

Abstract
Caffeine is a commonly used neurostimulant that also produces cerebral vasoconstriction by antagonizing adenosine receptors. Chronic caffeine use results in an adaptation of the vascular adenosine receptor system presumably to compensate for the vasoconstrictive effects of caffeine. We investigated the effects of caffeine on cerebral blood flow (CBF) in increasing levels of chronic caffeine use. Low (mean = 45 mg/day), moderate (mean = 405 mg/day), and high (mean = 950 mg/day) caffeine users underwent quantitative perfusion magnetic resonance imaging on four separate occasions: twice in a caffeine abstinent state (abstained state) and twice in a caffeinated state following their normal caffeine use (native state). In each state, there were two drug conditions: participants received either caffeine (250 mg) or placebo. Gray matter CBF was tested with repeated‐measures analysis of variance using caffeine use as a between‐subjects factor, and correlational analyses were conducted between CBF and caffeine use. Caffeine reduced CBF by an average of 27% across both caffeine states. In the abstained placebo condition, moderate and high users had similarly greater CBF than low users; but in the native placebo condition, the high users had a trend towards less CBF than the low and moderate users. Our results suggest a limited ability of the cerebrovascular adenosine system to compensate for high amounts of daily caffeine use. Hum Brain Mapp 2009. © 2009 Wiley‐Liss, Inc.
Keywords: caffeine, cerebral blood flow, perfusion, magnetic resonance, chronic drug use
INTRODUCTION
Caffeine (1,3,7‐trimethylxanthine), a naturally occurring plant alkaloid, is the most widely used neurostimulant in the world, being present in coffee, tea, soft drinks, chocolate, analgesics, and dietary supplements. A recent review of national dietary consumption between 1994 and 1998 estimated that 87% of the population consumes caffeine, with an average of 193 mg/day. Among adults 18 years and older, the daily intake ranges between 166 and 336 mg/day [Frary et al.,2005].
A 250 mg dose of caffeine has been shown to reduce resting cerebral blood flow (CBF) between 22% and 30% [Cameron et al.,1990; Field et al.,2003; Lunt et al.,2004]. Caffeine, at normal dietary concentrations, acts as a competitive antagonist of adenosine A2A and A2B receptors located on vascular smooth muscle [Ngai et al.,2001]. Adenosine is an essential cellular constituent involved in energy metabolism and also acts as an extracellular signaling molecule by binding to its receptors that are found on virtually every cell in the body. When bound to the A2A and A2B receptor subtypes located on cerebrovascular smooth muscle, adenosine produces vasodilation [Coney and Marshall,1998; Ngai et al.,2001] by opening ATP‐ dependent K+ channels and decreasing the conductance ofCa2+ (for review see [Standen and Quayle,1998]), although there may be other mechanisms of action as well.
Evidence from receptor binding, physiology, and behavioral studies suggest an adaptation to the effects of caffeine following chronic intake. This adaptation presumably accounts for the development of tolerance to the vasoconstrictive and neurostimulant effects of caffeine. Some evidence exists for the sensitization (e.g. conversion to high affinity state) of adenosine receptors following chronic caffeine use [Green and Stiles,1986], whereas receptor binding studies suggest that the majority of this adaptation is due to receptor upregulation (e.g. increase in receptor number) [Johansson et al.,1997; Shi and Daly,1999]. Although these studies have been conducted in rodents, A2A receptors located on human platelets have been shown to be upregulated following 2 weeks of 400 mg/day of caffeine, or following 1 week of 600 mg/day [Varani et al.,1999]. Indirect evidence for receptor upregulation comes from studies that have reported the development of tolerance to the effects of caffeine on blood pressure, heart rate [Denaro et al.,1991], and subjective mood [Evans and Griffiths,1992] following repeated dosing. Withdrawal symptoms following a period of abstention also indicate a physiological adaptation to chronic caffeine use. Symptoms such as headache, fatigue, and impaired concentration emerge between 12 and 24 hours after caffeine cessation [Lane and Phillips‐Bute,1998; Nehlig,1999], and may be present from as little as 100 mg/day of chronic caffeine use [Griffiths et al.,1990]. More importantly, withdrawal headaches among caffeine users correspond with increased CBF following 20–24 hours of abstention from caffeine [Couturier et al.,1997; Jones et al.,2000]. This may be the result of adenosine binding to an increased number of vascular receptors that are normally antagonized by chronic caffeine levels.
To what extent adenosine receptors can upregulate to compensate for the effects of caffeine is a pertinent health question. There has been concern regarding the potential negative effects of caffeine, especially on cardiovascular health [James,2004; Nawrot et al.,2003]. Studies have suggested that incomplete, or partial, tolerance develops to the hypertensive effects of caffeine because complete tolerance has not been demonstrated in all study participants [Denaro et al.,1991; Lovallo et al.,2004] and because hypertensive effects persist following repeated doses of caffeine [Lane and Manus,1989]. This indicates there may be limits in the ability of adenosine receptors to adapt to chronic caffeine levels; however, no study has reported the development of cerebrovascular tolerance to caffeine in humans. Another area of concern is the impact of cerebrovascular tone on the blood oxygenation level dependent (BOLD) signal [Honey and Bullmore,2004]. Changes in vascular tone affect the magnitude and dynamics of the BOLD response [Cohen et al.,2002]. Given its widespread use, caffeine is a potential confound in all imaging studies, because participants' CBF may be abnormally low due to recent caffeine ingestion, or abnormally high due to overnight withdrawal from caffeine. Therefore, it is essential to understand the development, and the limits, of tolerance to caffeine to minimize differences in CBF in a subject sample that includes chronic caffeine consumers.
Previously, 24 hours of caffeine abstinence increased CBF among high caffeine users (mean = 986 mg/day) but not among low caffeine users (mean = 126 mg/day) [Mathew and Wilson,1985]. In an earlier study from our laboratory, we measured CBF among low (mean = 41 mg/day) and high (mean = 648 mg/day) caffeine users following 30 hours of caffeine abstinence [Field et al.,2003]. There was greater CBF among the high users after placebo, and daily caffeine intake positively correlated with CBF. The goal of the present study was to further extend our understanding of the interaction between chronic caffeine use and CBF. In particular, we measured the effects of caffeine and placebo in low, moderate, and high caffeine users following 30 hours of caffeine abstinence and following their normal caffeine consumption, conditions that we refer to as abstained placebo (AP), abstained caffeine (AC), native placebo (NP), and native caffeine (NC) in the remainder of this manuscript. Salivary caffeine concentrations were also obtained to confirm participants' compliance with the study requirements and to compare caffeine concentrations with CBF. We hypothesized that there would be an increase in CBF following caffeine abstention that is proportional to daily caffeine use, and that in the native caffeinated state there would be no differences in CBF between caffeine use groups.
METHODS
Participants
Forty‐five adult volunteers (25 women, aged 18–50 years) participated in this study between 2005 and 2007. The Institutional Review Board of Wake Forest University School of Medicine approved this study, and all participants gave informed consent to the study procedures. Potential volunteers were recruited from the Winston‐Salem, North Carolina area with flyers, newspaper advertisements, and by word of mouth. Respondents who met the following inclusion criteria were invited to participate: (1) no history of migraines, diabetes, stroke, hypertension, or any neurologic or vascular disease, (2) no current symptoms of depression or an anxiety disorder, (3) no previous brain surgery or serious CNS trauma, (4) no use of vasoactive medications, (5) no current abuse of alcohol or illicit drugs, (6) visual acuity able to be corrected to 20/40, and (7) no MR incompatible implanted devices. Participants' average daily caffeine intake was estimated from their responses to a 7‐day caffeine consumption diary (7‐day CCD; Addicott et al.,2008) modified from the Caffeine Consumption Questionnaire [Landrum,1992] and from data on the caffeine content of common beverages (Center for Science in the Public Interest web site, http://www.cspinet.org, published July 1997). Participants were then identified by their daily caffeine use as “low users” (n = 18; < 200 mg/day), “moderate users” (n = 9; 200–600 mg/day), and “high users” (n = 18; > 600 mg/day). Although there is no currently accepted cut offs for categorizing caffeine use levels, group divisions used here are consistent with previous work [Schuh and Griffiths,1997; Tinley et al.,2003]. In addition, we present analyses that correlate caffeine use with CBF to supplement findings based on these groupings. The average daily caffeine use (mg/day) recorded in the participants' 7‐day CCD was converted into body‐weight adjusted caffeine use (mg/kg/day) using participants' weight taken at the first visit.
Participant characteristics appear in Table I. There were expected differences between groups in daily caffeine use (mg/day) (one‐way analysis of variance, ANOVA: F(2,42) = 146.90, p < 0.001; Bonferroni t‐tests: low < moderate < high, p < 0.001) and in body‐weight adjusted caffeine use (mg/kg/day) (F(2,42) = 54.17, p < 0.001; t‐tests: low < moderate, p < 0.05; low and moderate < high, p < 0.001). Differences between groups in weight (1‐way ANOVA), gender, race, cigarette use, and oral birth control use (χ2 tests) were not significant. Groups differed slightly in age (F(2,42) = 6.56, p < 0.005) with low users being younger than high users (p< 0.005). Since age correlated with some measures of CBF, it was used as a covariate in CBF analyses.
Table I.
Participant characteristics
| Mean (SD) | Low users | Moderate users | High users |
|---|---|---|---|
| n = 18 | n = 9 | n = 18 | |
| Age | 27 (5) | 32 (10) | 37 (10) |
| Weight (lbs) | 144 (23) | 176 (27) | 168 (47) |
| Caffeine use | 45 (38) | 405 (103) | 950 (237) |
| mg/day | |||
| mg/kg/day | 0.8 (0.7) | 5.1 (1.2) | 13.3 (5.6) |
| Frequencies | |||
| Male/female | 8/10 | 3/6 | 9/9 |
| Race (C/AA/H/A) | 10/5/2/1 | 6/2/1/0 | 15/1/1/1 |
| Cigarette users | 0 | 3 | 3 |
| Oral birth control | 4 | 3 | 3 |
C, Caucasian; AA, African American; H, Hispanic; A, Asian.
Study Design
Each participant underwent quantitative perfusion imaging on four separate occasions in a randomized, double‐blind study design. Test sessions were at least 1 week apart to allow participants to resume normal caffeine use between scans. Two of these scans followed participants' normal caffeine use (native state) and two followed abstention from caffeine (abstained state). For the native state, participants were instructed to continue their normal daily caffeine usage until 15 min before their scheduled visit. For the abstained state, participants were instructed to avoid consuming any foods, beverages, or drugs that contain caffeine for 30 hours before their scheduled visit. This duration of abstinence was selected so that caffeine concentrations would most likely be below detection threshold (0.2 μg/ml) at the time of testing. Caffeine use or abstention from use during the 30 hours before a scan visit was verified with a 3‐day CCD. In addition, a saliva sample was obtained upon arrival to the scan visit to verify compliance with study requirements. Saliva samples were collected using Salivettes® (Sarstedt, Inc. Newton, NC), and caffeine concentrations were assayed by HPLC (Global Lifescience Solutions, LLC, Ann Arbor, MI).
Following the first saliva sample, participants ingested a capsule containing placebo or caffeine (250 mg, anhydrous). A second saliva sample was obtained 1 hour after drug administration to measure the varied levels of caffeine in each participant. Respiration rate, end‐tidal CO2, oxygen saturation (Capnocheck® II, SIMS BCI Inc., Waukesha, WI), heart rate, and blood pressure (IVAC vital check model 4410, ALARIS medical systems, Dublin, OH) were also recorded when the saliva samples were collected. Immediately following collection of the second saliva sample, participants were placed in the MRI scanner. Structural, functional (results to be reported elsewhere), and perfusion imaging were completed. The perfusion scan was collected approximately 1.5–2 hours after drug administration. During the perfusion sequence, participants were instructed to maintain their gaze on a fixation cross (E‐Prime, Psychology Software Tools, Pittsburgh, PA) delivered through MR‐compatible virtual reality goggles (Resonance Technology Inc., Los Angeles, CA). Participants' wakefulness throughout the scan was monitored via an infrared camera embedded in the goggles. Participants were scanned within the 3 hours of their peak caffeine use as determined by the 7‐day CCD.
Image Acquisition
Imaging experiments were performed on a 1.5 T GE scanner (Twin‐speed; GE Healthcare, Milwaukee, WI) with a 4‐channel neurovascular MR coil array (MEDRAD, Inc. USA, Warrendale, PA). The imaging protocol consisted of a high‐resolution anatomical image for tissue classification and a perfusion‐weighted image to measure CBF.
A T1‐weighted structural scan was acquired using an Inversion Prepared 3D Spoiled Gradient Echo; IR‐3DSPGR [Mugler and Brookeman,1990]. The parameters for this scan were as follows: TE = 2.60 msec, TR = 9.00 msec, TI = 600 msec, bandwidth = 16.93 kHz, flip angle = 20 degrees, field of view = 240 mm (frequency) × 240 mm (phase), and an acquisition matrix of 256 (frequency) × 256 (phase), 124 slices, 1.5 mm thickness, 0 mm slice gap, and an in‐plane resolution of 0.94 mm.
CBF was measured with QUantitative Imaging of Perfusion using Single Subtraction with Thin Slice TI1 Periodic Saturation: QUIPSS II TIPS, also known as Q2TIPS [Luh et al.,1999] with Flow‐sensitive Alternative Inversion Recovery (FAIR) encoding [Kim and Tsekos,1997]. Images were acquired with a single shot gradient echo Echo Planar Imaging (EPI) sequence [Mansfield,1977]. To improve perfusion sensitivity by minimizing slice imperfections [Frank et al.,1997; Yongbi et al.,1999], a C‐shaped frequency offset corrected inversion pulse (β = 1361, μ = 6) [Ordidge et al.,1996] was used for tagging the blood. Very selective saturation (VSS) pulses were used [Tran et al.,2000] and applied every 25 msec between 800 ms (TI1) [Wang et al.,2002] and 1,200 msec (TI1s) to minimize the uncertainty of the labeled blood's transit time to the imaging slice [Wong et al.,1998a]. Three VSS pulses were also applied immediately before and after the inversion pulses to suppress tissue signal in the imaging plane. The Q2TIPS‐FAIR sequence consisted of 80 alternating slice‐selective and non‐selective radiofrequency inversion pulses (label/control pairs) to improve the signal to noise ratio (imaging time 8 min 36 seconds). These label/control pairs are pair‐wise subtracted then averaged to generate a perfusion‐weighted image. The first 36 seconds (10 volumes with an initial 6 second quiescent delay) were used to establish steady state. After the initial 6 seconds, a single‐shot EPI proton density (M 0) image was acquired. The mean white matter signal from the M0 image was used to scale perfusion‐weighted images to accurately calculate absolute quantitative CBF maps according to the general kinetic model [Buxton et al.,1998]. Immediately after the EPI 90 degree excitation RF pulse, a bipolar diffusion gradient with an equivalent b value of 5.25 mm2/sec was used to suppress intraarterial spins [Yang et al.,1998]. The 10 oblique slices were parallel to the AC/PC line (2 slices below and 8 slices above the AC/PC line, 8 mm thickness, 0 mm slice gap) and were acquired inferior to superior. Additional parameters of interest for the Q2TIPS‐FAIR‐EPI sequence used in this experiment are as follows: TE = 30.4msec, TR = 3,000 sec, TI = 2,000 msec, bandwidth = 62.5 kHz, flip angle = 90 degrees, field of view = 240 mm (frequency) × 240 mm (phase), and an acquisition matrix of 64 (frequency) × 40 (phase).
Data Analysis
Perfusion data were preprocessed using SPM99 software (Wellcome Department of Imaging Neuroscience, Institute of Neurology, London). The reconstructed control and label images were motion corrected with a six‐parameter rigid body transformation using SPM99. After motion correction, control/label images were pair wise subtracted; the different images were averaged; and quantitative perfusion maps were calculated from the equation:
where CBF is the cerebral blood flow, ΔM(TI2) is the mean difference in the signal intensity between the label and control images, M 0,blood is the equilibrium magnetization of blood, α is the tagging efficiency, TI1 is the time duration of the tagging bolus, TI2 is the inversion time of each slice, and T 1,blood is the longitudinal relaxation time of blood, q p is a correction factor that accounts for the difference between the T 1 of blood and the T 1 of brain tissue [Wong et al.,1998a]. For this study, the correction factor q p was assumed to be 1, which is a reasonable approximation when the T 1 of blood and the T 1 of brain tissue are similar. Based on the literature, the T 1 of blood was assumed to be 1,200 ms at 1.5T [Simonetti et al.,1996]. The inversion efficiency was measured to be 0.95 from another experiment (data not shown). The M 0,blood is approximated from the M 0,white matter, which is measured directly from the M 0 image acquired with the perfusion weighted images [Wong et al.,1998a].
Tissue probability maps were created using an SPM99 automated segmentation of the T 1‐weighted structural scan. The T 1‐weighted image was coregistered to the M 0 image and the same transformation was applied to the tissue probability maps. The coregistered tissue probability maps were used to create a white matter mask (81% probability) and a gray matter mask (51% probability). Then the average signal intensity of white matter was calculated. The signal intensity of white matter and global scaling factors were applied to convert the perfusion‐weighted image to a quantitative CBF map that represents the magnitude of perfusion (ml/100 g tissue/min) for each voxel. Mean gray matter CBF values were calculated by averaging the perfusion values from all the voxels contained in the gray matter mask. Images were normalized within SPM5 by warping the M 0 to the SPM EPI template. The transformation was then applied to the perfusion maps. Perfusion maps were visually inspected, and no errors were detected. The normalized images were used to perform voxel‐wise comparisons, which showed no regionally‐specific changes after correcting for global blood flow changes. These results are not depicted, but the mean normalized images are shown in the figures to demonstrate perfusion changes.
Primary analyses were conducted using univariate analyses of variance (ANOVA). The pre‐ and post‐drug salivary caffeine concentrations were analyzed separately with repeated‐measures 2 (state) × 2 (drug) × 3 (caffeine use) ANOVAs. The same analyses were performed on pre‐ and post‐drug measures for respiration rate, end‐tidal CO2, O2 saturation, heart rate, and blood pressure. Potential interactions between gender and gray matter CBF were investigated with a 2 (state) × 2 (drug) × 2 (sex) ANOVA. There were no significant interactions between gender and any of the main effects or interactions; therefore, the genders were combined in further analyses. The signal to noise ratio of the M 0 images were analyzed with a 2 (state) × 2 (drug) × 3 (caffeine use) repeated‐measures ANOVA, and no significant effects were found. Gray matter CBF was analyzed with a repeated‐measures 2 (state) × 2 (drug) × 3 (caffeine use) analysis of covariance (ANCOVA) using age as the covariate. The difference from the mean age was used instead of the raw value, to avoid interfering with the main effects [Delaney and Maxwell,1981]. Difference measures in CBF between caffeine states and drug conditions were analyzed with a 2 (state) × 3 (caffeine use) and a 2 (drug) × 3 (caffeine use) ANCOVA, respectively. Follow‐up analyses included two‐tailed independent‐samples t‐tests to measure between‐caffeine use group effects, and paired‐samples t‐tests to measure within‐ caffeine use group effects. Correlations were performed using two‐tailed Pearson's product‐moment correlation. Analyses were conducted with SPSS version 15.0 (SPSS Inc., Chicago, IL) with alpha set to 0.05. CBF images were created with MRIcro v1.40 [Rorden and Brett,2000].
RESULTS
Salivary Caffeine Concentrations
Pre‐drug salivary caffeine concentrations showed participant compliance with the study requirements (see Table II). Caffeine concentrations were near zero in the abstained state and were not different between the caffeine use groups. In the native state there were differences in caffeine concentrations between caffeine use groups (state x caffeine use interaction effect [F(2,42) = 18.26, p < 0.001], Between‐subjects' Caffeine Use effect [F(2,42) = 18.53, p < 0.001]). Since there was no effect of drug condition, caffeine concentrations were averaged from the two native state measures and significant differences were found between caffeine use groups (low < moderate [t(25) = 3.71, p < 0.05]; low < high [t(34) = 5.92, p < 0.001]; moderate < high [t(25) = 2.35, p < 0.05].
Table II.
Salivary caffeine concentrations for pre‐ and post‐drug administration in the placebo and caffeine drug conditions
| Placebo | Caffeine | |||
|---|---|---|---|---|
| Pre‐ | Post‐ | Pre‐ | Post‐ | |
| Abstained state | ||||
| Low users | 0.14 (0.39) | 0.22 (0.53) | 0.07 (0.13) | 4.03 (2.00) |
| Moderate users | 0.20 (0.31) | 0.18 (0.26) | 0.17 (0.23) | 3.69 (2.01) |
| High users | 0.08 (0.12) | 0.09 (0.14) | 0.18 (0.33) | 3.79 (1.84) |
| Native state | ||||
| Low users | 0.28 (0.36) | 0.28 (0.38) | 0.36 (0.45) | 4.26 (1.45) |
| Moderate users | 1.68 (1.30) | 1.72 (1.33) | 1.85 (1.96) | 5.72 (2.33) |
| High users | 3.76 (2.77) | 3.44 (2.90) | 4.27 (2.90) | 7.29 (4.16) |
Shown are the group means (SD) in μg/ml.
As expected, post‐drug salivary caffeine concentrations increased from pre‐drug levels in the caffeine conditions, but not in the placebo conditions (drug effect [F(1,42) = 189.08, p < 0.001]). Post‐drug caffeine concentrations were greater in the native state than in the abstained state and depended on caffeine use group (state x caffeine use interaction effect [F(2,42) = 11.86, p < 0.001]). Differences between caffeine use groups were observed in the NP condition (low < moderate [t(25) = 4.30, p < 0.001], low < high [t(34) = 4.58, p < 0.001]). The state x drug interaction did not reach significance, but there were greater caffeine concentrations in the NC than the AC condition for moderate (t(8) = 3.21, p < 0.05) and high users (t(17) = 4.01, p < 0.005).
There were no main effects or interactions in the pre‐ and post‐drug measures of respiration rate, end‐tidal CO2, or O2 saturation, nor did any of these post‐drug measures correlate with CBF in each of the four state × drug conditions. The average pre‐drug levels were: respiration rate: 16.1 ± 3.5 breaths/min, end‐tidal CO2: 36.6% ± 4.5%, and oxygen saturation: 97.1% ± 1.1%. The average post‐drug levels were: respiration rate: 15.6 ± 3.4 breaths/min, end‐tidal CO2: 35.9% ± 5.2%, and oxygen saturation: 97.3% ± 1.2%. There were expected changes in heart rate and blood pressure. Following caffeine administration, heart rate decreased by an average of 8 beats/min from a resting rate of 74 beats/min, and blood pressure increased by an average of 3.0/2.4 mmHg from a mean resting value of 121/70 mmHg. These changes are similar to previously published reports on the effects of caffeine on blood pressure [Nurminen et al.,1999].
Cerebral Perfusion
Caffeine reduced gray matter CBF by 27% to approximately 60 ml/100 g of tissue/min across all caffeine users, although the decrease was larger in the abstained state (33%) than in the native state (20%; see Fig. 1). The effects of caffeine on CBF depended on the caffeine state (F(1,41) = 21.81, p < 0.001), drug condition (F(1,41) = 190.73, p < 0.001), and the level of caffeine use (state x drug x caffeine use interaction effect [F(2,41) = 8.26, p < 0.005]). Caffeine reduced CBF within each caffeine use group in both the abstained state (low [t(17) = 7.85, p < 0.001], moderate [t(8) = 5.78, p < 0.001], and high [t(17) = 8.88, p < 0.001]) and the native state (low [t(17) = 6.77, p < 0.001], moderate [t(8) = 4.36, p < 0.005], and high [t(17) = 3.13, p < 0.05]). Differences in CBF between caffeine users in the AP and NP conditions appear to be driving the interaction effect. The low users exhibited virtually no difference between these conditions, whereas there was a considerable increase in CBF in the AP condition relative to the NP condition among the moderate users (t(8) = 4.78, p < 0.005) and high users (t(17) = 5.90, p < 0.001). Compared with the low users, high users had greater CBF in the AP condition (t(34) = 2.46, p < 0.05) and had a trend for greater CBF in the AC condition (t(34) = 1.97, p = 0.057); high users also had a trend for less CBF in the NP condition (t(34) = 1.67, p = 0.104). The difference between moderate and low users in the AP condition approached significance as well (t(25) = 1.91, p = 0.068). Potential contributions of other between‐group differences to the 3‐way interaction effect were investigated with further analyses.
Figure 1.
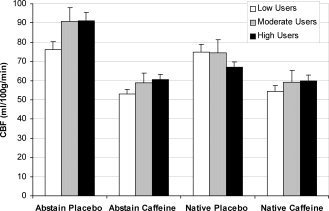
Gray matter CBF means for placebo and caffeine conditions in the abstained and native caffeinated states among all subjects. Caffeine, relative to placebo, reduced CBF in both states within each caffeine use group. High users had greater CBF than low users in the abstained placebo condition (p < 0.05), and there was greater CBF in the abstained placebo condition than in the native placebo condition for the moderate and high (p < 0.05), but not the low users. The repeated‐measures ANOVA interaction between drug condition, caffeine state, and caffeine use was significant (p < 0.005). Error bars are SEM.
To investigate how the caffeine states contributed to the state x drug x caffeine use interaction effect, differences in CBF between drug conditions in each state (abstained placebo − abstained caffeine, AP‐AC; native placebo − native caffeine, NP‐NC) were calculated (see Fig. 2). The effects of caffeine on CBF differed between caffeine users in the native state but not in the abstained state (state x caffeine use [F(2,41) = 8.26, p < 0.005]). In the native state, high users exhibited a smaller caffeine‐induced reduction in CBF than low users (t(34) = 3.53, p < 0.005) and there was a trend for high users to have a smaller reduction than moderate users (t(25) = 2.06, p = 0.050). There was also a greater reduction in CBF following caffeine in the abstained state than in the native state for moderate (t(8) = 5.90, p < 0.001) and high users (t(17) = 5.36, p < 0.001). Figure 3 illustrates the average whole brain CBF in the NP and NC conditions. The reduced CBF following caffeine administration is most evident in the low user group.
Figure 2.
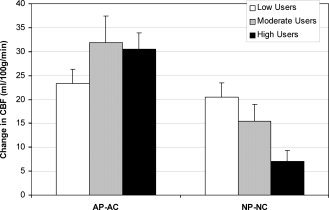
Shown are the differences in CBF between drug conditions in each caffeine state to explore the repeated‐measures ANOVA interaction between caffeine use and caffeine state. In the abstained state (AP‐AC), caffeine reduced CBF equally among the caffeine use groups. In the native state (NP‐NC), caffeine reduced CBF more among the low users than the high users (p < 0.005). Reductions in CBF following caffeine were smaller among the moderate and high users in the native state compared to the abstained state (p < 0.001). Error bars are SEM.
Figure 3.
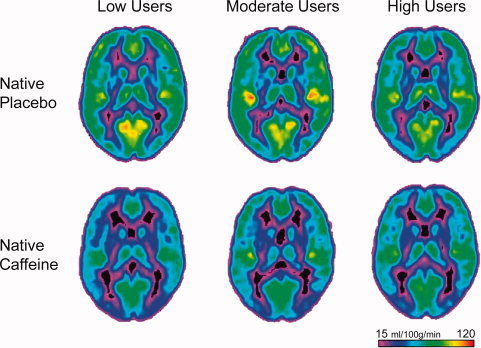
Average CBF maps (at MNI coordinates z = 14) of the low, moderate, and high caffeine users in the native placebo and native caffeine conditions. The difference in CBF following caffeine was greatest among the low users, followed by the moderate users, and the smallest difference was among the high users.
Since the quantification of caffeine use groups was arbitrary, correlational analyses were conducted between changes in CBF and body‐weight adjusted caffeine use (mg/kg/day). Caffeine use was normalized by taking the square root of the usage in mg/kg/day to produce a more uniform distribution along the abscissa. There was a negative correlation between caffeine use and the difference between placebo and caffeine conditions in the native state (NP‐NC; r = −0.48, p < 0.005; see Fig. 4). Correlations between the percent change and caffeine use were similar (r = −0.48, p < 0.005).
Figure 4.
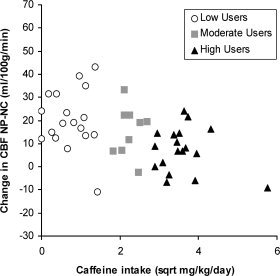
Correlation between the reduction in CBF following caffeine in the native state (NP‐NC) and caffeine use. Caffeine produced smaller reductions in CBF as daily caffeine use increased (r = −0.48, p < 0.005).
To investigate how the drug conditions contributed to the state x drug x caffeine use interaction effect, difference in CBF between states for each drug condition (abstained placebo − native placebo, AP‐NP; abstained caffeine − native caffeine, AC‐NC) were calculated (see Fig. 5). Differences in CBF varied within the placebo condition, but not within the caffeine condition (drug x caffeine use [F(2,41) = 8.26, p < 0.005]), and the effects of caffeine on CBF were dependent on the caffeine use group (between‐group effect [F(2,41) = 5.72, p < 0.05]). There was a greater difference in CBF between placebo conditions in the high users compared to the low users (t(34) = 4.23, p < 0.001) and in the moderate users compared to the low users (t(25) = 2.66, p < 0.05). Additionally, there was a greater difference in CBF in the AP‐NP condition than in the AC‐NC condition among moderate (t(8) = 5.90, p < 0.001) and high users (t(17) = 5.36, p < 0.001). Figure 6 illustrates the average whole brain CBF for the three groups in the AP and NP conditions. The increased CBF in the high users following caffeine abstention compared to the low and high users' native state is evident. There was a positive correlation between caffeine use and the difference between abstained state and native state during the placebo condition (AP‐NP; r = 0.62, p < 0.001; see Fig. 7). Correlations between the percent change and caffeine use were similar (r = 0.60, p < 0.001).
Figure 5.
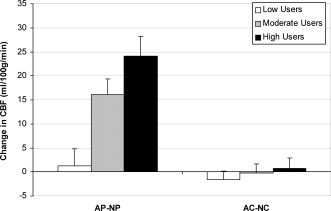
Shown are the differences in CBF between caffeine states in each drug condition to explore the repeated‐measures ANOVA interaction between caffeine use and caffeine state. Differences in CBF between the abstained and native state are apparent in the placebo condition (AP‐NP). The difference in CBF in the placebo condition was greater among the high users than the low users (p < 0.001), and the difference in the placebo condition was greater than the difference in the caffeine condition for the moderate and high users (p < 0.001). Error bars are SEM.
Figure 6.
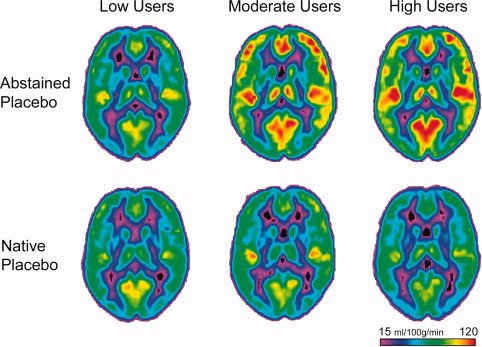
Average CBF maps (at MNI coordinates z = 14) of the low, moderate, and high caffeine users in the abstained placebo and native placebo conditions. Although there is virtually no difference in CBF between the two states among the low users, increased CBF following abstention is apparent among the moderate and high users.
Figure 7.
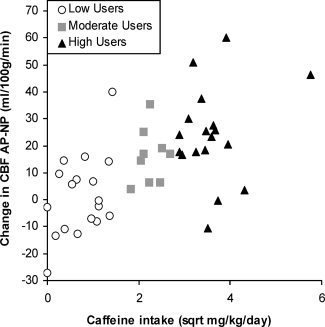
Correlation between the reduction in CBF from the abstained state to the native state (AP‐NP) and caffeine use. The difference in CBF increased as daily caffeine use increased (r = 0.62, p < 0.001).
Correlations between caffeine use and CBF in each of the four state by drug conditions were also performed. Similar to the previous study by Field et al. [2003], there was a significant relationship between CBF and caffeine use in the AP condition (r = 0.42, p < 0.005). A correlation was also found in the AC condition (r = 0.31, p < 0.05). This relationship did not reach significance in the native state. There were no correlations between CBF and salivary caffeine concentrations in any of the state by drug conditions.
DISCUSSION
We investigated the effects of caffeine and placebo in a native caffeinated state and following 30 hours of caffeine abstention among low, moderate, and high caffeine users. As expected, caffeine (250 mg) reduced gray matter CBF in both the abstained and native states, and the extent of this reduction in the native state depended on the amount of daily caffeine use. We hypothesized that there would be a step‐wise increase in CBF across low, moderate, and high users in the AP condition, and that CBF would be equal among the three groups in the NP condition. There was a positive correlation between daily caffeine use and CBF in the AP condition, but the average CBF was similar between moderate and high users, and in the NP condition, there was a trend for high users to have slightly less CBF than moderate and low users. These differences contributed to the 3‐way interaction effect found between caffeine state, drug condition, and caffeine use group.
Our interpretation is that, during chronic caffeine use, vascular adenosine receptors are upregulated to preserve the CBF at a level that would have existed if the individual had remained caffeine‐naïve. In the AP condition, this upregulation is evidenced by the increased CBF among the moderate and high users and the positive correlation between the absolute CBF and daily caffeine use. However, equal CBF between the moderate and high users suggests that receptor upregulation is not entirely proportional to caffeine use history. Either adenosine upregulation has reached an upper threshold following moderate use, or the binding of adenosine to an increased number of free receptors has produced maximum vasodilation of the blood vessels.
In the AC condition, the 250 mg dose of caffeine resulted in slightly greater CBF in the high users than the low users, and this supports our hypothesis that high users have more adenosine receptors. Alternatively, a dose of caffeine that had been adjusted for caffeine use history may have equalized post‐dose CBF, which would be advantageous if caffeine use history were a possible confound in the study. However, caffeine reduced CBF similarly across the three groups and the post‐dose CBF values were not significantly different. Therefore, it appears that this bolus dose of caffeine either produced maximum binding of available adenosine receptors, or it produced maximum vasoconstriction of the blood vessels. This is further supported by the fact that a dose of caffeine in the native state did not reduce CBF to a lower level than in the abstained state.
In the NP condition, there was similar CBF despite a step‐wise increase in salivary [CAF] across the three groups, which indicates the development of tolerance. However, high users had a trend towards less CBF than the low users, and this trend contributed to the significant relationship between caffeine use history and the difference in CBF from the AP to the NP condition. This suggests that a withdrawal, or “rebound,” effect on CBF is roughly proportional to daily caffeine use. Last, there was no difference in CBF between groups in the NC condition, nor were there differences in [CAF], possibly because of higher rates of metabolism among the moderate and high users [Berthou et al.,1995]. Interestingly, there was no difference in CBF between the AC and NC conditions, even though there was greater [CAF] in the NC condition among the moderate and high users. This supports the previous conclusion that a bolus dose of 250 mg (or possibly less) will produce maximum vasoconstriction. The difference in CBF from the NP to the NC condition may be indicative of the availability of adenosine receptors following normal caffeine use. As daily caffeine use increases, the number of free receptors decreases, and the amount of vasoconstriction possible is reduced. Similarly, a recent study showed a greater venous signal reduction in caffeine abstinent volunteers compared with regular caffeine users in a nonwithdrawn state after a 200 mg dose of caffeine [Sedlacik et al.,2008]. In the current study, the high users appear to have fewer free receptors than the low and moderate users, suggesting an incomplete adaptation.
The results of this study have health relevance. As James [2004] noted, the concerns regarding health risks associated with caffeine usage are often dampened by the belief that tolerance develops to the hypertensive and vasoconstrictive effects of caffeine [Robertson et al.,1981]. Specifically, caffeine reliably increases blood pressure when plasma caffeine concentrations are low [Smits et al.,1985], but has less consistent effects on blood pressure after the second dose of caffeine [Lane and Manus,1989]. As one study reported, some individuals develop tolerance to the pressor effects of caffeine following a second dose, whereas others do not [Lovallo et al.,2004]. In the cerebral vasculature, we found that caffeine reliably reduces CBF following both abstention and normal caffeine use. Therefore, peripheral and central vascular circulatory systems may adapt to caffeine by different mechanisms, and blood pressure may not be a good predictor of the effects of caffeine on CBF. Our results suggest that cerebral vascular adenosine receptors can compensate for moderate caffeine use (in this case <600 mg/day) because moderate users' average CBF in the NP condition was identical to that of the low users'. Higher levels of caffeine use may not be entirely compensated for given the weak correlation between caffeine use and CBF in the AP condition and since the average CBF of the high users is not greater than moderate users' following abstention.
The results of this study also have methodological significance. The vasoconstrictive effects of caffeine along with adenosine receptor upregulation following chronic use may produce unexpected effects in perfusion studies, thus making the results difficult to interpret. This should be a consideration in functional imaging studies as well, since the BOLD signal is also affected by changes in vascular tone [Cohen et al.,2002] and, in addition to its vascular effects, caffeine antagonizes neural adenosine receptors and indirectly produces neural stimulation [Fredholm et al.,1999]. Indeed, adenosine itself is thought to play a role in the coupling of the hemodynamic response to neural activity (for review see [D'Esposito et al.,2003]) and caffeine has been found to decrease the coupling ratio between CBF and the cerebral metabolic rate of oxygen consumption [Chen and Parrish,2008]. The problem of receptor upregulation is complicated because the extent of receptor upregulation is dependent on the amount of caffeine used per day and therefore is unique to each individual. In fact, the effects of caffeine on the BOLD signal response amplitude in an abstained state have been shown to positively correlate with daily caffeine use [Laurienti et al.,2002]. This is possibly due to the neural effects of caffeine outweighing the vascular effects following chronic high caffeine use [Laurienti et al.,2002].
These inconsistencies raise the methodological issue of whether to instruct study participants to refrain from caffeine use before an imaging scan, although this may result in abnormally high CBF due to caffeine withdrawal, or to instruct them to continue their normal caffeine use. Ideally, we would have perfusion measurements at several time points from the time of the last caffeine ingestion to determine when differences in CBF between caffeine users are most reduced. At the time of our perfusion measurement in the NP condition, which was at least 2 hours after the last caffeine ingestion, there were no significant differences in CBF between caffeine users. However, the mean CBF for high users exhibits a trend to be lower, and depending on the variance in the sample, this difference could approach significance. To avoid this potential error, perhaps daily caffeine use that exceeds 600 mg/day should be an exclusionary criterion for imaging studies. This should not substantially affect recruitment, since the average caffeine consumption falls into the low‐to‐moderate range [Frary et al.,2005].
Previous studies also investigating the effects of a dose of caffeine on CBF have asked participants to refrain from dietary use of caffeine anywhere from 8 to 24 hours before the experiment and have reported decreases in CBF between 10% to 30% using positron emission tomography [Cameron et al.,1990], 133Xenon inhalation [Mathew and Wilson,1985], and pulsed arterial spin labeling [Bendlin et al.,2007; Mulderink et al.,2002]. Although invasive measures such as positron emission tomography have been considered the “gold standard” of CBF quantification, the use of paramagnetic or radioactive tracers poses a greater safety risk to participants than arterial spin labeling (ASL). ASL is a practical and widely used method of measuring CBF. Continuous ASL (CASL) has slightly better signal to noise, is less sensitive to transit time, and has better control of the bolus width compared with pulsed ASL (PASL) methods, but CASL requires nonstandard hardware that is not available on most clinical scanners. A newly developed method, pseudocontinuous ASL, may improve tagging efficiency and signal to noise ratio without additional hardware in future studies [Wu et al.,2007]. Currently, PASL techniques have higher inversion efficiency and lower radiofrequency (RF) power deposition than CASL. However, a limitation of PASL was the poorly defined distal edge of the labeling plane, which introduces a systematic bias to the CBF quantification. This limitation was overcome with the inventions of QUIPPS and Q2TIPS [Luh et al.,1999; Wong et al.,1998a]. QUIPPS and Q2TIPS apply a saturation pulse at a specified time after the inversion pulse to sharply define the labeling plane's distal edge, which eliminates the systematic bias. For this study, VSS pulses were used to precisely define the distal edge of the blood bolus [Tran et al.,2000]. A potential problem is if the transit of the blood is completed before the saturation pulse is applied after the inversion pulse. For this study, we used a saturation time of 800 msec to maximize the perfusion signal while defining the distal edge of the blood bolus [Campbell and Beaulieu,2006; Wang et al.,2002; Wang and Licht,2006; Wong et al.,1998b]. According to Yang et al. [2000], the transit times of the blood across the tagging plane in healthy adults is greater than 1,200 msec when measuring CBF with PASL FAIR methods. Although Wong et al. [1998b] has reported shorter transit times of 700–800 msec across the tagging plane when using either echo‐planar imaging with signal targeting and alternating radiofrequency (EPISTAR) or proximal inversion with a control for off‐resonance effects (PICORE). These smaller transit times are a direct result of the shorter tagging region in EPISTAR and PICORE than in FAIR. In the unlikely event that the saturation time of 800 msec is longer than the blood's transit time, the measured perfusion signal would be underestimated and this would result in an underestimation of the effect being studied.
In a previous study examining participants following caffeine abstention, high caffeine users given caffeine had similar anterior gray matter CBF levels as low caffeine users given placebo [Field et al.,2003] suggesting full tolerance. In the current study, this is not the case. High caffeine users in the AC condition have less CBF than low users in the AP condition. This discrepancy is noted because both studies were conducted in our laboratory and are methodologically similar. An important difference is that the results from the Field et al. [2003] study were based on a single perfusion slice segmented into anterior and posterior gray matter, whereas our results are based on the average whole brain gray matter perfusion from 10 slices. Additionally, physiological caffeine concentrations were not measured in the Field et al. [2003] study to confirm dietary caffeine abstinence. Given our larger sample of data and more thorough methodology, our current results are probably better estimates of overall CBF.
The effects of caffeine in a nonwithdrawn state are rarely studied. One impediment to this research is deciding what constitutes a nonwithdrawn or native state, given the variability of caffeine use between caffeine consumers as well as day‐to‐day differences in caffeine consumption within individuals. Some studies have pre‐treated participants with caffeine or allowed normal dietary use before the experiment to prevent caffeine withdrawal [Heatherley et al.,2005; Warburton et al.,2001]. However, these methods either do not account for individual differences in caffeine consumption or rely on unsupervised self‐administration [James and Rogers,2005]. Another method is randomized controlled trials where participants ingest defined doses of caffeine on a daily basis while abstaining from regular dietary use [Lovallo et al.,2004]. In this case, caffeine must be administered long enough for tolerance to develop and compliance must be monitored. This drastically increases the length of time participants must remain in the study, which could compromise feasibility. Another drawback of this methodology may be that the dose administered may have different physiological effects than the one that is self‐selected. Genetic polymorphisms of the A2A receptor are associated with different levels of chronic caffeine consumption [Cornelis et al.,2007], and may be related to unique sensitivities to caffeine, especially the negative side effects such as anxiety [Alsene et al.,2003]. Hypothetically, individuals will consume an amount of caffeine that will maximize its positive effects (i.e. alertness) and minimize its negative effects (i.e. anxiety). In addition to being a more feasible methodology, there may be unique physiological features of an individual's true “native” caffeine state that are not preserved in a randomized controlled trial.
We obtained measures of salivary caffeine concentrations to monitor participant compliance with the caffeine consumption requirements. Overall, participants followed our instructions, and there was an excellent agreement between caffeine concentrations and self‐reported caffeine use before each scan. Caffeine concentrations were also measured with the intention of correlating them with CBF. Previously, we found a correlation between daily caffeine use and CBF [Field et al.,2003], and in the present study, we hypothesized that caffeine concentrations would also predict CBF. However, in each of the four state by drug conditions, there was limited variability in CBF or salivary caffeine concentrations, or both. Only in the AP condition was there a large spread of CBF values. Here, there was a modest relationship with daily caffeine use, but since salivary caffeine concentrations were near 0 μg/ml, there could not be a correlation.
There are several limitations to the present study. First, although the caffeine state and drug administration did not affect respiration rate, oxygen saturation, or end‐tidal CO2, there was an effect on heart rate and blood pressure. Although our study was not designed to test a potential interaction between blood pressure and CBF, it is unlikely that the small changes in blood pressure reported here would influence CBF independently of the effects of caffeine [Paulson et al.,1990]. Second, because our subjects were scanned on four separate occasions, differences in perfusion slice prescriptions between scans are possible. However, since the state and drug conditions were randomized and double‐blind, there is unlikely to be any systematic bias in the data. Third, our designation of low, moderate, and high caffeine use groups is somewhat arbitrary, since there is no standard definition of these groups used in research. These categories were created with the intention of producing statistically significant differences between groups, and were not based on the national average of caffeine consumption. However, we quantified an 8 oz cup of coffee as containing 135 mg caffeine; therefore, moderate caffeine use is approximately 2–4 cups of coffee per day, which encompasses estimates of the national average [Frary et al.,2005]. Last, we did not monitor subjective symptoms of withdrawal or caffeine‐induced agitation among our subjects. This dose has not been reported to produce anxiogenic side‐effects [Cysneiros et al.,2007; Lieberman et al.,1987), but withdrawal symptoms probably occurred in some of our subjects. It is unlikely that subjective symptoms would interfere with our measurement of CBF, but the relationship between changes in self‐reported symptoms and CBF following caffeine administration would be an interesting future direction.
Acknowledgements
This work was supported by NIH grants EB03880, EB004673, DA024950, NS042568, and RR07122. We thank the Center for Biomolecular Imaging, the General Clinical Research Center, and Debra Hege, for their assistance with the collection of data.
REFERENCES
- Addicott MA,Yang LL,Peiffer AM,Laurienti PJ ( 2008): Methodological considerations for the quantification of self‐reported caffeine use. Psychopharmacology (in press). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alsene K,Deckert J,Sand P,de Wit H ( 2003): Association between A(2a) receptor gene polymorphisms and caffeine‐induced anxiety. Neuropsychopharmacology 28: 1694–1702. [DOI] [PubMed] [Google Scholar]
- Bendlin BB,Trouard TR,Ryan L ( 2007): Caffeine attenuates practice effects in word stem completion as measured by fMRI BOLD signal. Hum Brain Mapp 28: 654–662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berthou F,Goasduff T,Dreano Y,Menez JF ( 1995): Caffeine increases its own metabolism through cytochrome P4501A induction in rats. Life Sci 57: 541–549. [DOI] [PubMed] [Google Scholar]
- Buxton RB,Frank LR,Wong EC,Siewert B,Warach S,Edelman RR ( 1998): A general kinetic model for quantitative perfusion imaging with arterial spin labeling. Magn Reson Med 40: 383–396. [DOI] [PubMed] [Google Scholar]
- Cameron OG,Modell JG,Hariharan M ( 1990): Caffeine and human cerebral blood flow: a positron emission tomography study. Life Sci 47: 1141–1146. [DOI] [PubMed] [Google Scholar]
- Campbell AM,Beaulieu C ( 2006): Pulsed arterial spin labeling parameter optimization for an elderly population. J Magn Resn Imaging 23: 398–403. [DOI] [PubMed] [Google Scholar]
- Chen Y,Parrish TB ( 2008): Caffeine's effects on cerebrovascular reactivity and coupling between cerebral blood flow and oxygen metabolism. Neuroimage: Article in Press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen ER,Ugurbil K,Kim SG ( 2002): Effect of basal conditions on the magnitude and dynamics of the blood oxygenation level‐dependent fMRI response. J Cereb Blood Flow Metab 22: 1042–1053. [DOI] [PubMed] [Google Scholar]
- Coney AM,Marshall JM ( 1998): Role of adenosine and its receptors in the vasodilatation induced in the cerebral cortex of the rat by systemic hypoxia. J Physiol 509 (Pt 2): 507–518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornelis MC,El‐Sohemy A,Campos H ( 2007): Genetic polymorphism of the adenosine A(2A) receptor is associated with habitual caffeine consumption. Am J Clin Nutr 86: 240–244. [DOI] [PubMed] [Google Scholar]
- Couturier EG,Laman DM,van Duijn MA,van Duijn H ( 1997): Influence of caffeine and caffeine withdrawal on headache and cerebral blood flow velocities. Cephalalgia 17: 188–190. [DOI] [PubMed] [Google Scholar]
- Cysneiros RM,Farkas D,Harmatz JS,von Moltke LL,Greenblatt DJ ( 2007): Pharmacokinetic and pharmacodynamic interactions between zolpidem and caffeine. Clin Pharmacol Ther 82: 54–62. [DOI] [PubMed] [Google Scholar]
- D'Esposito M,Deouell LY,Gazzaley A ( 2003): Alterations in the bold FMRI signal with ageing and disease: A challenge for neuroimaging. Nat Rev Neurosci 4: 863–872. [DOI] [PubMed] [Google Scholar]
- Delaney HD,Maxwell SE ( 1981): On using analysis of covariance in repeated measures designs. Multivariate Behav Res 16: 105–123. [DOI] [PubMed] [Google Scholar]
- Denaro CP,Brown CR, Jacob P,III ,Benowitz NL ( 1991): Effects of caffeine with repeated dosing. Eur J Clin Pharmacol 40: 273–278. [DOI] [PubMed] [Google Scholar]
- Evans SM,Griffiths RR ( 1992): Caffeine tolerance and choice in humans. Psychopharmacology (Berl) 108: 51–59. [DOI] [PubMed] [Google Scholar]
- Field AS,Laurienti PJ,Yen YF,Burdette JH,Moody DM ( 2003): Dietary caffeine consumption and withdrawal: confounding variables in quantitative cerebral perfusion studies? Radiology 227: 129–135. [DOI] [PubMed] [Google Scholar]
- Frank LR,Wong EC,Buxton RB ( 1997): Slice profile effects in adiabatic inversion: Application to multislice perfusion imaging. Magn Reson Med 38: 558–564. [DOI] [PubMed] [Google Scholar]
- Frary CD,Johnson RK,Wang MQ ( 2005): Food sources and intakes of caffeine in the diets of persons in the United States. J Am Diet Assoc 105: 110–113. [DOI] [PubMed] [Google Scholar]
- Fredholm BB,Battig K,Holmen J,Nehlig A,Zvartau EE ( 1999): Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol Rev 51: 83–133. [PubMed] [Google Scholar]
- Green RM,Stiles GL ( 1986): Chronic caffeine ingestion sensitizes the A1 adenosine receptor‐adenylate cyclase system in rat cerebral cortex. J Clin Invest 77: 222–227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffiths RR,Evans SM,Heishman SJ,Preston KL,Sannerud CA,Wolf B,Woodson PP ( 1990): Low‐dose caffeine physical dependence in humans. J Pharmacol Exp Ther 255: 1123–1132. [PubMed] [Google Scholar]
- Heatherley SV,Hayward RC,Seers HE,Rogers PJ ( 2005): Cognitive and psychomotor performance, mood, and pressor effects of caffeine after 4, 6 and 8 h caffeine abstinence. Psychopharmacology (Berl) 178: 461–470. [DOI] [PubMed] [Google Scholar]
- Honey G,Bullmore E ( 2004): Human pharmacological MRI. Trends Pharmacol Sci 25: 366–374. [DOI] [PubMed] [Google Scholar]
- James JE ( 2004): Critical review of dietary caffeine and blood pressure: A relationship that should be taken more seriously. Psychosom Med 66: 63–71. [DOI] [PubMed] [Google Scholar]
- James JE,Rogers PJ ( 2005): Effects of caffeine on performance and mood: withdrawal reversal is the most plausible explanation. Psychopharmacology (Berl) 182: 1–8. [DOI] [PubMed] [Google Scholar]
- Johansson B,Georgiev V,Lindstrom K,Fredholm BB ( 1997): A1 and A2A adenosine receptors and A1 mRNA in mouse brain: effect of long‐term caffeine treatment. Brain Res 762: 153–164. [DOI] [PubMed] [Google Scholar]
- Jones HE,Herning RI,Cadet JL,Griffiths RR ( 2000): Caffeine withdrawal increases cerebral blood flow velocity and alters quantitative electroencephalography (EEG) activity. Psychopharmacology 147: 371–377. [DOI] [PubMed] [Google Scholar]
- Kim SG,Tsekos NV ( 1997): Perfusion imaging by a flow‐sensitive alternating inversion recovery (FAIR) technique: application to functional brain imaging. Magn Reson Med 37: 425–435. [DOI] [PubMed] [Google Scholar]
- Landrum RE ( 1992): College students' use of caffeine and its relationship to personality. Coll Stud J 26: 151–155. [Google Scholar]
- Lane JD,Manus DC ( 1989): Persistent cardiovascular effects with repeated caffeine administration. Psychosom Med 51: 373–380. [DOI] [PubMed] [Google Scholar]
- Lane JD,Phillips‐Bute BG ( 1998): Caffeine deprivation affects vigilance performance and mood. Physiol Behav 65: 171–175. [DOI] [PubMed] [Google Scholar]
- Laurienti PJ,Field AS,Burdette JH,Maldjian JA,Yen YF,Moody DM ( 2002): Dietary caffeine consumption modulates fMRI measures. Neuroimage 17: 751–757. [PubMed] [Google Scholar]
- Lieberman HR,Wurtman RJ,Emde GG,Roberts C,Coviella IL ( 1987): The effects of low doses of caffeine on human performance and mood. Psychopharmacology (Berl) 92: 308–312. [DOI] [PubMed] [Google Scholar]
- Lovallo WR,Wilson MF,Vincent AS,Sung BH,McKey BS,Whitsett TL ( 2004): Blood pressure response to caffeine shows incomplete tolerance after short‐term regular consumption. Hypertension 43: 760–765. [DOI] [PubMed] [Google Scholar]
- Luh WM,Wong EC,Bandettini PA,Hyde JS ( 1999): QUIPSS II with thin‐slice TI1 periodic saturation: a method for improving accuracy of quantitative perfusion imaging using pulsed arterial spin labeling. Magn Reson Med 41: 1246–1254. [DOI] [PubMed] [Google Scholar]
- Lunt MJ,Ragab S,Birch AA,Schley D,Jenkinson DF ( 2004): Comparison of caffeine‐induced changes in cerebral blood flow and middle cerebral artery blood velocity shows that caffeine reduces middle cerebral artery diameter. Physiol Meas 25: 467–474. [DOI] [PubMed] [Google Scholar]
- Mansfield P ( 1977): Multi‐planar image‐formation using nmr spin echoes. J Phys C Solid State Phys 10: L55–L58. [Google Scholar]
- Mathew RJ,Wilson WH ( 1985): Caffeine consumption, withdrawal and cerebral blood‐flow. Headache 25: 305–309. [DOI] [PubMed] [Google Scholar]
- Mugler JP,III ,Brookeman JR ( 1990): Three‐dimensional magnetization‐prepared rapid gradient‐echo imaging (3D MP RAGE). Magn Reson Med 15: 152–157. [DOI] [PubMed] [Google Scholar]
- Mulderink TA,Gitelman DR,Mesulam MM,Parrish TB ( 2002): On the use of caffeine as a contrast booster for BOLD fMRI studies. Neuroimage 15: 37–44. [DOI] [PubMed] [Google Scholar]
- Nawrot P,Jordan S,Eastwood J,Rotstein J,Hugenholtz A,Feeley M ( 2003): Effects of caffeine on human health. Food Addit Contam 20: 1–30. [DOI] [PubMed] [Google Scholar]
- Nehlig A ( 1999): Are we dependent upon coffee and caffeine? A review on human and animal data. Neurosci Biobehav Rev 23: 563–576. [DOI] [PubMed] [Google Scholar]
- Ngai AC,Coyne EF,Meno JR,West GA,Winn HR ( 2001): Receptor subtypes mediating adenosine‐induced dilation of cerebral arterioles. Am J Physiol Heart Circ Physiol 280: H2329–2335. [DOI] [PubMed] [Google Scholar]
- Nurminen ML,Niittynen L,Korpela R,Vapaatalo H ( 1999): Coffee, caffeine and blood pressure: a critical review. Eur J Clin Nutr 53: 831–839. [DOI] [PubMed] [Google Scholar]
- Ordidge RJ,Wylezinska M,Hugg JW,Butterworth E,Franconi F ( 1996): Frequency offset corrected inversion (FOCI) pulses for use in localized spectroscopy. Magn Reson Med 36: 562–566. [DOI] [PubMed] [Google Scholar]
- Paulson OB,Strandgaard S,Edvinsson L ( 1990): Cerebral autoregulation. Cerebrovas Brain Metab Rev 2: 161–192. [PubMed] [Google Scholar]
- Robertson D,Wade D,Workman R,Woosley RL,Oates JA ( 1981): Tolerance to the humoral and hemodynamic effects of caffeine in man. J Clin Invest 67: 1111–1117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rorden C,Brett M ( 2000): Stereotaxic display of brain lesions. Behav Neurol 12: 191–200. [DOI] [PubMed] [Google Scholar]
- Schuh KJ,Griffiths RR ( 1997): Caffeine reinforcement: the role of withdrawal. Psychopharmacology 130: 320–326. [DOI] [PubMed] [Google Scholar]
- Sedlacik J,Helm K,Rauscher A,Stadler J,Mentzel HJ,Reichenbach JR ( 2008): Investigations on the effect of caffeine on cerebral venous vessel contrast by using susceptibility‐weighted imaging (SWI) at 1.5, 3 and 7T. Neuroimage 40: 11–18. [DOI] [PubMed] [Google Scholar]
- Shi D,Daly JW ( 1999): Chronic effects of xanthines on levels of central receptors in mice. Cell Mol Neurobiol 19: 719–732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simonetti OP,Finn JP,White RD,Laub G,Henry DA ( 1996): “Black blood” T2‐weighted inversion‐recovery MR imaging of the heart. Radiology 199: 49–57. [DOI] [PubMed] [Google Scholar]
- Smits P,Thien T,Vantlaar A ( 1985): Circulatory effects of coffee in relation to the pharmacokinetics of caffeine. Am J Cardiol 56: 958–963. [DOI] [PubMed] [Google Scholar]
- Standen NB,Quayle JM ( 1998): K+ channel modulation in arterial smooth muscle. Acta Physiol Scand 164: 549–557. [DOI] [PubMed] [Google Scholar]
- Tinley EM,Yeomans MR,Durlach PJ ( 2003): Caffeine reinforces flavour preference in caffeine‐dependent, but not long‐term withdrawn, caffeine consumers. Psychopharmacology 166: 416–423. [DOI] [PubMed] [Google Scholar]
- Tran TKC,Vigneron DB,Sailasuta N,Tropp J,Le Roux P,Kurhanewicz J, et al.( 2000): Very selective suppression pulses for clinical MRSI studies of brain and prostate cancer. Magn Reson Med 43: 23–33. [DOI] [PubMed] [Google Scholar]
- Varani K,Portaluppi F,Merighi S,Ongini E,Belardinelli L,Borea PA ( 1999): Caffeine alters A2A adenosine receptors and their function in human platelets. Circulation 99: 2499–2502. [DOI] [PubMed] [Google Scholar]
- Wang J,Licht DJ ( 2006): Pediatric perfusion MR imaging using arterial spin labeling. Neuroimaging Clin N Am 16: 149–167 ix. [DOI] [PubMed] [Google Scholar]
- Wang JJ,Alsop DC,Li L,Listerud J,Gonzalez‐At JB,Schnall MD,Detre JA ( 2002): Comparison of quantitative perfusion imaging using arterial spin labeling at 1.5 and 4.0 tesla. Mag Res Med 48: 242–254. [DOI] [PubMed] [Google Scholar]
- Warburton DM,Bersellini E,Sweeney E ( 2001): An evaluation of a caffeinated taurine drink on mood, memory and information processing in healthy volunteers without caffeine abstinence. Psychopharmacology 158: 322–328. [DOI] [PubMed] [Google Scholar]
- Wong EC,Buxton RB,Frank LR ( 1998a): Quantitative imaging of perfusion using a single subtraction (QUIPSS and QUIPSS II). Magn Reson Med 39: 702–708. [DOI] [PubMed] [Google Scholar]
- Wong EC,Buxton RB,Frank LR ( 1998b): A theoretical and experimental comparison of continuous and pulsed arterial spin labeling techniques for quantitative perfusion imaging. Magn Reson Med 40: 348–355. [DOI] [PubMed] [Google Scholar]
- Wu WC,Fernandez‐Seara M,Detre JA,Wehrli FW,Wang J ( 2007): A theoretical and experimental investigation of the tagging efficiency of pseudocontinuous arterial spin labeling. Mag Res Med 58: 1020–1027. [DOI] [PubMed] [Google Scholar]
- Yang Y,Engelien W,Xu S,Gu H,Silbersweig DA,Stern E ( 2000): Transit time, trailing time, and CBF during brain activation: measurement using multislice, pulsed spin‐labeling perfusion imaging. Magn Reson Med 44: 680–685. [DOI] [PubMed] [Google Scholar]
- Yang YH,Frank JA,Hou L,Ye FQ,McLaughlin AC,Duyn JH ( 1998): Multislice imaging of quantitative cerebral perfusion with pulsed arterial spin labeling. Magn Reson Med 39: 825–832. [DOI] [PubMed] [Google Scholar]
- Yongbi MN,Yang Y,Frank JA,Duyn JH ( 1999): Multislice perfusion imaging in human brain using the C‐shaped frequency offset corrected inversion pulse: comparison with hyperbolic secant. Magn Reson Med 42: 1098–1105. [DOI] [PubMed] [Google Scholar]