

Abstract
We sought to understand the basis of performance variability and perceptual learning in saccadic visual search. Four subjects searched for a target based on its shape in a linear array of densely packed, regularly spaced items, a configuration used to simplify the analysis of performance and to minimize search strategy variability. We measured the dependence of performance—search speed—on the oculomotor variables of fixation duration and saccade amplitude, both within and across experimental sessions. We also measured perceptual span, the area in visual space in which subjects could identify the target above chance, with a modified version of the task using a gaze-contingent display with transiently appearing targets. The principal finding of this study was that both within and across sessions, saccade metrics accounted for much more of the variability and improvement in performance than did fixation duration. Increases in search speed were due primarily to the processing information from a greater area of the visual field, rather than processing information from a fixed area more quickly, though there was a small but consistent decrease in fixation duration across sessions. The increase in performance derived from an increase in perceptual span and not merely an increase in subjects’ efficiency in ‘tiling’ the search array with regions of visibility.
Keywords: Saccade, visual search, human, perceptual learning, fixation, attention
INTRODUCTION
Visual information is acquired rapidly by redirecting the eye using saccadic eye movements. Saccades occur 2–4 times per second; the frequency and precision of these movements, along with their relative ease of measurement, makes saccadic behavior not only a worthy subject of study in its own right, but also a good model for investigating how the brain processes visual information during goal-directed behavior. Our particular interest in this paper concerns the oculomotor strategy used during visual search, how performance depends upon metrics of saccadic eye movements, eye fixations and perceptual discrimination, and how changes in these metrics with practice affect search performance.
A priori, visual search performance depends both on the duration of eye fixations and the size of saccades, and previous work has shown that both these oculomotor variables correlate broadly with overall performance across search tasks of varying difficulty. (Jacobs 1986; Zelinsky & Sheinberg 1995). These oculomotor variables are, in turn, dependent on what useful visual information can be acquired during a single fixation and how fast this information can be processed. Previous work has indicated that the size of saccades made while acquiring visual information reflects “perceptual span” (Rayner 1998), defined as the spatial region about a fixation from which task-relevant information is obtained during scanning. Related concepts in the literature are “decision region” (Rayner & Fisher 1987), “conspicuity area” (Motter & Belky 1998a, b), “discrimination window” (Geisler & Chou 1995), “visual span” (Jacobs 1986; Jacobs & O’Regan 1987), and “visual lobe” (Sekuler & Ball 1986; Ball, Beard, Roenker, Miller, & Griggs, D. S. 1988; Gramopadhye, Drury, Jiang, & Sreenivasan, 2002). The last three of these concepts, however, differ from perceptual span as defined by Rayner in that they are based on measures taken during a static, no-saccade task in which stimuli are briefly presented. To distinguish between perceptual measures obtained during saccadic behavior and extended fixation, we will define saccadic perceptual span as the region about the point of fixation within which the target can be identified in a saccadic visual search task, and use fixational perceptual span to refer to this region as defined in a static task. Saccadic perceptual span has been found to correlate with visual search time (Bertera & Rayner 2000); researchers who have measured the fixational perceptual span have found that, not surprisingly, it shrinks if target/distracter similarity is increased, and that correspondingly, visual search time increases (Gordon 1969; Jacobs 1986, Jacobs & O’Regan 1987; Geisler & Chou 1995; Courtney & Chan 1986). In particular, Geisler and Chou (1995) and Jacobs (1986) were able to account for 80–90% of the variance in search time on the basis of fixational perceptual span size. Other studies of visual search using eye movements have also supported the idea that searchers adapt their eye behavior to the size of their saccadic perceptual span (Motter & Belky 1998a; Nasanen, Ojanpaa, & Kojo 2001).
While saccade size during visual search reflects both fixational and saccadic perceptual span—though this has been shown less definitively in the latter case—fixation duration appears to depend less on the visual composition of the area to be searched. Motter & Belky (1998a) monitored the eye movements of two macaque monkeys while they performed classical feature and conjunction search tasks. They found that as the density of the search array was varied, fixation duration changed much less than saccade size. Jacobs (1986) varied target/distracter similarity widely in a linear search task, producing a 6.5-fold change in mean forward saccade amplitude, while merely doubling mean fixation duration. Moreover, the variability of forward saccade amplitude in Jacobs’ data exceeded that of fixation duration in all but the most difficult search condition (in which the target differed from the distracters by one pixel). A subsequent study (Jacobs & O’Regan 1987) found qualitatively similar results between saccade size measured in degrees of visual angle and fixation duration, although manipulations of different experimental variables were found to affect fixation duration and saccade size to different degrees. McCarley, Kramer, Boot, Peterson, Wang, & Irwin (2006) found, for searches of displays containing multiple targets, that total search time correlated much more strongly with the number of gazes (comprising all successive fixations to a single stimulus item) than it did with fixation duration. They hypothesized that the differences in reaction time across conditions were due to corresponding changes in saccadic perceptual span. Zelinsky & Sheinberg (1997) found no relation between the duration of the initial fixation and response time in a visual search task, while correlating response time significantly with the number of fixations made. Pelz & Canosa (2001) used a wearable eyetracker to measure eye movements in mobile, unconstrained subjects as they walked to a restroom, washed their hands, and walked back. Our visual inspection of their data indicates again that fixation durations are less variable than saccade amplitudes. Thus while researchers routinely modulate fixation duration through manipulation of experimental variables such as target/distracter similarity (Hooge & Erkelens 1999; Shen et al. 2003), viewing distance (Jacobs & O’Regan 1987) and others, there is evidence that, at least for all but the most demanding searches, fixation durations constitute a relatively stable component of the oculomotor strategy.
However, these findings leave several questions unanswered regarding the relative contributions of fixation duration and saccade-related metrics to performance. For search at a fixed difficulty level, some searches will be faster and some slower, and the oculomotor basis of this variability is not known. Do subjects turn out particularly fast trials by using fewer fixations (and longer saccades), fixations of shorter duration, or both? Similarly, the oculomotor basis of visual learning in a search task across sessions is unknown. Does improvement in performance derive from an increase in saccadic perceptual span? Or does it result instead from the subject ‘tiling’ the search array more efficiently with regions of saccadic perceptual span—i.e. shifting gaze such that there is less overlap between the areas visible at each fixation. This latter hypothesis is motivated by findings indicating that subjects do not, at least initially, pursue a ‘pure’ tiling strategy in visual search (Geisler & Chou, 1995; Bertera & Rayner, 2000), nor are fixations in reading distributed this way (Rayner 1998). Rather, the extent of saccadic perceptual span is larger than the amplitude of the average saccade.
These questions are addressed in the present study with a linear search task, in which subjects searched from top to bottom through a dense, regular column of visual stimuli for one of four targets. Given our goal in this study—to systematically measure oculomotor variables and saccadic perceptual span—a linear search task, and not a two dimensional task, is appropriate for four reasons. First, a search task with a well-defined search path allows the use of search speed, and not merely mean reaction time, as a performance variable. Search speed is well-defined for a single trial, whereas mean reaction time is only well-defined over a relatively large number of trials. Therefore, a linear search array allows for between-trial comparisons within a session. Second, a regular, linear array reduces the amount of variability in the search path by eliminating a degree of freedom in the search path trajectory. Third, since the array is homogenous, the coarse configurational properties of the stimuli as they appear on a given fixation are known in advance. Thus, the variability of saccade amplitude in this task should be relatively low both within and across trials, making the effects of perceptual learning easier to observe. Finally, using a dense linear array simplifies the measurement of perceptual span. Psychophysical thresholds are easier to obtain when there is only one spatial dimension to assess, and the regularity of the array allows us to probe target discriminability at different distances from the point of fixation in a controlled way.
Using this task, we determined whether saccade- and fixation-related metrics change as a function of performance, both within and across sessions. Thus we determined whether the visual system learns by ‘seeing more’ in a single fixation, rather than by reducing the amount of time spent processing a set amount of information. Answers to these questions should contribute to our understanding of the dynamics of visual information processing during everyday visual search. Our results have been previously published in abstract form (Phillips & Edelman 2005).
METHODS
General
Eye movements of 4 subjects were recorded at 500Hz using video oculography (EyeLink II, Sensorimotor Instruments). Stimuli were presented on a 22” Compaq P1220 CRT monitor, set to 1024×768 resolution with a refresh rate of 85Hz. The computer ran an in-house program written in C using the Vision Shell libraries (Comtois, 2003). Subjects sat 62cm from the monitor and had their heads stabilized with a full impression bite bar. The research was performed under a protocol approved by the Institutional Review Board of The City College of New York and the CUNY Medical School.
Subjects
4 subjects participated in this experiment. 2 subjects (hh and jj) were the authors, and the other two (pp and zz) were naïve to the purposes of the experiment. All subjects were experienced psychophysical observers.
Stimuli and procedure
Subjects ran 2–4 sessions a week for three weeks, each completing a total of 10 sessions. Each session consisted of 10 blocks of 25 trials.
At the beginning of each trial, subjects fixated a small square at the top of the screen; after a 700–1000ms variable delay it disappeared and 30 rows of 3 boxes appeared. The target was one of four “Landolt C”-like boxes with a notch on one of the four sides (figure 1a). Subjects did not know in advance which of the four targets would be presented on a given trial. All stimuli were 0.45deg long per segment and segments had a thickness of 0.074deg. The distance from the center of one box to the center of its horizontal and vertical neighbors was 0.62deg.
Figure 1.

a) The four targets. b) Example stimuli and scanpaths, from subject zz. The initial fixation square (top) disappeared when stimuli were shown. The target is shown in red for expository purposes. The strikethrough black square on the scanpath indicates the keypress, immediately after which the second fixation square was displayed. The blue bars over each fixation record the duration of that fixation in ms (1deg visual angle corresponds to 100ms). The strikethrough red disk on the right scanpath indicates the position of the eye when the transient target was displayed, i.e. when a distracter was replaced with the target. The target was replaced by a distracter during the next saccade (not marked). In all trials, target appearance was restricted to the center column.
Subjects were instructed to find the target and report its orientation as quickly as possible while minimizing the number of errors (figure 1b). They were instructed to scan from top to bottom only, and received an error message displayed on the screen and heard a ‘beep’ from the computer speaker if they made a regressive (upward) saccade of more than 5deg. Subjects were instructed to guess the target orientation if they reached the bottom of the list without having seen the target. Target placement was restricted to the center column of boxes, and subjects were apprised of this. The flanking columns were added in order to help equalize the discriminability of the 4 target orientations, as well as to increase task difficulty. Responses (4AFC) were recorded by key press, after which a second fixation square appeared for 750ms. Subjects received a feedback screen after every trial. On correctly performed trials, this screen displayed their speed for the trial just completed and their cumulative error rate for that block. On error trials, the screen displayed an appropriate error message and subjects heard a ‘beep’ from the computer. Subjects controlled when the program advanced to the next trial by key press.
Trial types
There were two trial types, trials with stationary (80%) and with transient (20%) targets. In the stationary trials the target was in the display in a fixed location for the entire trial. In transient trials, the target was displayed only for the duration of a single fixation; otherwise the location contained a distracter. The change was made in mid-saccade. Transient targets were distributed ahead and behind the point of fixation in equal proportions. Otherwise, the two trial types were identical, although they were analyzed differently (see Results--Saccadic perceptual span and search speed).
Calibration
Subjects were calibrated at the beginning of each session using the 9-point calibration routines built into the EyeLink II and Vision Shell software packages in succession. In addition, a drift correction was performed between each block of trials. In post hoc analysis, a MATLAB program was used to identify the fixation made to the second fixation square (See ‘Stimuli and procedure’), and the scanpath from each trial was translated so that this fixation overlaid the square.
Data Analysis
Our offline analysis program identified a saccade as occurring when the eye velocity (lowpass filtered, 200Hz Butterworth) exceeded 40deg/sec until it fell below 10deg/sec, with a total duration of at least 8 ms. We did not use the entire eye position trace in our analysis. Rather, we defined the ‘scanpath’, the trajectory of eye position during search, to begin at the beginning of the first fixation within the top 2deg of the list, and to end at the end of the fixation prior to the fixation during which the key press was made. Hence, the fixation during which the key press was made, as well as the saccade immediately prior to it, were not considered in the calculation of any of the oculomotor data. We did this because the duration of the fixation during which the keypress was made tended to be much longer than the average fixation duration, and its duration varied widely, from <100ms to >1000ms. This variation includes variation derived from variability in the manual response. The last saccade prior to the keypress was generally much shorter than average and so it too was excluded. Possibly as a result of these measures we found very few (0.13%) corrective saccades (latency < 60ms).
Search time, then, was defined as the amount of time used to traverse the scanpath. Of course, search time will be faster for targets located near the top of the array. Therefore, the metric of performance we used was search speed, defined as search time divided by the number of items from the beginning of the list to the target, inclusive. This quantity, items per second, describes the rate at which the subject traverses the list during search and is thus distinct from the traditional measure of search time or reaction time used to assay performance in visual search tasks. We correlated this performance measure with the following three oculomotor variables: items scanned per fixation, defined as the number of fixations in the scanpath divided by the number of items from the beginning of the list to the target, inclusive; forward saccade amplitude, defined as the mean amplitude of the saccades made in the top-to-bottom direction, and fixation duration. We considered items scanned per fixation and forward saccade amplitude to be saccade-related metrics and fixation duration to be a fixation-related metric. Only trials with a stationary target (80% of all trials) were used in the calculation of these quantities. Both forward saccade amplitude and items scanned per fixation have been used as indirect measures of span in the literature (Jacobs 1986; Ojanpaa, Nasanen & Kojo 2002). They are however different metrics; items scanned per fixation but not forward saccade amplitude will reflect regressive saccades that were made, placement of the initial fixation within the search array, and the distance of the final fixation in the scanpath to the target. Rather than decide a priori which measure is a better indicator of oculomotor strategy and saccadic perceptual span, we use both in our analysis. They are, as we will see, empirically distinct. Our direct measurement of span using transient target trials will be described in Results—Saccadic perceptual span and search speed below.
Of all stationary trials, many (hh: 32%, jj: 39%, pp: 32%, zz: 34%) were excluded from analysis. The reasons for excluding these trials included: trials in which the scanpath contained only one fixation, or subject made a single saccade directly to the target (hh: 17%, jj: 13%, pp: 17%, zz: 18%), in which case forward saccade amplitude was undefined; trials in which an incorrect keypress was made (hh: 5%, jj: 11%, pp: 5%, zz: 5%); trials in which the keypress occurred when the eye was fixated more than 5 degrees below the target (hh: 7%, jj: 5%, pp: 0.5%, zz: 1%), a distance at which the ability to discriminate was almost completely lost, and which usually arose from the subject having passed the target on the way down, reached the end, and (presumably) guessed; and trials in which the cumulative amplitude of regressive saccades was over 5deg (hh: 2%, jj: 3%, pp: 1%, zz: 1%). Remaining excluded trials consisted of trials in which there was either a blink or an initial fixation error.
All analysis of the data was performed using routines written in the MATLAB (MathWorks) software, and Sigma Plot (SPSS) programming environments.
RESULTS
Subjects’ search strategies
Subjects systematically made saccades in the downward direction until they reached the target, typically making 3–5 fixations along the way (figure 1b). The percentage of regressive saccades was low across subjects and sessions (hh: 4%, jj: 2%, pp: 9%, zz: 3%). The percentage of trials within a session which contained at least one regressive saccade was correspondingly low, though not negligible (hh: 7%, jj: 5%, pp: 18%, zz: 5%). There were no significant changes in regressive saccade frequency across sessions. Thus, regressive saccades were not a consistent part of subjects’ search strategy. Rather, as expected, subjects’ search strategy was to make saccades downward through the search array until the target was found.
Within-session comparison of overall search speed with saccade and fixation metrics
Within every session and for each subject, the correlation between search speed and items scanned per fixation, forward saccade amplitude, and fixation duration was examined. For 3 of the 4 subjects there was no significant change in these correlations across sessions; for subject pp modest increases in r value (Pearson product-moment), on the order of 0.1, were observed. Therefore, we collapsed data across sessions and compared average correlations for the different independent variables for the entire data set.
Items scanned per fixation had by far a stronger and more consistent correlation with search speed than did saccade size or fixation duration (figure 2). Across all subjects and sessions, the correlation between search speed and rows per fixation was the strongest (minimum r = 0.82, mean r = 0.92) and exhibited the least variation. The correlation between mean forward saccade amplitude and search speed was substantially weaker (mean r = 0.50), but still much stronger than the correlation between mean fixation duration and search speed (mean absolute r = 0.17). The correlation between fixation duration and search speed, moreover, was usually not significant at the 0.05 level; for 15 of 40 sessions across the four subjects these correlations were significant, vs. 40/40 and 39/40 for items scanned per fixation and forward saccade amplitude, respectively. The ordinal relationship between correlation strengths between search speed and the three measures (rows scanned per fixation—highest, forward saccade amplitude—intermediate, and fixation duration—lowest) present in session averages also held across all individual sessions.
Figure 2.

Mean correlations of oculomotor variables with performance for each subject, averaged across sessions. Error bars = 1 SE. The correlations between items scanned per fixation and performance were always (40/40) significant at the 0.05 level, and the correlations between forward saccade amplitude and performance were almost always (39/40) significant, whereas the correlations between fixation duration and performance typically were not (15/40).
In addition, the comparison for each subject of fast and slow trials from all 10 sessions combined also indicates that search speed depends on variability in saccade-based metrics rather than variability in fixation duration (figure 3). We considered here trials in which the target occurred in the bottom half of the array, as there was a clear dependency between target placement and search speed when we considered all trials (slow trials tended to be ones where the target was near the beginning of the array). This analysis shows in a qualitative way that the fastest trials exhibit substantially larger mean forward saccade amplitude and fewer fixations per unit distance along the search array to the target. The difference in fixation duration between fast and slow trials, while perceptible for some subjects, is visibly much smaller in magnitude.
Figure 3.

Schematic average scanpaths of the slowest 25% and fastest 25% of all trials in which the target occurred in the bottom half of the search array, for each subject. Only mean fixation locations are shown; eye position during the saccade is not. Abscissa markings now designate fixation duration. Search speed was defined as time spent searching divided by the distance of the target from the beginning of the list; hence trials with equivalently placed targets could differ with respect to the amount of time taken to find the target. The gray box indicates the average target location for the corresponding trials. Horizontal bars denote the mean duration of the corresponding fixation; whiskers are 1 SD of the mean. The dotted vertical line at 200ms is for reference purposes.
There was considerable variability in performance across trials, as well as in the independent variables we measured; this is illustrated by the correlation data from a representative session (figure 4). It seems unlikely that this variability is due to subjects making a speed-accuracy tradeoff, e.g. subjects increasing their search speed in the high-performance trials by processing only portions of the array and ‘skipping over’ the rest, as there was no corresponding difference in error rates among subjects for fast versus slow trials analyzed in figure 3. Incorrect keypress errors changed significantly by t-test between slow and fast trials for only one subject, hh, and in this case decreased from 9% to 3%; error rates for other subjects increased by <3%, non-significantly. Errors due to either too much cumulative regressive saccade amplitude or to a keypress made over 5deg beyond the target, <9% across subjects and sessions, were more likely or equally likely to occur in slow trials rather than fast trials; this is no doubt due to the deleterious effect on search speed of failing to make a saccade that lands near the target. Furthermore, a) subjects generally performed the task correctly—in particular, the total error rate of the naïve subjects, pp and zz, was 7% or less, b) the incidence of regressive saccades was low, and c) saccadic perceptual span was generally larger than mean forward saccade amplitude (see Results—Across-session comparisons below). For all of these reasons we assume that subjects’ performance variability reflects genuine variability in visual search efficacy from trial to trial when performing the task correctly.
Figure 4.
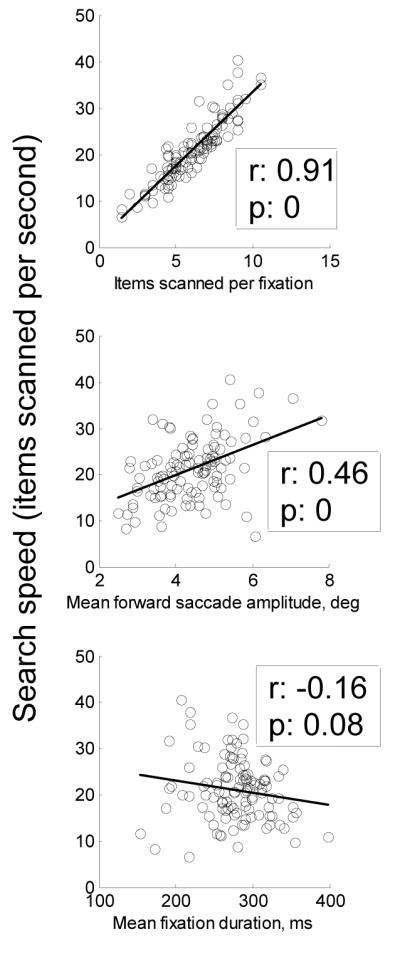
Correlations between performance on the search task, defined as search speed (items scanned per second), and oculomotor variables for representative session (jj, session 10). The ordinal relationship between correlation strengths—strongest for items scanned per fixation, moderate for mean forward saccade amplitude, and weakest for mean fixation duration—held for every individual session.
We think the picture of within-session variability that arises from these data is quite clear. Subjects achieve the highest search speeds when few fixations are made and the target is detected peripherally at large eccentricities. Significant shortening of fixation duration to increase performance does not occur on a trial-to-trial basis. These results are consistent with the hypothesis that variability in search speed from trial to trial is due to variability in the spatial extent of perception from trial to trial—how much visual information the visual system processes—as opposed to variability in how quickly visual information from a fixed area is processed.
Across-session comparisons of overall search speed and saccade and fixation metrics
Comparing performance for the last of the ten sessions with that of the first, search speed improved significantly in all four subjects (figure 5). We measured the proportional change from the first session to the last, taking the corresponding values from the best linear fit. Consistent with our results for within-session comparisons, the magnitude of the change in the saccade-based variables was much larger than the magnitude of the change for fixation duration (figure 6). All subjects showed either a significant increase in these two measures (including both naïve subjects) or a trend in this direction. Fixation duration decreased significantly for all subjects, but the magnitude of this change, 10.5% averaged across subjects, was small compared to the change in saccade-based variables, 22.2% and 40.5% for items scanned per fixation and forward saccade amplitude, respectively. Therefore, learning in this task involved increasing the area scanned in a given fixation much more than it did increasing the time spent fixating. Nonetheless, the consistent decline in mean fixation duration is notable given the absence of any significant effect of fixation duration on performance within a session, and suggests that a very gradual, long-term decrease in the time required to process information acquired from a single fixation took place. From the first to the last session, the decreases were as follows: hh, 12ms; jj, 25ms; pp, 47ms; zz, 28ms, calculated from the best linear fit to the data.
Figure 5.

Across-session correlations. Performance as well as all of the oculomotor variables in the search task correlated significantly with session number for at least 2 of the 4 subjects. The correlations were always significant for both naïve subjects. hh: triangle, jj: star, pp: square, zz: disk.
Figure 6.

Proportional change across sessions in performance and the oculomotor variables. Change was calculated using the equation (Xend − Xbegin)/Xend, where Xbegin is the value of the best linear fit (least squares) at session 1 and Xend is the value of this function for session 10. Asterisks indicate where the correlation was significant at the 0.05 level.
Saccadic perceptual span and search speed
The findings from across-session comparsions described above imply the saccadic perceptual span increases with task experience. To examine this directly, we measured saccadic perceptual span by presenting the target transiently for a single fixation on 20% of all trials. The target appeared (replacing a distracter) while the eye was in mid-saccade, stayed in the display for the duration of the subject’s following fixation, and was replaced with a distracter when the subject made the next saccade. In contrast to stationary target trials, no transient target trials were excluded on the basis of scanpath. As described in Methods, the location of the target relative to fixation was chosen randomly about the current fixation position; off-line analysis showed that targets placed ahead of fixation appeared on average 2.8deg ahead of fixation (hh: 2.3deg, jj: 2.8deg, pp: 3.1deg, zz: 3.0deg), and targets placed behind fixation appeared on average 2.5deg behind fixation (hh: 2.4deg, jj: 2.6deg, pp: 2.6deg, zz: 2.4deg). For all subjects, at least 94% of targets appeared within 6deg of fixation. We measured forward span—the region of visibility below fixation, i.e. in the forward direction of search—and backward span, the region of visibility above fixation, simultaneously by fitting a function composed of two Weibull cumulative distribution functions which modeled the respective forward and backward halves of the data set:
This procedure ensured that the model function was continuous at x=0, while allowing the slope (β1, β2) and inflection (α1, α2) parameters of the two component cdfs to vary independently. Parameter γ was fixed at chance (0.25), and λ was restricted to the interval [0, 0.75]. This function was fit to the corresponding data sets in each session, illustrated with an example session (figure 7). The spatial region comprising the subject’s saccadic perceptual span for that session was then defined as the portion of the visual field within which target identification was 62.5%, i.e. halfway between ceiling (100%) and chance on the 4-AFC task (25%), or better. In 5 of 40 sessions across the four subjects the fitting algorithm identified the threshold as eccentric to the most eccentric data point in either the forward or backward directions, due to insufficient or noisy data; these sessions were not analyzed.
Figure 7.

Measuring saccadic perceptual span directly. Transient targets (20% of trials) appeared for the duration of a single fixation; they replaced a distracter during a saccade, and were replaced by a distracter during the next saccade. The retinal eccentricity of the target, and the response (correct/incorrect) were collected, and the data were then fit with cumulative normal distribution functions—one for targets appearing ‘ahead’ of fixation and one for targets appearing ‘behind’ fixation. The shaded region comprising the functional field of view was defined arbitrarily by the point at which the psychophysical curves crossed 62.5% correct, halfway between chance and ceiling. Small x’s near the horizontal baseline denote eccentricities at which no data was collected. The bin at 8° within which the response rate is 100% contains 1 trial.
Saccadic perceptual span was usually (20/35 sessions) larger than mean forward saccade amplitude; on average, saccadic perceptual span was 1.06 times the size of forward saccade amplitude (maximum 1.62, maximum 0.65). This result depends on our arbitrarily chosen measure of span threshold (62.5%), but nonetheless suggests that subjects managed a fairly efficient tiling of the search array. Moreover, the ratio of forward saccade amplitude to saccadic perceptual span—search efficiency—did not improve significantly in any of the subjects across sessions, nor was there any correlation in the pooled, z-transformed data (p=0.79).
The saccadic perceptual span was approximately centered around fixation. Mean backward and forward spans for each subject were as follows: hh, (−2.9, 2.3); jj, (−2.3, 1.3); pp, (−2.7, 3.0); zz, (−2.8, 2.7); all measurements in degrees. For all subjects except jj, the mean difference between (absolute) forward and backward span was not significantly different from 0 by t-test; in this subject’s case backwards span was larger than forwards span. There was also no significant change in the difference between forward and backward span across sessions, neither for individual subjects nor for group data pooled by z-transformation.
There was a general increase in span from the first to the last session in which span was measured. The increase in span was significant across sessions in 2 of 4 subjects, with a positive trend in the other two subjects (figure 8a). The increase in span was also strongly significant when subjects’ z-transformed data were pooled together (figure 8b).
Figure 8.

Saccadic perceptual span across sessions. The correlation between saccadic perceptual span and task experience was calculated a) for each individual subject (hh: triangle, jj: star, pp: square, zz: disk), and b) across subjects using the z-transformed, pooled data.
In conclusion, we found that saccadic perceptual span was on average only slightly larger than forward saccade amplitude, and that it increased with experience. In the absence of any tendency for search efficiency to increase over time among subjects, we rule out the possibility that subjects improve on the task by minimizing the extent to which regions of saccadic perceptual span overlap, i.e. ‘tiling’ the search array more effectively. We also found no evidence that span was biased in the direction of search; subjects were no better at detecting the target when it appeared ahead of fixation as they were when it appeared behind.
DISCUSSION
To better understand the perceptual and oculomotor determinants of visual search performance, we employed a linear-array search task in which the overall speed of search, measured as items scanned per second, could be measured on individual trials. Overall search speed was compared to three oculomotor variables: (i) items scanned per fixation, the number of fixations in the scanpath divided by the number of items from the beginning of the list to the target, inclusive; (ii) forward saccade amplitude, the mean amplitude of the saccades made in the top-to-bottom direction; and (iii) fixation duration. Within sessions, we found with striking regularity that the overall speed of search correlated much more strongly with saccade-based metrics of oculomotor behavior—items scanned per fixation and forward saccade amplitude—than with fixation duration. Across sessions, search speed increased significantly in all subjects. The overall improvement in performance across the ten sessions appeared to depend on the size of saccades during search and the number of items scanned per fixation and was reflected in an increase in saccadic perceptual span—the visual area in which targets could be detected during a single fixation—as subjects gained additional experience in the task.
Although our two span-related oculomotor metrics, forward saccade amplitude and items scanned per fixation, were both more determinative of search speed than fixation duration, they were not identical. Forward saccade amplitude increased more than items scanned per fixation across sessions despite the fact that, as in within-session comparisons, it correlated to performance somewhat less strongly than did items scanned per fixation (mean r’s of 0.88 and 0.95, respectively). We attribute this to the fact that forward saccade amplitude is one of several factors which determines items scanned per fixation, and the others—placement of initial fixation, distance of the target to the last fixation, and proportion of regressive saccades—did not change significantly in any of the subjects across sessions. Thus although items scanned per fixation is a better predictor of performance both within and across sessions, it is the increase in forward saccade amplitude which drives the increase in performance. Particularly since the proportion of regressive saccades did not change across sessions, we take this finding, along with the lack of improvement in search efficiency (see Results—Saccadic perceptual span and search speed) to be evidence that improvement on this task derives primarily from an increase in saccadic perceptual span rather than any change in search strategy.
The dependence of performance on the spatial determinants of oculomotor performance—items scanned per fixation and forward saccade size, rather than fixation duration—is consistent with previous analyses of search behavior. The correlation we found between items scanned per fixation and search speed is mathematically equivalent to a correlation between total search time and number of fixations; this correlation has been previously observed, within sessions, by Zelinsky & Sheinberg (1997) and by Motter & Belky (1998a). Hooge & Erkelens (1996) found that subjects performing a search of 7 items located in a circular array made many (~25% of total) return saccades, in which the subject fixated the target, moved their eyes to a new location, then refixated the target. As they noted, return saccades should be rare if fixation duration is tailored to the information processing needs of a particular fixation. Hooge & Erkelens (1996) also found that 2 of their 4 subjects adjusted their fixation duration to the difficulty level of the task (small, medium, or large target/distracter similarity) to a much greater degree in experimental sessions in which trials of different difficulty were mixed together within a block, as opposed to sessions in which trials were blocked by difficulty level. The remaining two subjects (the authors) exhibited less overall dependency of fixation duration on task difficulty.
In addition, Motter & Belky (1998a) monitored the eye movements of two macaque monkeys while they performed classical feature and conjunction searches of arrays of varying density and size. They did find that fixation duration was modulated somewhat by the density of the array; a conjunction search through the most sparse array required a mean of 215 ms/fixation while the monkeys averaged 175 ms/fixation to search through a dense array, but this difference was due primarily to skewness in the duration distribution at small array sizes and was not reflected in the median values. A subsequent analysis showed that fixation duration for a given array size was independent of the number of nearby search items, suggesting that fixation duration did not depend on the local stimulus properties in the neighborhood of the fixation. Altogether, these results provide support for the idea that, as Hooge & Erkelens (1996) have suggested, fixation durations in visual search do not vary substantially, at least relative to array type.
The results described here are also consistent with the relatively few studies of perceptual learning that have investigated the spatial extent of perception, in the form of either saccadic or fixational perceptual span. There have been several studies which investigated the effect of learning on fixational perceptual span, with many (Chung, Legge, & Cheung 2004; Green & Bavelier 2003; Gramopadhye et al. 2002; Sekuler & Ball 1986; Ball et al. 1988), but not all (Leachtenauer 1978) finding an increase in fixational perceptual span with training. No prior study we know of has investigated the effect of learning on saccadic perceptual span per se.
On the other hand, our finding that span does not exhibit a forward bias may seem surprising in light of previous studies of the relationship between perception and saccades (Kowler, Andersen, Dosher & Blaser 1995; Hoffman & Subramaniam 1995; Gersch, Kowler & Dosher 2004; Deubel & Schneider 1996), in which a perceptual advantage in the area of the impending saccade is shown. Such results suggest the hypothesis that span would exhibit a forward bias in our task. However, in those tasks, unlike ours, saccade targets are instructed by the experimenter rather than chosen by the subject. The attentional demands created by the instruction itself may result in a perceptual advantage at or near the saccade target. In addition, in our task, foveal proximity was the determining factor as to whether a given search item was perceived, overwhelming any perceptual advantage caused by attentional enhancement at the impending saccade goal. Likewise, we speculate that task differences explain the contrast between our finding with respect to the shape of span and the asymmetry in saccadic perceptual span that has been found for reading, in which subjects’ span is skewed sharply in the direction of eye movement (Rayner 1998). Reading is a highly structured and extensively practiced behavior, and involves high-level cognitive processes as well; any of these factors could account for the particularly asymmetric span it produces.
We conclude that it is variability in the spatial aspects of visual processing and saccade programming that drive performance in visual search, far more so than the temporal programming of saccades and fixations. It is easier for the visual system to extend attention spatially than it is to compress processing temporally. Subjects improved on our search task primarily by seeing more in a relatively fixed amount of time, rather than by processing visual information from a relatively fixed area more quickly.
Using a linear, familiar search array, we greatly simplified the process of saccade planning relative to typical 2- or 3-dimensional visual search. Such a simplified design facilitated the analysis of the relations between saccadic and perceptual task metrics during performance improvement and variability in a visuomotor task. While it is uncertain whether these results of one-dimensional search will extend to more complex search, it is notable that we found significant plasticity in saccadic perceptual span for stimuli in which variability in the stimulus array was kept to a minimum. In more complex, less predictable arrays, the analysis of information in the visual periphery becomes more important, as it is needed to decide where to make the next saccade. For these reasons, we predict more rather than less plasticity in saccadic perceptual span for more complex, less predictable arrays. This prediction has already been borne out for arrays of different difficulty levels, using non-homogenous distracters (Phillips & Edelman 2006).
One relevant question which remains unanswered by this study is why modulation of saccadic perceptual span is so relevant for performance whereas modulation of fixation duration, such as it is, is not. Our data are consistent with the following sketch of the timing of visual processing during search. First, within a fixation, the visual system analyzes the scene in roughly parallel fashion (Motter & Belky 1998a; Zenon, Ben Hamed, Duhamel, & Olivier 2005; Duncan, Ward & Shapiro 1994; Ward, Duncan & Shapiro 1996; Theeuwes, Godijn, & Pratt 2004), despite the fact that subjects’ time-per-item speeds (30–63 ms/item) fall in what has traditionally been considered the serial search range (Treisman & Gelade 1980). Second, the extent to which further exposure to the stimulus array increases target discriminability during a fixation declines substantially after a certain point, in the neighborhood of 200ms for our stimuli. Thus, our data suggest what can be grasped during a fixation is grasped either quickly or not at all, and that performance depends upon how much can be perceived in this small fraction of a second. The rate of saccadic eye movements during natural visuomotor behavior may strongly reflect these perceptual constraints.
Acknowledgments
NIGMS GM00816-26 (SCORE) and NIH/NCRR 5G12 Rr03060 (RCMI).
BIBLIOGRAPHY
- Ahissar M, Hochstein S. Task difficulty and the specificity of perceptual learning. Nature. 1997;387(6631):401–6. doi: 10.1038/387401a0. [DOI] [PubMed] [Google Scholar]
- Ball KK, Beard BL, Roenker DL, Miller RL, Griggs DS. Age and visual search: Expanding the useful field of view. Journal of the Optical Society of America A. 1988;5:2210–2219. doi: 10.1364/josaa.5.002210. [DOI] [PubMed] [Google Scholar]
- Bertera JH, Rayner K. Eye movements and the span of the effective stimulus in visual search. Perception & Psychophysics. 2000;62(3):576–85. doi: 10.3758/bf03212109. [DOI] [PubMed] [Google Scholar]
- Chung ST, Legge GE, Cheung SH. Letter-recognition and reading speed in peripheral vision benefit from perceptual learning. Vision Research. 2004;44(7):695–709. doi: 10.1016/j.visres.2003.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Courtney AJ, Chan HS. Visual lobe dimensions and search performance for targets on a competing homogneous background. Perception & Psychophysics. 2001;40(1):39–44. doi: 10.3758/bf03207592. [DOI] [PubMed] [Google Scholar]
- Comtois R. Vision Shell software library. Vision Sciences Laboratory, Harvard University; Cambridge MA: 2003. [Google Scholar]
- Deubel H, Schneider WX. Saccade target selection and object recognition: evidence for a common attentional mechanism. Vision Research. 1996;36(12):1827–37. doi: 10.1016/0042-6989(95)00294-4. [DOI] [PubMed] [Google Scholar]
- Duncan J, Ward R, Shapiro K. Direct measurement of attentional dwell time in human vision. Nature. 1994;369(6478):313–5. doi: 10.1038/369313a0. [DOI] [PubMed] [Google Scholar]
- Fahle M. Perceptual learning: a case for early selection. Journal of Vision. 2004;4(10):879–90. doi: 10.1167/4.10.4. [DOI] [PubMed] [Google Scholar]
- Fahle M. Learning to tell apples from oranges. Trends in Cognitive Science. 2005;9(10):455–7. doi: 10.1016/j.tics.2005.07.005. [DOI] [PubMed] [Google Scholar]
- Geisler WS, Chou KL. Separation of low-level and high-level factors in complex tasks: visual search. Psychological Review. 1995;102(2):356–378. doi: 10.1037/0033-295x.102.2.356. [DOI] [PubMed] [Google Scholar]
- Gersch TM, Kowler E, Dosher B. Dynamic allocation of visual attention during the execution of sequences of saccades. Vision Research. 2004;44(12):1469–83. doi: 10.1016/j.visres.2003.12.014. [DOI] [PubMed] [Google Scholar]
- Gordon IE. Eye movements during search through printed lists. Perceptual and Motor Skills. 1969;29(3):683–686. doi: 10.2466/pms.1969.29.3.683. [DOI] [PubMed] [Google Scholar]
- Gramopadhye AK, Drury CG, Jiang X, Sreenivasan R. Visual search and visual lobe size: can training one affect the other? International Journal of Industrial Ergonomics. 2002;30:181–195. [Google Scholar]
- Green CS, Bavelier D. Action video game modifies visual selective attention. Nature. 2003;423(6939):534–7. doi: 10.1038/nature01647. [DOI] [PubMed] [Google Scholar]
- Hoffman JE, Subramaniam B. The role of visual attention in saccadic eye movements. Perception & Psychophysics. 1995;57(6):787–95. doi: 10.3758/bf03206794. [DOI] [PubMed] [Google Scholar]
- Hooge IT, Erkelens CJ. Control of fixation duration in a simple search task. Perception & Psychophysics. 1996;58(7):969–76. doi: 10.3758/bf03206825. [DOI] [PubMed] [Google Scholar]
- Hooge ITC, Erkelens CJ. Peripheral vision and oculomotor control during visual search. Vision Research. 1999;39:1567–1575. doi: 10.1016/s0042-6989(98)00213-2. [DOI] [PubMed] [Google Scholar]
- Jacobs AM. Eye-movement control in visual search: how direct is visual span control? Perception & Psychophysics. 1986;39(1):47–58. doi: 10.3758/bf03207583. [DOI] [PubMed] [Google Scholar]
- Jacobs AM, O’Regan JK. Spatial and/or temporal adjustments of scanning behaviors to visibility changes. Acta Psychologica. 1987;65:133–146. doi: 10.1016/0001-6918(87)90023-0. [DOI] [PubMed] [Google Scholar]
- Kowler E, Anderson E, Dosher B, Blaser E. The role of attention in the programming of saccades. Vision Research. 1995;35(13):1897–916. doi: 10.1016/0042-6989(94)00279-u. [DOI] [PubMed] [Google Scholar]
- Leachtenauer JC. Peripheral acuity and photointerpretation performance. Human Factors. 1978;20:537–551. doi: 10.1177/001872087802000504. [DOI] [PubMed] [Google Scholar]
- McCarley JS, Kramer AF, Boot WR, Peterson MS, Wang RF, Irwin DE. Oculomotor behaviour in visual search for multiple targets. Visual Cognition. 2006;14:685–703. [Google Scholar]
- Motter BC, Belky EJ. The guidance of eye movements during active visual search. Vision Research. 1998a;38(12):1805–15. doi: 10.1016/s0042-6989(97)00349-0. [DOI] [PubMed] [Google Scholar]
- Motter BC, Belky EJ. The zone of focal attention during active visual search. Vision Research. 1998b;38(7):1007–22. doi: 10.1016/s0042-6989(97)00252-6. [DOI] [PubMed] [Google Scholar]
- Näsänen R, Ojanpää H, Kojo I. Effect of stimulus contrast on performance and eye movements in visual search. Vision Research. 2001;41(14):1817–24. doi: 10.1016/s0042-6989(01)00056-6. [DOI] [PubMed] [Google Scholar]
- Notman LA, Sowden PT, Ozgen E. The nature of learned categorical perception effects: a psychophysical approach. Cognition. 2005;95(2):B1–14. doi: 10.1016/j.cognition.2004.07.002. [DOI] [PubMed] [Google Scholar]
- Ojanpää H, Näsänen R, Kojo I. Eye movements in the visual search of word lists. Vision Research. 2002;42(12):1499–512. doi: 10.1016/s0042-6989(02)00077-9. [DOI] [PubMed] [Google Scholar]
- Pelz JB, Canosa R. Oculomotor behavior and perceptual strategies in complex tasks. Vision Research. 2001;41(25–26):3587–96. doi: 10.1016/s0042-6989(01)00245-0. [DOI] [PubMed] [Google Scholar]
- Petrov AA, Dosher BA, Lu ZL. The dynamics of perceptual learning: an incremental reweighting model. Psychological Review. 2005;112(4):715–43. doi: 10.1037/0033-295X.112.4.715. [DOI] [PubMed] [Google Scholar]
- Phillips MH, Edelman JA. Abstract Viewer/Itinerary Planner. Washington, DC: Society for Neuroscience; 2005. Performance and perceptual span in a constrained visual search task . Program No. 165.15. 2005. Online. [Google Scholar]
- Phillips MH, Edelman JA. Performance on a structured visual search task depends much more on perceptual span than fixation duration [Abstract] Journal of Vision. 2006;6(6):522, 522a . http://journalofvision.org/6/6/522/
- Rayner K. Eye movements in reading and information processing: 20 years of research. Psychological Bulletin. 1998;124(3):372–422. doi: 10.1037/0033-2909.124.3.372. [DOI] [PubMed] [Google Scholar]
- Rayner K, Fisher DL. Letter processing during eye fixations in visual search. Perception & Psychophysics. 1987;42(1):87–100. doi: 10.3758/bf03211517. [DOI] [PubMed] [Google Scholar]
- Sekuler R, Ball K. Visual localization: Age and practice. Journal of the Optical Society of America A. 1986;3:864–867. doi: 10.1364/josaa.3.000864. [DOI] [PubMed] [Google Scholar]
- Shen J, Reingold EM, Pomplun M, Williams DE. Saccadic selectivity during visual search: The influence of central processing difficulty. In: Hyona J, Radach R, Deubel H, editors. The mind’s eye: Cognitive and applied aspects of eye movement research. Amsterdam: Elsevier; 2003. pp. 65–88. [Google Scholar]
- Theeuwes J, Godijn R, Pratt J. A new estimation of the duration of attentional dwell time. Psychonomic Bulletin & Review. 2004;11(1):60–4. doi: 10.3758/bf03206461. [DOI] [PubMed] [Google Scholar]
- Treisman AM, Gelade G. A feature-integration theory of attention. Cognitive Psychology. 1980;12(1):97–136. doi: 10.1016/0010-0285(80)90005-5. [DOI] [PubMed] [Google Scholar]
- Ward R, Duncan J, Shapiro K. The Slow Time-Course of Visual Attention. Cognitive Psychology. 1996;30(1):79–109. doi: 10.1006/cogp.1996.0003. [DOI] [PubMed] [Google Scholar]
- Zelinsky G, Sheinberg D. Why some search tasks take longer than others: using eye movements to redefine reaction times. In: Findlay JM, Walker R, Kentridge RW, editors. Eye Movement Research: mechanisms, processes, and applications. North-Holland: 1995. pp. 325–336. [Google Scholar]
- Zelinsky GJ, Sheinberg DL. Eye movements during parallel-serial visual search. Journal of Experimental Psychology: Human Perception & Performance. 1997;23:244–262. doi: 10.1037//0096-1523.23.1.244. [DOI] [PubMed] [Google Scholar]
- Zenon A, Ben Hamed S, Duhamel J, Olivier E. 2005 Abstract Viewer/Itinerary Planner. Washington, DC: Society for Neuroscience; 2005. Association of focal attention and parallel filtering furing an inefficient visual search. Program No. 165.10. Online. [Google Scholar]