

Abstract
Chronic wasting disease (CWD) is a neurodegenerative prion disease of cervids. Some animal prion diseases, such as bovine spongiform encephalopathy, can infect humans; however, human susceptibility to CWD is unknown. In ruminants, prion infectivity is found in central nervous system and lymphoid tissues, with smaller amounts in intestine and muscle. In mice, prion infectivity was recently detected in fat. Since ruminant fat is consumed by humans and fed to animals, we determined infectivity titers in fat from two CWD-infected deer. Deer fat devoid of muscle contained low levels of CWD infectivity and might be a risk factor for prion infection of other species.
Prion diseases are fatal neurodegenerative diseases that include Creutzfeldt-Jakob disease in humans, bovine spongiform encephalopathy (BSE) in cattle, scrapie in sheep, and chronic wasting disease (CWD) in cervids. Cross-species prion infection can occur and is responsible for the spread of BSE to humans (2). Since spread is likely due to exposure to infected tissues, it is vital to know which tissues contain infectivity. In animals such as cattle, sheep, and cervids, whose tissues are part of both the human and domestic-animal food chains, the central nervous system (CNS) has the highest propensity for infectivity. Lymphoid organs and muscles can also be positive for the disease agent, but this varies among species (1, 4, 7). We recently found prion infectivity in brown and white fat of scrapie agent-infected mice (13) and wanted to determine if fat from animals actually consumed by humans may also carry infectivity. To answer this question, we inoculated fat from two CWD agent-infected deer into susceptible transgenic mice expressing deer prion protein (TgDeerPrP mouse) (10).
Mule deer tissues.
Tissues were collected by the Colorado Division of Wildlife (DOW) from two deer (deer 1720 and 1721) infected with CWD from natural exposure. Deer 1720 was a 4.3-year-old male, and deer 1721 was a 2.3-year-old female. Clinical records indicated that deer 1720 was at a less advanced stage of CWD than deer 1721; additional clinical data were not available. Tissue samples were collected in a manner to ensure no cross-contamination. Immunohistochemistry (IHC) for PrPres using monoclonal antibody F99/97.6.1 (12) was completed by the Colorado DOW and showed that both deer had PrPres in their retropharyngeal lymph nodes (RPLN). The Colorado DOW also tested the RPLN tissue of each deer by using a Bio-Rad CWD enzyme-linked immunosorbent assay (8). Tissue from deer 1720 had an optical density (OD) score of 1.4, and tissue from deer 1721 had an OD of 1.9. These scores are consistent with deer in an early clinical phase of disease. PRNP genotypes for these deer were not determined. Uninfected tissue (perimuscular fat) was collected from a hunter-harvested free-ranging adult male mule deer in Montana. To date, no CWD has been detected in free-ranging cervids of Montana.
In our laboratory, CWD was confirmed in deer 1720 and 1721 by the presence of disease-associated proteinase K-resistant prion protein (PrPres) as shown by routine Western blotting (13) using anti-PrP monoclonal antibody L42 (R-Biopharm) or 6H4 (Prionics) and an enhanced chemiluminescence substrate (GE Healthcare). Deer 1721 had PrPres in brain and tonsil (Fig. 1A, lanes 2 and 3), and deer 1720 had PrPres present in tonsil but not in brain (Fig. 1A, lanes 8 and 9), suggesting that CWD was less advanced in deer 1720. Perimuscular, omental, perirenal, and mediastinal fat samples from both deer were negative for PrPres by Western blotting (Fig. 1A, lanes 4 to 7 for 1721). In addition to our routine protocol, we used a second protocol where PrPres is concentrated from a larger tissue sample (5). Using the latter method, we were able to load 66 mg of tissue equivalents onto each gel for each fat sample, but we still obtained negative results (data not shown). The same conditions can detect PrPres in brain and lymphoid tissues from highly positive deer with less than 0.25 mg of tissue.
FIG. 1.
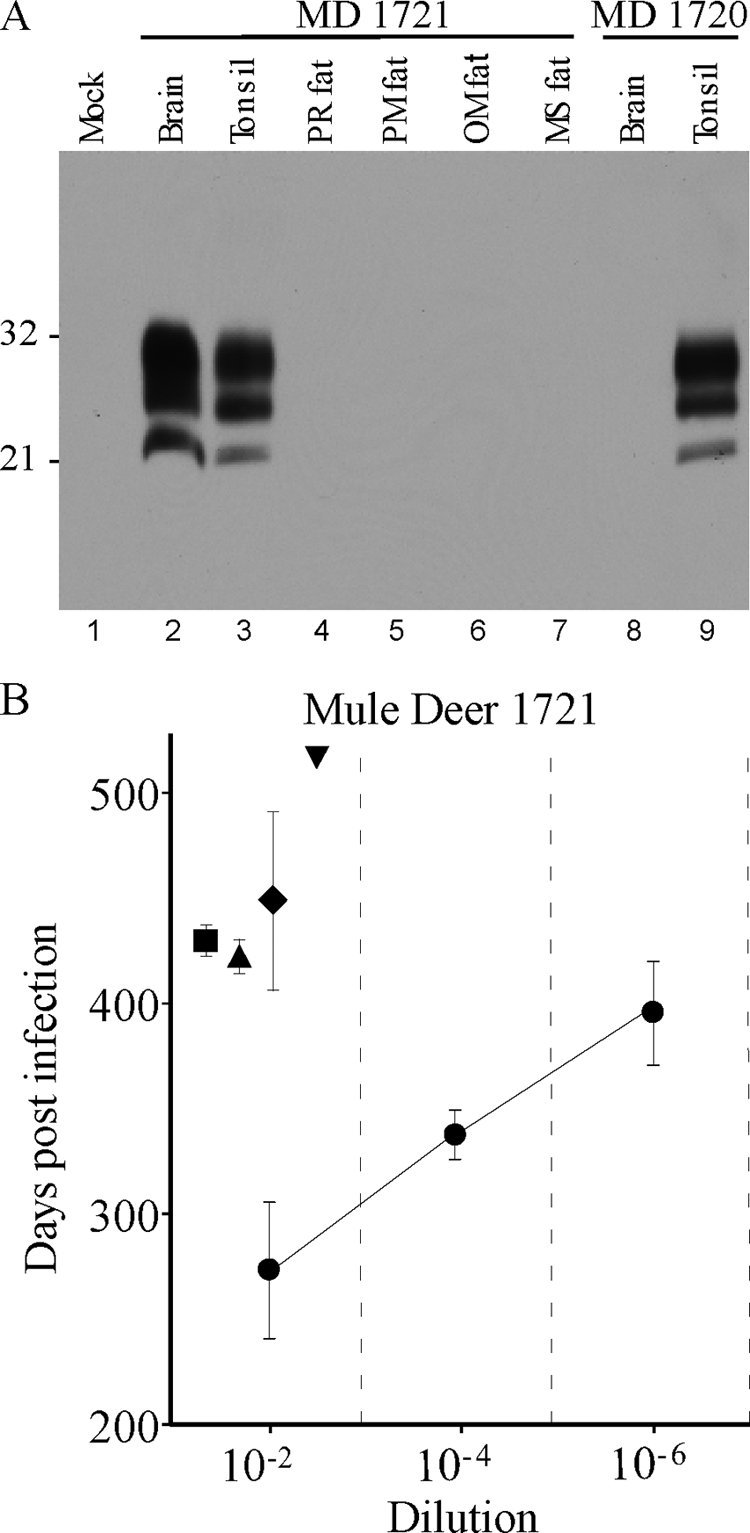
(A) Western blot of PrPres from CWD agent-infected mule deer. All samples were treated with proteinase K as described in the text. Lanes 1 and 4 to 8 were loaded with 2-mg tissue equivalents. Lanes 2, 3, and 9 were loaded with 0.5-mg tissue equivalents. Lane 1 contains uninfected deer brain, lanes 2 to 7 show tissues from mule deer (MD) 1721, and lanes 8 and 9 show tissues from mule deer 1720. The blot was probed using the anti-PrP antibody 6H4 (Prionics) by enhanced-chemiluminescence detection. Film exposure was 14 min. Numbers at the left are molecular weights (in thousands). PR, perirenal; PM, perimuscular; OM, omental; MS, mediastinal. (B) CWD incubation periods in TgDeerPrP mice following intracerebral injection of tissue homogenates from mule deer 1721. Symbols: •, brain; ▪, perirenal fat; ▴, perimuscular fat; ♦, omental fat; ▾, mediastinal fat (only one of four mice developed CWD, so there are no error bars). Note that the incubation periods found in fat at a 10−2 dilution are slightly longer than the incubation periods following inoculation of a 10−6 dilution of brain, suggesting 10,000- to 100,000-fold-lower infectivity in fat than in brain.
Bioassay of TgDeerPrP mice.
Bioassays for infectivity are considered more sensitive than immunoblotting for detection of prion agent infectivity. Therefore, dilutions of brain and fat homogenates from these deer were inoculated intracerebrally into TgDeerPrP mice. All mice were housed at the Rocky Mountain Laboratories (RML) in an AAALAC International-accredited facility, and experimentation followed NIH RML Animal Care and Use Committee-approved protocols. Fat tissues (perirenal, perimuscular, omental, and mediastinal) were finely dissected to remove any grossly evident lymphatic and neural tissues. Homogenization of each tissue was done using new, sterile, disposable tubes and homogenizer rotors and either a mini-bead beater (Biospec Products) or an OMNI tissue homogenizer (OMNI International). Inocula were diluted in phosphate-buffered balanced salt solution plus 2% fetal bovine serum, and a 50-μl volume was injected into the cerebrum of 4- to 6-week-old mice. Following inoculation, the mice were monitored for onset of clinical disease as described previously (10), and PrPres was confirmed by Western blot analysis of brain samples from infected mice. All four fat samples from deer 1721 had detectable infectivity (Fig. 1B and Table 1). Based on comparative titrations with brain tissues from deer 1721, there was 10,000- to 100,000-fold less infectivity in fat than in brain (Fig. 1B). In contrast, in deer 1720, no infectivity was found in four fat samples (Table 1). The lack of infectivity and PrPres in the brain of deer 1720 indicated that disease in this animal was at an early stage. This result also suggested that prion infectivity may occur in fat only at later stages of disease.
TABLE 1.
Bioassay of CWD agent-infected mule deer tissues in TgDeerPrP micea
| Donor | Tissue (inoculum concn) | CWD incidenceb | Incubation periodc |
|---|---|---|---|
| Deer 1720 | Brain | 0/3 | NA |
| Omental fat | 0/4 | NA | |
| Perimuscular fat | 0/3 | NA | |
| Perirenal fat | 0/4 | NA | |
| Mediastinal fat | 0/4 | NA | |
| Deer 1721 | Brain (1%) | 3/3 | 273 ± 19 |
| Brain (0.01%) | 4/4 | 338 ± 6 | |
| Brain (0.0001%) | 4/4 | 395 ± 12 | |
| Omental fat | 3/3 | 466 ± 18 | |
| Perimuscular fat | 4/4 | 423 ± 4 | |
| Perirenal fat | 4/4 | 430 ± 2 | |
| Mediastinal fat | 1/3 | 518 | |
| Uninfected deer | Perimuscular fat | 0/4 | NA |
Tissue homogenates from two CWD agent-infected and one uninfected mule deer were inoculated intracerebrally into CWD agent-susceptible TgDeerPrP mice. Unless otherwise indicated, the concentration of the inoculum was 1% (10−2 dilution). Brain and fat homogenates from mule deer 1720 and perirenal fat from mule deer 1721 were also tested at a 10−4 dilution, but no mice developed CWD (data not shown). Perimuscular, omental, and mediastinal fat from mule deer 1721 were not tested at the 10−4 dilution. Mice were observed for a minimum of 600 days.
The numerator is the number of CWD-positive mice, and the denominator is the number of mice inoculated.
Mean numbers of days of incubation ± standard errors are shown. NA, not applicable.
The highest risk of human contact with CWD might be through exposure to high-titer CNS tissue through accidental skin cuts or corneal contact at the time of harvest and butchering. However, the likelihood of a human consuming fat infected with a low titer of the CWD agent is much higher. It is impossible to remove all the fat present within muscle tissue, and fat consumption is inevitable when eating meat. Of additional concern is the fact that meat from an individual deer harvested by a hunter is typically consumed over multiple meals by the same group of people. These individuals would thus have multiple exposures to the CWD agent over time, which might increase the chance for transfer of infection.
In the Rocky Mountain region of North America, wild deer are subject to predation by wolves, coyotes, bears, and mountain lions. Although canines such as wolves and coyotes are not known to be susceptible to prion diseases, felines definitely are susceptible to BSE (9) and might also be infected by the CWD agent. Deer infected with the CWD agent are more likely to be killed by predators such as mountain lions (11). Peripheral tissues, including lymph nodes, muscle, and fat, which harbor prion infectivity are more accessible for consumption than CNS tissue, which has the highest level of infectivity late in disease. Therefore, infectivity in these peripheral tissues may be important in potential cross-species CWD transmissions in the wild.
The present finding of CWD infectivity in deer fat tissue raises the possibility that prion infectivity might also be found in fat tissue of other infected ruminants, such as sheep and cattle, whose fat and muscle tissues are more widely distributed in both the human and domestic-animal food chains. Although the infectivity in fat tissues is low compared to that in the CNS, there may be significant differences among species and between prion strains. Two fat samples from BSE agent-infected cattle were reported to be negative by bioassay in nontransgenic RIII mice (3, 6). However, RIII mice are 10,000-fold-less sensitive to BSE agent infection than transgenic mice expressing bovine PrP (4). It would be prudent to carry out additional infectivity assays on fat from BSE agent-infected cattle and scrapie agent-infected sheep using appropriate transgenic mice or homologous species to determine the risk from these sources.
Acknowledgments
We thank Byron Caughey, Sue Priola, and Mikael Klingeborn for critical review of the manuscript; Anita Mora and Gary Hettrick for graphics assistance; and the Colorado DOW for providing CWD-infected tissues.
Funding for this work was provided by the National Institute of Allergy and Infectious Diseases, Division of Intramural Research.
Footnotes
Published ahead of print on 1 July 2009.
REFERENCES
- 1.Angers, R. C., S. R. Browning, T. S. Seward, C. J. Sigurdson, M. W. Miller, E. A. Hoover, and G. C. Telling. 2006. Prions in skeletal muscles of deer with chronic wasting disease. Science 311:1117. [DOI] [PubMed] [Google Scholar]
- 2.Beringue, V., J. L. Vilotte, and H. Laude. 2008. Prion agent diversity and species barrier. Vet. Res. 39:47. [DOI] [PubMed] [Google Scholar]
- 3.Bradley, R. 1996. Bovine spongiform encephalopathy distribution and update on some transmission and decontamination studies, p. 11-28. In C. J. Gibbs, Jr. (ed.), Bovine spongiform encephalopathy. The BSE dilemma. Springer-Verlag, New York, NY.
- 4.Buschmann, A., and M. H. Groschup. 2005. Highly bovine spongiform encephalopathy-sensitive transgenic mice confirm the essential restriction of infectivity to the nervous system in clinically diseased cattle. J. Infect. Dis. 192:934-942. [DOI] [PubMed] [Google Scholar]
- 5.Chesebro, B., M. Trifilo, R. Race, K. Meade-White, C. Teng, R. Lacasse, L. Raymond, C. Favara, G. Baron, S. Priola, B. Caughey, E. Masliah, and M. Oldstone. 2005. Anchorless prion protein results in infectious amyloid disease without clinical scrapie. Science 308:1435-1439. [DOI] [PubMed] [Google Scholar]
- 6.Fraser, H., and J. D. Foster. 1994. Transmission to mice, sheep and goats and bioassay of bovine tissues, p. 145-159. In R. Bradley and B. Marchant (ed.), Transmissible spongiform encephalopathies. Proceedings of a consultation on BSE with the Scientific Veterinary Committee of the Commission of the European Communities. CEC, Brussels, Belgium.
- 7.Hadlow, W. J., R. C. Kennedy, and R. E. Race. 1982. Natural infection of Suffolk sheep with scrapie virus. J. Infect. Dis. 146:657-664. [DOI] [PubMed] [Google Scholar]
- 8.Hibler, C. P., K. L. Wilson, T. R. Spraker, M. W. Miller, R. R. Zink, L. L. DeBuse, E. Andersen, D. Schweitzer, J. A. Kennedy, L. A. Baeten, J. F. Smeltzer, M. D. Salman, and B. E. Powers. 2003. Field validation and assessment of an enzyme-linked immunosorbent assay for detecting chronic wasting disease in mule deer (Odocoileus hemionus), white-tailed deer (Odocoileus virginianus), and Rocky Mountain elk (Cervus elaphus nelsoni). J. Vet. Diagn. Investig. 15:311-319. [DOI] [PubMed] [Google Scholar]
- 9.Kirkwood, J. K., and A. A. Cunningham. 1994. Epidemiological observations on spongiform encephalopathies in captive wild animals in the British Isles. Vet. Rec. 135:296-303. [DOI] [PubMed] [Google Scholar]
- 10.Meade-White, K., B. Race, M. Trifilo, A. Bossers, C. Favara, R. Lacasse, M. Miller, E. Williams, M. Oldstone, R. Race, and B. Chesebro. 2007. Resistance to chronic wasting disease in transgenic mice expressing a naturally occurring allelic variant of deer prion protein. J. Virol. 81:4533-4539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Miller, M. W., H. M. Swanson, L. L. Wolfe, F. G. Quartarone, S. L. Huwer, C. H. Southwick, and P. M. Lukacs. 2008. Lions and prions and deer demise. PLoS. One 3:e4019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Miller, M. W., and E. S. Williams. 2002. Detection of PrP(CWD) in mule deer by immunohistochemistry of lymphoid tissues. Vet. Rec. 151:610-612. [DOI] [PubMed] [Google Scholar]
- 13.Race, B., K. Meade-White, M. B. Oldstone, R. Race, and B. Chesebro. 2008. Detection of prion infectivity in fat tissues of scrapie-infected mice. PLoS. Pathog. 4:e1000232. [DOI] [PMC free article] [PubMed] [Google Scholar]