

Abstract
We evaluated the safety and efficacy of the mast cell activator compound 48/80 (C48/80) when used as an adjuvant delivered intradermally (ID) with recombinant anthrax protective antigen (rPA) in comparison with two well-known adjuvants. Mice were vaccinated in the ear pinnae with rPA or rPA + C48/80, CpG oligodeoxynucleotides (CpG), or cholera toxin (CT). All adjuvants induced similar increases in serum anti-rPA IgG and lethal toxin neutralizing antibodies. C48/80 induced a balanced cytokine production (Th1/Th2/Th17) by antigen-restimulated splenocytes, minimal injection site inflammation, and no antigen-specific IgE. Histological analysis demonstrated that vaccination with C48/80 reduced the number of resident mast cells and induced an injection-site neutrophil influx within 24 hours. Our data demonstrate that C48/80 is a safe and effective adjuvant, when used by the intradermal route, to induce protective antibody and balanced Th1/Th2/Th17 responses.
Keywords: Compound 48/80, adjuvant safety, intradermal vaccination
2. Introduction
Immunization programs have led to the eradication of smallpox, the near eradication of polio, and the control of other infectious diseases, including measles, mumps, rubella, and diphtheria. While there are many success stories, HIV, pandemic Influenza, and a variety of emerging infectious diseases (West Nile Virus, Dengue, etc.) provide active reminders that safe and effective vaccines continue to be needed to combat infectious diseases.
Adjuvants are substances that, when combined with vaccine antigens, enhance the induction of the desired immune responses [1, 2]. For example, the addition of adjuvants to vaccines may increase vaccine potency by enhancing the magnitude of antibody or cellular responses induced, reducing the time to seroprotection or selectively inducing CD4+ Th1, Th2, Th17, or CD8+ T cell responses [1, 2]. The mechanism of action of adjuvants varies depending on the adjuvant used, but the end result is thought to include the activation and migration of dendritic cells as well as the expression of antigen presenting molecules, providing superior induction of antigen-specific T and B cell responses [1, 2]. Despite increasing information on the mechanism of action of adjuvants, there are currently very few vaccine adjuvants licensed for human use. Aluminum-based adjuvants are currently the only vaccine adjuvants approved for use worldwide [3], although others, such as MPL (GlaxoSmithKline) and MF59 (Novartis) [4], have been approved for use in the European Union. New classes of adjuvants including cytokines, toll-like receptor (TLR) agonists, and compounds targeting specific cell populations are currently being explored [1-3].
We recently reported that the mast cell activator compound 48/80 (C48/80) was an effective adjuvant for the induction of anthrax lethal toxin neutralizing antibody responses when delivered intranasally or via the footpad to mice with anthrax protective antigen (PA; 5 μg or 0.5 μg, respectively) [5]. The adjuvant activity of C48/80 was associated with its ability to induce dendritic cell migration via a mechanism that required mast cells and mast cell-derived TNF but did not involve activation of TLR2,3,4,5,7,8,9 or require MyD88 [5]. The prevalence of mast cells in the dermis [6] suggests that the use of mast cell-activating adjuvants in intradermal vaccines could further increase the potency of this immunization route. Despite our demonstration that C48/80 provided effective adjuvant activity when delivered by the nasal route, we had no information regarding the adjuvant activity of C48/80 when delivered by the intradermal route.
Safety is an important issue that must be evaluated for any new adjuvant [1, 7]. For example, although Complete Freund's adjuvant (CFA) and Incomplete Freund's adjuvant (IFA) are potent adjuvants that have been used for many years in research settings, they often induce excessive injection site inflammation when used in humans and are therefore too toxic for routine human use [8-10]. Additionally, some adjuvants induce undesirable immune responses such as antigen-specific IgE that could sensitize the host to allergic or anaphylactic responses upon antigen challenge [11-16]. In the current study, the adjuvant activity and safety profile of C48/80 was evaluated when administered intradermally into the ear pinnae of mice with Bacillus anthracis protective antigen. CpG, a TLR9 ligand adjuvant known to induce Th1 type responses [17, 18] that has a history of use in humans [19-22] and cholera toxin (CT), a known Th2 adjuvant [23, 24] were used as control adjuvants.
3. Materials and Methods
3.1 Mice
Female C3H/HeN mice were obtained from the Charles River/National Cancer Institute. Mice were housed in filter top cages and provided food and water ad libitum. All procedures were approved by the Duke University Institutional Animal Care and Use Committee.
3.2 Vaccination
Mice were immunized i.d. in the dorsal side of the left ear pinnae with 10 μl of vaccine (diluted in PBS) delivered with a Gastight syringe using a 31-gauge needle (Hamilton Co., Reno, Nev.). Mice were anesthetized with ketamine-xylazine prior to immunization and ear tagged in the right ear following immunization. Mice were divided into groups of five. All mice, except naïve mice, received 0.5 μg of rPA (List Biologicals) as immunogen, either with or without adjuvant. Adjuvants included 3, 10, or 30 μg C48/80 (Sigma), 0.1 or 1.0 μg CT (List Biologicals), and 1 or 10 μg CpG DNA (CpG ODN 1826; Invivogen). CT and CpG doses were similar to those used intradermally by other groups [14, 25, 26]. Mice were immunized on days 0 and +21. Serum samples were collected on days +35 and +42.
3.3 Ear Swelling Assay
Ear thickness measurements were taken of the left ear immediately prior to and 24 hours post-vaccination with a dial thickness gauge (Mitutoyo, model no. 7326). The results are expressed as “vaccine-induced ear swelling” by subtracting the ear thickness prior to immunization from the ear thickness 24 hours post-immunization. Ear swelling is expressed in units of millimeters.
3.4 Sample Collection
Blood samples were collected from anesthetized mice by orbital sinus or maxillary venipuncture. Samples were collected into 1.5 ml centrifuge tubes, allowed to clot and centrifuged at 13,000 rpm at 4°C for 25 minutes in a Heraeus Biofuge fresco centrifuge. The serum was transferred to a new tube and stored at -20°C until tested.
3.5 Ex-vivo Restimulation of Spleen Cells
Mice were euthanized on day +42 using CO2 overdose, their spleens were immediately harvested, and a single cell suspension of spleen cells was prepared. Splenocyte restimulation was done as previously described [27] with the following exception: 2.5 × 106 cells per well were plated in 250 μl into 48-well plates. 250 μl of either T cell media or a solution of 2 μg/ml rPA in media (to yield a final concentration of 1 μg/ml) was then added to the cells. The plates were incubated at 37 °C for 60 hours. Supernatants were harvested to 96-well deep well plates and stored at -80 °C until analyzed.
3.6 Cytokine Profiles
Spleen cell restimulation cytokine profiles were determined using a multiplex bead assay from R&D (Minneapolis, MN). Analytes measured included IL-4, IL-5, IL-6, IL-17, and IFNγ. Samples with analyte concentrations that fell below the low standard were assigned a value equal to half the low standard for statistical analysis.
3.7 Lethal Toxin Neutralization Assay
This procedure was performed as outlined by Staats et al [16] with the following exceptions. Serum collected from mice on day +42 post-immunization was used to measure the titer of anthrax lethal toxin neutralizing antibodies in an anthrax macrophage toxicity assay. The amount of toxin used was 4-fold of the dose required for killing 100% of the cells. Serum samples were first diluted 1:64 in media and then serially diluted 1:2. rPA and LF were added at concentrations of 0.75 μg/ml and 0.375 μg/ml, respectively for a final concentration of 0.1875 μg/ml. Seventy-five percent neutralization titers (NT75) were calculated by plotting percent neutralization versus serum dilution and using linear regression to calculate the dilution at which 75% of the cells were viable. Samples with an NT75 less than 1:128 were below our tested range and were assigned a value of 1:2 for graphical representation and statistical evaluation.
3.8 Enzyme-linked Immunosorbent Assay
ELISAs were performed as outlined in Bradney et al. [28] and Nordone et al. [29] except that ELISA plates were coated with rPA at 2 μg/ml in CBC buffer. The log2 endpoint titers were used for statistical analysis.
3.9 IgE ELISA
ELISA plates were coated with 15 μl purified anti-mouse IgE (clone R35-72; BD Pharmingen Cat. # 02111D) at 5 μg/ml in CBC buffer. After overnight incubation, non-specific binding was blocked by adding 30 μl/well dry milk in CBC buffer and incubated for at least 2 hours. Plates were washed in ELISA wash buffer (PBS, 0.1% Kathon, 0.05% Tween20) and diluted samples (1:16) were plated in complete sample diluent (10% 10× PBS, 1% W/V bovine serum albumin, 1% W/V non-fat dry milk, 5% normal goat serum, 0.05% Tween20, 0.5% Kathon, dH20) for overnight incubation. Plates were washed and biotinylated rPA (15 μl/well) was added at 2 μg/ml diluted in secondary antibody diluent (% 10× PBS, 1% W/V bovine serum albumin, 5% normal goat serum, 0.05% Tween20, 0.5% Kathon, dH20) and incubated for at least 2 hours at room temperature. Plates were washed, streptavidin-AP diluted in secondary antibody diluent was added (15 μl/well), and plates were incubated for at least 2 hours. Plates were washed with ELISA wash buffer and 15 μl Attophos substrate (Promega) was added to each well and incubated for 15 minutes before reading at 440/560 nm.
3.10 Histology
Vaccinations were performed as described above. Mice were euthanized 4 and 24 hours after vaccination and ears were removed and fixed in 10% formalin prior to paraffin embedding. Sections were cut at a thickness of 5 μm and stained with hematoxylin and eosin (H&E) or Toluidine Blue O. Sections were evaluated by a pathologist blinded to the treatment groups. Mast cells were counted at 40× in at least 14 0.16mm2 fields per ear section. Degranulating mast cells were defined as having at least three granules evident outside the cell while hypogranulated mast cells were defined as having granules reduced in density throughout entire cell and nucleus clearly visible. The numbers of degranulating and hypogranulated mast cells were added to yield the number of activated mast cells. The percent of activated mast cells was determined by dividing the number of activated mast cells by the total number of mast cells counted in each ear section.
3.11 Statistics
Paired two sample T-tests were used to calculate significance between serum IgG1 and IgG2a (within groups), and media restimulated vs. rPA restimulated spleen cell cytokines (within groups). P values < 0.05 were considered significant. ANOVAs for multiple comparisons (Tukey) were performed in GraphPad Prism 5 for serum IgG and IgG subclasses, as well as ear swelling data and total mast cell numbers. Non-parametric ANOVAs (Kruskal-Wallis) with Dunn's Multiple Comparison Test were performed in GraphPad Prism 5 for serum IgE and spleen cell restimulation cytokines.
4. Results
4.1 Compound 48/80 is an effective adjuvant when administered by the intradermal route with anthrax protective antigen
To evaluate the adjuvant activity of C48/80 (a mast cell activator) when delivered by the intradermal route, female C3H/HeN mice were intradermally vaccinated with 0.5 μg rPA alone or combined with C48/80 (3, 10 or 30 μg) on days 0 and 21. Control adjuvants included CpG (1 or 10 μg) due to its Th1-polarizing activity [17, 18] and CT (0.1 or 1 μg) due to its well-known Th2-polarizing effects [23, 24]. Immunization with rPA alone induced moderate serum anti-rPA IgG titers by day +42 (Figure 1). The use of 3 μg C48/80 as an adjuvant did not provide significant adjuvant activity and was not pursued further. Increasing the dose of C48/80 to 10 or 30 μg augmented the induction of anti-rPA IgG antibodies and increased GMTs 53- and 177-fold (1:2,522,926 and 1:8,388,608, respectively) relative to rPA alone on day +42 (p < 0.001). The serum day +42 anti-rPA IgG titers induced by 30 μg C48/80 were not significantly different than those induced by CpG (1 or 10 μg; 1:11,068,835 and 1:67,108,864, respectively) or CT (0.1 or 1 μg; 1:25,429,504 and 1:25,429,504, respectively) (Figure 1). Therefore, C48/80 provided adjuvant activity comparable to that provided by CpG or CT when delivered intradermally.
Figure 1.
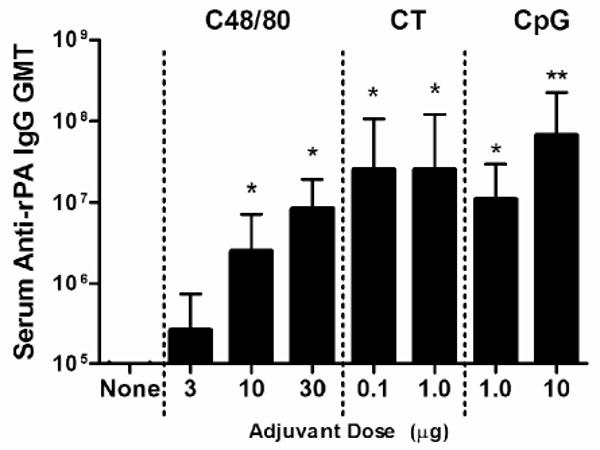
Serum anti-rPA IgG geometric mean titers after intradermal immunization in the ear pinna with 0.5 μg rPA with or without adjuvant on days 0 and +21. Serum samples taken on day +42 were tested by ELISA. Bars represent the geometric mean titers for each group on day +42 for all replicates, with error bars representing the 95% confidence level (CL). 0.1 μg CT, 1.0 μg CT, 1.0 μg CpG, and 10 μg CpG groups have an n = 5. 3.0 μg C48/80 group n = 10. rPA alone, 10 μg C48/80, and 30 μg C48/80 groups have an n = 15. * p < 0.05 over rPA alone and rPA plus 3 μg C48/80. ** p < 0.01 over rPA plus 10 μg C48/80.
Antigen-specific serum IgG subclasses reflect the subset of CD4+ T helper cells that are induced by vaccination, with IgG1 and IgG2a corresponding with Th2 and Th1 responses, respectively [30]. It is still unclear what effects Th17 CD4+ T cells have on B cell class switching and IgG subclass profiles [31]. To determine how C48/80 influenced the antigen-specific IgG subclass responses, day +42 serum samples were tested for rPA-specific IgG1, IgG2a, IgG2b, and IgG3. IgG1 titers were significantly higher than the IgG2a, IgG2b, and IgG3 titers in groups immunized with rPA alone (p < 0.001) or rPA plus C48/80 for all doses tested (p < 0.05). Mice vaccinated with rPA plus 1 μg or 10 μg CpG had anti-PA IgG2a titers that were greater than the anti-PA IgG1 titers, although these increases were not significant. Mice vaccinated with rPA plus 1 μg CT also had similar levels of IgG1 and IgG2a. IgG1 titers were 12-fold greater than IgG2a titers in mice vaccinated with rPA plus 0.1 μg CT (p = 0.0033) (Figure 2). These data demonstrate that C48/80 and CT induced Th2-biased IgG antibody responses while CpG induced a Th1-biased response.
Figure 2.

Serum anti-rPA IgG1, IgG2a, IgG2b, and IgG3 geometric mean titers after intradermal immunization in the ear pinna with 0.5 μg rPA in PBS with or without adjuvant on days 0 and +21. Bars represent the geometric mean titers for each group on day +42 for all replicates, with error bars representing the 95% CL. 1.0 μg CT has an n = 3. 0.1 μg CT, 1 μg CpG, and 10 μg CpG have an n = 5. 3.0 μg C48/80 group n = 10. rPA alone, 10 μg C48/80, and 30 μg C48/80 groups have an n = 15. Serum samples were tested by ELISA. 2 p < 0.05 over IgG2a; 3 p < 0.05 over IgG2b; *IgG3 was significantly lower than all other subclasses (p < 0.01).
IgG2b levels were also increased by vaccination. All adjuvant groups induced significantly higher titers of IgG2b than PA alone (1:213; p < 0.001). Vaccination with 10 or 30μg of C48/80 increased IgG2b titers to 1:7,061 and 1:217,904, respectively. Mice vaccinated with 0.1 or 1 μg CT induced IgG2b titers of 1:456,419 and 1:330,281, respectively. 10 μg CpG induced the greatest increased in IgG2b with a titer of 1:2,097,152, while 1 μg CpG induced a titer of 1:301,124. IgG3 levels were nearly nonexistent in the rPA alone group and very low in all mice vaccinated with C48/80, 0.1 μg CT, or 1 μg CpG as adjuvants (GMT < 1:41). Groups vaccinated with 1 μg CT and 10 μg CpG developed anti-rPA IgG3 GMT of 1:5,161 and 1:21,619, respectively, which were significantly increased over all other groups (p < 0.05 and p < 0.001, respectively).
4.2 Compound 48/80 augments the induction of lethal toxin neutralizing antibodies after intradermal immunization with anthrax protective antigen
Induction of antigen-specific IgG measured by ELISA does not always correlate with protective antibody responses [32, 33]. It has been shown, however, that anthrax lethal toxin (LeTx) neutralizing antibody responses correlate with survival [34, 35]. Therefore, we tested day +42 serum from mice immunized i.d. with rPA plus or minus adjuvant for its ability to neutralize LeTx using a macrophage protection assay. Of the twelve serum samples tested from mice vaccinated with 0.5 μg rPA alone, only one had a detectable level LeTx of neutralizing antibodies (Figure 3). C48/80 induced LeTx neutralizing antibodies in a dose-dependent fashion with 10 μg and 30 μg C48/80 augmenting significantly increased LeTx neutralizing antibody titers of 1:31 (p < 0.05) and 1:1,062 (p < 0.001), respectively. 0.1 μg and 1.0 μg CT induced LeTx neutralizing antibody responses of 1:624 and 1:1,886, respectively, while 1.0 μg and 10 μg CpG induced titers of 1:1,752 and 1:2,494, respectively. The adjuvant groups 30 μg C48/80, 0.1 μg CT, 1 μg CT, 1 μg CpG, and 10 μg CpG induced neutralizing antibody titers that were significantly greater than rPA alone and rPA plus 10 μg C48/80 (p < 0.001), but were not significantly different from each other. This data demonstrates that intradermal immunization with any of the three tested adjuvants induced significant increases in LeTx neutralizing antibodies over antigen alone that were of sufficient magnitudes to protect mice from LeTx challenge, as previous work has demonstrated that an NT50 of 1:1,250 is sufficient to protect animals from LeTx challenge [16, 36]. However, their ability to induce LeTx-neutralizing antibodies relative to ELISA-binding antibodies varied. Relative to the serum anti-PA IgG titers at day 42, 30 μg C48/80 induced the greatest proportion of neutralizing antibodies (i.e., higher NT75 with lower serum ELISA titer) of all adjuvants tested. Although C48/80 did not induce the highest serum IgG titer in our study, it had the greatest ability to induce functional antibodies, with an efficiency 1.7-fold greater than 1 μg CT and almost 3.5-fold greater than that of 10 μg CpG.
Figure 3.
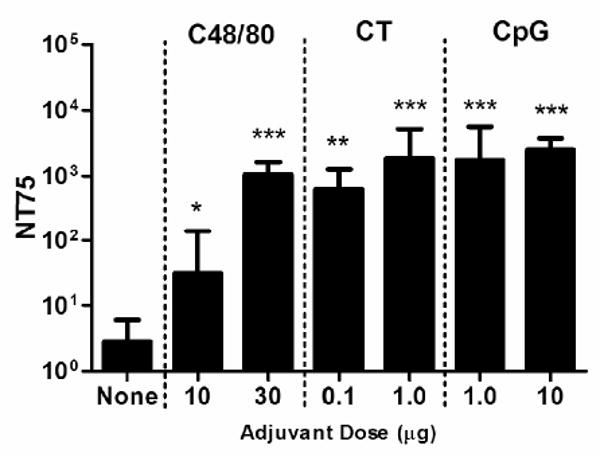
Serum lethal toxin neutralization titer 75 on day +42 after intradermal immunization in the ear pinna with 0.5 μg rPA with or without adjuvant on days 0 and +21. Bars represent the geometric mean titers for each group on day +42 for all replicates, with error bars representing the 95% CL. rPA alone n = 12. 10 μg C48/80 and 30 μg C48/80 groups have an n = 15. 0.1 μg CT, 1.0 μg CT, 1 μg CpG, and 10 μg CpG groups have an n = 5. Error bars represent 95% CL. Groups with neutralization titers below 1:128 were assigned a value of 2 for graphical representation and statistical analysis. * p < 0.05 over rPA alone. ** p < 0.05 over rPA alone and rPA plus 10 μg C48/80. *** p < 0.001 over rPA alone and rPA plus 10 μg C48/80.
4.3 Compound 48/80 does not induce PA-specific IgE
Adjuvant-induced, antigen-specific IgE responses are considered a safety hazard for vaccines. For example, given orally as an adjuvant, CT can induce the production of antigen-specific IgE resulting in anaphylaxis upon antigen challenge [37]. We therefore monitored rPA-specific IgE responses induced by intradermal vaccination with rPA alone or combined with adjuvant (Figure 4). C48/80 (10 or 30 μg) and CpG (1 or 10 μg) did not significantly induce the production of IgE as anti-PA IgE geometric mean titers were lower than the limit of sensitivity of our assay (1:16). Our positive control Th2 adjuvant, CT, (0.1 μg and 1 μg) induced anti-PA IgE titers that were significantly greater than all other groups (p < 0.01; 1:128 and 1:91, respectively). These results demonstrate that C48/80 provides effective adjuvant activity while not inducing potentially detrimental IgE.
Figure 4.

Serum anti-rPA IgE geometric mean titers after intradermal immunization in the ear pinna with 0.5 μg rPA in PBS with or without adjuvant on days 0 and +21. Lines represent the geometric mean titers for each group on day +42 for all replicates. Serum samples were tested by ELISA. The Y axis is set at the threshold of detection for this assay, 16. 0.1 and 1.0 μg CT groups have an n = 5 and n = 4, respectively. All mice vaccinated with CT had detectable IgE. rPA alone mice had no detectable IgE. 1 of 15 mice vaccinated with 10 μg C48/80 had detectable IgE, while 8 of 15 mice vaccinated with 30 μg C48/80 had detectable levels. 1.0 and 10 μg CpG induced IgE in 2 of 5 and 2 of 4 mice, respectively. ** p < 0.01 over rPA alone and rPA plus 10 μg C48/80.
4.4 Compound 48/80 induces splenocyte production of Th1, Th2, and Th17 cytokines when used as an adjuvant for intradermally administered vaccines
Adjuvants are useful tools for directing the immune response toward the desired CD4+ T helper cell response to combat different types of pathogens [38]. To evaluate the ability of the C48/80 to influence antigen-specific T cell responses induced by vaccination, spleens harvested at the end of each experiment were restimulated with rPA and supernatants were evaluated for the presence of Th1 (IFNγ), Th2 (IL-4 and IL-5), and Th17 (IL-6 and IL-17) cytokines (Table 1). Antigen restimulation of splenocytes from naïve mice or mice immunized with rPA alone did not result in significant cytokine production upon spleen cell restimulation when compared to media-treated splenocytes from naïve mice. Vaccination with the highest dose of each adjuvant tested (30 μg C48/80, 1 μg CT, or 10 μg CpG), resulted in the production of statistically equivalent amounts of IL-4, -6, -17, and IFNγ, which were significantly greater than the amounts produced by mice vaccinated with rPA alone (p < 0.01 and 0.001, respectively). 30 μg C48/80-induced IL-5 production was also significantly greater than 1.0 μg CpG-induced IL-5 production (p < 0.05). The highest concentrations of IL-4, IL-5, IL-6 and IL-17 in this study were produced by splenocytes from mice immunized with 1.0 μg CT as adjuvant. CT-induced IL-4 production was significantly greater than that induced by PA alone and all of the low-dose adjuvant groups (p < 0.05). Of the high-dose adjuvant groups, 30 μg C48/80 induced the lowest absolute production of the Th17-related cytokines, IL-6 and IL-17, but the levels were significantly increased over PA alone (p < 0.001 and 0.05, respectively) and were not significantly less than the levels induced by 1.0 μg CT or 10 μg CpG. As expected, vaccination with rPA plus 10 μg CpG produced the absolute highest concentration of IFNγ, 3.67 ng/ml IFNγ, nearly 2.5-fold greater than the level induced by 1 μg CT (1.5 ng/ml) and 3.1-fold greater than the level induced by 30 μg C48/80 (1.1 ng/ml), but these increases were not significant. Of the low adjuvant doses, 1.0 μg CpG was the most effective, inducing significant increases in all but IL-5 production over PA alone (p < 0.05). 0.1 μg CT induced a significant increase in IL-6 production (p < 0.05). These data indicate that C48/80 induces an antigen-specific cytokine response between that induced by a known Th1 adjuvant (CpG) and a known Th2 adjuvant (CT), when used as an adjuvant for intradermally-administered vaccines.
Table 1.
Antigen-specific cytokine production after 60 hr splenocytes restimulation with 1 μg rPA.
| Group | Treatment | Cytokine Production (pg/ml) | ||||
|---|---|---|---|---|---|---|
| IL-4 | IL-5 | IL-6 | IL-17 | IFNγ | ||
| a | Naïve | 0 ± 0 | 1 ± 2 | 2 ± 3 | 2 ± 6 | 11 ± 15 |
| b | PA alone | 0 ± 0 | 9 ± 17 | 2 ± 3 | 0 ± 1 | 26 ± 46 |
| c | PA + 10 μg C48/80 | 1 ± 2 | 59 ± 52 | 16 ± 14 | 6 ± 11 | 230 ± 226 |
| d | PA + 30 μg C48/80 | 10 ± 10a,b | 258 ± 149a,b,g | 52 ± 17a,b | 37 ± 29b | 1,184 ± 475a,b |
| e | PA + 0.1 μg CT | 3 ± 6 | 68 ± 69 | 64 ± 40b | 65 ± 72 | 878 ± 681 |
| f | PA + 1.0 μg CT | 51 ± 30a,b,c,e,g | 513 ± 592a,b | 213 ± 116a,b,c | 657 ± 443a,b,c | 1,503 ± 330a,b |
| g | PA + 1.0 μg CpG | 0 ± 0 | 12 ± 24 | 99 ± 20a,b | 73 ± 42a,b | 3,321 ± 624a,b,c |
| h | PA + 10 μg CpG | 13 ± 6a,b,c | 78 ± 36 | 112 ± 61a,b | 138 ± 116a,b,c | 3,686 ± 421a,b,c |
Superscript letters indicate treatment groups that are significantly different (p < 0.05).
4.5 Compound 48/80 induces minimal injection site swelling
Injection site reactions are a common adverse event observed after vaccination [39]. We therefore monitored vaccine-induced ear swelling 24 hours after vaccination as a measure of adverse events. Using the anti-rPA serum IgG titers and serum LeTx neutralization titers, we chose the dose of each adjuvant that induced the greatest serum IgG titer. Mice immunized with rPA alone had a vaccine-specific ear swelling thickness of 0.0046 ± 0.0097mm at 24 hours after immunization (Figure 5). All three adjuvant groups induced significant levels of ear swelling. The swelling induced by 30 μg C48/80 (0.0427 ± 0.018 mm) was significantly increased vs. mice immunized with rPA alone (p < 0.001). The amount of swelling induced by 30 μg C48/80 was similar to that induced by 10 μg CpG (0.0406 ± 0.024 mm), which was also significantly greater than rPA alone (p < 0.01). However, 1.0 μg CT induced over twice as much ear swelling (0.0955 ± 0.025 mm) as the other groups tested (rPA alone, 30 μg C48/80, and 10 μg CpG; p < 0.001). Our results demonstrate that C48/80 does not induce excessive injection site swelling when used as an adjuvant for intradermally-administered vaccines.
Figure 5.
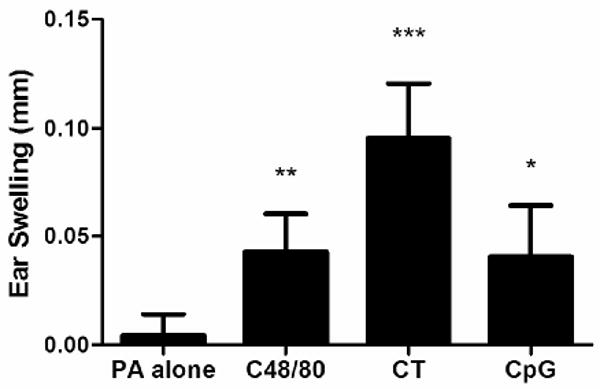
Ear swelling in mm 24 hours after intradermal immunization in the ear pinna with 0.5 μg rPA in PBS with or without adjuvant on day 0. Bars represent the mean swelling for each group for all replicates. 1.0 μg CT and 10 μg CpG groups have an n = 5. rPA alone and 30 μg C48/80 groups have an n = 15. Measurements were made using a dial thickness gauge. Error bars represent standard deviation. * p < 0.01 over rPA alone. ** p < 0.001 over rPA alone. *** p < 0.001 over all other groups.
4.6 Compound 48/80 induced cellular influx into the injection site
It was also of interest to assess cellular infiltrates at the injection site at 24 hours as an additional indicator of injection site inflammation. Mice were vaccinated ID in the ear pinnae at time 0 and ears were harvested 4 and 24 hours later. Ear sections taken from mice immunized with PA alone showed little inflammation (Figures 6B/G) compared to unvaccinated ears (Figures 6A/F). Vaccination with PA plus 30 μg C48/80 induced edema by 4 hours that persisted through 24 hours, at which point cellular infiltrate was present in all sections examined. The cellular infiltrate contained primarily neutrophils (Figure 6H). Edema was also present in ears from mice vaccinated with rPA plus 10 μg CpG at both 4 and 24 hours (Figures 6D/I) with both mononuclear cells and neutrophils present at 24 hours (Figure 6I). Ears taken from mice vaccinated with rPA plus 1 μg CT showed less edema than that induced by the other two adjuvants and minimal cellular infiltrate (Figures 6E/J). Separation of the ear tissue was visible in all ear sections taken from mice vaccinated with CT (Figure 6E), a histologic indicator of the massive swelling noted grossly. Sections were also stained with Toluidine Blue to highlight the mast cells in the ear tissue. Fully granulated mast cells are evident in sections from naïve mice or mice immunized with PA alone (Figures 6K and 6L). However, mast cells could not be identified in most of the ear tissue examined from mice vaccinated with C48/80 (Figure 6M). The total number of mast cells in the tissue at 4 hours after vaccination decreased from 49 ± 12 in ears vaccinated with PA alone to 20 ± 11 MC/mm2 in ears vaccinated with PA + C48/80 (p < 0.05). Mast cells were almost totally absent in the regions with maximal edema and infiltrate (Figure 7a). A similar picture was seen at 24 hours after vaccination, with 51 ± 12 MC/mm2 in ears vaccinated with PA alone and 16 ± 4 MC/mm2 in ears vaccinated with PA + C48/80 (p < 0.01) (Figure 7b). The total number of mast cells present in the ears of mice vaccinated with either PA + CT or PA + CpG did not differ from the number present in the ears of mice vaccinated with PA alone (Figure 7). Although the percentage of activated mast cells (defined as degranulating + hypogranulated mast cells) did not differ between vaccination groups (data not shown), the decrease in mast cell number induced by C48/80 was likely due to their activation. The histological examination therefore suggests that although C48/80 and CpG are inducing measurable ear swelling, they are encouraging the migration of neutrophils or neutrophils and monocytes to the site of injection, respectively, and that activation of mast cells by C48/80 results in their absence from the sections. Although this analysis did not determine what immune parameter induced the ear swelling, we are confident that an innate immune mechanism is responsible, since ear swelling measurements were taken just 24 hours after the initial vaccination, which is not sufficient time to induce antigen-specific immune responses such as antibody production or a cellular immune response.
Figure 6.
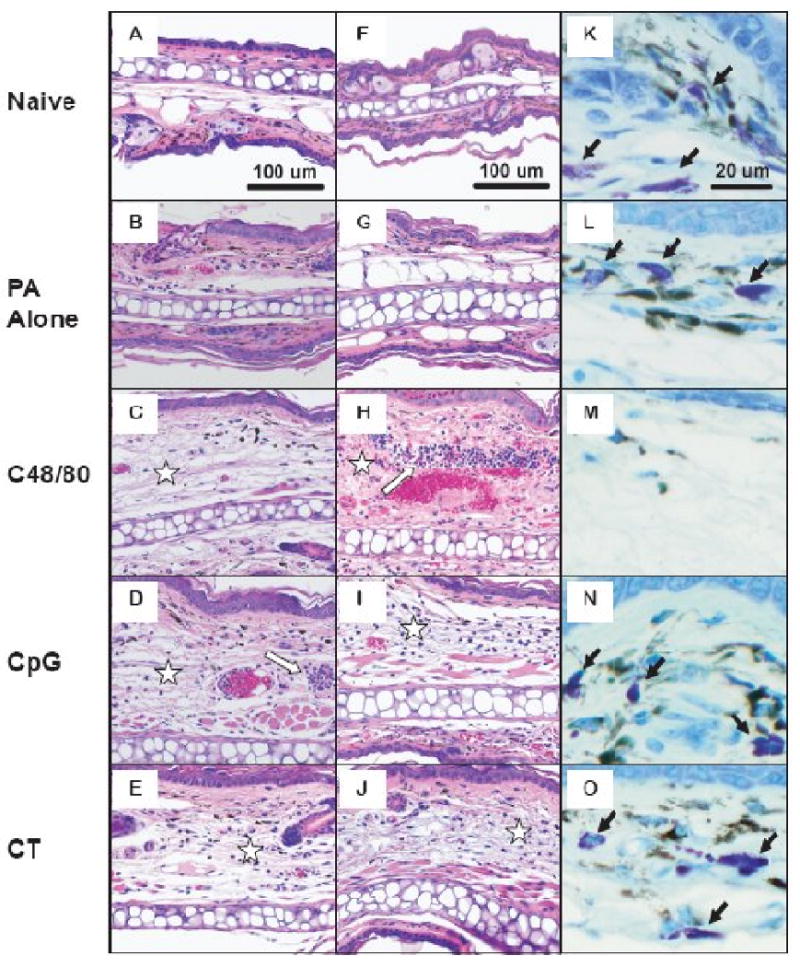
Histological changes in the ear pinna following intradermal vaccination. Mice were immunized with 0.5 mg rPA in PBS with or without the indicated adjuvant. Hematoxylin and eosin- stained sections are shown 4 hours (A-E) or 24 hrs (F-J) after immunization. Stars indicate areas of prominent edema and white arrows indicate inflammatory infiltrates. Toluidine Blue-stained sections (K-O) show that mast cells (black arrows) are readily detected following immunization with PA alone or with CT or CpG adjuvants, but not following immunization with c48/80.
Figure 7.
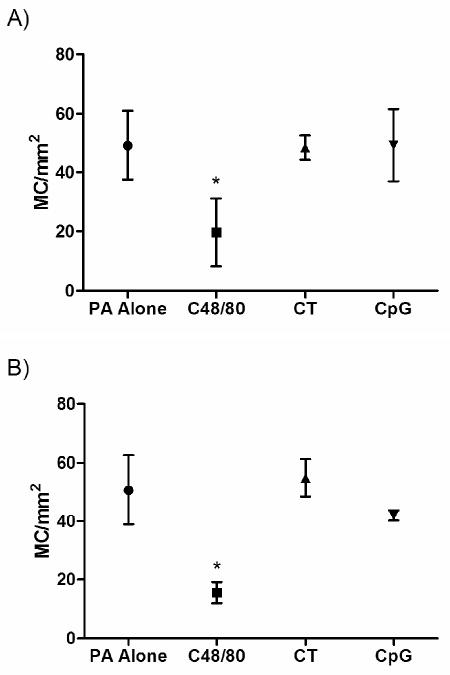
Total number of mast cells present per mm2 in ear sections taken at 4 (a) or 24 (b) hours after intradermal vaccination in the ear pinnae. Sections were stained with Toluidine Blue and mast cells were counted in a minimum of 14 (0.16mm2) fields at 40×. Error bars represent standard deviation. * p < 0.05 against all other groups.
5. Discussion
In this study, we have demonstrated that C48/80 provides adjuvant activity when co-administered with Bacillus anthracis protective antigen by the intradermal route. Vaccination using C48/80 as an adjuvant was associated with increases in serum IgG, serum lethal toxin neutralizing antibodies, and antigen-specific Th1/Th2/Th17 responses. Unlike CT, C48/80 did not induce antigen-specific IgE when used as an adjuvant. In addition, C48/80 encouraged the migration of inflammatory cells into the ear pinna while inducing less injection site inflammation than CT. C48/80 was the only adjuvant to activate mast cells.
This study is the first to demonstrate that an intradermally administered mast cell activator is able to provide vaccine adjuvant activity and confirms our recent observation that mast cell activators provide effective adjuvant activity [5]. Others have previously documented that mast cells are able to regulate adaptive immune responses in the skin. For example, Mazzoni et al demonstrated that the application of some mast cell activators in addition to vaccination with OVA and LPS suppressed the ability of DCs to induce Th1 CD4+ T cells [40]. Although their study involved the use of a second adjuvant (LPS) and did not focus on the induction of humoral immunity, they did demonstrate the ability of C48/80 to influence the development of T cell responses, much like we saw in our study. However, we noted an increase in both Th1 and Th2 cytokines. Although their study did not focus on the ability of mast cell activators to provide adjuvant activity or induce humoral immunity, it did demonstrate the important role that mast cells play in shaping the adaptive immune response. Our study builds on this study and others to demonstrate the potency of mast cell activators as vaccine adjuvants when delivered intradermally.
Our results confirm the observations of others [41-47] that intradermal immunization is an effective route of immunization that requires reduced antigen doses for the induction of protective immunization. Others have utilized rPA as an immunogen by the intramuscular [34, 48] or nasal [49, 50] routes and used a total of 30-60 or 15-120 μg of antigen, respectively, with three immunizations. Although all four studies induced LeTx neutralizing antibodies, the intradermal route used in our study required 15-120-fold less antigen. After just two immunizations, totaling 1 μg of antigen, we induced LeTx neutralizing antibody titers (NT50) in excess of 1:1,900. In contrast, Boyaka et al [49] and Matyas et al [48] both induced LeTx neutralizing titers of ∼1:1,000, but they required two nasal immunizations totaling 20 μg of antigen and three intramuscular immunizations for a total of 30 μg of rPA, respectively. By comparison, our results demonstrate the ability of intradermal immunization to induce potent immune responses using reduced antigen doses.
Adjuvants are useful tools for directing the immune response toward the desired CD4+ T helper cell response to combat different types of pathogens [38]. With regard to our study, vaccine-induced Th2 responses are likely to be the most effective in combating B. anthracis as antibody-mediated immune responses have been shown by others to provide protection against anthrax lethal toxin challenge [16, 51] and B. anthracis spore challenge [52, 53]. C48/80 was second only to CT in IL-5 production, yet it produced much greater levels of IFNγ than it did IL-4, -5, -6, or -17 and reduced the IgG1/IgG2a ratio to 1.1. Although mast cells are generally considered to play a role in the induction of Th2 effector immune responses [54], connective tissue mast cells, such as those found in the mouse ear pinna, have the ability to produce the Th1 cytokine IFNγ [54]. It is therefore possible that C48/80 is acting through the connective tissue mast cells to stimulate an environment favorable for the development of Th1 immune responses. Of the three adjuvants tested, C48/80 induced the most balanced cytokine profile.
Although adjuvants are useful for increasing and directing the immune response to vaccine antigens, several well-known adjuvants have been shown to induce strong inflammatory reactions. For instance, complete Freund's adjuvant (CFA) induces fibrosis at the site of injection [55] and incomplete Freund's adjuvant has been associated with injection site inflammation when used in humans [56, 57]. In addition to inflammatory reactions, the induction of antigen-specific IgE is also an important consideration in vaccine safety, as it has the potential to induce anaphylactic reactions [37, 58, 59]. To monitor these potential side-effects, we measured adjuvant-induced injection site inflammation (i.e., ear swelling) and antigen-specific serum IgE. In addition to its role as a strong Th2 adjuvant, CT is known to be a potent inducer of vaccine antigen-specific IgE when given as a mucosal or intradermal adjuvant, causing such serious reactions as anaphylaxis upon antigen challenge [23, 24, 37, 60]. As expected, CT induced the greatest amounts of both injection site swelling and antigen-specific IgE. In agreement with our previous study [5], vaccination with C48/80 did not induce antigen-specific IgE, and in fact, it induced similar injection site swelling as 10 μg CpG, which has been safely used in several clinical trials [9, 19-22]. Although mast cells are thought to be involved in the IgE-mediated allergic response [8], it has been shown that IgE-mediated anaphylaxis also occurs in mast cell-deficient mice [61], demonstrating that mast cells are not required for the induction of antigen-specific IgE. Our results confirm our previous observations that the mast cell activator C48/80 does not induce antigen-specific IgE.
Histological evaluation was necessary to more closely examine the injection site reactions. The large neutrophil influxes seen when mice were vaccinated i.d. with rPA plus C48/80 were likely due to the mast cell activating capacity of C48/80, as several studies have demonstrated the ability of mast cells to rapidly recruit neutrophils [11, 62, 63]. In a study by Malayvia et al, it was demonstrated that neutrophils migrated to the bladder of mice after E. coli challenge in a mast cell-dependent fashion [64]. The reduction in the number of mast cells observed in ear tissues is presumably due to C48/80-induced mast cell degranulation. Until recently, neutrophils were considered to only play a role in the innate immune response, where they acted as important mediators of several processes, including bacterial clearance in infection [62, 65]. However, it is becoming increasingly apparent that neutrophils do play a role in inducing the adaptive immune response [66, 67]. Two groups have demonstrated that neutrophil depletion alters the balance of Th1/Th2 cytokines in response to infection in favor of Th2 [68, 69]. It has also been shown that neutrophils can interact with and modulate the maturation of dendritic cells [70, 71], as well as migrate to lymph nodes [72] and prime naïve T cells in vivo [67]. Such evidence suggests that recruiting neutrophils to the site of vaccination may be beneficial for driving immune responses.
C48/80 has safely been used in human subjects, and our data are consistent with the few studies that have applied C48/80 cutaneously or intradermally to humans for allergy studies, in which it did not induce any serious long term side effects [73-77]. In one of these studies, researchers intradermally injected 700 μg of C48/80 into human subjects and no significant side effects were noted beyond the traditional wheal response [77]. This demonstrates that judicious application of MC activators appears to have no significant adverse effects and may provide a novel class of compounds for use as vaccine adjuvants.
In summary, our findings demonstrate that the mast cell activator C48/80 is a safe and potent adjuvant for intradermally delivered anthrax protective antigen. C48/80 induced enhanced antibody- and cell-mediated adaptive immune responses and lethal toxin neutralizing antibodies while inducing undetectable antigen-specific IgE and resulting in minimal injection site inflammation. Our results suggest that mast cell activators represent a new class of adjuvants that may be safely administered with intradermally administered vaccines.
Acknowledgments
The following reagents were obtained through the NIH Biodefense and Emerging Infections Research Resources Repository, NIAID, NIH: Anthrax Protective Antigen (PA), Recombinant from Bacillus anthracis, NR-140 and Anthrax Lethal Factor (LF), Recombinant from Bacillus anthracis, NR-570. Supported by NIH/NIAID R21 AI-059591.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Guy B. The perfect mix: recent progress in adjuvant research. Nat Rev Micro. 2007;5(7):505. doi: 10.1038/nrmicro1681. [DOI] [PubMed] [Google Scholar]
- 2.McKee AS, Munks MW, Marrack P. How Do Adjuvants Work? Important Considerations for New Generation Adjuvants. 2007;27(5):687–90. doi: 10.1016/j.immuni.2007.11.003. [DOI] [PubMed] [Google Scholar]
- 3.WHO. Proceedings of the third global vaccine research forum; Geneva: WHO; 2002. [Google Scholar]
- 4.Sesardic D, Dobbelaer R. European union regulatory developments for new vaccine adjuvants and delivery systems. Vaccine. 2004;22(19):2452. doi: 10.1016/j.vaccine.2003.11.071. [DOI] [PubMed] [Google Scholar]
- 5.McLachlan JB, Shelburne CP, Hart JP, Pizzo SV, Goyal R, Brooking-Dixon R, et al. Mast cell activators: a new class of highly effective vaccine adjuvants. Nat Med. 2008;14(5):536–41. doi: 10.1038/nm1757. [DOI] [PubMed] [Google Scholar]
- 6.Gersch C, Dewald O, Zoerlein M, Michael L, Entman M, Frangogiannis N. Mast cells and macrophages in normal C57/BL/6 mice. Histochemistry and Cell Biology. 2002;118(1):41–9. doi: 10.1007/s00418-002-0425-z. [DOI] [PubMed] [Google Scholar]
- 7.O'Hagan DT, Valiante NM. Recent advances in the discovery and delivery of vaccine adjuvants. Nat Rev Drug Discov. 2003;2(9):727–35. doi: 10.1038/nrd1176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Harissios Vliagoftis ADB. Rapidly changing perspectives about mast cells at mucosal surfaces. 2005:190–203. doi: 10.1111/j.0105-2896.2005.00279.x. [DOI] [PubMed] [Google Scholar]
- 9.Speiser DE, Liénard D, Rufer N, Rubio-Godoy V, Rimoldi D, Lejeune F, et al. Rapid and strong human CD8+ T cell responses to vaccination with peptide, IFA, and CpG oligodeoxynucleotide 7909. Journal of Clinical Investigation. 2005;115:739–46. doi: 10.1172/JCI23373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Choi IH, Shin YM, Park JS, Lee MS, Han EH, Chai OH, et al. Immunoglobulin E-dependent Active Fatal Anaphylaxis in Mast Cell-deficient. Mice. 1998:1587–92. doi: 10.1084/jem.188.9.1587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Malaviya R, Ikeda T, Ross E, Abraham SN. Mast cell modulation of neutrophil influx and bacterial clearance at sites of infection through TNF-[alpha] Nature. 1996;381(6577):77–80. doi: 10.1038/381077a0. [DOI] [PubMed] [Google Scholar]
- 12.Peachman KK, Rao M, Alving CR, Burge R, Leppla SH, Rao VB, et al. Correlation between Lethal Toxin-Neutralizing Antibody Titers and Protection from Intranasal Challenge with Bacillus anthracis Ames Strain Spores in Mice after Transcutaneous Immunization with Recombinant Anthrax. 2006:794–7. doi: 10.1128/IAI.74.1.794-797.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sawamura D, Abe R, Goto M, Akiyama M, Hemmi H, Akira S, et al. Direct injection of plasmid DNA into the skin induces dermatitis by activation of monocytes through toll-like receptor. 2005;9:664–71. doi: 10.1002/jgm.709. [DOI] [PubMed] [Google Scholar]
- 14.Wack A, Baudner BC, Hilbert AK, Manini I, Nuti S, Tavarini S, et al. Combination adjuvants for the induction of potent, long-lasting antibody and T-cell responses to influenza vaccine in mice. Vaccine. 2008;26(4):552–61. doi: 10.1016/j.vaccine.2007.11.054. [DOI] [PubMed] [Google Scholar]
- 15.Yamada H, Ishii K, Klinman D. Suppressive oligodeoxynucleotides inhibit CpG-induced inflammation of the mouse lung. Crit Care Med. 2004;32(10):2045–9. doi: 10.1097/01.ccm.0000142397.38134.ef. [DOI] [PubMed] [Google Scholar]
- 16.Staats HF, Alam SM, Scearce RM, Kirwan SM, Zhang JX, Gwinn WM, et al. In Vitro and In Vivo Characterization of Anthrax Anti-Protective Antigen and Anti-Lethal Factor Monoclonal Antibodies after Passive Transfer in a Mouse Lethal Toxin Challenge Model To Define Correlates of Immunity. 2007:5443–52. doi: 10.1128/IAI.00529-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tengvall S, Josefsson A, Holmgren J, Harandi AM. CpG oligodeoxynucleotide augments HSV-2 glycoprotein D DNA vaccine efficacy to generate T helper 1 response and subsequent protection against primary genital herpes infection in mice. Journal of Reproductive Immunology. 2005;68(12):53. doi: 10.1016/j.jri.2005.06.010. [DOI] [PubMed] [Google Scholar]
- 18.Ling Liu, X Z, H L, L X, Z Y. CpG motif acts as a ‘danger signal’ and provides a T helper type 1-biased microenvironment for DNA vaccination. 2005:223–30. doi: 10.1111/j.1365-2567.2005.02150.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Halperin SA, Van Nest G, Smith B, Abtahi S, Whiley H, Eiden JJ. A phase I study of the safety and immunogenicity of recombinant hepatitis B surface antigen co-administered with an immunostimulatory phosphorothioate oligonucleotide adjuvant. Vaccine. 2003;21(1920):2461–7. doi: 10.1016/s0264-410x(03)00045-8. [DOI] [PubMed] [Google Scholar]
- 20.Cooper CL, Davis HL, Morris ML, Efler SM, Krieg AM, Li Y, et al. Safety and immunogenicity of CPG 7909 injection as an adjuvant to Fluarix influenza vaccine. Vaccine. 2004;22(2324):3136–43. doi: 10.1016/j.vaccine.2004.01.058. [DOI] [PubMed] [Google Scholar]
- 21.Cooper CL, Davis HL, Angel JB, Morris ML, Elfer SM, Seguin I, et al. CpG 7909 adjuvant improves hepatitis B virus vaccine seroprotection in antiretroviral-treated HIV-infected adults. AIDS. 2005;19:1473–9. doi: 10.1097/01.aids.0000183514.37513.d2. [DOI] [PubMed] [Google Scholar]
- 22.Cooper CL, Davis HL, Morris ML, Efler SM, Adhami MA, Krieg AM, et al. CPG 7909, an Immunostimulatory TLR9 Agonist Oligodeoxynucleotide, as Adjuvant to Engerix-B® HBV Vaccine in Healthy Adults: A Double-Blind Phase I/II Study. Journal of Clinical Immunology. 2004;24(6):693–701. doi: 10.1007/s10875-004-6244-3. [DOI] [PubMed] [Google Scholar]
- 23.Bo Su S, Silver PB, Wang P, Chan CC, Caspi RR. Cholera Toxin Prevents Th1-Mediated Autoimmune Disease by Inducing Immune Deviation. J Immunol. 2004 July 15;173(2):755–61. doi: 10.4049/jimmunol.173.2.755. [DOI] [PubMed] [Google Scholar]
- 24.Marinaro M, Staats HF, Hiroi T, Jackson RJ, Coste M, Boyaka PN, et al. Mucosal adjuvant effect of cholera toxin in mice results from induction of T helper 2 (Th2) cells and IL-4. J Immunol. 1995 November 15;155(10):4621–9. [PubMed] [Google Scholar]
- 25.Akhiani AA, Nilsson LA, Ouchterlony O. Effect of cholera toxin on vaccine-induced immunity and infection in murine schistosomiasis mansoni. 1993:4919–24. doi: 10.1128/iai.61.11.4919-4924.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Uddowla S, Freytag LC, Clements JD. Effect of adjuvants and route of immunizations on the immune response to recombinant plague antigens. Vaccine. 2007;25(47):7984–93. doi: 10.1016/j.vaccine.2007.09.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Staats HF, Ennis FA., Jr IL-1 Is an Effective Adjuvant for Mucosal and Systemic Immune Responses When Coadministered with Protein Immunogens. J Immunol. 1999 May 15;162(10):6141–7. [PubMed] [Google Scholar]
- 28.Bradney CP, Sempowski GD, Liao HX, Haynes BF, Staats HF. Cytokines as Adjuvants for the Induction of Anti-Human Immunodeficiency Virus Peptide Immunoglobulin G (IgG) and IgA Antibodies in Serum and Mucosal Secretions after Nasal Immunization. J Virol. 2002 January 15;76(2):517–24. doi: 10.1128/JVI.76.2.517-524.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Nordone SK, Peacock JW, Kirwan SM, Staats HF. Capric Acid and Hydroxypropylmethylcellulose Increase the Immunogenicity of Nasally Administered Peptide Vaccines. AIDS Research and Human Retroviruses. 2006;22(6):558–68. doi: 10.1089/aid.2006.22.558. [DOI] [PubMed] [Google Scholar]
- 30.McKee AS, Munks MW, Marrack P. How Do Adjuvants Work? Important Considerations for New Generation Adjuvants. Immunity. 2007;27(5):687. doi: 10.1016/j.immuni.2007.11.003. [DOI] [PubMed] [Google Scholar]
- 31.Takagi R, Higashi T, Hashimoto K, Nakano K, Mizuno Y, Okazaki Y, et al. B Cell Chemoattractant CXCL13 Is Preferentially Expressed by Human Th17 Cell Clones. 2008:186–9. doi: 10.4049/jimmunol.181.1.186. [DOI] [PubMed] [Google Scholar]
- 32.Duc LH, Hong HA, Atkins HS, Flick-Smith HC, Durrani Z, Rijpkema S, et al. Immunization against anthrax using Bacillus subtilis spores expressing the anthrax protective antigen. Vaccine. 2007;25(2):346–55. doi: 10.1016/j.vaccine.2006.07.037. [DOI] [PubMed] [Google Scholar]
- 33.Berthold I, Pombo ML, Wagner L, Arciniega JL. Immunogenicity in mice of anthrax recombinant protective antigen in the presence of aluminum adjuvants. Vaccine. 2005;23(16):1993–9. doi: 10.1016/j.vaccine.2004.10.014. [DOI] [PubMed] [Google Scholar]
- 34.Peachman KK, Rao M, Alving CR, Burge R, Leppla SH, Rao VB, et al. Correlation between Lethal Toxin-Neutralizing Antibody Titers and Protection from Intranasal Challenge with Bacillus anthracis Ames Strain Spores in Mice after Transcutaneous Immunization with Recombinant Anthrax Protective Antigen. Infect Immun. 2006 January 1;74(1):794–7. doi: 10.1128/IAI.74.1.794-797.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Staats HF, Alam SM, Scearce RM, Kirwan SM, Zhang JX, Gwinn WM, et al. In Vitro and In Vivo Characterization of Anthrax Anti-Protective Antigen and Anti-Lethal Factor Monoclonal Antibodies after Passive Transfer in a Mouse Lethal Toxin Challenge Model To Define Correlates of Immunity. Infect Immun. 2007 November 1;75(11):5443–52. doi: 10.1128/IAI.00529-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zeng M, Xu Q, Pichichero ME. Protection against anthrax by needle-free mucosal immunization with human anthrax vaccine. Vaccine. 2007;25(18):3588–94. doi: 10.1016/j.vaccine.2007.01.075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Snider DP, Marshall JS, Perdue MH, Liang H. Production of IgE antibody and allergic sensitization of intestinal and peripheral tissues after oral immunization with protein Ag and cholera toxin. J Immunol. 1994 July 15;153(2):647–57. [PubMed] [Google Scholar]
- 38.Korn T, Oukka M, Kuchroo V, Bettelli E. Th17 cells: Effector T cells with inflammatory properties. Seminars in Immunology. 2007;19(6):362–71. doi: 10.1016/j.smim.2007.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kimmel SR. Vaccine adverse events: separating myth from reality. American Family Physician. 2002;66(1):2113–20. [PubMed] [Google Scholar]
- 40.Mazzoni A, Siraganian RP, Leifer CA, Segal DM. Dendritic Cell Modulation by Mast Cells Controls the Th1/Th2 Balance in Responding T Cells. J Immunol. 2006 September 15;177(6):3577–81. doi: 10.4049/jimmunol.177.6.3577. [DOI] [PubMed] [Google Scholar]
- 41.Lodmell DL, Ewalt LC, Parnell MJ, Rupprecht CE, Hanlon CA. One-time intradermal DNA vaccination in ear pinnae one year prior to infection protects dogs against rabies virus. Vaccine. 2006 Jan 23;24(4):412–6. doi: 10.1016/j.vaccine.2005.08.003. [DOI] [PubMed] [Google Scholar]
- 42.Belshe RB, Newman FK, Cannon J, Duane C, Treanor J, Van Hoecke C, et al. Serum Antibody Responses after Intradermal Vaccination against Influenza. N Engl J Med. 2004 November 25;351(22):2286–94. doi: 10.1056/NEJMoa043555. [DOI] [PubMed] [Google Scholar]
- 43.Kenney RT, Frech SA, Muenz LR, Villar CP, Glenn GM. Dose Sparing with Intradermal Injection of Influenza Vaccine. N Engl J Med. 2004 November 25;351(22):2295–301. doi: 10.1056/NEJMoa043540. [DOI] [PubMed] [Google Scholar]
- 44.La Montagne JR, Fauci AS. Intradermal Influenza Vaccination -- Can Less Be More? N Engl J Med. 2004 November 25;351(22):2330–2. doi: 10.1056/NEJMe048314. [DOI] [PubMed] [Google Scholar]
- 45.Roukens AH, Vossen AC, Bredenbeek PJ, van Dissel JT, Visser LG. Intradermally Administered Yellow Fever Vaccine at Reduced Dose Induces a Protective Immune Response: A Randomized Controlled Non-Inferiority Trial. PLoS ONE. 2008;3(4):e1993. doi: 10.1371/journal.pone.0001993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Sugimura T, Ito Y, Tananari Y, Ozaki Y, Maeno Y, Yamaoka T, et al. Improved antibody responses in infants less than 1 year old using intradermal influenza vaccination. Vaccine. 2008;26(22):2700. doi: 10.1016/j.vaccine.2008.03.016. [DOI] [PubMed] [Google Scholar]
- 47.Auewarakul P, Kositanont U, Sornsathapornkul P, Tothong P, Kanyok R, Thongcharoen P. Antibody responses after dose-sparing intradermal influenza vaccination. Vaccine. 2007;25(4):659. doi: 10.1016/j.vaccine.2006.08.026. [DOI] [PubMed] [Google Scholar]
- 48.Matyas GR, Friedlander AM, Glenn GM, Little S, Yu J, Alving CR. Needle-Free Skin Patch Vaccination Method for Anthrax. Infect Immun. 2004 February 1;72(2):1181–3. doi: 10.1128/IAI.72.2.1181-1183.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Boyaka PN, Tafaro A, Fischer R, Leppla SH, Fujihashi K, McGhee JR. Effective Mucosal Immunity to Anthrax: Neutralizing Antibodies and Th Cell Responses Following Nasal Immunization with Protective Antigen. J Immunol. 2003 June 1;170(11):5636–43. doi: 10.4049/jimmunol.170.11.5636. [DOI] [PubMed] [Google Scholar]
- 50.Sloat B, Cui Z. Nasal Immunization with Anthrax Protective Antigen Protein Adjuvanted with Polyriboinosinic–Polyribocytidylic Acid Induced Strong Mucosal and Systemic Immunities. Pharmaceutical Research. 2006;23(6):1217. doi: 10.1007/s11095-006-0206-9. [DOI] [PubMed] [Google Scholar]
- 51.Cui X, Li Y, Moayeri M, Choi Gil H, Subramanian GÂM, Li X, et al. Late Treatment with a Protective Antigen Directed Monoclonal Antibody Improves Hemodynamic Function and Survival in a Lethal Toxin Infused Rat Model of Anthrax Sepsis. 2005:422–34. doi: 10.1086/427189. [DOI] [PubMed] [Google Scholar]
- 52.Beedham RJ, Turnbull PCB, Williamson ED. Passive transfer of protection against Bacillus anthracis infection in a murine model. Vaccine. 2001;19(31):4409–16. doi: 10.1016/s0264-410x(01)00197-9. [DOI] [PubMed] [Google Scholar]
- 53.Mohamed N, Clagett M, Li J, Jones S, Pincus S, D'Alia G, et al. A High-Affinity Monoclonal Antibody to Anthrax Protective Antigen Passively Protects Rabbits before and after Aerosolized Bacillus anthracis. Spore Challenge. 2005:795–802. doi: 10.1128/IAI.73.2.795-802.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kataoka TR, Komazawa N, Morii E, Oboki K, Nakano T. Involvement of connective tissue-type mast cells in Th1 immune responses via Stat4 expression. 2005:1016–20. doi: 10.1182/blood-2004-07-2811. [DOI] [PubMed] [Google Scholar]
- 55.Wiedemann Frank, L R, P K, J U, S HE, W KH, H RP, J G, B W, B T. Histopathological studies on the local reactions induced by complete fReund's adjuvant (CFA), bacterial lipopolysaccharide (LPS), and synthetic lipopeptide (P3C) conjugates. 1991:265–71. doi: 10.1002/path.1711640313. [DOI] [PubMed] [Google Scholar]
- 56.Alving CR. Design and selection of vaccine adjuvants: animal models and human trials. Vaccine. 2002 May 31;20(Supplement 3):S56–S64. doi: 10.1016/s0264-410x(02)00174-3. [DOI] [PubMed] [Google Scholar]
- 57.Barve M, Bender J, Senzer N, Cunningham C, Greco FA, McCune D, et al. Induction of immune responses and clinical efficacy in a phase II trial of IDM-2101, a 10-epitope cytotoxic T-lymphocyte vaccine, in metastatic non-small-cell lung cancer. J Clin Oncol. 2008 Sep 20;26(27):4418–25. doi: 10.1200/JCO.2008.16.6462. [DOI] [PubMed] [Google Scholar]
- 58.Sugai K, Shiga A, Okada K, Iwata T, Ogura H, Maekawa K, et al. Dermal testing of vaccines for children at high risk of allergies. Vaccine. 2007;25(17):3454–63. doi: 10.1016/j.vaccine.2006.12.057. [DOI] [PubMed] [Google Scholar]
- 59.Brotherton JML, Gold MS, Kemp AS, McIntyre PBM, Burgess MA, Campbell-Lloyd S, et al. Anaphylaxis following quadrivalent human papillomavirus vaccination. 2008:525–33. doi: 10.1503/cmaj.080916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Yamamoto S, Takeda Y, Yamamoto M, Kurazono H, Imaoka K, Yamamoto M, et al. Mutants in the ADP-ribosyltransferase Cleft of Cholera Toxin Lack Diarrheagenicity but Retain Adjuvanticity. 1997:1203–10. doi: 10.1084/jem.185.7.1203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Choi IH, Shin YM, Park JS, Lee MS, Han EH, Chai OH, et al. Immunoglobulin E-dependent Active Fatal Anaphylaxis in Mast Cell-deficient Mice. J Exp Med. 1998 November 2;188(9):1587–92. doi: 10.1084/jem.188.9.1587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Malaviya R, Abraham S. Role of mast cell leukotrienes in neutrophil recruitment and bacterial clearance in infectious peritonitis. J Leukoc Biol. 2000 June 1;67(6):841–6. doi: 10.1002/jlb.67.6.841. [DOI] [PubMed] [Google Scholar]
- 63.Echtenacher B, Mannel DN, Hultner L. Critical protective role of mast cells in a model of acute septic peritonitis. Nature. 1996;381(6577):75–7. doi: 10.1038/381075a0. [DOI] [PubMed] [Google Scholar]
- 64.Malaviya R, Ikeda T, Abraham SN, Malaviya R. Contribution of mast cells to bacterial clearance and their proliferation during experimental cystitis induced by type 1 fimbriated E. coli. Immunology Letters. 2004;91(23):103–11. doi: 10.1016/j.imlet.2003.10.005. [DOI] [PubMed] [Google Scholar]
- 65.Scapini P, Lapinet-Vera JA, Gasperini S, Calzetti F, Bazzoni F, Cassatella MA. The neutrophil as a cellular source of chemokines. Immunological Reviews. 2000;177(1):195–203. doi: 10.1034/j.1600-065x.2000.17706.x. [DOI] [PubMed] [Google Scholar]
- 66.Whale TA, Griebel PJ. A sheep in wolf's clothes: Can neutrophils direct the immune response? The Veterinary Journal. doi: 10.1016/j.tvjl.2007.12.030. In Press, Corrected Proof. [DOI] [PubMed] [Google Scholar]
- 67.Beauvillain C, Delneste Y, Scotet M, Peres A, Gascan H, Guermonprez P, et al. Neutrophils efficiently cross-prime naive T cells in vivo. 2007:2965–73. doi: 10.1182/blood-2006-12-063826. [DOI] [PubMed] [Google Scholar]
- 68.Tateda K, Moore TA, Deng JC, Newstead MW, Zeng X, Matsukawa A, et al. Early Recruitment of Neutrophils Determines Subsequent T1/T2 Host Responses in a Murine Model of Legionella pneumophila Pneumonia. 2001:3355–61. doi: 10.4049/jimmunol.166.5.3355. [DOI] [PubMed] [Google Scholar]
- 69.Romani L, Mencacci A, Cenci E, Del Sero G, Bistoni F, Puccetti P. An immunoregulatory role for neutrophils in CD4+ T helper subset selection in mice with candidiasis. 1997:2356–62. [PubMed] [Google Scholar]
- 70.van Gisbergen KPJM, Sanchez-Hernandez M, Geijtenbeek TBH, van Kooyk Y. Neutrophils mediate immune modulation of dendritic cells through glycosylation-dependent interactions between Mac-1 and DC-SIGN. 2005:1281–92. doi: 10.1084/jem.20041276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Bennouna S, Denkers EY. Microbial Antigen Triggers Rapid Mobilization of TNF-{alpha} to the Surface of Mouse Neutrophils Transforming Them into Inducers of High-Level Dendritic Cell TNF-{alpha} Production. 2005:4845–51. doi: 10.4049/jimmunol.174.8.4845. [DOI] [PubMed] [Google Scholar]
- 72.Maletto BA, Ropolo AS, Alignani DO, Liscovsky MV, Ranocchia RP, Moron VG, et al. Presence of neutrophil-bearing antigen in lymphoid organs of immune mice. 2006:3094–102. doi: 10.1182/blood-2006-04-016659. [DOI] [PubMed] [Google Scholar]
- 73.Fadel R, Ramboer I, Chatterjee N, Rihoux JP, Derde MP. Short communication. Cetirizine inhibits bradykinin-induced cutaneous wheal and flare in atopic and healthy subjects. Allergy. 2000;55(9):888–91. doi: 10.1034/j.1398-9995.2000.00322.x. [DOI] [PubMed] [Google Scholar]
- 74.Gold Y, Goldberg A, Sivan Y. Hyper-releasability of mast cells in family members of infants with sudden infant death syndrome and apparent life-threatening events. The Journal of Pediatrics. 2000;136(4):460–5. doi: 10.1016/s0022-3476(00)90008-8. [DOI] [PubMed] [Google Scholar]
- 75.Miloševic D, Janoševic L, Janoševic S, Invankovic Z, Dergenc R. Skin reactivity to vasomotor agents in non-eosinophilic and eosinophilic non-allergic rhinitis. Cambridge Journals Online. 2002:519–22. doi: 10.1258/002221502760132386. [DOI] [PubMed] [Google Scholar]
- 76.Rukwied R, Zeck S, Schmelz M, McGlone F. Sensitivity of human scalp skin to pruritic stimuli investigated by intradermal microdialysis in vivo. Journal of the American Academy of Dermatology. 2002;47(2):245–50. doi: 10.1067/mjd.2002.120461. [DOI] [PubMed] [Google Scholar]
- 77.Atkins P, Green GR, Zweiman B. Histologic studies of human skin test responses to ragweed, compound 48/80, and histamine. Journal of Allergy and Clinical Immunology. 1973;51(5):263–73. doi: 10.1016/0091-6749(73)90128-0. [DOI] [PubMed] [Google Scholar]