variation of isoprene biosynthesis-related genes in grey poplar leaves. Plant Physiol 139 474–484 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mgalobilishvili MP, Khetsuriani ND, Kalandadze AN, Sanadze GA (1978) Localization of isoprene biosynthesis in poplar leaf chloroplasts. Fiziol Rast 25 1055–1061 [Google Scholar]
- Miyake C, Miyata M, Shinzaki Y, Tomizawa K (2005) CO2 response of cyclic electron flow around PSI (CEF-PSI) in tobacco leaves: relative electron fluxes through PSI and PSII determine the magnitude of non-photochemical quenching (NPQ) of Chl fluorescence. Plant Cell Physiol 46 629–637 [DOI] [PubMed] [Google Scholar]
- Monson RK, Fall R (1989) Isoprene emission from aspen leaves: influence of environment and relation to photosynthesis and photorespiration. Plant Physiol 90 267–274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monson RK, Guenther AB, Fall R (1991) Physiological reality in relation to ecosystem- and global-level estimates of isoprene emission. In TD Sharkey, EA Holland, HA Mooney, eds, Trace Gas Emissions by Plants. Academic Press, San Diego, pp 185–207
- Monson RK, Harley PC, Litvak ME, Wildermuth M, Guenther AB, Zimmerman PR, Fall R (1994) Environmental and developmental controls over the seasonal pattern of isoprene emission from aspen leaves. Oecologia 99 260–270 [DOI] [PubMed] [Google Scholar]
- Monson RK, Trahan N, Rosenstiel TN, Veres P, Moore D, Wilkinson M, Norby RJ, Volder A, Tjoelker MG, Briske DD, et al (2007) Isoprene emission from terrestrial ecosystems in response to global change: minding the gap between models and observations. Philos Trans R Soc Lond A 365 1677–1695 [DOI] [PubMed] [Google Scholar]
- Murray AJS, Blackwell RD, Joy KW, Lea PJ (1987) Photorespiratory N donors, aminotransferase specificity and photosynthesis in a mutant of barley deficient in serine: glyoxylate aminotransferase activity. Planta 172 106–113 [DOI] [PubMed] [Google Scholar]
- Niinemets Ü, Tenhunen JD, Harley PC, Steinbrecher R (1999) A model of isoprene emission based on energetic requirements for isoprene synthesis and leaf photosynthetic properties for Liquidambar and Quercus. Plant Cell Environ 22 1319–1336 [Google Scholar]
- Nogués I, Brilli F, Loreto F (2006) Dimethylallyl diphosphate and geranyl diphosphate pools of plant species characterized by different isoprenoid emissions. Plant Physiol 141 721–730 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeffer M, Peisker M (1998) CO2 gas exchange and phosphoenolpyruvate carboxylase activity in leaves of Zea mays L. Photosynth Res 58 281–291 [Google Scholar]
- Platt SG, Plaut Z, Bassham JA (1977) Steady-state photosynthesis in alfalfa leaflets: effects of carbon dioxide concentration. Plant Physiol 60 230–234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plaxton WC (1996) The organization and regulation of plant glycolysis. Annu Rev Plant Physiol Plant Mol Biol 47 185–214 [DOI] [PubMed] [Google Scholar]
- Potosnak MJ (2002) Effects of growth carbon dioxide concentration on isoprene emissions from plants. PhD thesis. Columbia University, New York
- Rasulov B, Copolovici L, Laisk A, Niinemets Ü (2009) Postillumination isoprene emission: in vivo measurements of dimethylallyldiphosphate pool size and isoprene synthase kinetics in aspen leaves. Plant Physiol 149 1609–1618 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roeske CA, Widholm JM, Ogren WL (1989) Photosynthetic carbon metabolism in photoautotrophic cell suspension cultures grown at low and high CO2. Plant Physiol 91 1512–1519 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenstiel TN, Ebbets AL, Khatri WC, Fall R, Monson RK (2004) Induction of poplar leaf nitrate reductase: a test of extrachloroplastic control of isoprene emission rate. Plant Biol 6 12–21 [DOI] [PubMed] [Google Scholar]
- Rosenstiel TN, Fall R, Monson RK (2006) Substrate versus enzyme controls over isoprene emission from poplar leaves grown at elevated CO2 concentration. Geophysical Research Abstracts 8 01562 [Google Scholar]
- Rosenstiel TN, Fisher AJ, Fall R, Monson RK (2002) Differential accumulation of dimethylallyl diphosphate in leaves and needles of isoprene- and methylbutenol-emitting and nonemitting species. Plant Physiol 129 1276–1284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenstiel TN, Potosnak MJ, Griffin KL, Fall R, Monson RK (2003) Increased CO2 uncouples growth from isoprene emission in an agriforest ecosystem. Nature 421 256–259 [DOI] [PubMed] [Google Scholar]
- Samaras Y, Manetas Y (1988) Regulation of C4-phosphoenolpyruvate carboxylase activity by ambient CO2. Photosynth Res 18 299–305 [DOI] [PubMed] [Google Scholar]
- Sanadze GA (1966) Biosynthesis and light-dependent isoprene emission from leaves. DSc dissertation. Institute of Plant Biology, Moscow
- Sanadze GA (1969) Light-dependent excretion of molecular isoprene. Prog Photosynth Res 2 701–707 [Google Scholar]
- Sanadze GA (1990) The principle scheme of photosynthetic carbon conversion in cells of isoprene releasing plants. In M Baltscheffsky, ed, Current Research in Photosynthesis, Vol IV. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp 231–237
- Sanadze GA (2004) Biogenic isoprene (a review). Russ J Plant Physiol 51 729–741 [Google Scholar]
- Sanadze GA, Baazov DI (1985) Usileniye fotosinteza i vyhoda izoprena u listyev topolya. (Enhancement of photosynthesis and isoprene emission in poplar leaves). Soobshch Akad Nauk Gruz SSR 118 597–600 [Google Scholar]
- Sanadze GA, Dzhaiani GI (1972) Carbohydrate partition within isoprene molecule from photosynthetically assimilated CO2. Fiziol Rast 19 1082–1089 [Google Scholar]
- Sanadze GA, Dzhaiani GI, Tevzadze IM (1972) Incorporation into the isoprene molecule of carbon from 13CO2 assimilated during photosynthesis. Sov Plant Physiol 19 17–20 [Google Scholar]
- Sanadze GA, Tarhnishvili GM (1986) Deistviye molekulyarnogo kisloroda na protsess biosinteza izoprena v listyah pri nasyshtshayushtshei fotosintez intensivnosti sveta. (Effect of molecular oxygen on the biosynthesis of isoprene in leaves under conditions of photosynthesis-saturating light intensity). Dokl Akad Nauk SSSR 286 501–503 [Google Scholar]
- Sasaki K, Ohara K, Yazaki K (2005) Gene expression and characterization of isoprene synthase from Populus alba. FEBS Lett 579 2514–2518 [DOI] [PubMed] [Google Scholar]
- Schnitzler JP, Lehning A, Steinbrecher R (1997) Seasonal pattern of isoprene synthase activity in Quercus robur leaves and its significance for modeling isoprene emission rates. Bot Acta 110 240–243 [Google Scholar]
- Scholefield PA, Doick KJ, Herbert B, Hewitt CN, Schnitzler JP, Pinelli P, Loreto F (2004) Impact of rising CO2 on VOC emissions: isoprene emission from Phragmites australis growing at elevated CO2 in a natural carbon dioxide spring. Plant Cell Environ 27 393–401 [Google Scholar]
- Schulze-Siebert D, Heineke D, Scharf H, Schultz G (1984) Pyruvate-derived amino acids in spinach chloroplasts: synthesis and regulation during photosynthetic carbon metabolism. Plant Physiol 76 465–471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharkey TD (1985) Photosynthesis in intact leaves of C3 plants: physics, physiology and rate limitations. Bot Rev 51 53–105 [Google Scholar]
- Sharkey TD (1996) Emission of low molecular mass hydrocarbons from plants. Trends Plant Sci 1 78–82 [Google Scholar]
- Sharkey TD, Loreto F, Delwiche CF (1991. a) The biochemistry of isoprene emission from leaves during photosynthesis. In TD Sharkey, EA Holland, HA Mooney, eds, Trace Gas Emissions by Plants. Academic Press, San Diego, pp 153–184
- Sharkey TD, Loreto F, Delwiche CF (1991. b) High carbon dioxide and sun/shade effects on isoprene emission from oak and aspen tree leaves. Plant Cell Environ 14 333–338 [Google Scholar]
- Sharkey TD, Singsaas EL, Vanderveer PJ, Geron C (1996) Field measurements of isoprene emission from trees in response to temperature and light. Tree Physiol 16 649–654 [DOI] [PubMed] [Google Scholar]
- Sharkey TD, Stitt M, Heineke D, Gerhardt R, Raschke K, Heldt HW (1986) Limitation of photosynthesis by carbon metabolism. II. O2-insensitive CO2 uptake results from limitation of triose phosphate. Plant Physiol 81 1123–1129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharkey TD, Wiberley AE, Donohue AR (2008) Isoprene emission from plants: why and how. Ann Bot (Lond) 101 5–18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharkey TD, Yeh S, Wiberley AE, Falbel TG, Gong D, Fernandez DE (2005) Evolution of the isoprene biosynthetic pathway in kudzu. Plant Physiol 137 700–712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharkey TD, Yeh SS (2001) Isoprene emission from plants. Annu Rev Plant Physiol Plant Mol Biol 52 407–436 [DOI] [PubMed] [Google Scholar]
- Silver GM, Fall R (1991) Enzymatic synthesis of isoprene from dimethylallyl diphosphate in aspen leaf extracts. Plant Physiol 97 1588–1591 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stitt M (1986) Limitation of photosynthesis by carbon metabolism. I. Evidence for excess electron transport capacity in leaves carrying out photosynthesis in saturating light and CO2. Plant Physiol 81 1115–1122 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Streatfield SJ, Weber A, Kinsman EA, Hausler RE, Li JM, Post-Beittenmiller D, Kaiser WM, Pyke KA, Flugge UI, Chory J (1999) The phosphoenolpyruvate/phosphate translocator is required for phenolic metabolism, palisade cell development, and plastid-dependent nuclear gene expression. Plant Cell 11 1609–1621 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tingey DT, Evans R, Gumpertz M (1981) Effects of environmental conditions on isoprene emission from live oak. Planta 152 565–570 [DOI] [PubMed] [Google Scholar]
- Trainer M, Williams EJ, Parrish DD, Buhr MP, Allwine EJ, Westberg HH, Fehsenfeld FC, Liu SC (1987) Models and observations of the impact of natural hydrocarbons on rural ozone. Nature 329 705–707 [Google Scholar]
- Vahala J, Keinänen M, Schützendübel A, Polle A, Kangasjärvi J (2003) Differential effects of elevated ozone on two hybrid aspen genotypes predisposed to chronic ozone fumigation: role of ethylene and salicylic acid. Plant Physiol 132 196–205 [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Caemmerer S (2000) Biochemical Models of Leaf Photosynthesis. CSIRO Publishing, Collingwood, Australia
- von Caemmerer S, Edmondson DL (1986) Relationship between steady-state gas exchange, in vivo ribulose bisphosphate carboxylase activity and some carbon reduction cycle intermediates in Raphanus sativum. Aust J Plant Physiol 13 669–688 [Google Scholar]
- von Caemmerer S, Farquhar GD (1981) Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 153 376–387 [DOI] [PubMed] [Google Scholar]
- Wiberley AE, Donohue AR, Meier ME, Westphal MM, Sharkey TD (2008) Regulation of isoprene emission in Populus trichocarpa leaves subjected to changing growth temperature. Plant Cell Environ 31 258–267 [DOI] [PubMed] [Google Scholar]
- Wildermuth MC, Fall R (1996) Light-dependent isoprene emission: characterization of a thylakoid-bound isoprene synthase in Salix discolor chloroplasts. Plant Physiol 112 171–182 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wildermuth MC, Fall R (1998) Biochemical characterization of stromal and thylakoid-bound isoforms of isoprene synthase in willow leaves. Plant Physiol 116 1111–1123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilkinson MJ, Monson RK, Trahan N, Lee S, Brown E, Jackson RB, Polley HW, Fay PA, Fall R (2009) Leaf isoprene emission rate as a function of atmospheric CO2 concentration. Glob Change Biol 15 1189–1200 [Google Scholar]
- Wolfertz M, Sharkey TD, Boland W, Kuehnemann F, Yeh S, Weise SE (2003) Biochemical regulation of isoprene emission. Plant Cell Environ 26 1357–1364 [Google Scholar]