

Abstract
Purpose
The aim of this study was to investigate whether the nucleus parafascicularis (Pf) of the thalamus could be a relay of the control of epileptic seizures by the superior colliculus (SC). The Pf is one of the main ascending projection of the SC, the disinhibition of which has been shown to suppress seizures in different animal models and has been proposed as the main relay of the nigral control of epilepsy.
Methods
Rats with genetic absence seizures (Generalized Absence Epilepsy Rat from Strasbourg or GAERS) were used in this study. The effect of bilateral microinjection of picrotoxin, a GABA antagonist, in the SC on the glutamate and GABA extracellular concentration within the Pf was first investigated using microdialysis. In a second experiment, the effect of direct activation of Pf neurons on the occurrence of absence seizures was examined by microinjection of low doses of kainate, a glutamate agonist.
Results
Bilateral injection of picrotoxin (33 pmol/side) in the SC suppressed spike and wave discharges for 20 min. This treatment resulted in an increase of glutamate but not GABA levels in the Pf during the same time course. Bilateral injection of kainate (35 pmol/side) into the Pf significantly suppressed spike and wave discharges for 20 min, whereas such injection were without effects when at least one the site was located outside the Pf.
Conclusions
These data suggest that glutamatergic projections to the Pf could be involved in the control of seizures by the SC. Disinhibition of these neurons could lead to seizure suppression and may be involved in the nigral control of epilepsy.
Keywords: Animals; Disease Models, Animal; Electroencephalography; statistics & numerical data; Epilepsy, Absence; physiopathology; prevention & control; Excitatory Amino Acid Agonists; pharmacology; Extracellular Fluid; chemistry; drug effects; GABA Antagonists; pharmacology; Glutamates; analysis; drug effects; physiology; Intralaminar Thalamic Nuclei; chemistry; drug effects; physiopathology; Kainic Acid; administration & dosage; pharmacology; Male; Microdialysis; Microinjections; Neural Pathways; drug effects; physiopathology; Picrotoxin; administration & dosage; pharmacology; Rats; Rats, Inbred Strains; Substantia Nigra; physiopathology; Superior Colliculi; drug effects; physiopathology; gamma-Aminobutyric Acid; analysis; physiology
Keywords: epilepsy, animal model, control, substantia nigra, superior colliculus, parafascicular nucleus, microdialysis, microinjections, thalamus, absence epilepsy
Introduction
Several lines of evidence have suggested that the basal ganglia and especially the substantia nigra pars reticulata (SNR), their main output station, are involved in the control of epileptic seizures (1, 2). In particular, it has been shown that any pharmacological or electrical manipulation of the basal ganglia that results in inhibition of the SNR, suppresses the occurrence of most epileptic seizures (for review see 2). Yet, the mechanisms and circuits involved in this “nigral control of epilepsy” remain to be characterized.
Recently, we have shown that the neuronal populations of the intermediate and superficial layers of the caudal superior colliculus (SC) may constitute an important relay for the control of absence seizures by the basal ganglia, via the SNR (3). One of the main output of the SC is the ascending projection to the centromedial and the parafascicular nuclei (Pf) of the thalamus, two main components of the intralaminar nuclei (4, 5). Both experimental and clinical studies show that the intralaminar nuclei may play a role in the modulation and/or maintenance of seizures and neuronal oscillations (6, 7, 8).
The aim of this study was thus to investigate whether the Pf, the main intralaminar nucleus, could be a relay of the nigral control of epileptic seizures. To this aim, the genetic model of absence epilepsy in the rat (Genetic Absence Epilepsy Rat from Strasbourg or GAERS) was used and involvement of the collicular projection to the Pf during suppression of absence seizures was investigated by measuring variations in the extracellular levels of GABA and glutamate in the Pf in GAERS. In a second experiment, the consequences of Pf activation on the occurrence of absence seizures was studied by measuring the effects of local injection of a low, non toxic, dose of kainate on the occurrence of spike and wave discharges (SWD) in GAERS.
Methods
Adult male GAERS rats (250–350 g), a strain with spontaneous generalized non-convulsive seizures, bred at U398 Inserm in Strasbourg were kept in individual cages under a 12/12 h normal light/dark cycle, with food and water ad libitum. They were implanted under general anesthesia (diazepam 4 mg/kg + ketamine 1,000 mg/kg, i.p.) with four single-contact recording electrodes as previously described (3). Stainless-steel guide-cannulae (o.d. = 0.4 mm; i.d. = 0.3 mm) were aimed bilaterally at (i) the deep layers of the caudal part of the SC (A/P: −7.3 mm; M/L: 2.0 mm; D/V: −4.0 mm from bregma) or (ii) the Pf (A/P: −4.80 mm; M/L: 1.5 mm; D/V: 4.4 mm from bregma). A microdialysis guide was also aimed just above the parafascicular nucleus (A/P: −4.80 mm; M/L: 1.5 mm; D/V: 4.4 mm from bregma) in animals implanted in the SC. All experiments were carried out in accordance with the European Community Council Directive of 24 November 1986 (86/609/EEC).
After at least a week of recovery and habituation to handling, the electroencephalograms (EEGs) were recorded between 9.00 and 12.00 a.m (left frontal-parietal; right frontal-parietal) in awake freely moving animals using a digital acquisition system (Coherence 3NT, Deltamed, France). The animals were injected bilaterally in a volume of 200 nl/side (i) in the SC with saline or picrotoxin at a dose of 33 pmoles/side (Sigma, France) dissolved in NaCl 0.9 %, as in our previous study (3) or (ii) in the parafascicular nucleus with saline or a low non toxic dose of kainate (35 pmoles/side) dissolved in NaCl 0.9 %. This range of kainate dose is known to activate cell bodies for about 20–30 min, without lesioning effects, upon intracerebral microinjections (9). For each recording session, intracerebral injections were performed after a 20-min reference EEG recording. The EEG recording was then resumed for up to 60 min and the behaviour of the animal was observed.
Dialysis in the Pf was carried out 60 min before and 100 min after injection of picrotoxin or saline in the SC. The microdialysis probe (polycarbonate-polyether, 20-kDa molecular mass cutoff, 2 mm long and 0.52-mm diameter, CMA 12; CMA Microdialysis) was first inserted into the guide and was connected to a microinjection pump (CMA 100; CMA Microdialysis). The perfusion medium contained 147 mM NaCl, 4.0 mM KCl and 2.4 mM CaCl2, pH 6.5, filtered through 0.22 μM Millex Millipore filters before each experiment and was perfused at 1.0 μl/min (20-min collection). The GABA and glutamate contents of the dialysates were determined by fluorimetric detection (Waters fluorimeter 470; excitation, 345 nm; emission, 455 nm) of the o-phthaldehyde derivatives after their separation by HPLC, using a modification of the method of Allison et al. (1984) (10). All chromatographies were performed with a Nucleosil C 18 column (5 m, 25 × 0.4 cm), a Waters pump (model 626), a controller 600 S and a Millenium 32 integrator..
Upon completion of the experiments, the animals were killed with an overdose of nembutal and the brains were removed, cut in 20-μm coronal sections, stained with cresyl violet and each injection site and placement of the dialysis probe were localized with reference to the atlas of Paxinos and Watson (11) in the caudal aspect of the SC or near the Pf, respectively. When an injection or a microdialysis site were respectively located outside the SC or the parafascicular nucleus, the data were not included in the analysis.
The EEG data were expressed as mean ± SEM cumulated absence seizure duration (s) of spike-and-wave discharges (SWD) per 20-min period. Paired comparison versus control conditions (saline) were made using an ANOVA and were confirmed with a non parametric test (Wilcoxon). The microdialysis data were expressed as mean ± SEM of percentage of basal release of the 4 first baseline samples (i.e., before injection) and were compared to control conditions (saline) using a one-way ANOVA followed by a Student-Newman-Keuls as multiple comparisons test.
Results
Experiment 1: Effects of bilateral injection of picrotoxin in the caudal superior colliculus on amino acid release in the parafascicular nucleus
Bilateral injection of picrotoxin (33 pmoles/side; n=4) in the caudal part of the SC significantly decreased cumulated duration of SWD during the first 20 min post injection period as compared to saline injection, as previously reported (3, 12) (results not shown). When Pf microdialysis was performed in these animals, the mean baseline extracellular concentration was 37.8 ± 3.9 pmoles/20 min for glutamate and 3.91 ± 1.75 pmoles/20 min for GABA. Picrotoxin injection in the SC induced a significant increase of extracellular levels of glutamate in the Pf, 40 min post injection (+ 28.8 % of the basal level, p <0.05, Newman-Keuls; figure 1) as compared to saline injection, whereas no change was observed in the extracellular GABA level (figure 1). Since the dead volume of the catheter (20 μl) corresponded to a 20-min collection, the extracellular increase of glutamate observed at 40 min post injection of picrotoxin corresponded to the antiepileptic effect observed 20 min post injection.
Figure 1.

A. Effects of intracollicular injection (arrow) of picrotoxin (33 pmole/side, black diamonds) on the mean ± SEM of percentage of basal release of glutamate (upper graph) and GABA (lower graph) as compared to saline injection (open squares).
* p< 0.05 picrotoxin vs saline, ANOVA, n=4. B. Reconstruction of the injection sites in the caudal superior colliculus on an identified coronal section modified from the atlas of Paxinos and Watson. PAG: periaqueductal grey; SC: superior colliculus. C. Reconstruction of the location of the dialysis probes in the parafascicular nucleus on an identified coronal section modified from the atlas of Paxinos and Watson. PF: n. parafascicularis; VPM: ventroposteromedial nucleus of the thalamus.
Experiment 2: Effects of bilateral injection of kainate in the parafascicular nucleus on spike-and-wave discharges
Bilateral injection of kainate (35 pmole/side; n=8) in the Pf significantly decreased (> 70 %) the cumulated duration of SWD 20 min post-injection as compared to saline injection and to the reference period (p< 0.05 and p<0.0001 respectively, ANOVA; figure 2). At this dose no overt behavioral reaction was observed at any time after the kainate injection. No differences between kainate and saline conditions were observed at 40 and 60 min postinjection. No cell losses were observed in these animals in the region of the Pf following histological controls. When one of the injection sites was located outside the Pf (n=3), injection of KA did not result in significant suppression of SWD (Table 1).
Figure 2.
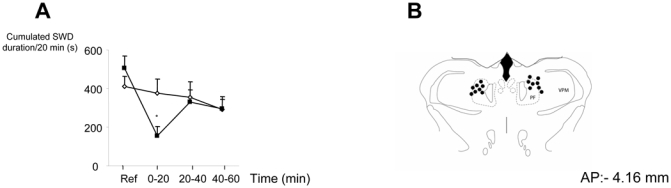
A. Effects of bilateral injection of kainate (35 pmol/side; black squares) into the parafascicular nucleus on mean ± SEM cumulated duration of SWD in GAERS (n=8) *: P< 0.05 as compared to saline (open diamonds), ANOVA.
B. Reconstruction of the injection sites in the parafascicular nucleus on an identified coronal section modified from the atlas of Paxinos and Watson. PF: n. parafascicularis; VPM: ventropostero medial nucleus of the thalamus.
Table 1.
Effects of bilateral injections of kainate (35 pmol/side) outside the parafascicular nucleus on absence seizures in the GAERS. The results are expressed as percent of cumulated duration of spike-and-wave discharges versus control conditions for each 20 min period postinjection. Pf: parafascicular nucleus; VPM: ventral posteromedial nucleus of the thalamus
| Location sites (right/left) | 0–20 min | 21–40 min | 41–60 min |
|---|---|---|---|
| Pf/VPM | 51 | 151 | 125 |
| VPM/Pf | 59 | 135 | 77 |
| No/Pf | 79 | 94 | 41 |
Discussion
The main results of the present study are that (i) disinhibition of the caudal part of the SC which results in suppression of absence seizures is associated with a significant increase of glutamate in the Pf whereas no change of GABA concentration was observed and (ii) direct pharmacological bilateral activation of the Pf suppresses absence seizures. Although preliminary, these data suggest that the glutamatergic projection from the caudal aspect of the SC to the Pf is a critical relay in the output circuit involved in the control of seizures by the basal ganglia via the SNR.
The data of the first experiment of this study confirmed previous results obtained in our laboratory showing that disinhibition of the intermediate layers of the caudal SC suppresses absence seizures without behavioural side effects (3, 12). It is also in agreement with data from the literature showing an antiepileptic effect in different animal models of seizures following activation or disinhibition of the SC (13). The microdialysis data of this experiment suggest that glutamatergic fibers are critically involved in mediating the antiepileptic effects of SC disinhibition. The involvement of these glutamatergic projections is further supported by the time course of the glutamate increase which is similar to the anti-epileptic effects. Little is known about the neurotransmitter nature of the SC-Pf projections (4, 5) and our data suggest that glutamatergic fibers are an important component of this pathway. It is not possible from our study to determine the origin of the GABA measured in the Pf, but our data suggest that GABAergic neurons may not be an important part of the SC-Pf projection. Further experiments are required to confirm the involvement of the glutamatergic neurons in the control of seizures by the disinhibition of the SC.
The second experiment of the present study further confirmed that activation of glutamate receptors in the Pf leads to absence-seizure suppression and is likely to mediate, at least in part, the suppressing effects of SNR inhibition and/or SC disinhibition. In the present study, microinjection of low doses of kainate was used to activate cell bodies, since intracerebral microinjection of this compound have been shown to result in behavioural or physiological effects lasting up to 30 min (9). The time course of suppressive effect on SWD is thus in agreement with data from the literature. In addition, at the dose used, no cell death is observed, especially in brain structures with moderate concentration of kainate receptors, as it is the case for the Pf (14). In our study, no overt loss of cells was observed in the area of the injection. These data remain to be confirmed with other pharmacogical or electrical means of activation. However, they are in agreement with clinical reports showing that electrical stimulation of the centromedian nucleus of the thalamus (which shares similar connections with Pf) results in potent anti-epileptic effects in human patients (7). Whether similar mechanisms are involved in seizure suppression in animal models and human patients and whether this control depends on the type of seizures remain to be determined. However, our data are also in agreement with experimental reports indicating that an increase of activity of intralaminar neurons is associated with the cortical desynchronized activity that characterizes wakefulness (15). Altogether, our results suggest that direct or indirect activation of Pf neurons could reduce the capacity of the cortical neurons to synchronize, thus limiting the occurrence of epileptic discharges.
Neurons of intralaminar thalamic nuclei such as the centrolateral and paracentral nuclei are recruited rythmically during SWDs through mechanisms underlying glutamatergic excitation modulated by GABAergic influences (8). These neurons may be involved in the maintenance of cortico-thalamic oscillations and a change in their excitability, as induced by activation of glutamatergic input or by direct activation of kainate receptor as in the present study, could thus interrupt SWD. Whether this suppressive effect results from a blockage of neuronal synchronization or oscillations in the frequency range of SWD remains to be determined. In addition, although injection of kainate at sites located outside the Pf did not result in significant effects in the present study, we cannot exclude the possibility that SWD suppression results from a change of excitability of adjacent thalamic nuclei. In particular, diffusion of kainate to neurons of the ventro-basal nuclei, known to be critical in the generation of SWD (16), could suppress their oscillating activity and result in seizure suppression. Further studies conducted in our laboratory are currently aimed to determine the specificity of the glutamatergic SC-Pf projection in the control of seizures by the SC and to better understand the mechanisms through which activation of these fibers suppress SWD.
Acknowledgments
The authors want to thank Viviane Hechler for her help in the HPLC analysis. KNB was the recipient of research fellowship from Université Louis Pasteur – CNRS – Eli Lilly.
References
- 1.Gale K. Mechanisms of seizure control mediated by gamma aminobutyric acid: role of the substantia nigra. Fed Proc. 1985;44:2414–2424. [PubMed] [Google Scholar]
- 2.Deransart C, Vercueil L, Marescaux C, Depaulis A. The role of basal ganglia in the control of generalized absence seizures. Epilepsy Res. 1998;32:213–223. doi: 10.1016/s0920-1211(98)00053-9. [DOI] [PubMed] [Google Scholar]
- 3.Nail-Boucherie K, Lê-Pham BT, Marescaux C, Depaulis A. Suppression of absence seizures by electrical and pharmacological activation of the caudal superior colliculus in a genetic model of absence epilepsy in the rat. Exp Neurol. 2002:503–514. doi: 10.1006/exnr.2002.7997. [DOI] [PubMed] [Google Scholar]
- 4.Yamasaki DS, Krauthamer GM, Rhoade RM. Superior collicular projection to intralaminar thalamus in rat. Brain Res. 1986;378:223–233. doi: 10.1016/0006-8993(86)90925-x. [DOI] [PubMed] [Google Scholar]
- 5.Krout KE, Loewy AD, Max Westby GW, Redgrave P. Superior colliculus projections to midline and intralaminar thalamic nuclei of the rat. J Comp Neurol. 2001;431:198–216. doi: 10.1002/1096-9861(20010305)431:2<198::aid-cne1065>3.0.co;2-8. [DOI] [PubMed] [Google Scholar]
- 6.Banerjee PK, Snead OC. Thalamic NMDA receptors in the gamma-hydroxybutyrate model of absence seizures: a cerebral microinjection study in rats. Neuropharmacology. 1995;34:43–53. doi: 10.1016/0028-3908(94)00134-e. [DOI] [PubMed] [Google Scholar]
- 7.Velasco M, Velasco F, Velasco AL. Centromedian-thalamic and hippocampal electrical stimulation for the control of intractable epileptic seizures. J Clin Neurophysiol. 2001;18:6495–513. doi: 10.1097/00004691-200111000-00001. [DOI] [PubMed] [Google Scholar]
- 8.Seidenbecher T, Pape HC. Contribution of intralaminar thalamic nuclei to spike-and-wave-discharges during spontaneous seizures in a genetic rat model of absence epilepsy. Eur J Neurosci. 2001;13:1537–46. doi: 10.1046/j.0953-816x.2001.01537.x. [DOI] [PubMed] [Google Scholar]
- 9.Bandler R, Depaulis A. Elicitation of intraspecific defence reactions in the rat from midbrain periaqueductal grey by microinjection of kainic acid, without neurotoxic effects. Neurosci Lett. 1988;7:291–296. doi: 10.1016/0304-3940(88)90226-1. [DOI] [PubMed] [Google Scholar]
- 10.Allison L, Mayer AGS, Shoup RE. O-Phtalaldehyde derivatives of amines for high-speed liquid chromatography/electrochemistry. Anal Chem. 1984;56:1089–1102. doi: 10.1021/ac00271a010. [DOI] [PubMed] [Google Scholar]
- 11.Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 2. Academic Press; San Diego: 1986. [DOI] [PubMed] [Google Scholar]
- 12.Depaulis A, Liu Z, Vergnes M, Marescaux C, Micheletti G, Warter J. Suppression of spontaneous generalized non-convulsive seizures in the rat by microinjection of GABA antagonists into the superior colliculus. Epilepsy Res. 1990;5:192–198. doi: 10.1016/0920-1211(90)90038-w. [DOI] [PubMed] [Google Scholar]
- 13.Redgrave P, Simkins M, Overton P, Dean P. Anticonvulsant role of nigrotectal projection in the maximal electroshock model of epilepsy - I. Mapping of dorsal midbrain with bicuculline. Neuroscience. 1992;46:379–90. doi: 10.1016/0306-4522(92)90059-b. [DOI] [PubMed] [Google Scholar]
- 14.Petralia RS, Wang YX, Wenthold RJ. Histological and ultrastructural localization of the kainate receptor subunits, KA2 and GlurR6/7, in the rat nervous system using selective antipeptide antibodies. J Comp Neurol. 1994;349:85–110. doi: 10.1002/cne.903490107. [DOI] [PubMed] [Google Scholar]
- 15.Glenn LL, Steriade M. Discharge rate and excitability of cortically projecting intralaminar thalamic neurons during waking and sleep states. J Neurosci. 1982;2:1387–404. doi: 10.1523/JNEUROSCI.02-10-01387.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Crunelli V, Leresche N. Childhood absence epilepsy: genes, channels, neurons and networks. Nat Rev Neurosci. 2002 May;3(5):371–382. doi: 10.1038/nrn811. [DOI] [PubMed] [Google Scholar]