

Key Points
This Review provides an overview on the spike (S) protein of severe acute respiratory syndrome-coronavirus (SARS-CoV) as a target for the development of vaccines and therapeutics for the prevention and treatment of SARS.
SARS is a newly emerging infectious disease, caused by SARS-CoV, a novel coronavirus that caused a global outbreak of SARS.
SARS-CoV S protein mediates binding of the virus with its receptor angiotensin-converting enzyme 2 and promotes the fusion between the viral and host cell membranes and virus entry into the host cell.
SARS-CoV S protein induces humoral and cellular immune responses against SARS-CoV.
SARS S protein is the target of new SARS vaccines. These vaccines are based on SARS-CoV full-length S protein and its receptor-binding domain, including DNA-, viral vector- and subunit-based vaccines
Peptides, antibodies, organic compounds and short interfering RNAs are additional anti-SARS-CoV therapeutics that target the S protein.
The work on SARS-CoV S protein-based vaccines and drugs will be useful as a model for the development of prophylactic strategies and therapies against other viruses with class I fusion proteins that can cause emerging infectious diseases.
The outbreaks of severe acute respiratory syndrome (SARS) between 2002 and 2004 killed hundreds of people. Vaccines against the SARS coronavirus (SARS-CoV) could protect the population during future outbreaks. In this Review, Shibo Jiang and colleagues describe such vaccines, as well as other therapeutics, based on the SARS-CoV spike protein.
Abstract
Severe acute respiratory syndrome (SARS) is a newly emerging infectious disease caused by a novel coronavirus, SARS-coronavirus (SARS-CoV). The SARS-CoV spike (S) protein is composed of two subunits; the S1 subunit contains a receptor-binding domain that engages with the host cell receptor angiotensin-converting enzyme 2 and the S2 subunit mediates fusion between the viral and host cell membranes. The S protein plays key parts in the induction of neutralizing-antibody and T-cell responses, as well as protective immunity, during infection with SARS-CoV. In this Review, we highlight recent advances in the development of vaccines and therapeutics based on the S protein.
Main
Severe acute respiratory syndrome (SARS) was the first new infectious disease identified in the twenty-first century. This acute, and often severe, respiratory illness originated in the Guangdong province of China in November 2002 (ref. 1). A global effort coordinated by WHO led to the identification, in April 2003, of a new coronavirus, SARS-coronavirus (SARS-CoV), as the agent that caused the outbreak2.
SARS-CoV is an enveloped, single and positive-stranded RNA virus2. Its genome RNA encodes a non-structural replicase polyprotein and structural proteins, including spike (S), envelope (E), membrane (M) and nucleocapsid (N) proteins3,4,5. SARS-CoV, a zoonotic virus, resides in hosts that form its natural reservoir, such as bats, but can also infect intermediate hosts, such as small animals (for example, palm civets), before being transmitted to humans6,7,8. SARS-CoV can infect and replicate in several cell types in the human body and causes serious pathological changes (Box 1, Fig 1). A further understanding of the life cycle and pathogenesis of SARS-CoV will help us to develop vaccines and therapeutics to prevent and treat SARS-CoV and SARS-like coronavirus (SL-CoV) infections in the future.
Figure 1. The life cycle of SARS-CoV in host cells.
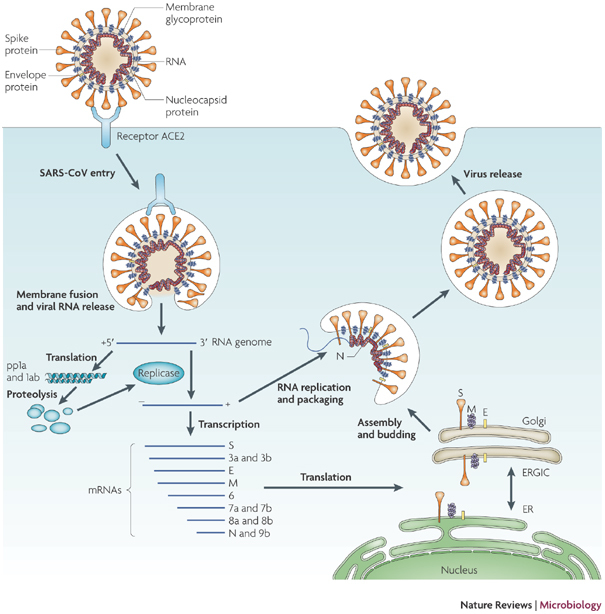
Severe acute respiratory syndrome-coronavirus (SARS-CoV) enters target cells through an endosomal pathway113,121,125,126,127. S protein first binds to the cellular receptor angiotensin-converting enzyme 2 (ACE2)129, and the ACE2–virus complex is then translocated to endosomes, where S protein is cleaved by the endosomal acid proteases (cathepsin L)105 to activate its fusion activity. The viral genome is released and translated into viral replicase polyproteins pp1a and 1ab, which are then cleaved into small products by viral proteinases. Subgenomic negative-strand templates are synthesized from discontinuous transcription on the plus-strand genome and serve as templates for mRNA synthesis. The full-length negative-strand template is made as a template for genomic RNA. Viral nucleocapsids are assembled from genomic RNA and N protein in the cytoplasm, followed by budding into the lumen of the ERGIC (endoplasmic reticulum (ER)–Golgi intermediate compartment)128. Virions are then released from the cell through exocytosis.
After its first occurrence, SARS rapidly spread around the world along international air-travel routes, reaching all five continents and 29 countries, resulting in 8,098 cases and 774 deaths by 23 September 2003 (ref. 9). The overall fatality of SARS is about 10% in the general population, but >50% in patients aged 65 years and older (WHO update 49; see Further information). The global outbreak of SARS was brought under control in July 2003 by effective quarantine, patient-isolation and travel restrictions. Four sporadic SARS cases caused by different SARS-CoV isolates than those that predominated in the 2002–2003 outbreak were reported in late 2003 and early 2004 (refs 10, 11, 12). The most recent epidemic of SARS occurred in Beijing and Anhui in China in April 2004 and originated from laboratory contamination (WHO update 7; see Further information). Since then, no new case of SARS has been reported, possibly because of continued global vigilance and surveillance and laboratory bio-safety practices, as well as the euthanizing or quarantining of animals that may have been exposed to SARS-CoV13,14. Although the outbreaks of SARS seem to be over, SARS is still a safety concern because of the possible reintroduction of a SL-CoV into humans and the risk of an escape of SARS-CoV from laboratories15,16.
Infection with SARS-CoV can trigger a series of humoral and cellular immune responses. Specific antibodies against SARS-CoV (immunoglobulin G (IgG) and IgM) were detectable approximately 2 weeks post-infection, reaching a peak 60 days post-infection and remaining at high levels until 180 days post-infection (ref. 17). High titres of neutralizing antibodies and SARS-CoV-specific cytotoxic T lymphocyte responses were detected in patients who had recovered from SARS18,19, and the levels of the responses correlated well with the disease outcome20. This suggests that both humoral and cellular immune responses are crucial for the clearance of infection by SARS-CoV.
Neutralizing antibodies and/or T-cell immune responses can be raised directly against several SARS-CoV proteins21,22,23, but mainly target the S protein20,24,25,26, suggesting that S protein-induced specific immune responses play important parts in the fight against SARS-CoV infection18. SARS-CoV S protein also has a key role in the ability of SARS-CoV to overcome the species barrier, as adaptive evolution of S protein can contribute to the animal-to-human transmission of SARS-CoV27. Because the S protein of SARS-CoV is involved in receptor recognition, as well as virus attachment and entry, it represents one of the most important targets for the development of SARS vaccines and therapeutics.
Structure of the SARS-CoV S protein
The spikes of SARS-CoV are composed of trimers of S protein, which belongs to a group of class I viral fusion glycoproteins that also includes HIV glycoprotein 160 (Env), influenza haemagglutinin (HA), paramyxovirus F and Ebola virus glycoprotein28. The SARS-CoV S protein encodes a surface glycoprotein precursor that is predicted to be 1,255 amino acids in length, and the amino terminus and most of the protein is predicted to be on the outside of the cell surface or the virus particles3. The predicted S protein consists of a signal peptide (amino acids 1–12) located at the N terminus, an extracellular domain (amino acids 13–1,195), a transmembrane domain (amino acids 1,196–1,215) and an intracellular domain (amino acids 1,216–1,255)29,30,31,32 (Fig. 2a). Similarly to other coronaviruses, the S protein of SARS-CoV can be cleaved into the S1 and S2 subunits by proteases, such as trypsin33, factor Xa34 and cathepsin L35. The trypsin cleavage site occurs at R667–S668 (ref. 36), whereas cathepsin L cleavage is mapped to T678–M679 in the S protein35. Cathepsin L cleaves the S protein of SARS-CoV upstream of, rather than adjacent to, the fusion peptide, and the cleavage is required for activation of the membrane fusion domain of the S protein following entry into target cells35.
Figure 2. SARS-CoV S protein structure and its complex with the receptor ACE2.
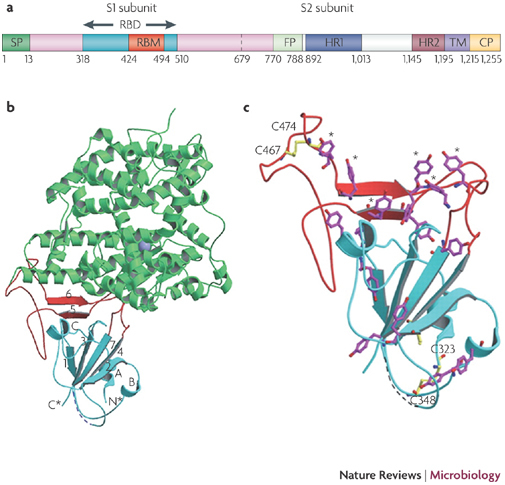
a | Schematic of the S protein29,30,31,32. The residue numbers of each region represent their positions in the S protein of severe acute respiratory syndrome-coronavirus (SARS-CoV). b | Crystal structures of the RBD complexed with the receptor. RBD (the core structure is cyan and the loop RBM is red) interacts with the receptor angiotensin-converting enzyme 2 (ACE2; green). A five-stranded anti-parallel β-sheet (β1–β4 and β7) that connects with three short α-helices (αA–αC) constitutes the core, whereas a two-stranded β-sheet (β5 and β6) forms the loop. N* and C* represent the amino and carboxyl termini of the RBD, respectively. c | The RBD tyrosine (magenta) and cysteine (yellow) residue distribution29. The asterisks represent six ACE2-contacting tyrosines on the RBD, and two disulphide bonds are shown to link C323 to C348 and C467 to C474. CP, cytoplasm domain; FP, fusion peptide; HR, heptad repeat; RBD, receptor-binding domain; RBM, receptor-binding motif; SP, signal peptide; TM, transmembrane domain. Parts b and c are adapted, with permission, from Ref. 29 © (2005) American Association for the Advancement of Science.
Angiotensin-converting enzyme 2 (ACE2) has been identified as the receptor of SARS-CoV37. A fragment that is located in the S1 subunit and spans amino acids 318–510 is the minimal receptor-binding domain (RBD)30,38,39. Crystallographic studies have shown the structure of RBD complexed with its receptor ACE2 (refs 29, 40). During the interaction of RBD with the receptor, RBD presents a concave surface for the N terminus of the receptor peptidase, on which amino acids 445–460 anchor the entire receptor-binding loop of the RBD core (Fig. 2b). This loop (amino acids 424–494 of the RBD), which makes complete contact with the receptor ACE2, was referred to as receptor-binding motif (RBM) (Fig. 2a). The RBM region is tyrosine rich. Among the 14 residues of RBM that are in direct contact with ACE2, six are tyrosine, representing both the hydroxyl group and hydrophobic ring. The RBD region also contains multiple cysteine residues that are linked by disulphide bonds29 (Fig. 2c). Two residues in particular, those at positions 479 and 487, determine SARS disease progression and SARS-CoV tropism (host range)41,42. Any residue changes in these two positions might therefore enhance animal-to-human or human-to-human transmission29.
Human and animal SARS-CoVs depend on ACE2 for cell entry. Animal SARS-CoV could evolve to infect humans by a series of transmission events between animals and humans. For example, a chimeric recombinant SARS-CoV that bears the S protein of civet SARS-CoV (icSZ16-S) can adapt to human airway epithelial cells and displays enhanced affinity for human ACE2 (ref. 43). Changes of only a few residues in the RBD of the civet SARS-CoV S protein, which is responsible for binding with the peptidase domain of ACE2, result in enhanced human ACE2-binding affinity of SARS-CoVs from animals, including civets, mice and rats, facilitating efficient cross-species infections29. However, the SL-CoV from bats does not infect ACE2-expressing cells7,8, suggesting that, unlike SARS-CoVs from human and civets, the SL-CoV from bats does not use ACE2 as a cellular receptor. Thus, the SL-CoV from bats might be the precursor of animal SARS-CoVs, which may act as the intermediates for animal-to-human transmission.
Functions of the SARS-CoV S protein
SARS-CoV S protein has pivotal roles in viral infection and pathogenesis44,45. S1 recognizes and binds to host receptors, and subsequent conformational changes in S2 facilitate fusion between the viral envelope and the host cell membrane30,33.
Receptor binding. The RBD in S1 is responsible for virus binding to host cell receptors30,37,39. ACE2 from SARS-CoV-permissive Vero E6 cells efficiently binds S1, and its soluble form blocks S1 from associating with Vero E6 cells. In addition, SARS-CoV replicates efficiently in ACE2-transfected cells, and anti-ACE2 antibodies block virus entry and replication in Vero E6 cells. This shows that ACE2 is a functional receptor for SARS-CoV37,46,47. A total of 18 residues of ACE2 keep contact with 14 amino acids in the RBD of SARS-CoV S protein29. K341 of ACE2 and R453 of the RBD are important for the complex formation48. N479 and T487 of the RBD are important for the high-affinity association of S protein with ACE2 (ref. 42). A point mutation at R441 or D454 of the RBD disrupts the antigenic structure and binding activity of RBD to ACE2 (refs 30, 49).
SARS-CoV can also bind to host cells through alternative receptors, such as DC-SIGN (dendritic cell-specific intercellular adhesion molecule-3-grabbing non-integrin) and/or L-SIGN (liver/lymph node-SIGN)50,51. Seven asparagine-linked glycosylation sites in the S protein, including residues at positions 109, 118, 119, 158, 227, 589 and 699, are crucial for DC-SIGN- or L-SIGN-mediated virus entry. These residues differ from those of the ACE2-binding domain located at amino acids 318–510 (ref. 52). This would suggest that S protein can also use DC-SIGN or L-SIGN as a receptor, independently of ACE2. However, the actual function of DC-SIGN and L-SIGN needs to be further verified.
Viral fusion. The fusion process that is mediated by S protein of SARS-CoV is similar to that mediated by class I viral fusion proteins of other viruses, such as HIV-1 and murine hepatitis virus (MHV)53,54, but may occur in the acidic environment of the endosomes, rather than on the cell surface. S2 contains heptad repeat 1 (HR1) and HR2 domains, which play an important part in SARS-CoV fusion with target cells. Binding of the RBD of S1 to the receptor ACE2 triggers a conformational change of the S2 from a pre-fusion form to a post-fusion form, resulting in insertion of the putative fusion peptide (amino acids 770–788)31 into the target cell membrane and association of HR1 and HR2 domains to form a six-helix bundle fusion core structure. This brings the viral envelope and target cell membrane into close proximity for fusion. The crystal structure of the SARS-CoV fusion core is described in detail in ref. 55 (Fig. 3). Similarly to the S protein of MHV, but not gp41 of HIV-1, SARS-CoV S protein has a longer HR1 region than HR2 region. The six-helix bundle fusion core has a rod-shaped structure with a length of ∼70 Å and a diameter of ∼28 Å. Three HR1 helices form a parallel trimeric coiled-coil that is surrounded by three HR2 helices in an oblique, antiparallel manner55 (Fig. 3). A synthetic peptide derived from the HR2 region could interact with an HR1 peptide to form a stable six-helix bundle and inhibit SARS-CoV infection in a dose-dependent manner53. Consequently, both the HR1 and HR2 regions in the S2 domains are expected to participate in the viral fusion and entry processes and will serve as attractive targets for the development of anti-SARS-CoV therapeutics and vaccines.
Figure 3. The fusion core structure.
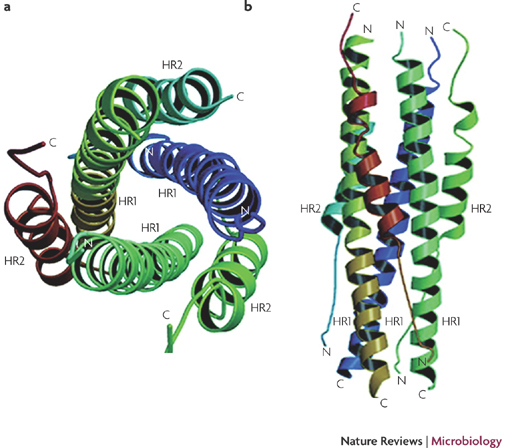
The fusion core is a six-helix bundle with three HR2 α-helices packed in an oblique antiparallel manner against the hydrophobic grooves on the surface of the central HR1 trimer55,130. A top (a) and side (b) view is shown of the severe acute respiratory syndrome-coronavirus (SARS-CoV) S protein six-helix bundle fusion core structure formed by the HR1 and HR2 domains in the S2 subunit. C, carboxyl; N, amino. Figure adapted, with permission, from Ref. 55 © (2004) American Society for Biochemistry and Molecular Biology.
Vaccines based on the SARS-CoV S protein
The roles of S protein in receptor binding and membrane fusion indicate that vaccines based on the S protein could induce antibodies to block virus binding and fusion or neutralize virus infection. Among all structural proteins of SARS-CoV, S protein is the main antigenic component that is responsible for inducing host immune responses, neutralizing antibodies and/or protective immunity against virus infection. S protein has therefore been selected as an important target for vaccine and antiviral development. A comparison of these approaches is provided in Table 1.
Table 1.
S protein-based vaccines and antiviral therapies against SARS-CoV
| Category | Advantages | Disadvantages | Refs |
|---|---|---|---|
| Vaccines * | |||
| Full-length S protein | Induces effective neutralizing-antibody and T-cell responses, as well as protective immunity | Might induce harmful immune responses that cause liver damage or enhanced infection | 64,65 |
| DNA-based | Easier to design; induces immunoglobulin G, neutralizing-antibody and T-cell responses and/or protective immunity | Might have low efficacy in humans; repeated doses may cause toxicity | 59,131 |
| Viral vector-based | Induces neutralizing-antibody responses, protective immunity and/or T-cell responses | Might induce ADE effect; possibly present pre-existing immunity | 60,61,65 |
| Recombinant S protein-based | Induces high neutralizing-antibody responses and protective immunity | Mainly humoral responses; need repeated doses and adjuvants | 62 |
| RBD | Induces highly potent neutralizing-antibody and T-cell responses and protective immunity | Not identified | 70,71,72,73 |
| DNA-based | Induces neutralizing-antibody and T-cell responses and/or protective immunity | Induces low responses; might not neutralize mutants | 132,133,134 |
| Viral vector-based | Induces neutralizing-antibody responses, protective immunity and/or T-cell responses | Possible genomic integration of foreign DNA; viral vector instability | 75,135 |
| Recombinant RBD protein-based | Safer and more effective than other RBD vaccines; induces neutralizing-antibody and T-cell responses, protective immunity and cross protection | Needs repeated doses and adjuvants | 26,70,71,72 |
| Therapeutics * | |||
| Peptides | Inhibits virus infection by preventing S protein-mediated receptor binding and blocking viral fusion and entry | Low antiviral potency | 53,82–84,136,137,138 |
| RBD–ACE2 blockers | Blocks RBD–ACE2 binding and S protein-mediated infection | Not identified | 82,83 |
| S cleavage inhibitors | Might interfere with S cleavage | Not identified | 84,136,137 |
| Fusion core blockers | Easy to design; inhibits virus infection with high specificity | Not identified | 53,89,90,138 |
| Neutralizing antibodies | Highly potent virus inhibition and/or neutralization activity against homologous and heterologous SARS-CoV isolates | Might enhance SARS-CoV entry; further studies needed | 139 |
| Neutralizing mouse antibodies | Easier to generate than human neutralizing antibodies; neutralizes SARS-CoV in vitro and prevents virus replication | Repeated use can cause HAMA response; might not recognize mutants with key substitutions in S protein | 65,94,140,141 |
| Neutralizing human antibodies | Inhibits virus entry, neutralizes virus infection, induces cross protection and reduces disease severity and viral burden; more suitable to development as human immunotherapeutics | Not identified | 97,142,143 |
| Small compounds | Oral bioavailability | Low antiviral potency | 103,104,105,107,108,109 |
| Protease inhibitors | Blocks virus entry and/or inhibits protease (cathepsin L) proteolysis | Not identified | 103,104,105 |
| S protein inhibitors | Specifically inhibits S protein-mediated SARS-CoV fusion and entry into the host cell | Not identified | 107,108,109 |
| Small interfering RNAs | Reduces virus replication and/or silences S gene expression | Low antiviral potency; limited usefulness | 113,114,115,116,117 |
| *All candidates are at the preclinical study stage. ACE2, angiotensin-converting enzyme 2; ADE, antibody-dependent enhancement; HAMA, human–anti-mouse antibody, RBD, receptor-binding domain, SARS-CoV; severe acute respiratory syndrome-coronavirus. | |||
It has been reported that antibodies raised to amino acids 485–625 in S1 or 1,029–1,192 in S2 neutralize infection by SARS-CoV strains (for example, Tor2 and Sin2774) in Vero E6 cells56,57. Vaccination of African green monkeys with an attenuated parainfluenza virus that encodes the full-length S protein of SARS-CoV Urbani strain resulted in the production of S protein-specific neutralizing antibodies, which protected vaccinated monkeys from subsequent homologous SARS-CoV challenge58, suggesting that immunization with the S protein of SARS-CoV is highly effective in the prevention of SARS.
Vaccines based on the full-length S protein. Several vaccines that are based on the full-length S protein of SARS-CoV have been reported. Yang et al.59 showed that a DNA vaccine encoding the full-length S protein SARS-CoV Urbani strain could induce both T-cell and neutralizing-antibody responses, as well as protective immunity, in a mouse model. Other groups have also shown that vaccination of mice or monkeys with highly attenuated modified vaccinia virus Ankara (MVA), which encodes the full-length S protein of the SARS-CoV Urbani strain or HKU39849 strain, elicited S-specific neutralizing antibodies and protective immunity, as evidenced by decreased virus titres in the respiratory tracts of animals after homologous SARS-CoV challenge60,61. Passive transfer of murine serum to naive mice also protected these mice from the challenge of homologous SARS-CoV60,61. In addition, vaccination of mice or hamsters with a full-length S protein trimer protected these animals from infection by homologous SARS-CoV (HKU39849 strain)62. Furthermore, a recombinant baculovirus-expressed full-length S protein of the Urbani strain and its trimer could induce sufficient neutralizing antibodies against human and palm civet SARS pseudoviruses that bore S proteins of homologous and heterologous SARS-CoV variants (for example, Tor2, GD03T13 and SZ3 strains) in vaccinated mice63. These reports suggest that the full-length S protein is highly immunogenic and induces protection against SARS-CoV challenge and that neutralizing antibodies alone may be able to suppress virus proliferation, further justifying the rationale that vaccines can be developed based on the S protein.
Although full-length S protein-based SARS vaccines can induce neutralizing antibody responses against SARS-CoV infection, they may also induce harmful immune responses that cause liver damage of the vaccinated animals or enhanced infection after challenge with homologous SARS-CoV64,65, raising concerns about the safety and ultimate protective efficacy of vaccines that contain the full-length SARS-CoV S protein.
Vaccines based on the RBD. Previous studies have shown that the RBDs of the S proteins of the coronaviruses MHV and HCoV-229E contain major antigenic determinants that can induce neutralizing antibodies66,67. We have discovered that the recombinant RBD (rRBD) antigen of SARS-CoV is highly reactive with the neutralizing antibodies against SARS pseudoviruses that bear S proteins of SARS-CoV (Tor2 strain) in the antisera of mice and rabbits immunized with inactivated SARS-CoV68. The RBD strongly reacts with the antisera from patients with SARS in the convalescent phase, and depletion of the RBD-specific antibodies from patients with SARS results in significant elimination of the neutralizing activity69. Chen et al.61 have also shown that most neutralizing antibodies of antisera of mice, rabbits and monkeys induced by a live-attenuated MVA virus that expressed the full-length S protein could be absorbed and removed by rRBD. Using a fusion protein that contained the RBD linked to human IgG1 Fc fragment (designated RBD-Fc) as an immunogen, we have successfully induced highly potent neutralizing antibodies against SARS-CoV BJ01 strain in immunized rabbits with neutralizing titres greater than 1:10,000 (ref. 70). The antibodies effectively cross-neutralize infection by SARS pseudoviruses that bear S proteins of both homologous and heterologous SARS-CoV isolates, including the representative strains of human 2002–2003 and 2003–2004 SARS-CoV (Tor2 and GD03, respectively) and palm civet SARS-CoV (SZ3)71. Immunization of mice with RBD-Fc induces long-term protective immunity against challenge with homologous SARS-CoV BJ01 strain70,72. Administration of an adeno-associated virus (AAV)-based vaccine that contains RBD (RBD-rAAV) by intramuscular and mucosal pathways elicits sufficient neutralizing antibodies to inhibit homologous SARS-CoV (GZ50) challenge in the established mouse model, and the immune responses can be enhanced by priming with RBD-rAAV and boosting with RBD-specific peptides73,74,75.
The SARS-CoV S protein can also induce CD8+ T-cell responses. One H-2(b)- and one H-2(d)-restricted T-cell epitope are mapped to RBD (S436–S443 and S366–S374, respectively)24. Immunization of mice with a RBD-based subunit vaccine (S318–S510) elicits both antibody and cellular immune responses against SARS-CoV26. The RBD of S protein contains multiple conformation-dependent epitopes and is the main domain that induces neutralizing antibody and T-cell immune responses against SARS-CoV infection76,77, making it an important target for vaccine development. The approaches for developing RBD-based vaccines against SARS-CoV have provided useful information for designing vaccines against other viruses with class I fusion proteins, as these proteins also contain RBDs in their S proteins.
It should be noted that the efficacy of these vaccine candidates is mainly tested in young-mouse and primate animal models. These models are usually less robust, providing virus replication but lacking clinical symptoms and diseases. It is necessary, therefore, to develop more-robust animal models of human diseases for evaluation of vaccine efficacy. Baric and colleagues78,79 have recently reported several lethal SARS-CoV challenge models in BALB/c mice that recapitulated the age-related SARS disease by using recombinant SARS-CoV that bore the S protein of early human and zoonotic strains (GZ02 and HC/SZ/61/03, respectively). They also developed another pathogenic model for young mice after 15 passages of the Urbani isolate in BALB/c mice, which resulted in a lethal virus, MA15, that replicates to high titres in the lungs of mice, causing clinical disease of SARS78,79. Other reports80,81 list examples for the use of senescent mouse models for vaccine evaluation. One candidate vaccine, Venezuelan equine encephalitis virus replicon particles, that expressed the Urbani SARS-CoV S protein partially protected the aged mice from challenge with a recombinant heterologous SARS-CoV that bore epidemic and zoonotic S proteins (icGDO3-S), providing a model to mimic the age-related susceptibility observed in the elder population80. The animal models discussed above can be used as valuable tools to evaluate the efficacy of SARS vaccines.
S protein-based therapeutics
Peptides that interrupt the RBD–ACE2 interaction. It has been shown that rRBD blocks S protein-mediated entry of lentivirus pseudotypes into ACE2-expressing 293T cells with a half maximal inhibitory concentration (IC50) of less than 10 nM30. Similarly, a peptide that overlaps the RBD sequence (amino acids 471–503) blocks the RBD–ACE2 interaction, inhibiting SARS-CoV entry into Vero cells with an IC50 of approximately 40 μM82. A polypeptide that contains two RBD-binding motifs of ACE2 (amino acids 22–44 and 351–357) linked by a glycine exhibits high potent inhibitory activity on SARS pseudovirus infection in ACE2-expressing HeLa cells with an IC50 of 100 nM83. These findings suggest that peptides derived from both RBD and ACE2 that block RBD–ACE2 binding could be developed as novel therapeutics against SARS-CoV infection. However, the in vivo inhibitory activity of these peptides should be evaluated in animal models before considering further development.
Peptides that interfere with the cleavage of S protein. Cleavage of the S protein trimer is an important event in infection, making the potential cleavage site between S1 and S2 domains another target for development of anti-SARS-CoV agents. Synthetic peptides, including P6 (amino acids 598–617) and P8 (amino acids 737–756), both of which are close to the S1–S2 connection and cleavage site, exhibit potent inhibitory activity against the GZ50 strain of SARS-CoV infection in fetal rhesus kidney (FRhK4) cells, and have IC90 values of approximately 100 and 25 μM84. This suggests that binding of the peptides to the S protein interferes with the cleavage of S1 and S2, inhibiting the production of functional S1 and S2 subunits and subsequent fusion of the viral envelope and the host cell membrane. Again, the in vivo antiviral efficacy of these peptides should be tested in animal models.
Peptides that block the HR1–HR2 interaction from forming a fusion-active core. In the early 1990s, Jiang et al.85 and Wild et al.86 discovered the highly potent anti-HIV peptides derived from the HIV-1 gp41 HR2 region. One of the HR2 peptides, T20 (enfuvirtide), was approved by the US Food and Drug Administration for the treatment of patients with HIV or AIDS, especially those who have failed to respond to the current antiretroviral drugs. These HR2 peptides could interact with the viral gp41 HR1 region at fusion-intermediate conformation and block six-helix bundle formation, resulting in the inhibition of HIV fusion at the nanomolar level87,88. Because the SARS-CoV S protein S2 domain also contains HR1 and HR2 sequences, we anticipated that peptides derived from the HR2 region of the SARS-CoV S protein S2 domain would also have antiviral activity against SARS-CoV. We designed and synthesized several peptides that overlapped the HR2 sequence and found that one of these, designated CP-1, could interact with an HR1 peptide to form a stable six-helix bundle and inhibited infection by SARS-CoV WHU strain in Vero E6 cells with an IC50 of approximately 20 μM53. Later, several other research groups also identified anti-SARS-CoV peptides from the S2 domain HR2 region that had viral fusion inhibitory activity at the micromolar level84,89,90. An NMR study has shown that in the pre-fusion intermediate state, the HR2 region forms a symmetric coiled-coil trimer, which has not been observed for other class I viral fusion proteins. The poor antiviral activity of anti-SARS-CoV peptides, compared with the anti-HIV peptides, could be attributed to the tendency of the SARS-CoV S protein HR2 region to form the trimeric coiled-coil. Replacement of the key residues in the HR2 peptide to reduce its ability to form the trimer, but increase its affinity of binding with the HR1 region, to form the six-helix bundle could lead to improvement of its antiviral efficacy. The peptidic antiviral drugs for SARS and other emerging infectious diseases with short incubation periods could have more advantages than the anti-HIV drug enfuvirtide, as enfuvirtide must be injected twice per day for the patient's lifetime. This results in an intolerable injection-site reaction and a high cost to patients, whereas a few injections of the peptidic drugs against SARS-CoV in the early stage of the acute phase could be enough to save patients' lives. One of the disadvantages of using HR2-based peptide inhibitors is the potential selection of escape mutants with altered host-range phenotypes91.
mAbs that target the S protein
Neutralizing mouse mAbs. Using rRBD and inactivated SARS-CoV as immunogens, we have successfully generated a panel of highly potent neutralizing mouse monoclonal antibodies (mAbs) that could block receptor binding and cross-neutralize infection by pseudoviruses that bore S proteins of the representative human SARS-CoV strains that caused the 2002–2003 and 2003–2004 outbreaks (Tor2 and GD03T13) and palm civet SARS-CoV (SZ3)63,69,71,92. Mouse mAbs that target other fragments of the SARS-CoV S protein (for example, amino acids 1,143–1,157) could also effectively inhibit SARS-CoV infection56,93. These neutralizing mouse mAbs can be administered to patients with SARS for early and urgent treatment of SARS-CoV infection9, but cannot be repeatedly used owing to the risk of a human–anti-mouse antibody response. Such a response could rapidly clear the murine antibody from the blood, thus preventing the mouse antibodies from producing the desired therapeutic effect and causing the patient to have an allergic reaction94. Some antibodies against trimeric S protein have the potential to mediate FcγRII-dependent entry into B cells in vitro and thereby cause antibody-dependent enhancement62.
Neutralizing human mAbs. A range of neutralizing human mAbs have been generated from B cells of patients infected with SARS-CoV95,96 or from human immunoglobulin transgenic mice immunized with full-length SARS-CoV S protein97,98,99. These S-specific mAbs, such as 80R and CR3014, could block SARS-CoV S protein binding with the ACE2 receptor and neutralize infection by human SARS-CoV strains Tor2 and HKU39849 and/or palm civet SARS-CoV strain SZ3 (refs 32, 100, 101). mAbs m396 and S230.15 neutralize human SARS-CoV and/or pseudoviruses that bear S proteins of human SARS-CoV strains (Urbani, Tor2 and GD03) and palm civet SARS-CoV strains (SZ3 and SZ16)97. Human anti-S mAbs S109.8, S215.17, S227.14 and S230.15 cross-neutralize infection by a panel of recombinant SARS-CoV strains bearing variant S proteins that are representative of human strains (GZ02, CUHK-W1 and Urbani) and zoonotic strains found in palm civet (HC/SZ/61/03) and raccoon dog (A031G)99. Some human mAbs, such as 80R32, m396 (ref. 97), 201 and 68 (ref. 102), exhibit potent antiviral effects against homologous SARS-CoV challenge in young-mouse replication models. However, others, such as S109.8, S227.14 and S230.15 (ref. 99), could induce broad protection against lethal homologous and heterologous SARS-CoV challenge in both young- and aged-mouse models, providing a strategy to minimize the emergence of mAb escape mutants.
Antiviral compounds and small molecules
Inhibitors of cathepsin L. Cathepsin L activates S protein-mediated membrane fusion by facilitating receptor-dependent and acid-dependent conformational changes in the S2 domain. This occurs in endosomes in which a low pH allows for optimal proteolytic activity35,103,104. Thus, cathepsin L inhibitors, such as E63c, E64d and MDL28170, can block viral entry or inhibit in vitro infection of SARS-CoV or SARS pseudoviruses103,105,106. These findings suggest that compounds which inhibit the activity of cathepsin L protease could be developed as therapeutics for the inhibition of SARS-CoV infection, but their in vivo antiviral activity should be further tested in animal models.
Other compounds and small molecules that target the S protein. Several other compounds and small molecules that target the S protein have been reported. For example, amiodarone blocks the in vitro spread of SARS-CoV by inhibiting virus infection at a post-endosomal level107. Yi et al.108 identified two small molecules, tetra-O-galloyl-beta-D-glucose (TGG) and luteolin, which have inhibitory activity, that blocked SARS-CoV or SARS pseudovirus entry into Vero E6 cells. Kao et al.109 identified 18 small molecules that targeted S protein–ACE2-mediated viral entry. One of these, VE607, exhibits potent inhibitory activity on SARS pseudovirus entry into ACE2-expressing 293T cells. These reports suggest that the small molecules discussed above can function as effective antiviral inhibitors against S protein-mediated viral entry. However, further studies are needed to determine the in vivo efficacy of these antiviral agents in animal models and select optimal formulations to deliver effective concentrations of the drugs to the target tissues.
Gene targeting with small interfering RNA. RNA interference induced by a small interfering RNA (siRNA) has been successfully used recently as a specific and efficient method for silencing specific viral genes, interrupting protein synthesis and suppressing virus replication110,111. It has been demonstrated that siRNAs directed against S sequences of SARS-CoV inhibited SARS-CoV replication in virus-infected Vero E6 cells112. Several research groups113,114,115,116,117 reported that S-specific siRNAs could reduce S protein expression by blocking S mRNA accumulation or reducing the number of copies of the viral genome in FRhK4 cells, indicating that S gene expression in SARS-CoV-infected cells can be effectively silenced by S-specific siRNAs. The in vivo study used a rhesus macaque model to indicate that siRNA duplexes (siSC2–5) that targeted the S protein and ORF1b of SARS-CoV could suppress SARS-like symptoms, inhibit virus replication in the monkey respiratory tract and protect lungs from acute damage118. The findings discussed above reveal the function of siRNA in the inhibition of SARS-CoV infection, replication and/or interruption of S gene expression, raising hopes for the development of effective, novel antiviral agents against SARS-CoV.
Conclusions and prospects
In summary, the S protein of SARS-CoV possesses some unique features that are different from other type I glycoproteins. Many class I fusion proteins, such as HIV Env, influenza HA and MHV S, are post-translationally cleaved at the N-proximal region of the fusion peptide by specific proteases into the surface and transmembrane subunits. By contrast, cleavage of the SARS-CoV S protein may occur far upstream of the predicted fusion peptide (Fig. 2a). Unlike the S proteins of coronaviruses cleaved by furin-like proteases, the S protein of SARS-CoV can be cleaved by cathepsin L at position T678 or by trypsin at R667. In contrast to the entrance mechanism of HIV, SARS-CoV can enter cells from an acidic environment of the endosome119. Nevertheless, SARS-CoV can also enter the target cell surface, which is mediated by proteases on the cell surface through a non-endosomal-dependent pathway120.
The interaction between the SARS-CoV S protein and ACE2 is essential for SARS-CoV entry. The natural evolution of the epidemic SARS-CoV strains probably occurred over a long period, through the repeated transmission of viruses from animals to humans and from humans to animals, resulting in mutations in both the SARS-CoV S protein and ACE2, so that human and animal SARS-CoVs could enter cells that bore human or animal ACE2. Further understanding of the tropism of the virus and the mechanism of the SARS-CoV S protein in receptor binding and entry is therefore important for the development of anti-SARS-CoV therapeutics and vaccines.
As the major component for the development of vaccines against SARS, S protein, and especially the RBD, has been shown to induce highly potent neutralizing antibodies to block virus binding and membrane fusion and/or protective immunity against virus infection. Owing to the absence of human SARS cases in recent years, future SARS epidemics will probably originate from zoonotic transmission. SARS vaccines should therefore protect against not only human SARS-CoV strains, including those from early, middle and late phases of the epidemic, but also those of zoonotic origin. Although current vaccine candidates effectively neutralize SARS-CoV in young-animal replication models without clinical symptoms, they may not protect an elderly population against SARS-CoV infection. Thus, it is essential to test the vaccine candidates in robust lethal-challenge models using aged animals. Future vaccines should effectively protect both the young and the elderly populations from infection by either human or animal SARS-CoV strains that may cause future SARS epidemics.
Peptides and non-peptidic small molecules that target the functional domain of the SARS-CoV S protein, particularly the RBD in the S1 subunit and the HR2 region in the S2 subunit, are mainly virus entry inhibitors and can be further developed as anti-SARS-CoV therapeutics. To develop these molecules as effective and safe antiviral drugs for the treatment of SARS, the urgent task is to improve their potency. Mouse and human mAbs that target the S protein of SARS-CoV have shown potent inhibition and/or neutralization to homologous and heterologous SARS-CoV isolates and can be further developed as immunotherapeutics or passive immunization agents for therapy and prophylaxis of SARS-CoV infection. Future studies are needed to test the in vivo efficacy of these antiviral agents in animal models.
Overall, the feasibility of using peptides and small molecules as anti-SARS therapeutics is partially limited by their low antiviral potency. Furthermore, the possibility of enhancing viral entry might restrict mAbs as immunotherapeutics for long-term use. It is likely, however, that S protein-based vaccines will bear fruit in the near future, as they have been proven to induce long-term and potent neutralizing antibodies and/or protective immunity against SARS-CoV. But the in vivo efficacy of these vaccine candidates in elderly and lethal-challenge models, and their protection against zoonotic virus infection, should be determined before a clinical study is initiated. To take these factors into full consideration, a combination of different strategies with multiple vaccines and antiviral therapeutics may be needed to induce broad and cross protection against various virus strains, especially isolates that have mutated quickly. Early clinical studies that were based on such strategies have been carried out, but it is difficult to push the clinical trials of these candidate vaccines and therapeutics forwards owing to a lack of SARS-CoV-infected subjects and insufficient financial support. Thus, most big pharmaceutical companies have no interest in developing SARS vaccines and therapeutics because of the concern of profitability. However, studies on SARS will provide important information for designing novel strategies for prophylaxis and therapies of other newly emerging infections caused by enveloped viruses with class I fusion proteins.
Box 1 | Pathology of SARS and the life cycle of SARS-CoV infection.
Severe acute respiratory syndrome-coronavirus (SARS-CoV) spreads primarily through droplets (respiratory secretions) and close person-to-person contact. After the virus enters into the body, it binds to primary target cells that express abundant virus receptor, the angiotensin-converting enzyme 2 (ACE2), including pneumocytes and enterocytes in the respiratory system. The virus enters and replicates in these cells. The matured virions are then released to infect new target cells121 (Fig. 1). SARS-CoV can also infect mucosal cells of intestines, tubular epithelial cells of kidneys, epithelial cells of renal tubules, cerebral neurons and immune cells122,123. Infectious viral particles in patients with SARS can be excreted through respiratory secretions, stool, urine and sweat. SARS-CoV infection damages lung tissues owing to elevated levels of production and activation of proinflammatory chemokines and cytokines124, resulting in atypical pneumonia with rapid respiratory deterioration and failure.
Acknowledgements
We thank all three anonymous reviewers for their constructive comments and informative suggestions. Our research was supported by the National Institutes of Health (NIH) of the United States (RO1 AI68002), by the Research Fund for the Control of Infectious Diseases, the Food and Health Bureau of the Hong Kong SAR Government, and by the National 973 Basic Research Program of China (2005CB523001).
Glossary
- Zoonotic virus
A virus that normally exists in vertebrate animals, but can also be transmitted to humans and can cause disease in both animals and humans.
- Adaptive evolution
A process that enables living organisms to cope with environmental stresses and pressures for survival in a new host. For example, under positive selective pressure, civet SARS-CoV can evolve and subsequently adapt to the human host.
- SARS pseudovirus
A synthetic virus that bears the SARS-CoV S protein and contains an Env-defective, luciferase-expressing genome of a retrovirus (for example, HIV), and can infect but does not replicate in cells that express receptors for SARS-CoV.
- Amiodarone
A medication commonly used to treat patients with irregular heart beats or cardiac arrhythmias, including ventricular tachycardia and ventricular fibrillation.
- Luteolin
A flavonoid extracted from Chinese herbs, including Prunella vulgaris and Saussurea lappa Clarks.
Biographies
Lanying Du received her PhD from the University of Hong Kong and is now a research fellow at the Viral Immunology Laboratory, the Lindsley F. Kimball Research Institute of New York Blood Center, USA. Her research interests focus on studies of the structure and function of the severe acute respiratory syndrome-coronavirus (SARS-CoV) spike protein and the development of spike protein-based SARS vaccines.
Yuxian He is an assistant member at the Viral Immunology Laboratory, the Lindsley F. Kimball Research Institute of New York Blood Center, USA, where he has made dedicated efforts to develop vaccines and therapeutics against emerging infectious viruses, including HIV, severe acute respiratory syndrome-coronavirus (SARS-CoV) and highly pathogenic influenza virus H5N1.
Yusen Zhou is Professor and Director of the Department of Pathogen Molecular Biology at the Beijing Institute of Microbiology and Epidemiology, China, where he has focused on studies of viral immunology and pathogenesis of hepatitis and emerging infectious diseases, including severe acute respiratory syndrome (SARS), H5N1 avian influenza and haemorrhagic fever with renal syndrome.
Shuwen Liu is Professor and Executive Director of the Antiviral Research Center, School of Pharmaceutical Sciences of the Southern Medical University, Guangzhou, China, where he has worked on the development of antiviral therapeutics against HIV, severe acute respiratory syndrome-coronavirus (SARS-CoV), influenza virus, hepatitis B virus and hepatitis C virus.
Bo-Jian Zheng is Professor and Director of the Vaccine Laboratory, Department of Microbiology, the University of Hong Kong, where he has worked on the epidemiology, immunology and anti-viral therapy of viral emerging infectious diseases, with a focus on severe acute respiratory syndrome (SARS) and H5N1 avian influenza.
Shibo Jiang is Head of the Viral Immunology Laboratory at the Lindsley F. Kimball Research Institute of the New York Blood Center, USA, where he has focused on the development of envelope glycoprotein-based therapeutics and vaccines against HIV and severe acute respiratory syndrome-coronavirus (SARS-CoV). He was the first to discover the highly potent anti-HIV peptide derived from the HIV-1 glycoprotein 41 HR2 region, and later licensed the patent to Trimeris Pharmaceuticals, who then developed the peptidic anti-HIV drug enfuvirtide, the first HIV fusion inhibitor approved by the US Food and Drug Administration. This discovery opened a new avenue to the development of viral fusion inhibitors against viruses with class I fusion proteins.
Related links
DATABASES
Entrez Genome Project
FURTHER INFORMATION
References
- 1.Zhong NS, et al. Epidemiology and cause of severe acute respiratory syndrome (SARS) in Guangdong, People's Republic of China, in February, 2003. Lancet. 2003;362:1353–1358. doi: 10.1016/S0140-6736(03)14630-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kuiken T, et al. Newly discovered coronavirus as the primary cause of severe acute respiratory syndrome. Lancet. 2003;362:263–270. doi: 10.1016/S0140-6736(03)13967-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Marra MA, et al. The genome sequence of the SARS-associated coronavirus. Science. 2003;300:1399–1404. doi: 10.1126/science.1085953. [DOI] [PubMed] [Google Scholar]
- 4.Peiris JS, et al. Coronavirus as a possible cause of severe acute respiratory syndrome. Lancet. 2003;361:1319–1325. doi: 10.1016/S0140-6736(03)13077-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Rota PA, et al. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science. 2003;300:1394–1399. doi: 10.1126/science.1085952. [DOI] [PubMed] [Google Scholar]
- 6.Guan Y, et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science. 2003;302:276–278. doi: 10.1126/science.1087139. [DOI] [PubMed] [Google Scholar]
- 7.Li W, et al. Bats are natural reservoirs of SARS-like coronaviruses. Science. 2005;310:676–679. doi: 10.1126/science.1118391. [DOI] [PubMed] [Google Scholar]
- 8.Lau SK, et al. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc. Natl Acad. Sci. USA. 2005;102:14040–14045. doi: 10.1073/pnas.0506735102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Skowronski DM, et al. Severe acute respiratory syndrome (SARS): a year in review. Annu. Rev. Med. 2005;56:357–381. doi: 10.1146/annurev.med.56.091103.134135. [DOI] [PubMed] [Google Scholar]
- 10.Che XY, et al. A patient with asymptomatic severe acute respiratory syndrome (SARS) and antigenemia from the 2003–2004 community outbreak of SARS in Guangzhou, China. Clin. Infect. Dis. 2006;43:e1–e5. doi: 10.1086/504943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Fleck F. SARS virus returns to China as scientists race to find effective vaccine. Bull. World Health Organ. 2004;82:152–153. [PMC free article] [PubMed] [Google Scholar]
- 12.Liang G, et al. Laboratory diagnosis of four recent sporadic cases of community-acquired SARS, Guangdong Province, China. Emerg. Infect. Dis. 2004;10:1774–1781. doi: 10.3201/eid1010.040445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Shi Z, Hu Z. A review of studies on animal reservoirs of the SARS coronavirus. Virus Res. 2008;133:74–87. doi: 10.1016/j.virusres.2007.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Liang WN, et al. Severe acute respiratory syndrome — retrospect and lessons of 2004 outbreak in China. Biomed. Environ. Sci. 2006;19:445–451. [PubMed] [Google Scholar]
- 15.Normile D. Infectious diseases. Mounting lab accidents raise SARS fears. Science. 2004;304:659–661. doi: 10.1126/science.304.5671.659. [DOI] [PubMed] [Google Scholar]
- 16.Orellana C. Laboratory-acquired SARS raises worries on biosafety. Lancet Infect. Dis. 2004;4:64. doi: 10.1016/S1473-3099(04)00911-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mo HY, et al. Evaluation by indirect immunofluorescent assay and enzyme linked immunosorbent assay of the dynamic changes of serum antibody responses against severe acute respiratory syndrome coronavirus. Chin. Med. J. 2005;118:446–450. [PubMed] [Google Scholar]
- 18.Xu X, Gao X. Immunological responses against SARS-coronavirus infection in humans. Cell. Mol. Immunol. 2004;1:119–122. [PubMed] [Google Scholar]
- 19.Zhong X, et al. B-cell responses in patients who have recovered from severe acute respiratory syndrome target a dominant site in the S2 domain of the surface spike glycoprotein. J. Virol. 2005;79:3401–3408. doi: 10.1128/JVI.79.6.3401-3408.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Li T, et al. Long-term persistence of robust antibody and cytotoxic T cell responses in recovered patients infected with SARS coronavirus. PLoS ONE. 2006;1:e24. doi: 10.1371/journal.pone.0000024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.See RH, et al. Severe acute respiratory syndrome vaccine efficacy in ferrets: whole killed virus and adenovirus-vectored vaccines. J. Gen. Virol. 2008;89:2136–2146. doi: 10.1099/vir.0.2008/001891-0. [DOI] [PubMed] [Google Scholar]
- 22.Dutta NK, et al. Search for potential target site of nucleocapsid gene for the design of an epitope-based SARS DNA vaccine. Immunol. Lett. 2008;118:65–71. doi: 10.1016/j.imlet.2008.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jin H, et al. Induction of Th1 type response by DNA vaccinations with N, M, and E genes against SARS-CoV in mice. Biochem. Biophys. Res. Commun. 2005;328:979–986. doi: 10.1016/j.bbrc.2005.01.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhi Y, et al. Identification of murine CD8 T cell epitopes in codon-optimized SARS-associated coronavirus spike protein. Virology. 2005;335:34–45. doi: 10.1016/j.virol.2005.01.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Buchholz UJ, et al. Contributions of the structural proteins of severe acute respiratory syndrome coronavirus to protective immunity. Proc. Natl Acad. Sci. USA. 2004;101:9804–9809. doi: 10.1073/pnas.0403492101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zakhartchouk AN, et al. Immunogenicity of a receptor-binding domain of SARS coronavirus spike protein in mice: implications for a subunit vaccine. Vaccine. 2007;25:136–143. doi: 10.1016/j.vaccine.2006.06.084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhang CY, Wei JF, He SH. Adaptive evolution of the spike gene of SARS coronavirus: changes in positively selected sites in different epidemic groups. BMC Microbiol. 2006;6:88. doi: 10.1186/1471-2180-6-88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Weissenhorn W, et al. Structural basis for membrane fusion by enveloped viruses. Mol. Membr. Biol. 1999;16:3–9. doi: 10.1080/096876899294706. [DOI] [PubMed] [Google Scholar]
- 29.Li F, Li W, Farzan M, Harrison SC. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science. 2005;309:1864–1868. doi: 10.1126/science.1116480. [DOI] [PubMed] [Google Scholar]
- 30.Wong SK, Li W, Moore MJ, Choe H, Farzan M. A 193-amino acid fragment of the SARS coronavirus S protein efficiently binds angiotensin-converting enzyme 2. J. Biol. Chem. 2004;279:3197–3201. doi: 10.1074/jbc.C300520200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sainz B, Jr, Rausch JM, Gallaher WR, Garry RF, Wimley WC. Identification and characterization of the putative fusion peptide of the severe acute respiratory syndrome-associated coronavirus spike protein. J. Virol. 2005;79:7195–7206. doi: 10.1128/JVI.79.11.7195-7206.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sui J, et al. Evaluation of human monoclonal antibody 80R for immunoprophylaxis of severe acute respiratory syndrome by an animal study, epitope mapping, and analysis of spike variants. J. Virol. 2005;79:5900–5906. doi: 10.1128/JVI.79.10.5900-5906.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Li F, et al. Conformational states of the severe acute respiratory syndrome coronavirus spike protein ectodomain. J. Virol. 2006;80:6794–6800. doi: 10.1128/JVI.02744-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Du L, et al. Cleavage of spike protein of SARS coronavirus by protease factor Xa is associated with viral infectivity. Biochem. Biophys. Res. Commun. 2007;359:174–179. doi: 10.1016/j.bbrc.2007.05.092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Bosch BJ, Bartelink W, Rottier PJ. Cathepsin L functionally cleaves the SARS-CoV class I fusion protein upstream of rather than adjacent to the fusion peptide. J. Virol. 2008;82:8887–8890. doi: 10.1128/JVI.00415-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lai L, et al. Quaternary structure, substrate selectivity and inhibitor design for SARS 3C-like proteinase. Curr. Pharm. Des. 2006;12:4555–4564. doi: 10.2174/138161206779010396. [DOI] [PubMed] [Google Scholar]
- 37.Li W, et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003;426:450–454. doi: 10.1038/nature02145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Xiao X, Chakraborti S, Dimitrov AS, Gramatikoff K, Dimitrov DS. The SARS-CoV S glycoprotein: expression and functional characterization. Biochem. Biophys. Res. Commun. 2003;312:1159–1164. doi: 10.1016/j.bbrc.2003.11.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Babcock GJ, Esshaki DJ, Thomas WD, Jr, Ambrosino DM. Amino acids 270 to 510 of the severe acute respiratory syndrome coronavirus spike protein are required for interaction with receptor. J. Virol. 2004;78:4552–4560. doi: 10.1128/JVI.78.9.4552-4560.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Prabakaran P, et al. Structure of severe acute respiratory syndrome coronavirus receptor-binding domain complexed with neutralizing antibody. J. Biol. Chem. 2006;281:15829–15836. doi: 10.1074/jbc.M600697200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Qu XX, et al. Identification of two critical amino acid residues of the severe acute respiratory syndrome coronavirus spike protein for its variation in zoonotic tropism transition via a double substitution strategy. J. Biol. Chem. 2005;280:29588–29595. doi: 10.1074/jbc.M500662200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Li W, et al. Receptor and viral determinants of SARS-coronavirus adaptation to human ACE2. EMBO J. 2005;24:1634–1643. doi: 10.1038/sj.emboj.7600640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sheahan T, Rockx B, Donaldson E, Corti D, Baric R. Pathways of cross-species transmission of synthetically reconstructed zoonotic severe acute respiratory syndrome coronavirus. J. Virol. 2008;82:8721–8732. doi: 10.1128/JVI.00818-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Holmes KV. SARS-associated coronavirus. N. Engl. J. Med. 2003;348:1948–1951. doi: 10.1056/NEJMp030078. [DOI] [PubMed] [Google Scholar]
- 45.Hofmann H, et al. S protein of severe acute respiratory syndrome-associated coronavirus mediates entry into hepatoma cell lines and is targeted by neutralizing antibodies in infected patients. J. Virol. 2004;78:6134–6142. doi: 10.1128/JVI.78.12.6134-6142.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kuhn JH, Li W, Choe H, Farzan M. Angiotensin-converting enzyme 2: a functional receptor for SARS coronavirus. Cell. Mol. Life Sci. 2004;61:2738–2743. doi: 10.1007/s00018-004-4242-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Prabakaran P, Xiao X, Dimitrov DS. A model of the ACE2 structure and function as a SARS-CoV receptor. Biochem. Biophys. Res. Commun. 2004;314:235–241. doi: 10.1016/j.bbrc.2003.12.081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zhang Y, Zheng N, Hao P, Cao Y, Zhong Y. A molecular docking model of SARS-CoV S1 protein in complex with its receptor, human ACE2. Comput. Biol. Chem. 2005;29:254–257. doi: 10.1016/j.compbiolchem.2005.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.He Y, Li J, Jiang S. A single amino acid substitution (R441A) in the receptor-binding domain of SARS coronavirus spike protein disrupts the antigenic structure and binding activity. Biochem. Biophys. Res. Commun. 2006;344:106–113. doi: 10.1016/j.bbrc.2006.03.139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Jeffers SA, et al. CD209L (L-SIGN) is a receptor for severe acute respiratory syndrome coronavirus. Proc. Natl Acad. Sci. USA. 2004;101:15748–15753. doi: 10.1073/pnas.0403812101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yang ZY, et al. pH-dependent entry of severe acute respiratory syndrome coronavirus is mediated by the spike glycoprotein and enhanced by dendritic cell transfer through DC-SIGN. J. Virol. 2004;78:5642–5650. doi: 10.1128/JVI.78.11.5642-5650.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Han DP, Lohani M, Cho MW. Specific asparagine-linked glycosylation sites are critical for DC-SIGN- and L-SIGN-mediated severe acute respiratory syndrome coronavirus entry. J. Virol. 2007;81:12029–12039. doi: 10.1128/JVI.00315-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Liu S, et al. Interaction between heptad repeat 1 and 2 regions in spike protein of SARS-associated coronavirus: implications for virus fusogenic mechanism and identification of fusion inhibitors. Lancet. 2004;363:938–947. doi: 10.1016/S0140-6736(04)15788-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Tripet B, et al. Structural characterization of the SARS-coronavirus spike S fusion protein core. J. Biol. Chem. 2004;279:20836–20849. doi: 10.1074/jbc.M400759200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Xu Y, et al. Crystal structure of severe acute respiratory syndrome coronavirus spike protein fusion core. J. Biol. Chem. 2004;279:49414–49419. doi: 10.1074/jbc.M408782200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Zhou T, et al. An exposed domain in the severe acute respiratory syndrome coronavirus spike protein induces neutralizing antibodies. J. Virol. 2004;78:7217–7226. doi: 10.1128/JVI.78.13.7217-7226.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Keng CT, et al. Amino acids 1055 to 1192 in the S2 region of severe acute respiratory syndrome coronavirus S protein induce neutralizing antibodies: implications for the development of vaccines and antiviral agents. J. Virol. 2005;79:3289–3296. doi: 10.1128/JVI.79.6.3289-3296.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Bukreyev A, et al. Mucosal immunisation of African green monkeys (Cercopithecus aethiops) with an attenuated parainfluenza virus expressing the SARS coronavirus spike protein for the prevention of SARS. Lancet. 2004;363:2122–2127. doi: 10.1016/S0140-6736(04)16501-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Yang ZY, et al. A DNA vaccine induces SARS coronavirus neutralization and protective immunity in mice. Nature. 2004;428:561–564. doi: 10.1038/nature02463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Bisht H, et al. Severe acute respiratory syndrome coronavirus spike protein expressed by attenuated vaccinia virus protectively immunizes mice. Proc. Natl Acad. Sci. USA. 2004;101:6641–6646. doi: 10.1073/pnas.0401939101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Chen Z, et al. Recombinant modified vaccinia virus Ankara expressing the spike glycoprotein of severe acute respiratory syndrome coronavirus induces protective neutralizing antibodies primarily targeting the receptor binding region. J. Virol. 2005;79:2678–2688. doi: 10.1128/JVI.79.5.2678-2688.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kam YW, et al. Antibodies against trimeric S glycoprotein protect hamsters against SARS-CoV challenge despite their capacity to mediate FcγRII-dependent entry into B cells in vitro. Vaccine. 2007;25:729–740. doi: 10.1016/j.vaccine.2006.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.He Y, Li J, Heck S, Lustigman S, Jiang S. Antigenic and immunogenic characterization of recombinant baculovirus-expressed severe acute respiratory syndrome coronavirus spike protein: implication for vaccine design. J. Virol. 2006;80:5757–5767. doi: 10.1128/JVI.00083-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Czub M, Weingartl H, Czub S, He R, Cao J. Evaluation of modified vaccinia virus Ankara based recombinant SARS vaccine in ferrets. Vaccine. 2005;23:2273–2279. doi: 10.1016/j.vaccine.2005.01.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Weingartl H, et al. Immunization with modified vaccinia virus Ankara-based recombinant vaccine against severe acute respiratory syndrome is associated with enhanced hepatitis in ferrets. J. Virol. 2004;78:12672–12676. doi: 10.1128/JVI.78.22.12672-12676.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Bonavia A, Zelus BD, Wentworth DE, Talbot PJ, Holmes KV. Identification of a receptor-binding domain of the spike glycoprotein of human coronavirus HCoV-229E. J. Virol. 2003;77:2530–2538. doi: 10.1128/JVI.77.4.2530-2538.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Kubo H, Yamada YK, Taguchi F. Localization of neutralizing epitopes and the receptor-binding site within the amino-terminal 330 amino acids of the murine coronavirus spike protein. J. Virol. 1994;68:5403–5410. doi: 10.1128/jvi.68.9.5403-5410.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.He Y, Zhou Y, Siddiqui P, Jiang S. Inactivated SARS-CoV vaccine elicits high titers of spike protein-specific antibodies that block receptor binding and virus entry. Biochem. Biophys. Res. Commun. 2004;325:445–452. doi: 10.1016/j.bbrc.2004.10.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.He Y, et al. Identification of a critical neutralization determinant of severe acute respiratory syndrome (SARS)-associated coronavirus: importance for designing SARS vaccines. Virology. 2005;334:74–82. doi: 10.1016/j.virol.2005.01.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.He Y, et al. Receptor-binding domain of SARS-CoV spike protein induces highly potent neutralizing antibodies: implication for developing subunit vaccine. Biochem. Biophys. Res. Commun. 2004;324:773–781. doi: 10.1016/j.bbrc.2004.09.106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.He Y, et al. Cross-neutralization of human and palm civet severe acute respiratory syndrome coronaviruses by antibodies targeting the receptor-binding domain of spike protein. J. Immunol. 2006;176:6085–6092. doi: 10.4049/jimmunol.176.10.6085. [DOI] [PubMed] [Google Scholar]
- 72.Du L, et al. Receptor-binding domain of SARS-CoV spike protein induces long-term protective immunity in an animal model. Vaccine. 2007;25:2832–2838. doi: 10.1016/j.vaccine.2006.10.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Du L, et al. Priming with rAAV encoding RBD of SARS-CoV S protein and boosting with RBD-specific peptides for T cell epitopes elevated humoral and cellular immune responses against SARS-CoV infection. Vaccine. 2008;26:1644–1651. doi: 10.1016/j.vaccine.2008.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Du L, et al. Recombinant adeno-associated virus expressing the receptor-binding domain of severe acute respiratory syndrome coronavirus S protein elicits neutralizing antibodies: implication for developing SARS vaccines. Virology. 2006;353:6–16. doi: 10.1016/j.virol.2006.03.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Du L, et al. Intranasal vaccination of recombinant adeno-associated virus encoding receptor-binding domain of severe acute respiratory syndrome coronavirus (SARS-CoV) spike protein induces strong mucosal immune responses and provides long-term protection against SARS-CoV infection. J. Immunol. 2008;180:948–956. doi: 10.4049/jimmunol.180.2.948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Qin C, et al. An animal model of SARS produced by infection of Macaca mulatta with SARS coronavirus. J. Pathol. 2005;206:251–259. doi: 10.1002/path.1769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.He Y, et al. Identification and characterization of novel neutralizing epitopes in the receptor-binding domain of SARS-CoV spike protein: revealing the critical antigenic determinants in inactivated SARS-CoV vaccine. Vaccine. 2006;24:5498–5508. doi: 10.1016/j.vaccine.2006.04.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Rockx B, et al. Synthetic reconstruction of zoonotic and early human severe acute respiratory syndrome coronavirus isolates that produce fatal disease in aged mice. J. Virol. 2007;81:7410–7423. doi: 10.1128/JVI.00505-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Roberts A, et al. A mouse-adapted SARS-coronavirus causes disease and mortality in BALB/c mice. PLoS Pathog. 2007;3:e5. doi: 10.1371/journal.ppat.0030005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Deming D, et al. Vaccine efficacy in senescent mice challenged with recombinant SARS-CoV bearing epidemic and zoonotic spike variants. PLoS Med. 2006;3:e525. doi: 10.1371/journal.pmed.0030525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Vogel LN, et al. Utility of the aged BALB/c mouse model to demonstrate prevention and control strategies for severe acute respiratory syndrome coronavirus (SARS-CoV) Vaccine. 2007;25:2173–2179. doi: 10.1016/j.vaccine.2006.11.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Hu H, et al. Screening and identification of linear B-cell epitopes and entry-blocking peptide of severe acute respiratory syndrome (SARS)-associated coronavirus using synthetic overlapping peptide library. J. Comb. Chem. 2005;7:648–656. doi: 10.1021/cc0500607. [DOI] [PubMed] [Google Scholar]
- 83.Han DP, Penn-Nicholson A, Cho MW. Identification of critical determinants on ACE2 for SARS-CoV entry and development of a potent entry inhibitor. Virology. 2006;350:15–25. doi: 10.1016/j.virol.2006.01.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zheng BJ, et al. Synthetic peptides outside the spike protein heptad repeat regions as potent inhibitors of SARS-associated coronavirus. Antivir. Ther. 2005;10:393–403. [PubMed] [Google Scholar]
- 85.Jiang S, Lin K, Strick N, Neurath AR. HIV-1 inhibition by a peptide. Nature. 1993;365:113. doi: 10.1038/365113a0. [DOI] [PubMed] [Google Scholar]
- 86.Wild CT, Shugars DC, Greenwell TK, Mcdanal CB, Matthews TJ. Peptides corresponding to a predictive alpha-helical domain of human immunodeficiency virus type 1 gp41 are potent inhibitors of virus infection. Proc. Natl Acad. Sci. USA. 1994;91:9770–9774. doi: 10.1073/pnas.91.21.9770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Liu S, Wu S, Jiang S. HIV entry inhibitors targeting gp41: from polypeptides to small-molecule compounds. Curr. Pharm. Des. 2007;13:143–162. doi: 10.2174/138161207779313722. [DOI] [PubMed] [Google Scholar]
- 88.Chan DC, Kim PS. HIV entry and its inhibition. Cell. 1998;93:681–684. doi: 10.1016/S0092-8674(00)81430-0. [DOI] [PubMed] [Google Scholar]
- 89.Bosch BJ, et al. Severe acute respiratory syndrome coronavirus (SARS-CoV) infection inhibition using spike protein heptad repeat-derived peptides. Proc. Natl Acad. Sci. USA. 2004;101:8455–8460. doi: 10.1073/pnas.0400576101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Yuan K, et al. Suppression of SARS-CoV entry by peptides corresponding to heptad regions on spike glycoprotein. Biochem. Biophys. Res. Commun. 2004;319:746–752. doi: 10.1016/j.bbrc.2004.05.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Mcroy WC, Baric RS. Amino acid substitutions in the S2 subunit of mouse hepatitis virus variant V51 encode determinants of host range expansion. J. Virol. 2008;82:1414–1424. doi: 10.1128/JVI.01674-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.He Y, Lu H, Siddiqui P, Zhou Y, Jiang S. Receptor-binding domain of severe acute respiratory syndrome coronavirus spike protein contains multiple conformation-dependent epitopes that induce highly potent neutralizing antibodies. J. Immunol. 2005;174:4908–4915. doi: 10.4049/jimmunol.174.8.4908. [DOI] [PubMed] [Google Scholar]
- 93.Lai SC, et al. Characterization of neutralizing monoclonal antibodies recognizing a 15-residues epitope on the spike protein HR2 region of severe acute respiratory syndrome coronavirus (SARS-CoV) J. Biomed. Sci. 2005;12:711–727. doi: 10.1007/s11373-005-9004-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Welt S, Ritter G. Antibodies in the therapy of colon cancer. Semin. Oncol. 1999;26:683–690. [PubMed] [Google Scholar]
- 95.Nie Y, et al. Neutralizing antibodies in patients with severe acute respiratory syndrome-associated coronavirus infection. J. Infect. Dis. 2004;190:1119–1126. doi: 10.1086/423286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Traggiai E, et al. An efficient method to make human monoclonal antibodies from memory B cells: potent neutralization of SARS coronavirus. Nature Med. 2004;10:871–875. doi: 10.1038/nm1080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Zhu Z, et al. Potent cross-reactive neutralization of SARS coronavirus isolates by human monoclonal antibodies. Proc. Natl Acad. Sci. USA. 2007;104:12123–12128. doi: 10.1073/pnas.0701000104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Coughlin M, et al. Generation and characterization of human monoclonal neutralizing antibodies with distinct binding and sequence features against SARS coronavirus using XenoMouse. Virology. 2007;361:93–102. doi: 10.1016/j.virol.2006.09.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Rockx B, et al. Structural basis for potent cross-neutralizing human monoclonal antibody protection against lethal human and zoonotic severe acute respiratory syndrome coronavirus challenge. J. Virol. 2008;82:3220–3235. doi: 10.1128/JVI.02377-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Sui J, et al. Potent neutralization of severe acute respiratory syndrome (SARS) coronavirus by a human mAb to S1 protein that blocks receptor association. Proc. Natl Acad. Sci. USA. 2004;101:2536–2541. doi: 10.1073/pnas.0307140101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Van Den Brink EN, et al. Molecular and biological characterization of human monoclonal antibodies binding to the spike and nucleocapsid proteins of severe acute respiratory syndrome coronavirus. J. Virol. 2005;79:1635–1644. doi: 10.1128/JVI.79.3.1635-1644.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Greenough TC, et al. Pneumonitis and multi-organ system disease in common marmosets (Callithrix jacchus) infected with the severe acute respiratory syndrome-associated coronavirus. Am. J. Pathol. 2005;167:455–463. doi: 10.1016/S0002-9440(10)62989-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Huang IC, et al. SARS coronavirus, but not human coronavirus NL63, utilizes cathepsin L to infect ACE2-expressing cells. J. Biol. Chem. 2006;281:3198–3203. doi: 10.1074/jbc.M508381200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Huang IC, et al. SARS-CoV, but not HCoV-NL63, utilizes cathepsins to infect cells: viral entry. Adv. Exp. Med. Biol. 2006;581:335–338. doi: 10.1007/978-0-387-33012-9_60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Simmons G, et al. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proc. Natl Acad. Sci. USA. 2005;102:11876–11881. doi: 10.1073/pnas.0505577102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Simmons G, Rennekamp AJ, Bates P. Proteolysis of SARS-associated coronavirus spike glycoprotein. Adv. Exp. Med. Biol. 2006;581:235–240. doi: 10.1007/978-0-387-33012-9_39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Stadler K, et al. Amiodarone alters late endosomes and inhibits SARS coronavirus infection at a post-endosomal level. Am. J. Respir. Cell Mol. Biol. 2008;39:142–149. doi: 10.1165/rcmb.2007-0217OC. [DOI] [PubMed] [Google Scholar]
- 108.Yi L, et al. Small molecules blocking the entry of severe acute respiratory syndrome coronavirus into host cells. J. Virol. 2004;78:11334–11339. doi: 10.1128/JVI.78.20.11334-11339.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Kao RY, et al. Identification of novel small-molecule inhibitors of severe acute respiratory syndrome-associated coronavirus by chemical genetics. Chem. Biol. 2004;11:1293–1299. doi: 10.1016/j.chembiol.2004.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Li T, et al. siRNA targeting the leader sequence of SARS-CoV inhibits virus replication. Gene Ther. 2005;12:751–761. doi: 10.1038/sj.gt.3302479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Gitlin L, Karelsky S, Andino R. Short interfering RNA confers intracellular antiviral immunity in human cells. Nature. 2002;418:430–434. doi: 10.1038/nature00873. [DOI] [PubMed] [Google Scholar]
- 112.Wu CJ, Huang HW, Liu CY, Hong CF, Chan YL. Inhibition of SARS-CoV replication by siRNA. Antiviral Res. 2005;65:45–48. doi: 10.1016/j.antiviral.2004.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Akerstrom S, Mirazimi A, Tan YJ. Inhibition of SARS-CoV replication cycle by small interference RNAs silencing specific SARS proteins, 7a/7b, 3a/3b and S. Antiviral Res. 2007;73:219–227. doi: 10.1016/j.antiviral.2006.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.He ML, et al. Kinetics and synergistic effects of siRNAs targeting structural and replicase genes of SARS-associated coronavirus. FEBS Lett. 2006;580:2414–2420. doi: 10.1016/j.febslet.2006.03.066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Zheng BJ, et al. Prophylactic and therapeutic effects of small interfering RNA targeting SARS-coronavirus. Antivir. Ther. 2004;9:365–374. [PubMed] [Google Scholar]
- 116.Qin ZL, et al. Silencing of SARS-CoV spike gene by small interfering RNA in HEK 293T cells. Biochem. Biophys. Res. Commun. 2004;324:1186–1193. doi: 10.1016/j.bbrc.2004.09.180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Zhang Y, et al. Silencing SARS-CoV spike protein expression in cultured cells by RNA interference. FEBS Lett. 2004;560:141–146. doi: 10.1016/S0014-5793(04)00087-0. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 118.Li BJ, et al. Using siRNA in prophylactic and therapeutic regimens against SARS coronavirus in rhesus macaque. Nature Med. 2005;11:944–951. doi: 10.1038/nm1280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Simmons G, et al. Characterization of severe acute respiratory syndrome-associated coronavirus (SARS-CoV) spike glycoprotein-mediated viral entry. Proc. Natl Acad. Sci. USA. 2004;101:4240–4245. doi: 10.1073/pnas.0306446101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Matsuyama S, Ujike M, Morikawa S, Tashiro M, Taguchi F. Protease-mediated enhancement of severe acute respiratory syndrome coronavirus infection. Proc. Natl Acad. Sci. USA. 2005;102:12543–12547. doi: 10.1073/pnas.0503203102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Qinfen Z, et al. The life cycle of SARS coronavirus in Vero E6 cells. J. Med. Virol. 2004;73:332–337. doi: 10.1002/jmv.20095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Guo Y, Korteweg C, Mcnutt MA, Gu J. Pathogenetic mechanisms of severe acute respiratory syndrome. Virus Res. 2008;133:4–12. doi: 10.1016/j.virusres.2007.01.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Gu J, Korteweg C. Pathology and pathogenesis of severe acute respiratory syndrome. Am. J. Pathol. 2007;170:1136–1147. doi: 10.2353/ajpath.2007.061088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Ding Y, et al. Organ distribution of severe acute respiratory syndrome (SARS) associated coronavirus (SARS-CoV) in SARS patients: implications for pathogenesis and virus transmission pathways. J. Pathol. 2004;203:622–630. doi: 10.1002/path.1560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Frieman M, Heise M, Baric R. SARS coronavirus and innate immunity. Virus Res. 2008;133:101–112. doi: 10.1016/j.virusres.2007.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Weiss SR, Navas-Martin S. Coronavirus pathogenesis and the emerging pathogen severe acute respiratory syndrome coronavirus. Microbiol. Mol. Biol. Rev. 2005;69:635–664. doi: 10.1128/MMBR.69.4.635-664.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Holmes KV. SARS coronavirus: a new challenge for prevention and therapy. J. Clin. Invest. 2003;111:1605–1609. doi: 10.1172/JCI18819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Stertz S, et al. The intracellular sites of early replication and budding of SARS-coronavirus. Virology. 2007;361:304–315. doi: 10.1016/j.virol.2006.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Inoue Y, et al. Clathrin-dependent entry of severe acute respiratory syndrome coronavirus into target cells expressing ACE2 with the cytoplasmic tail deleted. J. Virol. 2007;81:8722–8729. doi: 10.1128/JVI.00253-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Hakansson-Mcreynolds S, Jiang S, Rong L, Caffrey M. Solution structure of the severe acute respiratory syndrome-coronavirus heptad repeat 2 domain in the prefusion state. J. Biol. Chem. 2006;281:11965–11971. doi: 10.1074/jbc.M601174200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Zhao P, et al. DNA vaccine of SARS-CoV S gene induces antibody response in mice. Acta Biochim. Biophys. Sin. (Shanghai) 2004;36:37–41. doi: 10.1093/abbs/36.1.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Wang S, et al. Identification of two neutralizing regions on the severe acute respiratory syndrome coronavirus spike glycoprotein produced from the mammalian expression system. J. Virol. 2005;79:1906–1910. doi: 10.1128/JVI.79.3.1906-1910.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Huang J, et al. Priming with SARS-CoV S DNA and boosting with SARS-CoV S epitopes specific for CD4+ and CD8+ T cells promote cellular immune responses. Vaccine. 2007;25:6981–6991. doi: 10.1016/j.vaccine.2007.06.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Liu L, et al. Natural mutations in the receptor binding domain of spike glycoprotein determine the reactivity of cross-neutralization between palm civet coronavirus and severe acute respiratory syndrome coronavirus. J. Virol. 2007;81:4694–4700. doi: 10.1128/JVI.02389-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Taylor DR. Obstacles and advances in SARS vaccine development. Vaccine. 2006;24:863–871. doi: 10.1016/j.vaccine.2005.08.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Kliger Y, Levanon EY, Gerber D. From genome to antivirals: SARS as a test tube. Drug Discov. Today. 2005;10:345–352. doi: 10.1016/S1359-6446(04)03320-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Wu XD, et al. The spike protein of severe acute respiratory syndrome (SARS) is cleaved in virus infected Vero-E6 cells. Cell Res. 2004;14:400–406. doi: 10.1038/sj.cr.7290240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Ujike M, et al. Heptad repeat-derived peptides block protease-mediated direct entry from the cell surface of severe acute respiratory syndrome coronavirus but not entry via the endosomal pathway. J. Virol. 2008;82:588–592. doi: 10.1128/JVI.01697-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Yang ZY, et al. Evasion of antibody neutralization in emerging severe acute respiratory syndrome coronaviruses. Proc. Natl Acad. Sci. USA. 2005;102:797–801. doi: 10.1073/pnas.0409065102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Subbarao K, et al. Prior infection and passive transfer of neutralizing antibody prevent replication of severe acute respiratory syndrome coronavirus in the respiratory tract of mice. J. Virol. 2004;78:3572–3577. doi: 10.1128/JVI.78.7.3572-3577.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Mitsuki YY, et al. A single amino acid substitution in the S1 and S2 spike protein domains determines the neutralization escape phenotype of SARS-CoV. Microbes Infect. 2008;10:908–915. doi: 10.1016/j.micinf.2008.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Ter Meulen J, et al. Human monoclonal antibody as prophylaxis for SARS coronavirus infection in ferrets. Lancet. 2004;363:2139–2141. doi: 10.1016/S0140-6736(04)16506-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Roberts A, et al. Therapy with a severe acute respiratory syndrome-associated coronavirus-neutralizing human monoclonal antibody reduces disease severity and viral burden in golden Syrian hamsters. J. Infect. Dis. 2006;193:685–692. doi: 10.1086/500143. [DOI] [PMC free article] [PubMed] [Google Scholar]