

Abstract
The formation of bile depends on the structural and functional integrity of the bile-secretory apparatus and its impairment, in different situations, results in the syndrome of cholestasis. The structural bases that permit bile secretion as well as various aspects related with its composition and flow rate in physiological conditions will first be reviewed. Canalicular bile is produced by polarized hepatocytes that hold transporters in their basolateral (sinusoidal) and apical (canalicular) plasma membrane. This review summarizes recent data on the molecular determinants of this primary bile formation. The major function of the biliary tree is modification of canalicular bile by secretory and reabsorptive processes in bile-duct epithelial cells (cholangiocytes) as bile passes through bile ducts. The mechanisms of fluid and solute transport in cholangiocytes will also be discussed. In contrast to hepatocytes where secretion is constant and poorly controlled, cholangiocyte secretion is regulated by hormones and nerves. A short section dedicated to these regulatory mechanisms of bile secretion has been included. The aim of this revision was to set the bases for other reviews in this series that will be devoted to specific issues related with biliary physiology and pathology.
Keywords: Hepatocytes, Cholangiocytes, Bile flow, Bile acid, Transport
INTRODUCTION
In 1924 Cramer and Ludford[1] published a paper in which they indicated: "liver cells present an unusual problem of secretory cellular activity. The cells of all other secreting glands are functionally unipolar, but hepatocytes are bipolar". Furthermore they wrote that "all the known facts compel a return to the old conception of Claude Bernard that glycogenic function represents an internal secretion of the liver". It is clear that bile secretion is the external one. Finally, regarding this functional bipolarity, those authors reported "it is difficult to form a mental conception of the cellular mechanism which enables one cell to pass two different specific secretions in different directions"; towards sinusoids or towards canaliculi. In that paper they offered morphological evidence that the Golgi apparatus is involved in secretion by hepatocytes in both directions.
Since then our understanding of bile secretion has evolved, and differerent attempts have been made to resolve diverging conceptual theories. In the fifties we learned that bile flow is not the result of a hydrostatic perfusion[2] but the result of osmotic forces[3]. In the sixties we began to distinguish between canalicular and ductular bile[4,5] and showed that bile flow is related to the amount of bile salts secreted to the canaliculi[6]. Shortly after, it was shown that the canalicular bile may be either dependent or independent of bile salts[7,8].
The use of different methods and techniques, such as chronic bile fistula[9], isolated and perfused livers[10], couplets of hepatocytes[11], isolated bile duct units[12], separation of membranes from basolateral and apical domains[13] and several other experimental approaches have helped us to advance our knowledge of the different mechanism involved in the formation of bile flow.
Over the last two decades molecular biology techniques have allowed the cloning of different proteins expressed in cholangiocytes[14] and/or hepatocytes; such proteins may be involved in the transport of endogenous or exogenous organic anions and cations[15]. The overall role of these transporters depends on whether they are targeted from the Golgi apparatus to the apical or basolateral poles of the epithelial liver cells[16,17]. This may help to resolve the conceptual problem of bipolarity, pointed out by Cramer and Ludford. The discovery of nuclear receptors and plasma membrane receptors for bile acids (BAs)[18,19] has opened a new field of investigation regarding the role of these molecules in the control of secretory and metabolic mechanisms[20,21] that somehow links different aspects of endogenous and exogenous secretions to metabolic functions in parenchymal liver cells. This issue will be the matter of a separate review in this series.
In this introductory review, a brief revision is made of the structural bases that permit bile secretion as well as several aspects related to its composition and flow rate under certain physiological conditions. Knowledge of the role in bile flow generation of cholangiocytes has increased exponentially over the last few years[22,23] and is therefore reviewed here. Finally, a short section dedicated to the regulatory mechanisms of bile secretion is included. This should set the bases for other reviews in this series that will be devoted to specific issues related to biliary physiology and pathology.
STRUCTURAL BASES
The biliary apparatus is a convergent system of canals that begins in the canaliculi, followed by the bile ducts, and ending with the common bile duct (coledochus). Bile secretion depends on the function of membrane transport systems in hepatocytes and cholangiocytes and on the structural and functional integrity of the biliary tree. The hepatocytes, constituting the most abundant liver cell population (65%), generate the so-called primary bile in their canaliculi[4]. Biliary canaliculi are blind tubular structures, with a very high surface/volume ratio that favors-by means of osmotic gradients-the formation of bile flow[5]. Cholangiocytes, which constitute 3%-5% of the liver cells[24], modify the canalicular bile by secretory and reabsorptive processes as bile passes through the bile ducts[22], and they are responsible for approximately 30% of bile volume[25]. In contrast to hepatocytes, where secretion is constant and poorly controlled[26], cholangiocytes secretion is broadly regulated[22,27].
Hepatocytes exhibit structural and functional polarity, three different zones being distinguished in their membranes. The sinusoidal membrane, which faces Disse’s space, covers 37% of the total surface of the hepatocyte. There is no lamina basal between hepatocytes and endothelial cells, which are fenestrated and show abundant vacuole of endocytosis and exocytosis, accounting for the intense exchange of substances between blood and hepatocytes[28]. In the lateral membrane (50% of the total surface) there are specialized structures that allow adhesion (desmosomes and tight-junctions) and comunication (gap-junctions) between adjacent hepatocytes. The tight-junctions determine the exchange of fluids and electrolytes between Disse’s space and the canalicular space through the paracelular pathway[28]. Together, the sinusoidal and lateral membranes cover the basolateral surface (basolateral membrane). The canaliculi are tiny ducts delimited by the canalicular or apical membrane membrane of two adjacent hepatocytes[28] and they represent a small fraction of the total hepatocyte surface area.
Currently, the molecular biology and genetic characteristics of many transporters of the basolateral and apical membranes are known (Table 1). Such transporters take part in the transfer of substances between blood and hepatocytes and between hepatocytes and bile, respectively. Likewise, many transporters expressed in cholangiocyte membranes are also known (Table 2). In this review only those transporters with a clear role in the genesis of physiological bile flow are addressed. More comprehensive reviews are available elsewhere[29-31].
Table 1.
Membrane transporters in hepatocytes
| Abbreviation | Function |
| Basolateral membrane (sinusoidal) | |
| NTCP/SLC10A11 | Takes up BAs |
| OATP/SLC21A1 | Takes up BAs and OA- and exports BAs, GSH, HCO3- |
| MRP3/ABCC3 | Export OA- conjugates, GSH |
| MRP4/ABCC4 | Export BAs, GSH |
| OSTa /OSTb | Organic solute transporter: Exports BAs |
| NBC4c/SLC4A51 | Na+-HCO3- symporter, acid extruder |
| NHE1/SLC9A11 | Na+/H+ exchanger, acid extruder |
| SK2 | Potassium channel, potassium efflux |
| SLC12A2 | Na+-K+-Cl- symporter: Sodium, potassium, chloride uptake |
| Apical membrane (canalicular) | |
| BSEP/ABCB111, formerly SPGE | Bile salt export pump |
| MRP2/ABCC21, formerly cMOAT | Export non BAs OA-, GSH |
| MDR1 | Efflux of lipofilic cations |
| MDR3/ABCB4 | Phospholipid flipase |
| ABCG5/ABCG8 | Export sterols |
| AE2/SLC4A21 | Cl-/HCO3- anion exchanger: Acid loader |
| Cl- channel1 | Export chloride |
| AQP81 | Water channel |
| NHE3/SLC9A31 | Na+/H+ exchanger: Acid extruder |
Transporters relevant to bile flow under physiological basal conditions.
Table 2.
Membrane transporters in cholangiocytes: Abbreviations and function
| Abbreviation | Function |
| Basolateral membrane | |
| NDCBE/SLC4A81 | Na+-dependent Cl-/HCO3- exchanger: |
| Import HCO3- and Na+, Export H+ and Cl- | |
| NHE/SLC91 | Na+/H+ exchanger: Acid extruder |
| AQP41 | Water channel |
| tASBT/SLC10A2 | Export BAs and Na+ |
| SK2 | Potassium channel, potassium efflux |
| SLC12A2 | Import Na+-K+-2Cl- |
| MRP3/ABCC3 | Export OA- conjugates, GSH |
| MRP4/ABCC4 | Export BAs, GSH |
| Apical membrane | |
| AE2/SLC4A21 | Cl-/HCO3- anion exchanger: Acid loader |
| CFTR1 | Cl- channel: Export chloride |
| AQP11 | Water channel |
| NBCe/NBC4/SLC4A5 | Na+-HCO3- symporter: Acid extruder |
| ASBT/SLC10A2 | BAs-Na+, symport: Uptake of BAs and Na+ |
Transporters relevant to bile flow under physiological basal conditions.
The Golgi complex and the network of microtubules and microfilaments are important structures for the exocrine function of hepatocytes and also for the mechanisms of bile formation. The pericanalicular space, free of cellular organelles, contains actin microfilaments that reach to the microvilli of the canalicular membrane. Microtubules are distributed throughout the cytoplasm. The vesicles from the Golgi complex are vehicles for substances to be excreted in bile as well as plasma proteins, including transporters, to be placed at apical or basolateral membranes[17,32]. Newly synthesized apical ABC (ATP-binding cassette) transporters are transferred from the Golgi apparatus to the canalicular membrane[16]. This traffic is dependent on intact microtubule and microfilament systems[17,32].
The hepatocytes and the biliary system are closely related to the blood vascular elements, both forming a functional unit: the hepatic acinus[33]. The blood flow generates concentrative gradients of oxygen and nutrients along the sinusoids. These gradients allow a division of the hepatic acinus into three different zones according to its distance from the portal space: the periportal or zone I, zone II and the centrilobular or zone III. Moreover, these gradients cause functional heterogeneity between the hepatocytes of the different zones[34,35].
BILE COMPOSITION
Bile mainly consists of water, in which there are organic and inorganic substances in suspension, dissolved, or in equilibrium between both states. In bile samples, collected from the human common bile duct, the concentrations of the inorganic electrolytes sodium (Na+), potassium (K+), calcium (Ca2+) and bicarbonate (HCO3-) are slightly higher than their plasmatic concentrations, whereas biliary chloride (Cl-) concentrations are slightly lower than these found in plasma. BAs concentrations range between 2 and 45 mmol/L. The concentrations of biliary pigments range from 50 to 200 mg/100 mL. Proteins and peptides, such as glutathione, are also found in bile[36]. It is also possible to detect glucose and small amounts of endogenous substances such as thyroid and steroid hormones[37]. Human bile is rich in lipids. Thus, phospholipids concentrations seem to range between 25 and 810 mg/100 mL, whereas these of cholesterol vary between 60 and 320 mg/100 mL, with average ratios of phospholipids to BA of 0.3 and cholesterol to BA of 0.07 (Figure 1). Humans differ from other animals in the fact that our species eliminates cholesterol from the body to a greater extent as cholesterol itself rather than by converting it into BAs[38].
Figure 1.
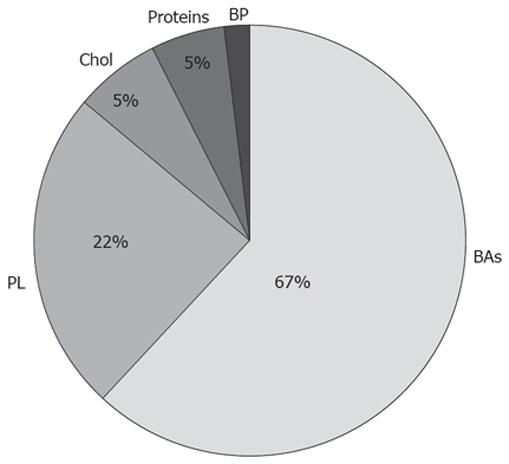
Organic composition of bile. Perceptual distribution of BAs, Phospholipids (PL), Cholesterol (Chol), Proteins and biliary pigments (BP).
BILE FLOW
The mean basal flow of bile in humans is approximately 620 mL/d. One portion of this flow (220 mL/d, 35%) is determined by the secretion of BAs and is called the BAs-dependent canalicular fraction -BADFc-[8]. In addition, there is a linear correlation between the amount of BAs secreted into bile and the amount of water that follows them (7-25 mL/mmol). This choleretic activity of individual BAs is species-dependent and varies according to its chemical structure, conjugational condition and relative concentrations. Choleretic activity is lower for BA species that have a higher tendency to form micellar aggregates in bile[39]. These findings explain the different contributions of BADFc to the bile flow among species (30%-60%)[3,40]. Certain BAs (ursodesoxycholic and the nor-derivatives of ursodesoxycholic and quenodeoxycholic acids) generate a volume of bile higher than that expected from their osmotic force. In order to explain such hypercholeresis, it has been proposed that these BAs would be reabsorbed, in their protonated form, by cholangiocytes. From there, they would be effluxed to blood to reach the sinusoids, where they would be taken up again by hepatocytes and re-secreted to bile, increasing the magnitude of the BADFc[41,42]. This phenomenon, so-called "the cholehepatic shunt pathway" is discussed below in the section devoted to ductular processes.
The amount of canalicular bile independent of the osmotic force of BAs (235 mL/d, 38%) has been designated the BA-independent canalicular fraction (BAIFc)[7,8]. Quantitatively, in humans the BAIFc represents up to a 40% of total primarily formed bile[43]. In other species it varies between 30% and 60%[5,40,44]. The ductular fraction of bile flow has a high value (30%) in humans[25], although it also varies among different species[8,24].
THE BA-DEPENDENT CANALICULAR FRACTION (BADFc)
BAs are supplied either by synthesis in liver cells or from the sinusoidal blood as part of de enterohepatic circulation (EHC).
Sinusoidal uptake
The BAs in sinusoidal blood are efficiently taken up by hepatocytes from Disse’s space despite being highly albumin bound, due to the existence in the basolateral membrane of transporters[45,46]. This uptake is carried out against an electrochemical gradient, is saturable[37] and depends on the structure of the BA. Thus, it is more efficient for trihydroxyl- than for dihydroxyl-BA and for conjugated more than for unconjugated BAs[47]. The sodium taurocholate-cotransporting polypeptide (NTCP), the main Na+-dependent BAs transporter[30,48] is only expressed in the basolateral membrane[49]. The uptake of BAs by Na+-independent mechanisms seems to be mediated by less specific transporters, known as organic anion-transporting polypeptides (OATPs), which exchange these molecules for other anions, such as HCO3-, glutathione (GSH) or even other BAs[30,47]. These transporters may take up BAs (mainly non conjugated forms), endogenous OA- (thyroid hormones, monoconjugated bilirubin) and xenobiotic compounds (toxins, drugs, food components, etc)[50]. The quantitative relevance of the different isoforms of these transporters in sodium-independent BA uptake by hepatocytes is still not completely understood (Figure 2). The heterodimeric protein OSTalpha/OSTbeta is expressed at the basal membrane of hepatocytes and cholangiocytes[51]. This is a sodium-independent BA transporter that may play a role in BA efflux from hepatocytes toward blood when these compounds get accumulated in cholestatic conditions. Moreover, in cholangiocytes, in addition to play a similar role, this transporter may also be involved in the cholehepatic shunting of BAs.
Figure 2.

Sinusoidal transport and canalicular secretion. Left: BA dependent canalicular fraction of bile (BADFc). Right: canalicular and sinusoidal secretion of anionic and cationic organic molecules.
Transcellular transport
In order to explain the transit of BAs from the sinusoidal membrane to the pericanalicular region, two different, not mutually excluding, mechanisms have been proposed: (1) simple diffusion of BAs bound to intracellular proteins[52]; (2) and/or vesicular transport of BAs driven by cytoskeleton contractile activity[53,54]. Two arguments have been raised against the role of the second mechanism. One is that hepatic transit of labeled BAs is too fast[54]. The second one is that the baseline secretion of BAs is not modified by microtubules disruption[53]. However, the overload of BAs intensify the vesicular trafficking from the Golgi complex to the pericanalicular zone[55], and under these circumstances the alteration in the functional integrity of the cytoskeleton results in impaired BA secretion[56] and subsequently cholestasis[57].
The quantity of ABC transporters in the apical membrane is regulated by the amount of biliary components available for secretion[58,59]. The regulated intracellular vesicular traffic of canalicular ABC transporters[59,60] is crucial for normal bile secretion. The bile salt export pump (BSEP, formerly SPGE, a sister of P-glycoprotein) is the main, if not the only, canalicular BA transporter[61], and it is also located in subcanalicular vesicles that may act as an intracellular pool. It is therefore probable that the impaired secretion of BAs observed in overloaded conditions would be an indirect result of the distortion of the increased vesicular traffic of transporters to the canalicular membrane[56]. These and other studies[17,62] have established not only the actual role of vesicular trafficking in hepatocytes, but also that a specific vesicle trafficking machinery is required for membrane polarity. The overall functions based on hepatocyte polarity are not attributable to the mere presence of transporters in both poles of these cells[63] but also to their intracellular trafficking and temporary anchorage to the different hepatocyte membranes (Figure 2).
Canalicular secretion
At the end of the eighties it was believed that BAs were extruded to the canalicular lumen by an electric gradient, being negative inside hepatocytes (around -37 mV)[64]. However, this gradient is not strong enough to impose higher concentrative differences, such as BAs up to 200 times more concentrated in canaliculus than in hepatocyte[26]. It is now known that the secretion of BAs to the canalicular lumen is a saturable phenomenon mediated by a transporter[65]. The energy-dependent bile secretion of BAs is mediated by BSEP[30,58,63]. This export pump was cloned in1998[54] and since them it has been studied in detail[66,67]. It is currently known that BSEP transports both conjugated and unconjugated BAs, sulfated lithocholyl conjugates[68], and a variety of drugs[69]. Other apical ATP binding cassette (ABC) transporters are required: multidrug resistance-P glycoprotein 3 (MDR3) for phospholipids, MDR1 for lipofilic cationic drugs, multidrug resistance-associated protein 2 (MRP2) for non-BA organic anions[70,71] and the heterodimeric protein ABCG5/G8 involved in the secretion of cholesterol and other sterols, such as fitosterols[72] (Figure 2).
BAIFc
The osmotic activity of BAs is not the only determinant of bile flow. Certain substances with osmotic activity, both endogenous and exogenous, may also play a role in canalicular bile generation, without modifying the BA secretion rate[73]. In certain situations, such as diabetes mellitus, there appears to be a reduction in bile flow, without impairment in BA secretion[74]. Glutathione and bicarbonate seem to play a similar quantitative role in BAIFc formation[13].
Multidrug resistance-associated protein 2 (MRP2, formerly cMOAT, canalicular multispecific organic-anion transporter) transports a broad range of amphipathic anionic substrates, including glutathione conjugates, glucuronosyl bilirubin and sulfated BA derivatives[63]. The ATP-dependent canalicular excretion of GSH is one of the main forces responsible for the generation of BAIFc[75]. Under basal conditions, the biliary levels of this tripeptide reache up to 5 mmol/L. This is a sufficient amount to account for the formation of bile by osmotic force[36]. Rats heterozygous and homozygous for inactivating mutations in Mrp2 secrete less GSH to bile, 37% and 99% below control levels, respectively[76].
The secretion of HCO3- is carried out by the canalicular antiporter named AE2[13,77]. This system functions in connection with the canalicular water channel aquaporin (AQP8) and the apical chloride channel, the cystic fibrosis transmembrane regulator (CFTR)[13,77]. However, the force that maintains favorable Cl- gradients remains to be defined. The AE2 antiporter requires the existence of suitable internal levels of HCO3- through cotransport of the anion with sodium in the basolateral membrane by the NBCe symporter[78,79] and/or by its formation activated by the carbonic anhydrase (CA) pathway[80]. This latter mechanism is linked to H+ extrusion via Na+/H+ antiporters (NHE) working in both the basolateral (NHE1)[81] and canalicular (NHE3)[82] membranes. In turn, sodium cations are extruded via a sodium pump. This is why bicarbonate secretion is said to be is a concentrative mechanism that indirectly requires metabolic energy. Canalicular bicarbonate excretion is upregulated by glucagon[83], which also enhances AQP8-mediated water permeability at the canaliculi[84]. These choleretic effects are microtubular-dependent and involve mobilization of intracellular vesicles[83,84]. The other osmotically active inorganic components of bile are not as important as HCO3- in generating BAIFc[37,85] (Figure 3).
Figure 3.
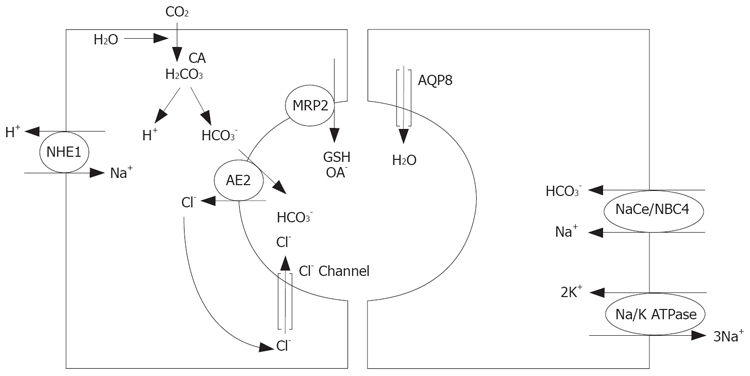
Sinusoidal transport and canalicular secretion. Left: BA independent canalicular fraction of bile (BAIFc). Right: Water an electrolyte movement. CA: carbonic anhydrase.
DUCTULAR PROCESSES
Cholangiocytes exert a series of reabsorptive and secretory processes that dilute and alkalinize the primary (canalicular) bile during its passage along the biliary tract[24,86,87]. Cholangiocytes secrete fluid, HCO3-, Cl- and carry out the reabsorption of glucose, BAs (cholehepatic shunt), glutamate, conjugated bilirubin, BSP, and small OAs-[87]. As a result, osmotic gradients generate an extra flow of bile known as the ductular BA-independent fraction (BAIFd). These processes are regulated by bile constituents, nerves and hormones[86,88]. The biliary transport of bicarbonate is a relevant function of the cholangiocytes. An electroneutral sodium-independent Cl-/HCO3- exchange activity (AE2) has been observed in the apical membrane of cholangiocytes[77]. There is also a cAMP-responsive Cl- channel (CFTR) that is coordinated with AE2 to play a role in biliary excretion of HCO3-[27,89]. These apical fluxes of anions, in the presence of aquaporins (AQP1) contribute to the BAIFd[90]. This coordinated function became more evident after discovering that a pool of AE2, CFTR and AQP1 is stored in cholangiocyte intracellular vesicles, which are co-redistributed to the apical membrane under secretin stimulation[90]. The CA pathway and an Cl-/HCO3- exchanger provide the required level of HCO3- and the H+ is subsequently extruded by a coupled carrier-mediated basolateral H+/Na+ exchanger (NHE)[91,92]. In humans, the import of HCO3- into cholangiocytes occurs mainly through electroneutral Na+-dependent Cl-/HCO3- anion exchange (NDCBE)[93] (Figure 4).
Figure 4.

Ductular secretion and reabsorption. BA independent ductular fraction of bile (BAIFd).
Lipophilic, unconjugated BAs, such as ursodeoxycholic acids, are passively reabsorbed in cholangiocytes, which constitutes the first essential step in the cholehepatic shunt model[94]. An active transport for conjugated BAs has been described in rat cholangiocytes[95], which expands the idea of cholehepatic shunting of BAS. With the identification of apical (ASBT) and basolateral (tASBT) BAs carriers in cholangiocytes[96], the hypothesis of the cholehepatic BA shunt received additional support[97].
Regulatory factors
Secretin receptors (SCTRs) are exclusively expressed at the basolateral membrane of cholangiocytes[98] and when they are stimulated intracellular levels of cAMP are increased[99]. The pool of AE2, CFTR and AQP1 stored in intracellular vesicles is redistributed to the apical membrane under cAMP or secretin stimulation[90] and secretin stimulation activates CFTR through cAMP. Both effects together explain the increase in HCO3-efflux[37]. Most experiments with rats and rabbits have used animals with induced bile duct proliferation[100], since normal rats[23,90] and rabbits[101] respond very poorly to secretin. ASBT activity increases acutely upon secretin stimulation[102], which may accentuate the cholehepatic BAs shunting in the postprandial period.
On cholangiocytes, acetylcholine increases both secretin-stimulated cAMP synthesis and Cl-/HCO3-exchanger activity[103,104]. Vagotomy in BDL rats inhibits secretin-stimulated ductal secretion and decrease cholangiocyte cAMP levels[88]. Bombesin can act either by increasing the secretin release in dogs[105], or inducing ductal secretion with activated Cl-/HCO3- exchange via secretin-independent mechanisms in isolated rat cholangiocytes[86]. VIP increases secretin-stimulated bile flow and HCO3- excretion in humans[106]. Dopamine, somatostatin and, gastrin to some extent, inhibit basal and secretin-stimulated bicarbonate-rich choleresis[86,107,108].
Footnotes
Peer reviewers: Milan Jirsa, PhD, Laboratory of Experimental Medicine-building Z1, Institute for Clinical and Experimental Medicine, Videnska 1958/9, Praha 414000, Czech; Liqing Yu, MD, PhD, Assistant Professor, Department of Pathology, Lipid Sciences, Director of Transgenic Mouse Core Facility Wake Forest University School of Medicine, Medical Center Blvd Winston-Salem, NC 27157-1040, United States
S- Editor Li DL E- Editor Lin YP
References
- 1.Cramer W, Ludford RJ. On the cellular mechanism of bile secretion and its relation to the Golgi apparatus of the liver cell. J Physiol. 1926;62:74–80. doi: 10.1113/jphysiol.1926.sp002337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Brauer RW, Leong GF, Holloway RJ. Mechanics of bile secretion; effect of perfusion pressure and temperature on bile flow and bile secretion pressure. Am J Physiol. 1954;177:103–112. doi: 10.1152/ajplegacy.1954.177.1.103. [DOI] [PubMed] [Google Scholar]
- 3.Sperber I. Secretion of organic anions in the formation of urine and bile. Pharmacol Rev. 1959;11:109–134. [PubMed] [Google Scholar]
- 4.Forker EL. Two sites of bile formation as determined by mannitol and erythritol clearance in the guinea pig. J Clin Invest. 1967;46:1189–1195. doi: 10.1172/JCI105612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wheeler HO, Ross ED, Bradley SE. Canalicular bile production in dogs. Am J Physiol. 1968;214:866–874. doi: 10.1152/ajplegacy.1968.214.4.866. [DOI] [PubMed] [Google Scholar]
- 6.Preisig R, Cooper H, Wheeler HO. The relationship between taurocholate secretion rate and bile production in the unanesthetized dog during cholinergic blockade and during secretin administration. J Clin Invest. 1962;41:1152–1162. doi: 10.1172/JCI104568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Boyer JL, Klatskin G. Canalicular bile flow and bile secretory pressure. Evidence for a non-bile salt dependent fraction in the isolated perfused rat liver. Gastroenterology. 1970;59:853–859. [PubMed] [Google Scholar]
- 8.Erlinger S, Dhumeaux D, Berthelot P, Dumont M. Effect of inhibitors of sodium transport on bile formation in the rabbit. Am J Physiol. 1970;219:416–422. doi: 10.1152/ajplegacy.1970.219.2.416. [DOI] [PubMed] [Google Scholar]
- 9.Jimenez R, Esteller A, Lopez MA. Biliary secretion in conscious rabbits: surgical technique. Lab Anim. 1982;16:182–185. doi: 10.1258/002367782781110250. [DOI] [PubMed] [Google Scholar]
- 10.Reichen J, Paumgartner G. Kinetics of taurocholate uptake by the perfused rat liver. Gastroenterology. 1975;68:132–136. [PubMed] [Google Scholar]
- 11.Graf J, Gautam A, Boyer JL. Isolated rat hepatocyte couplets: a primary secretory unit for electrophysiologic studies of bile secretory function. Proc Natl Acad Sci USA. 1984;81:6516–6520. doi: 10.1073/pnas.81.20.6516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Boyer JL. Isolated hepatocyte couplets and bile duct units--novel preparations for the in vitro study of bile secretory function. Cell Biol Toxicol. 1997;13:289–300. doi: 10.1023/a:1007435408208. [DOI] [PubMed] [Google Scholar]
- 13.Meier PJ, Knickelbein R, Moseley RH, Dobbins JW, Boyer JL. Evidence for carrier-mediated chloride/bicarbonate exchange in canalicular rat liver plasma membrane vesicles. J Clin Invest. 1985;75:1256–1263. doi: 10.1172/JCI111824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Tietz P, de Groen PC, Anderson NL, Sims C, Esquer-Blasco R, Meheus L, Raymackers J, Dauwe M, LaRusso NF. Cholangiocyte-specific rat liver proteins identified by establishment of a two-dimensional gel protein database. Electrophoresis. 1998;19:3207–3212. doi: 10.1002/elps.1150191825. [DOI] [PubMed] [Google Scholar]
- 15.Suzuki H, Sugiyama Y. Transporters for bile acids and organic anions. Pharm Biotechnol. 1999;12:387–439. doi: 10.1007/0-306-46812-3_14. [DOI] [PubMed] [Google Scholar]
- 16.Sai Y, Nies AT, Arias IM. Bile acid secretion and direct targeting of mdr1-green fluorescent protein from Golgi to the canalicular membrane in polarized WIF-B cells. J Cell Sci. 1999;112(Pt 24):4535–4545. doi: 10.1242/jcs.112.24.4535. [DOI] [PubMed] [Google Scholar]
- 17.Wakabayashi Y, Lippincott-Schwartz J, Arias IM. Intracellular trafficking of bile salt export pump (ABCB11) in polarized hepatic cells: constitutive cycling between the canalicular membrane and rab11-positive endosomes. Mol Biol Cell. 2004;15:3485–3496. doi: 10.1091/mbc.E03-10-0737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Makishima M, Okamoto AY, Repa JJ, Tu H, Learned RM, Luk A, Hull MV, Lustig KD, Mangelsdorf DJ, Shan B. Identification of a nuclear receptor for bile acids. Science. 1999;284:1362–1365. doi: 10.1126/science.284.5418.1362. [DOI] [PubMed] [Google Scholar]
- 19.Parks DJ, Blanchard SG, Bledsoe RK, Chandra G, Consler TG, Kliewer SA, Stimmel JB, Willson TM, Zavacki AM, Moore DD, et al. Bile acids: natural ligands for an orphan nuclear receptor. Science. 1999;284:1365–1368. doi: 10.1126/science.284.5418.1365. [DOI] [PubMed] [Google Scholar]
- 20.Chiang JY. Bile acid regulation of gene expression: roles of nuclear hormone receptors. Endocr Rev. 2002;23:443–463. doi: 10.1210/er.2000-0035. [DOI] [PubMed] [Google Scholar]
- 21.Chiang JY. Nuclear receptor regulation of lipid metabolism: potential therapeutics for dyslipidemia, diabetes, and chronic heart and liver diseases. Curr Opin Investig Drugs. 2005;6:994–1001. [PubMed] [Google Scholar]
- 22.Alpini G, Phillips JO, LaRusso NF. The biology of the biliary epithelia. In: Arias IM, Boyer JL, Fausto N, Jakoby WB, Schachter DA, et al., editors. The Liver: Biology and Pathobiology, 3rd edn. Vol. 6. New York: Raven Press; 1994. pp. 623–653. [Google Scholar]
- 23.Banales JM, Prieto J, Medina JF. Cholangiocyte anion exchange and biliary bicarbonate excretion. World J Gastroenterol. 2006;12:3496–3511. doi: 10.3748/wjg.v12.i22.3496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Tavoloni N. The intrahepatic biliary epithelium: an area of growing interest in hepatology. Semin Liver Dis. 1987;7:280–292. doi: 10.1055/s-2008-1040583. [DOI] [PubMed] [Google Scholar]
- 25.Nathanson MH, Boyer JL. Mechanisms and regulation of bile secretion. Hepatology. 1991;14:551–566. [PubMed] [Google Scholar]
- 26.Arrese M, Accatino L. From blood to bile: recent advances in hepatobiliary transport. Ann Hepatol. 2002;1:64–71. [PubMed] [Google Scholar]
- 27.Alvaro D, Cho WK, Mennone A, Boyer JL. Effect of secretion on intracellular pH regulation in isolated rat bile duct epithelial cells. J Clin Invest. 1993;92:1314–1325. doi: 10.1172/JCI116705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jones AL, Schmucker DL, Renston RH, Murakami T. The architecture of bile secretion. A morphological perspective of physiology. Dig Dis Sci. 1980;25:609–629. doi: 10.1007/BF01318875. [DOI] [PubMed] [Google Scholar]
- 29.Hagenbuch B, Meier PJ. The superfamily of organic anion transporting polypeptides. Biochim Biophys Acta. 2003;1609:1–18. doi: 10.1016/s0005-2736(02)00633-8. [DOI] [PubMed] [Google Scholar]
- 30.Meier PJ, Stieger B. Bile salt transporters. Annu Rev Physiol. 2002;64:635–661. doi: 10.1146/annurev.physiol.64.082201.100300. [DOI] [PubMed] [Google Scholar]
- 31.Trauner M, Boyer JL. Bile salt transporters: molecular characterization, function, and regulation. Physiol Rev. 2003;83:633–671. doi: 10.1152/physrev.00027.2002. [DOI] [PubMed] [Google Scholar]
- 32.Kipp H, Arias IM. Trafficking of canalicular ABC transporters in hepatocytes. Annu Rev Physiol. 2002;64:595–608. doi: 10.1146/annurev.physiol.64.081501.155793. [DOI] [PubMed] [Google Scholar]
- 33.Rappaport AM. The microcirculatory acinar concept of normal and pathological hepatic structure. Beitr Pathol. 1976;157:215–243. doi: 10.1016/s0005-8165(76)80083-2. [DOI] [PubMed] [Google Scholar]
- 34.Gonzalez J, Esteller A. Heterogeneity of rabbit hepatocytes for bile secretion after acinar zone 3 damage induced by bromobenzene. Effect of bilirubin and bile salt infusions. Biochem Pharmacol. 1985;34:507–514. doi: 10.1016/0006-2952(85)90181-9. [DOI] [PubMed] [Google Scholar]
- 35.Katz NR. Metabolic heterogeneity of hepatocytes across the liver acinus. J Nutr. 1992;122:843–849. doi: 10.1093/jn/122.suppl_3.843. [DOI] [PubMed] [Google Scholar]
- 36.Inoue M, Kinne R, Tran T, Arias IM. The mechanism of biliary secretion of reduced glutathione. Analysis of transport process in isolated rat-liver canalicular membrane vesicles. Eur J Biochem. 1983;134:467–471. doi: 10.1111/j.1432-1033.1983.tb07590.x. [DOI] [PubMed] [Google Scholar]
- 37.Erlinger S. Bile flow. In: Arias IM, Popper H, Jacoby WB, Schachter D, Shafritz DA, et al., editors. The Liver: Biology and Pathobiology. New York: Raven Press; 1988. pp. 643–664. [Google Scholar]
- 38.Quintao E, Grundy SM, Ahrens EH Jr. Effects of dietary cholesterol on the regulation of total body cholesterol in man. J Lipid Res. 1971;12:233–247. [PubMed] [Google Scholar]
- 39.Zsembery A, Thalhammer T, Graf J. Bile Formation: a Concerted Action of Membrane Transporters in Hepatocytes and Cholangiocytes. News Physiol Sci. 2000;15:6–11. doi: 10.1152/physiologyonline.2000.15.1.6. [DOI] [PubMed] [Google Scholar]
- 40.Erlinger S, Dhumeaux D. Mechanisms and control of secretion of bile water and electrolytes. Gastroenterology. 1974;66:281–304. [PubMed] [Google Scholar]
- 41.Dumont M, Erlinger S, Uchman S. Hypercholeresis induced by ursodeoxycholic acid and 7-ketolithocholic acid in the rat: possible role of bicarbonate transport. Gastroenterology. 1980;79:82–89. [PubMed] [Google Scholar]
- 42.Garcia-Marin JJ, Corbic M, Dumont M, de Couet G, Erlinger S. Role of H+ transport in ursodeoxycholate-induced biliary HCO-3 secretion in the rat. Am J Physiol. 1985;249:G335–G341. doi: 10.1152/ajpgi.1985.249.3.G335. [DOI] [PubMed] [Google Scholar]
- 43.Erlinger S. Does Na+-K+-atpase have any role in bile secretion? Am J Physiol. 1982;243:G243–G247. doi: 10.1152/ajpgi.1982.243.4.G243. [DOI] [PubMed] [Google Scholar]
- 44.Boyer JL. Canalicular bile formation in the isolated perfused rat liver. Am J Physiol. 1971;221:1156–1163. doi: 10.1152/ajplegacy.1971.221.4.1156. [DOI] [PubMed] [Google Scholar]
- 45.Frimmer M, Ziegler K. The transport of bile acids in liver cells. Biochim Biophys Acta. 1988;947:75–99. doi: 10.1016/0304-4157(88)90020-2. [DOI] [PubMed] [Google Scholar]
- 46.Hagenbuch B, Meier PJ. Sinusoidal (basolateral) bile salt uptake systems of hepatocytes. Semin Liver Dis. 1996;16:129–136. doi: 10.1055/s-2007-1007226. [DOI] [PubMed] [Google Scholar]
- 47.Meier PJ, Eckhardt U, Schroeder A, Hagenbuch B, Stieger B. Substrate specificity of sinusoidal bile acid and organic anion uptake systems in rat and human liver. Hepatology. 1997;26:1667–1677. doi: 10.1002/hep.510260641. [DOI] [PubMed] [Google Scholar]
- 48.Arrese M, Ananthananarayanan M, Suchy FJ. Hepatobiliary transport: molecular mechanisms of development and cholestasis. Pediatr Res. 1998;44:141–147. doi: 10.1203/00006450-199808000-00001. [DOI] [PubMed] [Google Scholar]
- 49.Hagenbuch B, Meier PJ. Molecular cloning, chromosomal localization, and functional characterization of a human liver Na+/bile acid cotransporter. J Clin Invest. 1994;93:1326–1331. doi: 10.1172/JCI117091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kullak-Ublick GA, Hagenbuch B, Stieger B, Schteingart CD, Hofmann AF, Wolkoff AW, Meier PJ. Molecular and functional characterization of an organic anion transporting polypeptide cloned from human liver. Gastroenterology. 1995;109:1274–1282. doi: 10.1016/0016-5085(95)90588-x. [DOI] [PubMed] [Google Scholar]
- 51.Ballatori N. Biology of a novel organic solute and steroid transporter, OSTalpha-OSTbeta. Exp Biol Med (Maywood) 2005;230:689–698. doi: 10.1177/153537020523001001. [DOI] [PubMed] [Google Scholar]
- 52.Kaplowitz N. Physiological significance of glutathione S-transferases. Am J Physiol. 1980;239:G439–G444. doi: 10.1152/ajpgi.1980.239.6.G439. [DOI] [PubMed] [Google Scholar]
- 53.Crawford JM, Berken CA, Gollan JL. Role of the hepatocyte microtubular system in the excretion of bile salts and biliary lipid: implications for intracellular vesicular transport. J Lipid Res. 1988;29:144–156. [PubMed] [Google Scholar]
- 54.Lamri Y, Roda A, Dumont M, Feldmann G, Erlinger S. Immunoperoxidase localization of bile salts in rat liver cells. Evidence for a role of the Golgi apparatus in bile salt transport. J Clin Invest. 1988;82:1173–1182. doi: 10.1172/JCI113714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Crawford JM, Vinter DW, Gollan JL. Taurocholate induces pericanalicular localization of C6-NBD-ceramide in isolated hepatocyte couplets. Am J Physiol. 1991;260:G119–G132. doi: 10.1152/ajpgi.1991.260.1.G119. [DOI] [PubMed] [Google Scholar]
- 56.Dubin M, Maurice M, Feldmann G, Erlinger S. Influence of colchicine and phalloidin on bile secretion and hepatic ultrastructure in the rat. Possible interaction between microtubules and microfilaments. Gastroenterology. 1980;79:646–654. [PubMed] [Google Scholar]
- 57.Trauner M, Meier PJ, Boyer JL. Molecular pathogenesis of cholestasis. N Engl J Med. 1998;339:1217–1227. doi: 10.1056/NEJM199810223391707. [DOI] [PubMed] [Google Scholar]
- 58.Gerloff T, Stieger B, Hagenbuch B, Madon J, Landmann L, Roth J, Hofmann AF, Meier PJ. The sister of P-glycoprotein represents the canalicular bile salt export pump of mammalian liver. J Biol Chem. 1998;273:10046–10050. doi: 10.1074/jbc.273.16.10046. [DOI] [PubMed] [Google Scholar]
- 59.Kipp H, Pichetshote N, Arias IM. Transporters on demand: intrahepatic pools of canalicular ATP binding cassette transporters in rat liver. J Biol Chem. 2001;276:7218–7224. doi: 10.1074/jbc.M007794200. [DOI] [PubMed] [Google Scholar]
- 60.Gatmaitan ZC, Nies AT, Arias IM. Regulation and translocation of ATP-dependent apical membrane proteins in rat liver. Am J Physiol. 1997;272:G1041–G1049. doi: 10.1152/ajpgi.1997.272.5.G1041. [DOI] [PubMed] [Google Scholar]
- 61.Strautnieks SS, Bull LN, Knisely AS, Kocoshis SA, Dahl N, Arnell H, Sokal E, Dahan K, Childs S, Ling V, et al. A gene encoding a liver-specific ABC transporter is mutated in progressive familial intrahepatic cholestasis. Nat Genet. 1998;20:233–238. doi: 10.1038/3034. [DOI] [PubMed] [Google Scholar]
- 62.Lapierre LA, Kumar R, Hales CM, Navarre J, Bhartur SG, Burnette JO, Provance DW Jr, Mercer JA, Bahler M, Goldenring JR. Myosin vb is associated with plasma membrane recycling systems. Mol Biol Cell. 2001;12:1843–1857. doi: 10.1091/mbc.12.6.1843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Paulusma CC, Bosma PJ, Zaman GJ, Bakker CT, Otter M, Scheffer GL, Scheper RJ, Borst P, Oude Elferink RP. Congenital jaundice in rats with a mutation in a multidrug resistance-associated protein gene. Science. 1996;271:1126–1128. doi: 10.1126/science.271.5252.1126. [DOI] [PubMed] [Google Scholar]
- 64.Weinman SA, Graf J, Boyer JL. Voltage-driven, taurocholate-dependent secretion in isolated hepatocyte couplets. Am J Physiol. 1989;256:G826–G832. doi: 10.1152/ajpgi.1989.256.5.G826. [DOI] [PubMed] [Google Scholar]
- 65.Sippel CJ, Ananthanarayanan M, Suchy FJ. Isolation and characterization of the canalicular membrane bile acid transport protein of rat liver. Am J Physiol. 1990;258:G728–G737. doi: 10.1152/ajpgi.1990.258.5.G728. [DOI] [PubMed] [Google Scholar]
- 66.Stieger B, Meier Y, Meier PJ. The bile salt export pump. Pflugers Arch. 2007;453:611–620. doi: 10.1007/s00424-006-0152-8. [DOI] [PubMed] [Google Scholar]
- 67.Suchy FJ, Ananthanarayanan M. Bile salt excretory pump: biology and pathobiology. J Pediatr Gastroenterol Nutr. 2006;43 Suppl 1:S10–S16. doi: 10.1097/01.mpg.0000226385.71859.5f. [DOI] [PubMed] [Google Scholar]
- 68.Hayashi H, Takada T, Suzuki H, Onuki R, Hofmann AF, Sugiyama Y. Transport by vesicles of glycine- and taurine-conjugated bile salts and taurolithocholate 3-sulfate: a comparison of human BSEP with rat Bsep. Biochim Biophys Acta. 2005;1738:54–62. doi: 10.1016/j.bbalip.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 69.Hirano M, Maeda K, Hayashi H, Kusuhara H, Sugiyama Y. Bile salt export pump (BSEP/ABCB11) can transport a nonbile acid substrate, pravastatin. J Pharmacol Exp Ther. 2005;314:876–882. doi: 10.1124/jpet.105.084830. [DOI] [PubMed] [Google Scholar]
- 70.Nies AT, Gatmaitan Z, Arias IM. ATP-dependent phosphatidylcholine translocation in rat liver canalicular plasma membrane vesicles. J Lipid Res. 1996;37:1125–1136. [PubMed] [Google Scholar]
- 71.Thiebaut F, Tsuruo T, Hamada H, Gottesman MM, Pastan I, Willingham MC. Cellular localization of the multidrug-resistance gene product P-glycoprotein in normal human tissues. Proc Natl Acad Sci USA. 1987;84:7735–7738. doi: 10.1073/pnas.84.21.7735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Kidambi S, Patel SB. Cholesterol and non-cholesterol sterol transporters: ABCG5, ABCG8 and NPC1L1: a review. Xenobiotica. 2008;38:1119–1139. doi: 10.1080/00498250802007930. [DOI] [PubMed] [Google Scholar]
- 73.Aza MJ, Gonzalez J, Esteller A. Effect of diethyl maleate pretreatment on biliary excretion and choleretic action of sulfobromophthalein in rats. Arch Int Pharmacodyn Ther. 1986;281:321–330. [PubMed] [Google Scholar]
- 74.Garcia-Marin JJ, Villanueva GR, Esteller A. Diabetes-induced cholestasis in the rat: possible role of hyperglycemia and hypoinsulinemia. Hepatology. 1988;8:332–340. doi: 10.1002/hep.1840080224. [DOI] [PubMed] [Google Scholar]
- 75.Ballatori N, Truong AT. Glutathione as a primary osmotic driving force in hepatic bile formation. Am J Physiol. 1992;263:G617–G624. doi: 10.1152/ajpgi.1992.263.5.G617. [DOI] [PubMed] [Google Scholar]
- 76.Paulusma CC, van Geer MA, Evers R, Heijn M, Ottenhoff R, Borst P, Oude Elferink RP. Canalicular multispecific organic anion transporter/multidrug resistance protein 2 mediates low-affinity transport of reduced glutathione. Biochem J. 1999;338(Pt 2):393–401. [PMC free article] [PubMed] [Google Scholar]
- 77.Martinez-Anso E, Castillo JE, Diez J, Medina JF, Prieto J. Immunohistochemical detection of chloride/bicarbonate anion exchangers in human liver. Hepatology. 1994;19:1400–1406. [PubMed] [Google Scholar]
- 78.Fitz JG, Persico M, Scharschmidt BF. Electrophysiological evidence for Na+-coupled bicarbonate transport in cultured rat hepatocytes. Am J Physiol. 1989;256:G491–G500. doi: 10.1152/ajpgi.1989.256.3.G491. [DOI] [PubMed] [Google Scholar]
- 79.Renner EL, Lake JR, Scharschmidt BF, Zimmerli B, Meier PJ. Rat hepatocytes exhibit basolateral Na+/HCO3- cotransport. J Clin Invest. 1989;83:1225–1235. doi: 10.1172/JCI114005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Buanes T, Grotmol T, Veel T, Landsverk T, Ridderstrale Y, Raeder MG. Importance of carbonic anhydrase for canalicular and ductular choleresis in the pig. Acta Physiol Scand. 1988;133:535–544. doi: 10.1111/j.1748-1716.1988.tb08438.x. [DOI] [PubMed] [Google Scholar]
- 81.Moseley RH, Meier PJ, Aronson PS, Boyer JL. Na-H exchange in rat liver basolateral but not canalicular plasma membrane vesicles. Am J Physiol. 1986;250:G35–G43. doi: 10.1152/ajpgi.1986.250.1.G35. [DOI] [PubMed] [Google Scholar]
- 82.Mennone A, Biemesderfer D, Negoianu D, Yang CL, Abbiati T, Schultheis PJ, Shull GE, Aronson PS, Boyer JL. Role of sodium/hydrogen exchanger isoform NHE3 in fluid secretion and absorption in mouse and rat cholangiocytes. Am J Physiol Gastrointest Liver Physiol. 2001;280:G247–G254. doi: 10.1152/ajpgi.2001.280.2.G247. [DOI] [PubMed] [Google Scholar]
- 83.Benedetti A, Strazzabosco M, Ng OC, Boyer JL. Regulation of activity and apical targeting of the Cl-/HCO3- exchanger in rat hepatocytes. Proc Natl Acad Sci USA. 1994;91:792–796. doi: 10.1073/pnas.91.2.792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Gradilone SA, Garcia F, Huebert RC, Tietz PS, Larocca MC, Kierbel A, Carreras FI, Larusso NF, Marinelli RA. Glucagon induces the plasma membrane insertion of functional aquaporin-8 water channels in isolated rat hepatocytes. Hepatology. 2003;37:1435–1441. doi: 10.1053/jhep.2003.50241. [DOI] [PubMed] [Google Scholar]
- 85.Scharschmidt BF, Van Dyke RW. Mechanisms of hepatic electrolyte transport. Gastroenterology. 1983;85:1199–1214. [PubMed] [Google Scholar]
- 86.Kanno N, LeSage G, Glaser S, Alpini G. Regulation of cholangiocyte bicarbonate secretion. Am J Physiol Gastrointest Liver Physiol. 2001;281:G612–G625. doi: 10.1152/ajpgi.2001.281.3.G612. [DOI] [PubMed] [Google Scholar]
- 87.Strazzabosco M. New insights into cholangiocyte physiology. J Hepatol. 1997;27:945–952. doi: 10.1016/s0168-8278(97)80338-8. [DOI] [PubMed] [Google Scholar]
- 88.LeSagE G, Alvaro D, Benedetti A, Glaser S, Marucci L, Baiocchi L, Eisel W, Caligiuri A, Phinizy JL, Rodgers R, et al. Cholinergic system modulates growth, apoptosis, and secretion of cholangiocytes from bile duct-ligated rats. Gastroenterology. 1999;117:191–199. doi: 10.1016/s0016-5085(99)70567-6. [DOI] [PubMed] [Google Scholar]
- 89.Cohn JA, Strong TV, Picciotto MR, Nairn AC, Collins FS, Fitz JG. Localization of the cystic fibrosis transmembrane conductance regulator in human bile duct epithelial cells. Gastroenterology. 1993;105:1857–1864. doi: 10.1016/0016-5085(93)91085-v. [DOI] [PubMed] [Google Scholar]
- 90.Tietz PS, Marinelli RA, Chen XM, Huang B, Cohn J, Kole J, McNiven MA, Alper S, LaRusso NF. Agonist-induced coordinated trafficking of functionally related transport proteins for water and ions in cholangiocytes. J Biol Chem. 2003;278:20413–20419. doi: 10.1074/jbc.M302108200. [DOI] [PubMed] [Google Scholar]
- 91.Henry RP. Multiple roles of carbonic anhydrase in cellular transport and metabolism. Annu Rev Physiol. 1996;58:523–538. doi: 10.1146/annurev.ph.58.030196.002515. [DOI] [PubMed] [Google Scholar]
- 92.Spirli C, Granato A, Zsembery K, Anglani F, Okolicsanyi L, LaRusso NF, Crepaldi G, Strazzabosco M. Functional polarity of Na+/H+ and Cl-/HCO3- exchangers in a rat cholangiocyte cell line. Am J Physiol. 1998;275:G1236–G1245. doi: 10.1152/ajpgi.1998.275.6.G1236. [DOI] [PubMed] [Google Scholar]
- 93.Grubman SA, Perrone RD, Lee DW, Murray SL, Rogers LC, Wolkoff LI, Mulberg AE, Cherington V, Jefferson DM. Regulation of intracellular pH by immortalized human intrahepatic biliary epithelial cell lines. Am J Physiol. 1994;266:G1060–G1070. doi: 10.1152/ajpgi.1994.266.6.G1060. [DOI] [PubMed] [Google Scholar]
- 94.Hofmann AF. Biliary secretion and excretion in health and disease: current concepts. Ann Hepatol. 2007;6:15–27. [PubMed] [Google Scholar]
- 95.Lazaridis KN, Pham L, Tietz P, Marinelli RA, deGroen PC, Levine S, Dawson PA, LaRusso NF. Rat cholangiocytes absorb bile acids at their apical domain via the ileal sodium-dependent bile acid transporter. J Clin Invest. 1997;100:2714–2721. doi: 10.1172/JCI119816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Lazaridis KN, Tietz P, Wu T, Kip S, Dawson PA, LaRusso NF. Alternative splicing of the rat sodium/bile acid transporter changes its cellular localization and transport properties. Proc Natl Acad Sci USA. 2000;97:11092–11097. doi: 10.1073/pnas.200325297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Alpini G, Glaser S, Francis H, Marzioni M, Venter J, LeSage G. Bile acid interaction with cholangiocytes. In: Alpini G, Alvaro D, Marzioni M, LeSage G, LaRusso N, et al., editors. The Pathophysiology of the Biliary Epithelia, Georgetown, TX: Landes Bioscience; 2004. pp. 112–126. [Google Scholar]
- 98.Farouk M, Vigna SR, McVey DC, Meyers WC. Localization and characterization of secretin binding sites expressed by rat bile duct epithelium. Gastroenterology. 1992;102:963–968. doi: 10.1016/0016-5085(92)90183-y. [DOI] [PubMed] [Google Scholar]
- 99.Levine RA, Hall RC. Cyclic AMP in secretin choleresis. Evidence for a regulatory role in man and baboons but not in dogs. Gastroenterology. 1976;70:537–544. [PubMed] [Google Scholar]
- 100.Jimenez R, Torres D, Gomez-Bautista M, Esteller A. Basal and secretin induced hipercholeresis in experimental biliary cirrhosis: The role of ductular-duct proliferation. J Hepatol. 1991:13 Sup 2, S38. [Google Scholar]
- 101.Esteller A, Lopez MA. The effect of secretin and cholecystokinin-pancreozymin on the secretion of bile in the anaesthetized rabbit. Q J Exp Physiol Cogn Med Sci. 1977;62:353–359. doi: 10.1113/expphysiol.1977.sp002407. [DOI] [PubMed] [Google Scholar]
- 102.Alpini G, Glaser S, Baiocchi L, Francis H, Xia X, Lesage G. Secretin activation of the apical Na+-dependent bile acid transporter is associated with cholehepatic shunting in rats. Hepatology. 2005;41:1037–1045. doi: 10.1002/hep.20653. [DOI] [PubMed] [Google Scholar]
- 103.Hirata K, Nathanson MH. Bile duct epithelia regulate biliary bicarbonate excretion in normal rat liver. Gastroenterology. 2001;121:396–406. doi: 10.1053/gast.2001.26280. [DOI] [PubMed] [Google Scholar]
- 104.Nathanson MH, Burgstahler AD, Mennone A, Boyer JL. Characterization of cytosolic Ca2+ signaling in rat bile duct epithelia. Am J Physiol. 1996;271:G86–G96. doi: 10.1152/ajpgi.1996.271.1.G86. [DOI] [PubMed] [Google Scholar]
- 105.Kaminski DL, Deshpande YG. Effect of somatostatin and bombesin on secretin-stimulated ductular bile flow in dogs. Gastroenterology. 1983;85:1239–1247. [PubMed] [Google Scholar]
- 106.Nyberg B, Einarsson K, Sonnenfeld T. Evidence that vasoactive intestinal peptide induces ductular secretion of bile in humans. Gastroenterology. 1989;96:920–924. [PubMed] [Google Scholar]
- 107.Glaser S, Alvaro D, Roskams T, Phinizy JL, Stoica G, Francis H, Ueno Y, Barbaro B, Marzioni M, Mauldin J, et al. Dopaminergic inhibition of secretin-stimulated choleresis by increased PKC-gamma expression and decrease of PKA activity. Am J Physiol Gastrointest Liver Physiol. 2003;284:G683–G694. doi: 10.1152/ajpgi.00302.2002. [DOI] [PubMed] [Google Scholar]
- 108.Tietz PS, Alpini G, Pham LD, Larusso NF. Somatostatin inhibits secretin-induced ductal hypercholeresis and exocytosis by cholangiocytes. Am J Physiol. 1995;269:G110–G118. doi: 10.1152/ajpgi.1995.269.1.G110. [DOI] [PubMed] [Google Scholar]