

Abstract
RNA interference (RNAi) is an endogenous process that regulates expression of genes and corresponding proteins to maintain homeostasis in diverse organisms. Non-coding RNAs (ncRNAs) including both long and short ncRNAs are widely expressed and levels of some specific microRNAs are different in tumor and non-tumor tissues. RNAi has been invaluable for unraveling critical pathways involved in cancer development, growth and metastasis and has identified critical tumor-type specific gene targets for chemotherapy. In addition, the development of new derivatized small inhibitory RNAs and more efficient methods of their delivery will facilitate the future development of these ribonucleotides as cancer chemotherapeutic agents.
KEYWORDS: RNAi, siRNA, miRNA, ncRNA, cancer, oncology, VEGF
INTRODUCTION
RNA interference (RNAi) is a relatively new discovery for inhibiting gene expression and is considered as one of the most important recent discoveries in molecular oncology. RNAi is already a valuable and well-used research tool in the analysis of molecular mechanisms for many diseases including cancer and this is because it allows researchers to silence the expression of specific genes, much like antisense technology, but with higher specificity and efficacy. RNAi was first identified as a defense mechanism against the invasion of foreign genes in the nematode Caenorhabditis elegans (Fire et al, 1998) and has subsequently been discovered in diverse eukaryotes such as insects, plants, fungi and vertebrates. RNAi is a post-transcriptional process that can effect gene silencing through chromatin remodeling, blocking protein synthesis and cleaving specifically targeted mRNA. The manner by which RNAi produces gene-silencing is primarily dependent on the structure of the initiating RNA [complete (siRNA) or incomplete (miRNA)] homology to the target mRNA (Figure 1). The classical RNAi pathway is initiated by double-stranded RNAs (dsRNAs) that are homologous to the gene being silenced. These dsRNAs are processed by an enzyme called Dicer (a cellular ribonuclease III), which generates duplexes of approximately 21 nucleotides with 3′-overhangs. These are referred to as small inhibitory or interfering RNAs (siRNA). These fragments activate a protein complex called RISC (RNA-Induced Silencing Complex) and lead to degradation of the specific mRNA through the action of a protein called Slicer (Argonaute-2)(Liu et al, 2004b; Song et al, 2004) and it has been shown recently in cytoplasmic extracts from human HEK293 and Drosophila S2 cell that these processes of dsRNA unwinding, RISC assembly as well as target RNA cleavage are ATP-independent (Matranga et al, 2005; Rand et al, 2005).
Figure 1.
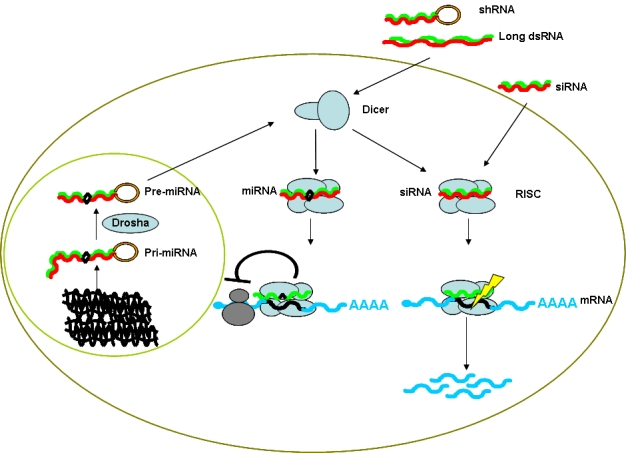
The classical RNAi and miRNA pathways: Long dsRNAs [and short hairpin RNA (shRNA)] are processed by Dicer to give siRNAs. The duplexed siRNAs are unwound as they assemble to the RNA-induced silencing complex (RISC). The single-stranded siRNA guides the endonuclease activity of the activated RISC to the homologous site on the mRNA, cleaving the mRNA. In miRNA pathway: The pre-miRNA is expressed in the nucleus from endogenous long transcripts and is processed into ∼70-nt hairpins by the RNase III family member Drosha to become the pre-miRNA. The pre-miRNA is exported to the cytoplasm by Exportin 5 and is further cleaved by Dicer into the mature miRNA. The miRNA is loaded into RISC where it guides the protein complex to a site on the mRNA that has only partial sequence complementarity to the miRNA, leading to repression of translation.
In mammalian cells, siRNA molecules are capable of 70-100 % silencing of a specific gene and has become a new and powerful alternative to other nucleic-acid-based tools such as antisense oligonucleotides and ribozymes to analyze loss-of function phenotypes. Although the specificity of gene silencing is a significant advantage of RNAi technology over other gene-targeting approaches, progress in the areas of delivery and duration of the effect are still required. Conventional methods of physical or chemical methods for transfection can be used for delivering siRNA duplexes into mammalian cells, however the resulting efficiency of the RNAi can be highly variable.
More prolonged gene silencing has been achieved by expressing siRNAs from plasmid vectors that contain specific promoters (Lee et al, 2002; Paul et al, 2002; Sui et al, 2002). The most effective plasmid construct is the one that expresses the siRNA as a short hairpin (shRNA) precursor structure of around 70 nucleotides (Paddison et al, 2002). These shRNA precursors expressed from RNA polymerase promoters II or III are processed in to a fully functional siRNA by an enzyme called Drosha (RNase III enzyme involved in processing pri-microRNA and shRNA) (Brummelkamp et al, 2002b). Viral vectors have also been developed for more stable and long-term expression of shRNAs. For example, a recombinant adenovirus system has been used to target p53 mRNA in MCF-7 breast cancer and A549 lung carcinoma cells and this was a proof of principle for a prospective siRNA applications in functional genomics and cancer gene therapy (Shen et al, 2003).
RNAi AND CANCER
RNAi is a highly promising technology for gene therapy application in treatment of different cancers and preliminary results of trials with siRNAs targeted against VEGF for treatment of macular degeneration disease are encouraging. However, in systemic disease, RNAi, like other forms of gene therapy, is challenged by the method of delivery and stability in vivo. In oncology, the initial applications of RNAi have focused on targeting dominant mutant oncogenes, amplified, translocated and viral oncogenes in order to elucidate their function and interactions with other genes and this will be discussed in the next section of this review. The use of RNAi in probing the functions of various genes in several tumors has also facilitated the systematic search for new drug targets. On the other hand, the effects of existing drugs such as imatinib and rapamycin have been enhanced by silencing their resistance-associated gene BCR-ABL using RNAi technology (Chen et al, 2004).
Role of RNAi in tumorgenesis
Cancer is a multistep genetic and epigenetic disease with a complex etiology. Cancer cells have been characterized with several defects such as mutations, down-regulation, over-expression and deletions of oncogenes and tumor suppressor genes. Different experimental approaches have been used to compare tumor cells to normal cells and have shown that defects in non-coding RNAs (ncRNAs) might be important. The first example of large ncRNA associated with tumors was the H19 gene (Hao et al, 1993) and there is now a growing list of ncRNA transcripts implicated in different types of cancer. NcRNAs that are implicated in cancer include both small and large RNAs and it has been reported that large ncRNAs can generate one or more microRNAs (miRNA). For example, the ncRNA BIC which has been implicated in growth control and oncogenesis in cancer cells generates two miRNA, one of which miR155, is over expressed in in Burkitt and B cell lymphomas (Metzler et al, 2004; Eis et al, 2005). For this reason both long and short ncRNAs implicated in cancer will be discussed in this review.
Long ncRNA
It has been reported that Prostate Specific Gene 1 (PCGEM1) ncRNA gene is over expressed in prostate tumors compared to normal and primary tumor specimens (Srikantan et al, 2000). In addition, over expression of PCGEM1 in LNCaP and in NIH3T3 cells enhanced cell proliferation and dramatically increased in colony formation suggesting a biological role for PCGEM1 in prostate tumorigenesis (Petrovics et al, 2004). Interestingly, some of these ncRNAs have been used as very sensitive and specific markers for the detection of specific tumors. For example, one study with patient urine samples suggested that DD3 ncRNA gene which is overexpressed in prostate cancer can be used as a molecular marker for the diagnosis of this disease (Tinzl et al, 2004). In addition, another study has considered this ncRNA as a target for further cancer therapies because it is not expressed in any normal human tissues, including breast, bladder, testis, gastrointestinal organ, and musculoskeletal tissue and this has been confirmed by reverse transcription-polymerase chain reaction analysis (Schalken et al, 2003). Metastasis Associated in Lung Adenocarcinoma Transcript 1 ncRNA gene or the MALAT-1 is over expressed in NSCLC (Non Small Cell Lung Carcinoma) and has been characterized as a highly reliable prognostic parameter for NSCLC patient survival, MALAT-1 has been used for identification of early-stage NSCLC patients who are at high risk to develop metastases (Ji et al, 2003). Some ncRNAs such as small dendritic non-translatable RNA BC1 are selectively expressed in tumor cells, but not in the corresponding normal tissues (Chen et al, 1997b). In a similar study, BC200 ncRNA, a brain-specific small cytoplasmic RNA, is expressed in human carcinomas of the breast, cervix, esophagus, lung, ovary, parotid, and tongue, but not in their corresponding non-tumor tissues (Chen et al, 1997a).
Short ncRNA
At present, there are no reports of endogenously expressed siRNAs in mammals. However, miRNAs have been identified from various organisms and cell types (Lagos-Quintana et al, 2003; Lim et al, 2003a; Lim et al, 2003b). Differential expression of miRNAs has been reported in several human cancers including chronic lymphocytic leukemia (Calin et al, 2002), colorectal neoplasia (Michael et al, 2003) and Burkitt's lymphoma (Metzler et al, 2004) and human miRNAs are usually located at fragile sites and genomic loci involved in several cancers (Calin et al, 2004).
Some of these miRNA, have been identified based on their homology and proximity to known genes (Pasquinelli et al, 2000); other miRNAs have been characterized by scoring candidate miRNA stem loops and pairing with known miRNA genes (Ambros et al, 2003; Lim et al, 2003b). Currently, there are 326 confirmed miRNAs in humans, and computational searches estimate that the total count might be as high as one thousand (Bentwich et al, 2005; Berezikov et al, 2005). In another recent study, Bead-based hybridization method has been used as a profiling method for the analysis of expression of all known miRNAs across a large number of samples of human normal and tumor tissues including bladder, breast, follicular lymphoma, kidney, liver, melanoma, mesothelioma, pancreas, prostate, stomach, uterus, acute myelogenous leukaemia and others (Lu et al, 2005). The miRNA profiles were highly informative, they showed a general downregulation of miRNAs in tumors compared with normal tissues and a differential expression of nearly all miRNAs across cancer types which suggest that unlike mRNA expression, a small number of miRNAs might be sufficient to classify human cancers and reflects the differentiation state of the tumors.
Application of oligonucleotide microchip methods have identified a total of 281 miRNA precursor sequences and have also quantified their expression (Liu et al, 2004a). This approach can also be utilized to compare miRNA expression in normal or malignant tissues. For example, miR-143 and miR-145 consistently display lower steady-state levels at the adenomatous and cancerous stages of colorectal neoplasia (Michael et al, 2003). Importantly, deletion and/or down-regulation of miR-15a and miR-16a has frequently been detected in patients with B-CLL (Calin et al, 2002). Recently, it was suggested that both miR-15a and miR-16 potentiate the normal apoptotic response by targeting the anti-apoptotic gene BCL-2 (Cimmino et al, 2005). In this case, these miRNAs function as tumor suppressors, and re-expression of miR-15a–16 in lymphoma cells promotes apoptosis. Expression of let-7 miRNAs were decreased in lung cancer patients and were an independent factor for prediction of patient survival (Takamizawa et al, 2004). Another study showed that over-expression of let-7 miRNAs in lung adenocarcinoma cell lines inhibits lung cancer cell growth in vitro (Takamizawa et al, 2004) indicating an important role for let-7 miRNAs in lung tumorigenesis. A recent study, showed an elevated expression of the mature miRNAs collectively denoted as mir-17–92 in primary lymphomas and a wide range of tumor derived cell lines (He et al, 2005; Tagawa and Seto, 2005).
Transcriptional regulation of miRNA expression and modulation of their target genes has been investigated. One study has shown that c-Myc transcription factor activates expression of a cluster of six miRNA on human chromosome 13 and two of these, miR-17-5p and miR-20a, negatively regulate E2F, another transcription factor. E2F1 is a down stream target of c-Myc that promotes cell cycle progression. This report identifies miRNAs as targets of c-Myc that tightly regulates c-Myc-mediated cellular proliferation by decreasing E2F1 expression (O'Donnell et al, 2005).
The recent identification of new classes of ncRNAs implicated in important steps of cancer initiation and progression reinforces the importance of these transcripts in the process of tumorigenesis. Moreover, some of the newly identified ncRNAs can now be used as targets for drug development and for early diagnosis and prognosis of cancer suggesting that some of these transcripts may be as important as protein-coding genes in cancer biology.
Application of RNAi in studying multistep carcinogenesis pathways
RNAi has become the method of choice for specific gene silencing in cell culture. The success of this approach depends on the effectiveness of siRNA delivery and design of the siRNA sequence (McManus and Sharp, 2002; Bantounas et al, 2004). Different biochemical, pharmacological, and histological assays have been used to determine the effects of siRNA inhibition of specific genes and to analyze the phenotypic changes in cells. Currently, the use of siRNA to characterize gene function, and potential therapeutic drug targets is a highly promising application for this technology (Lu et al, 2003). Selective gene knockdown can be used to identify critical genes and/or pathways that can be targeted by siRNA, drugs and their combinations for treatment of various disease including cancers. To reach these goals, RNAi has been used to investigate many of the important stages in tumor formation, growth and metastasis and some of these applications are highlighted below.
Oncogenes and tumor suppressor genes
RNAi has been employed to investigate gene function and molecular mechanisms of tumorgenesis and progression. For example, cellular oncogenes (c-onc) are normal cellular genes with the potential to enhance cell growth and tumor formation. Numerous proteins such as growth factors and their receptors, signal transducers and transcription factors associated with tumor initiation and progression are encoded by c-onc. For example the Ras gene family is frequently mutated in human cancers, particularly in pancreatic and colon carcinoma. Ras genes are guanine nucleotide binding proteins which include K-RAS, H-RAS, and N-RAS and these proteins are associated with the inner plasma membrane and transduce external signals to regulate intracellular functions. In CAPAN-1 human pancreatic cells down-regulation of K-RAS protein by RNAi leads to loss of anchorage-independent growth and tumorigenesis (Brummelkamp et al, 2002a). Single and combinatorial c-onc siRNAs combined with cationic lipid complexes have been transfected into HeLa, lung adenocarcinoma, hepatoma, ovarian carcinoma, and melanoma cells. The results showed that siRNAs-dependent downregulation of bcl-2, cdk-2, mdm-2, pkc-α, tgf-β1, H-ras, vegf, and gfp mRNA expression differentially suppressed proliferation of these cancer cell lines (Yin et al, 2003) demonstrating that many oncogenes are potential molecular targets for human cancer treatment by siRNAs.
Tumour suppressor genes (TSGs) generally inhibit cell proliferation and tumor formation and mutation or loss-of-function of these genes drives normal cells toward cancer cell phenotype. For example the somatic inactivation of Retinoblastoma (Rb) tumor suppressor gene leads to enhanced tumor growth. RNAi technology has been used to investigate a new program of dE2F/RBF- dependent transcription, in which dE2F2/RBF complexes repressed oncogene expression in SL2 cells. Each component of the dE2F/dDP/RBF pathway was silenced by RNAi and examination of changes in gene expression showed a remarkable division of labor between family members. Classic E2F targets, encoding functions required for cell cycle progression, are expressed in cycling cells and are primarily dependent on dE2F1and RBF1 (Dimova et al, 2003).
Another TSG locus that has been studied using RNAi is INK4A. This locus is often inactivated in human cancer and encodes for p14ARF and p16INK4A that inhibit cell growth through p53 and pRb, respectively. The function of the human INK4A genes has been investigated in normal and tumorigenic growth and it has been shown that the suppression of p16INK4A expression did not affect cell proliferation but promoted their growth and transformation with loss of p53 expression (Agami, 2002).
DNA damage and genome stability
Biological responses to DNA damage are mediated by multiple DNA repair pathways, or cell cycle checkpoint responses that inhibit cellular pathways leading to cell death. Cells routinely experience several kinds of DNA lesions and these include mismatches that occur during replication, various types of base damage and single and double DNA strand breaks (Jeggo et al, 1991; Hoeijmakers, 2001; Bedford and Dewey, 2002). Cells deal with DNA double strand breaks by two known processes, namely non-homologous end joining (NHEJ) which can lead to deletions and chromosomal rearrangements (Dai et al, 2003) and homology-directed repair (HDR) or repair by homologous recombination [Reviewed in (Jeggo et al, 1991; Hoeijmakers, 2001; Bedford and Dewey, 2002)].
Defects in DNA repair enzymes may not affect cell viability, but result in genomic instability, and thereby increase the rate of genetic changes and the rate of tumor formation (Vogelstein and Kinzler, 2004). SiRNA technology has been applied to the DNA repair field through knockdown of the ATR interacting protein (ATRIP)(Cortez et al, 2001). ATR (Ataxia-Telangiectasia and Rad3-Related) is a member of the phosphatidylinositol kinase family which is involved in DNA damage signaling and cell cycle checkpoints. While the related cell cycle checkpoint protein ATM (Ataxia-Telangiectasia Mutated) from AT patients (characterized by cerebellar ataxia, x-ray hypersensitivity and a predisposition to malignancy) has been investigated, studies on ATR were not well characterized due to lack of ATR−/− cell lines since ATR knock-out mice exhibit embryonic lethality (Brown and Baltimore, 2000). Three different siRNAs targeting different regions of the ATRIP mRNA were used for gene silencing and the results reveal that ATRIP is phosphorylated by ATR, regulates ATR expression, and is an essential component of the DNA damage checkpoint pathway (Cortez et al, 2001). SiRNAs against ATR have also been used in human lymphoblast cell lines and show participation of ATR in chromosome fragile site stability (Casper et al, 2002). SiRNA has also been used to knock down the p53-binding protein (53BP1) and showed that this protein plays a role in checkpoint responses and phosphorylation of 53BP1 after DNA damage by ionizing radiation leading to rapid relocalization, along with Mre11, and gammaH2AX, to the sites of DNA damage (Morales et al, 2003).
Rad-51 gene is another gene of interest in DNA repair field that has been studied using the RNAi technology. Suppression of rad-51 in C. elegans showed that rad51 played a roles in sister chromatids repair after both endogenous and exogenous double-strand breaks (DSB) formation during meiosis, but not as ‘pairing centers’ for meiotic synapsis (Takanami et al, 2003). Rad-51 RNAi did not affect chromosome appearance if DSB was prevented by spo-11 or mre-11 mutations. Chromosomal DNA degradation is a hallmark of apoptosis and represents a key point in cell death execution. Recently, seven additional cell death-related nucleases were identified using an RNAi-based functional genomic approach (Parrish and Xue, 2003). For example CPS-6 and NUC-1 nucleases comprise two independent pathways that contribute to cell death induced by degrading chromosomal DNA. The application of RNAi in this area of research has elucidated the mechanism of genome instability and also it has suggested some molecular targets for clinical applications.
Angiogenesis and metastasis
Tumor angiogenesis involves new blood vessels formation and is mediated by changes in the balance between angiogenesis factors and angiogenesis inhibitors (Bergers and Benjamin, 2003). Angiogenesis can be suppressed by histone deacetylase (HADC) inhibitors. RECK is a membrane-anchored glycoprotein that negatively regulates matrix metalloproteinases (MMPs) and inhibits tumor metastasis and angiogenesis. Inhibition of RECK by siRNAs in CL-1 human lung cancer cells abolished the inhibitory effect of HDAC inhibitor trichostatin A (TSA) on MMP-2 activation (Liu et al, 2003). This study provided insights on how HDACs suppress tumor invasion and angiogenesis and provided a new strategy for cancer therapy. The CXC chemokine receptor-4 (CXCR4) is over-expressed in the highly invasive MDA-MB-231 human breast cancer cells and may play a role in the metastasis of this cell line in animal models. Using an inducible siRNAs expression system, it was shown that knockdown of CXCR4 in MDA-MB-231cells resulted in significant inhibition of breast cancer cell migration in vitro (Chen et al, 2003).
Expression of the angiogenic factor VEGF in cancer cell lines is regulated through Sp protein interactions with several proximal GC-rich motifs in the VEGF promoter (Shi et al, 2001; Abdelrahim et al, 2004). Sp proteins and Krüppel-like factors (KLFs) are a growing family of transcription factors that bind GC-rich motifs and modulate gene transcription (Safe and Abdelrahim, 2005). There is emerging evidence that Sp proteins are critical factors in tumor development, growth and metastasis; however, most studies have focused primarily on Sp1 protein/mRNA expression and not other member of this family. It was initially reported that Sp1 was overexpressed in pancreatic tumors compared to normal tissues, and there was a correlation between Sp and VEGF expression in pancreatic tumors and cancer cell lines (Shi et al, 2001). A recent study in this laboratory used siRNAs to further investigate the role of Sp1 and other Sp proteins in regulation of VEGF expression in pancreatic cancer cells (Abdelrahim et al, 2004). The results show that the proximal GC-rich sites in the VEGF promoter are required for expression of VEGF in Panc1 cells; however, results of RNA interference showed that multiple Sp proteins are involved in VEGF regulation (Figure 2). Sequential knockdown of Sp1, Sp3 or Sp4 by RNAi showed that all three proteins regulated transactivation in pancreatic cancer cells transfected with the pVEGF1, pVEGF2 and pVEGF3 constructs containing VEGF promoter inserts (Figure 2). Sp4 knockdown decreased activity (≥ 50%) in cells transfected with all three constructs, whereas Sp1 and Sp3 differentially regulated gene expression in cells transfected with pVEGF1 and pVEGF3, respectively. These results indicate that Sp1, Sp3 and Sp4 cooperatively regulate VEGF expression and the promoter analysis suggested differential activation by these proteins in the proximal and distal regions of the promoter. Sp4 expression has not been extensively investigated in cancer cells, and results from these RNAi studies suggest that the metastatic and growth potential of pancreatic tumors may also be dependent on levels of Sp4.
Figure 2.
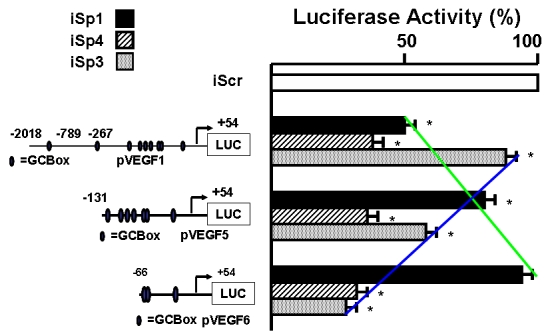
Regulation of VEGF expression in Panc-1 pancreatic cancer cells. Cells were transfected with various VEGF promoter constructs, transfected with scramble siRNA (non-specific) or with small inhibitory RNAs for Sp1 (iSp1), Sp3 (iSp3) or Sp4 (iSp4), and luciferase activity determined as previously reported (Abdelrahim et al, 2004).
Another important factor that regulates expression of VEGF and other angiogenic proteins is hypoxia-inducible factor (HIF). HIF is α/β heterodimeric DNA binding complex that directs an extensive transcriptional response involving the induction of genes with important roles in several aspects of tumor angiogenesis, invasion and metastasis. HIF-1α is the O2-regulated subunit while HIF-1 β is constitutively expressed. Overexpression of the HIF-1α subunit, due to intratumoral hypoxia or genetic alterations, has been demonstrated in common human cancers. One study has demonstrate that HIF-1α overexpression stimulates Matrigel invasion by HCT116 human colon carcinoma cells whereas this process is inhibited by a small interfering RNAs targeted against HIF-1α. The same siRNA approach was used to show that HIF-1 regulates the expression of genes encoding cathepsin D; matrix metalloproteinase 2; urokinase plasminogen activator receptor (uPAR); fibronectin 1; keratins 14, 18, and 19; vimentin; transforming growth factor α; and autocrine motility factor, which are proteins that play important roles in the pathophysiology of tumor invasion (Krishnamachary et al, 2003).
To date, three HIF-1 isoforms have been described, with the best characterized being HIF-1α and HIF-2α. The individual roles of these two alternative subunits are not fully understood due to close similarities between them. SiRNA approach has been used successfully to determine functional differences between HIF-1α and HIF-2α in different human cell lines (Warnecke et al, 2004). In this report, RNase protection assays revealed that HIF-1α knockdown in HeLa cells reduced hypoxia-stimulated mRNA induction of glucose transporter 1 (GLUT-1), lactate dehydrogenase A (LDH-A), VEGF, carbonic anhydrase IX (CA IX), and HIF prolyl hydroxylase 2 (PHD2) by 40–60% while HIF-2α knockdown had no effect on these HIF target genes. On the other hand, the study showed that mRNA induction of erythropoietin (EPO) after exposure of Hep3B cells to hypoxia was almost abolished by the HIF-2α siRNA whereas HIF-1α knockdown had no effect.
RNAi has also been used to study the viability of endothelial cells when Tie-2-dependent signaling is interrupted by RNAi. Tie-2 is a small molecule inhibitor of endothelial cell specific tyrosine kinases. Decreased expression of Tie-2 in endothelial cells by RNAi resulted in loss of cell viability and phosphotidylinisitol-3kinase (PI3K) activity and increased expression of thrombospondin (endogenous anti-angiogenic proteins) (Niu et al, 2004). These data show that Tie-2 mediated activation PI3Kinase/Akt is involved in angiopoietin signaling, which in turn represses thrombospondin expression suggesting that blocking of Tie-2 in endothelial cells might be an effective inhibitor of angiogenesis.
Other important proteins involved in tumor invasion and metastasis are matrix metalloproteases (MMP)-9 and cathepsin B which work by influencing matrix degradation. Cell-cell interactions between human microvascular endothelial cells werer significantly reduced when MMP-9 and cathepsin B genes expression were suppressed by hairpin RNAs in SNB19 cells (Lakka et al, 2004). It has been demonstrated that direct intratumoral injections of plasmid DNA expressing hairpin RNAs for MMP-9 and cathepsin B significantly inhibited established glioma tumor growth and invasion in vivo and intraperitoneal injections of these constructs completely repressed pre-established tumors for up to 4 months (Liu et al, 2003). This was the first report to demonstrate that simultaneous suppression of MMP-9 and cathepsin B genes by RNAi can be used for the treatment of human gliomas. These results show that RNAi can be used to selectively target genes that are involved in tumor growth, invasion and metastasis and this technique holds great promise for the clinical treatment of a wide variety of human tumors.
Tumor cell survival and apoptosis
The rate of tumor growth and expansion is controlled by the balance between cell proliferation/survival and apoptosis and in normal cells these processes are strictly regulated. However, apoptosis is disrupted in cancer cells and tumors (Wilda et al, 2002; Schmitt, 2003) and cell cycle proteins required for cell survival and proliferation are up-regulated or continuously activated.
Sp1 also plays a role in regulating hormone-responsiveness of several genes in breast cancer cells associated with cell proliferation and cycle progression (cyclin D1, E2F1, c-fos, transforming growth factor α), purine/pyrimidine synthesis and metabolism [thymidylate synthase, adenosine deaminase, DNA polymerase α, carbamylphosphate synthetase/aspartate carbamyltransferase/dihydroorotase (CAD)], angiogenesis (VEGF), and anti-apoptosis (bcl-2) [Reviewed in (Safe and Abdelrahim, 2005)]. These genes are regulated by ERα/Sp1-3 and knockdown of Sp1 and other Sp proteins resulted in loss of hormone induced gene expression. RNA interference using small inhibitory RNAs for Sp1, Sp3 and Sp4 has also used to investigate the role of these proteins in growth of Panc-1 pancreatic cancer cells (Abdelrahim et al, 2004). Results summarized in Figure 3 show that transfection of Panc-1 cells with nonspecific small inhibitory RNA (iScr) did not affect distribution of Panc-1 cells in G0/G1, G2/M or S phases of the cell cycle, whereas small inhibitory RNA for Sp1 (iSp1) decreased the percentage of cells in G2/M and S phase and increased the percentage in G0/G1. This response was also accompanied by decreased Rb phosphorylation and reflects the overall decrease in multiple genes associated with Sp1-dependent growth. In addition, dramatic response was observed in cells transfected with small inhibitory RNA for Sp3 (iSp3) in which the percentage of cells in G0/G1 and S phase increased by ∼20% and decreased by ∼17%, respectively. These results were explained after subsequent analysis of several cell cycle genes after iSp3 treatment. In Panc-1 and other pancreatic cancer cell lines transfected with iSp3 the cyclin-dependent kinase inhibitor p27 was upregulated showing that Sp3 enhanced cell growth through inhibiting p27 expression. On the other hand, transfection with small inhibitory RNA for Sp4 (iSp4) showed a minimum effect on cell cycle progression and p27 protein expression.
Figure 3.
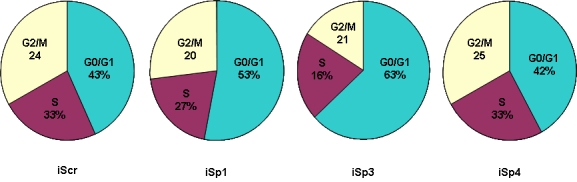
Effects of Sp proteins on cycle phase distribution in Panc-1 cells. Cells were transfected with Scramble siRNA (iScr, non-specific) or with small inhibitory RNA for iSp1, iSp3 or iSp4, and the percentage distribution of Panc-1 cells in G0/G1, G2/M and S phases of the cell cycle were determined by FACS analysis (Abdelrahim et al, 2004). Sp3 knockdown increased G0/G1 and decreased S phase due to increased expression of p27.
Apoptosis has also been extensively studied by RNAi. Antiapoptotic factors have been over expressed in many tumors and that might explain the resistance of some tumors to apoptosis induced by cytotoxic drugs (Schmitt, 2003). For example, the livin (ML-IAP, KIAP) gene is an antiapoptotic factor and silencing of livin in MeWo melanoma and HeLa cervical carcinoma cells strongly increased apoptotic rates in response to different proapoptotic stimuli and this was linked to caspase-3 activation (Crnkovic-Mertens et al, 2003; Schmitt, 2003). This study showed that functional inhibition of specific antiapoptotic factors by siRNA may provide a rational basis for the development of novel therapeutic strategies and the livin gene was identified as a promising molecular target for therapeutic inhibition.
Bcl-2 and xIAP are antiapoptotic factors and the sensitivity of MCF-7 breast cancer cells to treatment with the drugs etoposide and doxorubicin is increased after silencing of bcl-2 or xIAP by siRNA (Lima et al, 2004). Treatment of these cells with both siRNAs decreases the number of viable cells and increased cellular apoptosis. These studies suggest that combination therapy that includes traditional chemotherapies coadministered with specific siRNAs may be a highly effective therapy for cancer.
RNAi has also been used to determine which apoptotic genes regulate the survival of HCT116 colorectal carcinoma cells. Bcl-2 and Bcl-xL are silenced in isogenic clones of p53+/+, p53−/−, Bax+/−, Bax−/− cells. In this study authors identified a novel proapoptotic function of p53 that does not require activation by genotoxic agents and it appeared to be constitutively suppressed by Bcl-2. In addition Bcl-2 induced massive p53-dependent apoptosis which required Bax and caspase 2 as essential apoptotic mediators. RNAi showed that this Bcl-2/p53 functional interface represents a key regulator of apoptosis which can be activated by targeting Bcl-2 in colorectal carcinoma cells (Jiang and Milner, 2003).
It is clear that RNAi has greatly facilitated the identification and the study of components of apoptosis and survival pathways and this will facilitate identification of specific gene targets for the improving the effectiveness of cancer therapies.
Therapeutic potential of siRNA in cancer
The ability of RNAi to silence disease-associated genes in cell culture and animal models has spurred development of RNAi-based reagents for clinical applications to treat diseases including cancer (Howard, 2003; Wall and Shi, 2003). SiRNAs are readily synthesized with low production costs compared to protein or antibody therapies. In addition, siRNAs have favorable pharmacokinetic properties and can be delivered to a wide range of organs (Braasch et al, 2004). However, their stability in blood and delivery methods are challenges that must be solved for developing effective RNAi reagents for cancer therapy. Several groups have been investigating the use of alternative backbone and nucleotide modifications to improve the clinical properties of these reagents. For example, by conjugating the 3′-end of the sense strand of siRNA with cholesterol through a pyrrolidine linker has markedly improved the pharmacological properties of siRNA molecules (Soutschek et al, 2004). The Cholesterol-conjugated siRNA are more resistant to nuclease degradation, exhibit increased stability in blood by increasing binding to human serum albumin and show increased uptake into liver. Another study has shown that boranophosphate modified siRNAs were 10 times more nuclease resistant than unmodified siRNAs (Hall et al, 2004). In addition, boranophosphate siRNAs were more potent than unmodified siRNAs and appeared to act through the standard RNAi pathway. As an alternative to backbone modifications other groups have improved siRNA stability and delivery in vivo by complexing them with polyethyleneimine (PEI) (Urban-Klein et al, 2005) or atelocollagen (Minakuchi et al, 2004). For example, in a subcutaneous mouse tumor model, intraperitoneal administration of siRNAs complexed with PEI led to the delivery of intact siRNAs into tumors (Urban-Klein et al, 2005). Another group has constructed self-assembling nanoparticles with siRNA and PEI PEGylated with an Arg-Gly-Asp peptide ligand attached to the distal end of the polyethylene glycol (PEG) as a means to target tumor neovasculature-expressing integrins (Schiffelers et al, 2004). Treatment of tumor-bearing mice with this complex by intravenous administration resulted in selective tumor uptake, siRNA sequence-specific inhibition of protein expression within the tumor, and inhibition of both tumor angiogenesis and growth. The results showed that siRNAs can be targeted at two levels: tumor tissue-selective delivery via the nanoparticle ligand and gene pathway selectivity via the siRNA oligonucleotide.
Topical gels have also been used to deliver siRNAs to cells and could open the way for dermatological applications, as well as the treatment for cervical cancer (Jiang et al., 2004). Intradermal administration of nucleic acids via gene guns have also been used to deliver siRNA in vivo to enhance cancer vaccine potency (Kim et al., 2005). Coadministration of DNA vaccines encoding human papillomavirus type 16 E7 with siRNAs targeting key proapoptotic proteins Bak and Bax prolonged the lives of antigen-expressing dendritic cells in the draining lymph nodes, enhanced antigen-specific CD8(+) T-cell responses, and elicited potent antitumor effects against an E7-expressing tumor model in vaccinated mice.
Systemic administration of siRNA has also been used to target tumor growth and angiogenesis. Efficient delivery of siRNA targeting VEGF into tumors by systemic intrperitoneal administration at low doses (125 μg/kg/day) in saline was observed (Filleur et al., 2003). This treatment resulted in a robust inhibition of the endogenous VEGF expression and produced a marked inhibition of fibrosarcoma tumor cells growth. No side-effects were detected in the animals or in specific tissue/organs suggesting that this route of siRNA administration is highly effective.
CONCLUSIONS
RNA interference and the function of non-coding RNAs represent one of the major scientific discoveries of the past decade. The endogenous functions of these ribunucleotides remain to be further discovered and the application of specific gene targeting by small inhibitory RNAs will play an important future role as another important treatment option for cancer and other diseases.
ACKNOWLEDGEMENTS
The authors wish to acknowledge M.D. Anderson Pancreatic Cancer SPORE (P20-CA10193) and the National Institute of Health (ES09106, CA104116 and CA108718) for the supports.
LIST OF ABBREVIATIONS
- shRNA
Short hairpin RNA
- miRNA
MicroRNA
- ncRNA
Non-coding RNA
- RISC
RNA-induced silencing complex
- VEGF
Vascular endothelial growth factor
- TSGs
Tumor suppressor genes
- iSp
Small inhibitory RNA for Sp protein
- iScr
Scrambled small inhibitory RNA
STATEMENT OF COMPETING INTERESTS
The authors declared no competing interests.
REFERENCES
- Abdelrahim M, Smith R, 3rd, Burghardt R, Safe S. Role of Sp proteins in regulation of vascular endothelial growth factor expression and proliferation of pancreatic cancer cells. Cancer Res. 2004;64:6740–6749. doi: 10.1158/0008-5472.CAN-04-0713. [DOI] [PubMed] [Google Scholar]
- Agami R. RNAi and related mechanisms and their potential use for therapy. Curr Opin Chem Biol. 2002;6:829–834. doi: 10.1016/s1367-5931(02)00378-2. [DOI] [PubMed] [Google Scholar]
- Ambros V, Lee RC, Lavanway A, Williams PT, Jewell D. MicroRNAs and other tiny endogenous RNAs in C. elegans. Curr Biol. 2003;13:807–818. doi: 10.1016/s0960-9822(03)00287-2. [DOI] [PubMed] [Google Scholar]
- Bantounas I, Phylactou LA, Uney JB. RNA interference and the use of small interfering RNA to study gene function in mammalian systems. J Mol Endocrinol. 2004;33:545–557. doi: 10.1677/jme.1.01582. [DOI] [PubMed] [Google Scholar]
- Bedford JS, Dewey WC. Radiation Research Society. 1952-2002. Historical and current highlights in radiation biology: has anything important been learned by irradiating cells? Radiat Res. 2002;158:251–291. doi: 10.1667/0033-7587(2002)158[0251:hachir]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- Bentwich I, Avniel A, Karov Y, Aharonov R, Gilad S, Barad O, Barzilai A, Einat P, Einav U, Meiri E, Sharon E, Spector Y, Bentwich Z. Identification of hundreds of conserved and nonconserved human microRNAs. Nat Genet. 2005;37:766–770. doi: 10.1038/ng1590. [DOI] [PubMed] [Google Scholar]
- Berezikov E, Guryev V, van de Belt J, Wienholds E, Plasterk RH, Cuppen E. Phylogenetic shadowing and computational identification of human microRNA genes. Cell. 2005;120:21–24. doi: 10.1016/j.cell.2004.12.031. [DOI] [PubMed] [Google Scholar]
- Bergers G, Benjamin LE. Tumorigenesis and the angiogenic switch. Nat Rev Cancer. 2003;3:401–410. doi: 10.1038/nrc1093. [DOI] [PubMed] [Google Scholar]
- Braasch DA, Paroo Z, Constantinescu A, Ren G, Oz OK, Mason RP, Corey DR. Biodistribution of phosphodiester and phosphorothioate siRNA. Bioorg Med Chem Lett. 2004;14:1139–1143. doi: 10.1016/j.bmcl.2003.12.074. [DOI] [PubMed] [Google Scholar]
- Brown EJ, Baltimore D. ATR disruption leads to chromosomal fragmentation and early embryonic lethality. Genes Dev. 2000;14:397–402. [PMC free article] [PubMed] [Google Scholar]
- Brummelkamp TR, Bernards R, Agami R. Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell. 2002a;2:243–247. doi: 10.1016/s1535-6108(02)00122-8. [DOI] [PubMed] [Google Scholar]
- Brummelkamp TR, Bernards R, Agami R. A system for stable expression of short interfering RNAs in mammalian cells. Science. 2002b;296:550–553. doi: 10.1126/science.1068999. [DOI] [PubMed] [Google Scholar]
- Calin GA, Dumitru CD, Shimizu M, Bichi R, Zupo S, Noch E, Aldler H, Rattan S, Keating M, Rai K, Rassenti L, Kipps T, Negrini M, Bullrich F, Croce CM. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci USA. 2002;99:15524–15529. doi: 10.1073/pnas.242606799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calin GA, Sevignani C, Dumitru CD, Hyslop T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M, Croce CM. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc Natl Acad Sci USA. 2004;101:2999–3004. doi: 10.1073/pnas.0307323101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casper AM, Nghiem P, Arlt MF, Glover TW. ATR regulates fragile site stability. Cell. 2002;111:779–789. doi: 10.1016/s0092-8674(02)01113-3. [DOI] [PubMed] [Google Scholar]
- Chen J, Wall NR, Kocher K, Duclos N, Fabbro D, Neuberg D, Griffin JD, Shi Y, Gilliland DG. Stable expression of small interfering RNA sensitizes TEL-PDGFbetaR to inhibition with imatinib or rapamycin. J Clin Invest. 2004;113:1784–1791. doi: 10.1172/JCI20673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen W, Bocker W, Brosius J, Tiedge H. Expression of neural BC200 RNA in human tumours. J Pathol. 1997a;183:345–351. doi: 10.1002/(SICI)1096-9896(199711)183:3<345::AID-PATH930>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- Chen W, Heierhorst J, Brosius J, Tiedge H. Expression of neural BC1 RNA: induction in murine tumours. Eur J Cancer. 1997b;33:288–292. doi: 10.1016/s0959-8049(96)00453-4. [DOI] [PubMed] [Google Scholar]
- Chen Y, Stamatoyannopoulos G, Song CZ. Down-regulation of CXCR4 by inducible small interfering RNA inhibits breast cancer cell invasion in vitro. Cancer Res. 2003;63:4801–4804. [PubMed] [Google Scholar]
- Cimmino A, Calin GA, Fabbri M, et al. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc Natl Acad Sci USA. 2005;102:13944–13949. doi: 10.1073/pnas.0506654102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cortez D, Guntuku S, Qin J, Elledge SJ. ATR and ATRIP: partners in checkpoint signaling. Science. 2001;294:1713–1716. doi: 10.1126/science.1065521. [DOI] [PubMed] [Google Scholar]
- Crnkovic-Mertens I, Hoppe-Seyler F, Butz K. Induction of apoptosis in tumor cells by siRNA-mediated silencing of the livin/ML-IAP/KIAP gene. Oncogene. 2003;22:8330–8336. doi: 10.1038/sj.onc.1206973. [DOI] [PubMed] [Google Scholar]
- Dai Y, Kysela B, Hanakahi LA, et al. Nonhomologous end joining and V(D)J recombination require an additional factor. Proc Natl Acad Sci USA. 2003;100:2462–2467. doi: 10.1073/pnas.0437964100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dimova DK, Stevaux O, Frolov MV, Dyson NJ. Cell cycle-dependent and cell cycle-independent control of transcription by the Drosophila E2F/RB pathway. Genes Dev. 2003;17:2308–2320. doi: 10.1101/gad.1116703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eis PS, Tam W, Sun L, et al. Accumulation of miR-155 and BIC RNA in human B cell lymphomas. Proc Natl Acad Sci USA. 2005;102:3627–3632. doi: 10.1073/pnas.0500613102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filleur S, Courtin A, Ait-Si-Ali S, et al. SiRNA-mediated inhibition of vascular endothelial growth factor severely limits tumor resistance to antiangiogenic thrombospondin-1 and slows tumor vascularization and growth. Cancer Res. 2003;63:3919–3922. [PubMed] [Google Scholar]
- Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 1998;391:806–811. doi: 10.1038/35888. [DOI] [PubMed] [Google Scholar]
- Hall AH, Wan J, Shaughnessy EE, Ramsay Shaw B, Alexander KA. RNA interference using boranophosphate siRNAs: structure-activity relationships. Nucleic Acids Res. 2004;32:5991–6000. doi: 10.1093/nar/gkh936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hao Y, Crenshaw T, Moulton T, Newcomb E, Tycko B. Tumour-suppressor activity of H19 RNA. Nature. 1993;365:764–767. doi: 10.1038/365764a0. [DOI] [PubMed] [Google Scholar]
- He L, Thomson JM, Hemann MT, et al. A microRNA polycistron as a potential human oncogene. Nature. 2005;435:828–833. doi: 10.1038/nature03552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoeijmakers JH. Genome maintenance mechanisms for preventing cancer. Nature. 2001;411:366–374. doi: 10.1038/35077232. [DOI] [PubMed] [Google Scholar]
- Howard K. Unlocking the money-making potential of RNAi. Nat Biotechnol. 2003;21:1441–1446. doi: 10.1038/nbt1203-1441. [DOI] [PubMed] [Google Scholar]
- Jeggo PA, Tesmer J, Chen DJ. Genetic analysis of ionising radiation sensitive mutants of cultured mammalian cell lines. Mutat Res. 1991;254:125–133. doi: 10.1016/0921-8777(91)90003-8. [DOI] [PubMed] [Google Scholar]
- Ji P, Diederichs S, Wang W, et al. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene. 2003;22:8031–8041. doi: 10.1038/sj.onc.1206928. [DOI] [PubMed] [Google Scholar]
- Jiang M, Milner J. Bcl-2 constitutively suppresses p53-dependent apoptosis in colorectal cancer cells. Genes Dev. 2003;17:832–837. doi: 10.1101/gad.252603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang M, Rubbi CP, Milner J. Gel-based application of siRNA to human epithelial cancer cells induces RNAi-dependent apoptosis. Oligonucleotides. 2004;14:239–248. doi: 10.1089/oli.2004.14.239. [DOI] [PubMed] [Google Scholar]
- Kim TW, Lee JH, He L, et al. Modification of professional antigen-presenting cells with small interfering RNA in vivo to enhance cancer vaccine potency. Cancer Res. 2005;65:309–316. [PubMed] [Google Scholar]
- Krishnamachary B, Berg-Dixon S, Kelly B, et al. Regulation of colon carcinoma cell invasion by hypoxia-inducible factor 1. Cancer Res. 2003;63:1138–1143. [PubMed] [Google Scholar]
- Lagos-Quintana M, Rauhut R, Meyer J, Borkhardt A, Tuschl T. New microRNAs from mouse and human. RNA. 2003;9:175–179. doi: 10.1261/rna.2146903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lakka SS, Gondi CS, et al. Inhibition of cathepsin B and MMP-9 gene expression in glioblastoma cell line via RNA interference reduces tumor cell invasion, tumor growth and angiogenesis. Oncogene. 2004;23:4681–4689. doi: 10.1038/sj.onc.1207616. [DOI] [PubMed] [Google Scholar]
- Lee NS, Dohjima T, Bauer G, et al. Expression of small interfering RNAs targeted against HIV-1 rev transcripts in human cells. Nat Biotechnol. 2002;20:500–505. doi: 10.1038/nbt0502-500. [DOI] [PubMed] [Google Scholar]
- Lim LP, Glasner ME, Yekta S, Burge CB, Bartel DP. Vertebrate microRNA genes. Science. 2003a;299:1540. doi: 10.1126/science.1080372. [DOI] [PubMed] [Google Scholar]
- Lim LP, Lau NC, Weinstein EG, et al. The microRNAs of Caenorhabditis elegans. Genes Dev. 2003b;17:991–1008. doi: 10.1101/gad.1074403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lima RT, Martins LM, Guimaraes JE, Sambade C, Vasconcelos MH. Specific downregulation of bcl-2 and xIAP by RNAi enhances the effects of chemotherapeutic agents in MCF-7 human breast cancer cells. Cancer Gene Ther. 2004;11:309–316. doi: 10.1038/sj.cgt.7700706. [DOI] [PubMed] [Google Scholar]
- Liu CG, Calin GA, Meloon B, et al. An oligonucleotide microchip for genome-wide microRNA profiling in human and mouse tissues. Proc Natl Acad Sci USA. 2004a;101:9740–9744. doi: 10.1073/pnas.0403293101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu J, Carmell MA, Rivas FV, et al. Argonaute2 is the catalytic engine of mammalian RNAi. Science. 2004b;305:1437–1441. doi: 10.1126/science.1102513. [DOI] [PubMed] [Google Scholar]
- Liu LT, Chang HC, Chiang LC, Hung WC. Histone deacetylase inhibitor up-regulates RECK to inhibit MMP-2 activation and cancer cell invasion. Cancer Res. 2003;63:3069–3072. [PubMed] [Google Scholar]
- Lu J, Getz G, Miska EA, et al. MicroRNA expression profiles classify human cancers. Nature. 2005;435:834–838. doi: 10.1038/nature03702. [DOI] [PubMed] [Google Scholar]
- Lu PY, Xie FY, Woodle MC. siRNA-mediated antitumorigenesis for drug target validation and therapeutics. Curr Opin Mol Ther. 2003;5:225–234. [PubMed] [Google Scholar]
- Matranga C, Tomari Y, Shin C, Bartel DP, Zamore PD. Passenger-strand cleavage facilitates assembly of siRNA into Ago2-containing RNAi enzyme complexes. Cell. 2005;123:607–620. doi: 10.1016/j.cell.2005.08.044. [DOI] [PubMed] [Google Scholar]
- McManus MT, Sharp PA. Gene silencing in mammals by small interfering RNAs. Nat Rev Genet. 2002;3:737–747. doi: 10.1038/nrg908. [DOI] [PubMed] [Google Scholar]
- Metzler M, Wilda M, Busch K, Viehmann S, Borkhardt A. High expression of precursor microRNA-155/BIC RNA in children with Burkitt lymphoma. Genes Chromosomes Cancer. 2004;39:167–169. doi: 10.1002/gcc.10316. [DOI] [PubMed] [Google Scholar]
- Michael MZ, SM OC, van Holst Pellekaan NG, Young GP, James RJ. Reduced accumulation of specific microRNAs in colorectal neoplasia. Mol Cancer Res. 2003;1:882–891. [PubMed] [Google Scholar]
- Minakuchi Y, Takeshita F, Kosaka N, et al. Atelocollagen-mediated synthetic small interfering RNA delivery for effective gene silencing in vitro and in vivo. Nucleic Acids Res. 2004;32:e109. doi: 10.1093/nar/gnh093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morales JC, Xia Z, Lu T, et al. Role for the BRCA1 C-terminal repeats (BRCT) protein 53BP1 in maintaining genomic stability. J Biol Chem. 2003;278:14971–14977. doi: 10.1074/jbc.M212484200. [DOI] [PubMed] [Google Scholar]
- Niu Q, Perruzzi C, Voskas D, Lawler J, Dumont DJ, Benjamin LE. Inhibition of Tie-2 signaling induces endothelial cell apoptosis, decreases Akt signaling, and induces endothelial cell expression of the endogenous anti-angiogenic molecule, thrombospondin-1. Cancer Biol Ther. 2004;3:402–405. doi: 10.4161/cbt.3.4.735. [DOI] [PubMed] [Google Scholar]
- O'Donnell KA, Wentzel EA, Zeller KI, Dang CV, Mendell JT. c-Myc-regulated microRNAs modulate E2F1 expression. Nature. 2005;435:839–843. doi: 10.1038/nature03677. [DOI] [PubMed] [Google Scholar]
- Paddison PJ, Caudy AA, Bernstein E, Hannon GJ, Conklin DS. Short hairpin RNAs (shRNAs) induce sequence-specific silencing in mammalian cells. Genes Dev. 2002;16:948–958. doi: 10.1101/gad.981002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parrish JZ, Xue D. Functional genomic analysis of apoptotic DNA degradation in C. elegans. Mol Cell. 2003;11:987–996. doi: 10.1016/s1097-2765(03)00095-9. [DOI] [PubMed] [Google Scholar]
- Pasquinelli AE, Reinhart BJ, Slack F, et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature. 2000;408:86–89. doi: 10.1038/35040556. [DOI] [PubMed] [Google Scholar]
- Paul CP, Good PD, Winer I, Engelke DR. Effective expression of small interfering RNA in human cells. Nat Biotechnol. 2002;20:505–508. doi: 10.1038/nbt0502-505. [DOI] [PubMed] [Google Scholar]
- Petrovics G, Zhang W, Makarem M, et al. Elevated expression of PCGEM1, a prostate-specific gene with cell growth-promoting function, is associated with high-risk prostate cancer patients. Oncogene. 2004;23:605–611. doi: 10.1038/sj.onc.1207069. [DOI] [PubMed] [Google Scholar]
- Rand TA, Petersen S, Du F, Wang X. Argonaute2 cleaves the anti-guide strand of siRNA during RISC activation. Cell. 2005;123:621–629. doi: 10.1016/j.cell.2005.10.020. [DOI] [PubMed] [Google Scholar]
- Safe S, Abdelrahim M. Sp transcription factor family and its role in cancer. Eur J Cancer. 2005;41:2438–2448. doi: 10.1016/j.ejca.2005.08.006. [DOI] [PubMed] [Google Scholar]
- Schalken JA, Hessels D, Verhaegh G. New targets for therapy in prostate cancer: differential display code 3 (DD3(PCA3)), a highly prostate cancer-specific gene. Urology. 2003;62:34–43. doi: 10.1016/s0090-4295(03)00759-3. [DOI] [PubMed] [Google Scholar]
- Schiffelers RM, Ansari A, Xu J, et al. Cancer siRNA therapy by tumor selective delivery with ligand-targeted sterically stabilized nanoparticle. Nucleic Acids Res. 2004;32:e149. doi: 10.1093/nar/gnh140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmitt CA. Senescence, apoptosis and therapy--cutting the lifelines of cancer. Nat Rev Cancer. 2003;3:286–295. doi: 10.1038/nrc1044. [DOI] [PubMed] [Google Scholar]
- Shen C, Buck AK, Liu X, Winkler M, Reske SN. Gene silencing by adenovirus-delivered siRNA. FEBS Lett. 2003;539:111–114. doi: 10.1016/s0014-5793(03)00209-6. [DOI] [PubMed] [Google Scholar]
- Shi Q, Le X, Abbruzzese JL, et al. Constitutive Sp1 activity is essential for differential constitutive expression of vascular endothelial growth factor in human pancreatic adenocarcinoma. Cancer Res. 2001;61:4143–4154. [PubMed] [Google Scholar]
- Song JJ, Smith SK, Hannon GJ, Joshua-Tor L. Crystal structure of Argonaute and its implications for RISC slicer activity. Science. 2004;305:1434–1437. doi: 10.1126/science.1102514. [DOI] [PubMed] [Google Scholar]
- Soutschek J, Akinc A, Bramlage B, et al. Therapeutic silencing of an endogenous gene by systemic administration of modified siRNAs. Nature. 2004;432:173–178. doi: 10.1038/nature03121. [DOI] [PubMed] [Google Scholar]
- Srikantan V, Zou Z, Petrovics G, et al. PCGEM1, a prostate-specific gene, is overexpressed in prostate cancer. Proc Natl Acad Sci USA. 2000;97:12216–12221. doi: 10.1073/pnas.97.22.12216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sui G, Soohoo C, Affar el B, et al. A DNA vector-based RNAi technology to suppress gene expression in mammalian cells. Proc Natl Acad Sci USA. 2002;99:5515–5520. doi: 10.1073/pnas.082117599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tagawa H, Seto M. A microRNA cluster as a target of genomic amplification in malignant lymphoma. Leukemia. 2005;19:2013–2016. doi: 10.1038/sj.leu.2403942. [DOI] [PubMed] [Google Scholar]
- Takamizawa J, Konishi H, Yanagisawa K, et al. Reduced expression of the let-7 microRNAs in human lung cancers in association with shortened postoperative survival. Cancer Res. 2004;64:3753–3756. doi: 10.1158/0008-5472.CAN-04-0637. [DOI] [PubMed] [Google Scholar]
- Takanami T, Mori A, Takahashi H, Horiuchi S, Higashitani A. Caenorhabditis elegans Ce-rdh-1/rad-51 functions after double-strand break formation of meiotic recombination. Chromosome Res. 2003;11:125–135. doi: 10.1023/a:1022863814686. [DOI] [PubMed] [Google Scholar]
- Tinzl M, Marberger M, Horvath S, Chypre C. DD3PCA3 RNA analysis in urine--a new perspective for detecting prostate cancer. Eur Urol. 2004;46:182–186. doi: 10.1016/j.eururo.2004.06.004. (discussion 187) [DOI] [PubMed] [Google Scholar]
- Urban-Klein B, Werth S, Abuharbeid S, Czubayko F, Aigner A. RNAi-mediated gene-targeting through systemic application of polyethylenimine (PEI)-complexed siRNA in vivo. Gene Ther. 2005;12:461–466. doi: 10.1038/sj.gt.3302425. [DOI] [PubMed] [Google Scholar]
- Vogelstein B, Kinzler KW. Cancer genes and the pathways they control. Nat Med. 2004;10:789–799. doi: 10.1038/nm1087. [DOI] [PubMed] [Google Scholar]
- Wall NR, Shi Y. Small RNA: can RNA interference be exploited for therapy? Lancet. 2003;362:1401–1403. doi: 10.1016/S0140-6736(03)14637-5. [DOI] [PubMed] [Google Scholar]
- Warnecke C, Zaborowska Z, Kurreck J, et al. Differentiating the functional role of hypoxia-inducible factor (HIF)-1alpha and HIF-2alpha (EPAS-1) by the use of RNA interference: erythropoietin is a HIF-2alpha target gene in Hep3B and Kelly cells. FASEB J. 2004;18:1462–1464. doi: 10.1096/fj.04-1640fje. [DOI] [PubMed] [Google Scholar]
- Wilda M, Fuchs U, Wossmann W, Borkhardt A. Killing of leukemic cells with a BCR/ABL fusion gene by RNA interference (RNAi) Oncogene. 2002;21:5716–5724. doi: 10.1038/sj.onc.1205653. [DOI] [PubMed] [Google Scholar]
- Yin JQ, Gao J, Shao R, Tian WN, Wang J, Wan Y. siRNA agents inhibit oncogene expression and attenuate human tumor cell growth. J Exp Ther Oncol. 2003;3:194–204. doi: 10.1046/j.1359-4117.2003.01092.x. [DOI] [PubMed] [Google Scholar]