

Abstract
Social species, from Drosophila melanogaster to Homo sapiens, fare poorly when isolated. Homo sapiens, an irrepressibly meaning-making species, are, in normal circumstances, dramatically affected by perceived social isolation. Research indicates that perceived social isolation (i.e., loneliness) is a risk factor for, and may contribute to, poorer overall cognitive performance, faster cognitive decline, poorer executive functioning, more negativity and depressive cognition, heightened sensitivity to social threats, a confirmatory bias in social cognition that is self-protective and paradoxically self-defeating, heightened anthropomorphism, and contagion that threatens social cohesion. These differences in attention and cognition impact emotions, decisions, behaviors, and interpersonal interactions that may contribute to the association between loneliness and cognitive decline and between loneliness and morbidity more generally.
Introduction
The health, life, and genetic legacy of members of social species are threatened when they finds themselves on the social perimeter. Social isolation decreases lifespan in the fruit fly1; promotes obesity and Type 2 diabetes in mice2; exacerbates infarct size and edema and decreases post-stroke survival rate following experimentally induced stroke in mice3; promotes activation of the sympatho-adrenomedullary response to an acute immobilization or cold stressor in rats4; delays the effects of exercise on adult neurogenesis in rats5; decreases open field activity, increases basal cortisol concentrations, and decreases lymphocyte proliferation to mitogens in pigs6; increases the 24 hr urinary catecholamines levels and evidence of oxidative stress in the aortic arch of rabbits7; and decreases the expression of genes regulating glucocorticoid response in the frontal cortex of piglets.8 Humans, born to the longest period of abject dependency of any species and dependent on conspecifics across the lifespan to survive and prosper, do not fare well, either, whether they live solitary lives or they simply perceive they live in relative isolation (see Box 1).
Box 1. Physiological and Health Effects.
Although objective social isolation can affect loneliness,55, 56 perceived social isolation (loneliness) is more closely related to the quality than quantity of social interactions.55 This is in part because loneliness is influenced by factors unrelated to objective isolation, including genetics,57 childhood environment,18 cultural norms58, social needs59, physical disabilities55, and discrepancies between actual and desired relationships.60 Accordingly, perceived social isolation predicts various outcomes above and beyond what is predicted by objective isolation. For instance, loneliness predicts elevated blood pressure61, morning rise in cortisol36, physical activity62, perceptions of the neighborhood environment63, and changes in life satisfaction across a lifetime17 beyond what could be predicted by social support or objective social isolation. In studies of cognitive functioning, Wilson et al. 16 found no evidence for an influence of social network size or frequency of social activity on cognitive decline or on risk for Alzheimer's Disease (AD), whereas loneliness persisted in predicting each of these outcomes even when social network size and frequency of social activity were statistically held constant. Similarly, loneliness has been found to predict lifetime change in IQ17 and changes in depressive symptoms39 beyond what could be predicted by objective isolation. Experimental manipulation of loneliness30 and imagined future isolation64 result in cognitive changes even though objective isolation is not altered in these experimental studies. Perceived (but not objective) social isolation has even been associated with gene expression—specifically, the under-expression of genes bearing anti-inflammatory glucocorticoid response elements and over-expression of genes bearing response elements for pro-inflammatory NK-κB/Rel transcription factors [37; See Figure 1]. This finding is paralleled by decreased lymphocyte sensitivity to physiological regulation by the hypothalamic pituitary adrenocortical (HPA) axis in lonely individuals65, which together with evidence of increased activity of the HPA axis24, 66, 67, suggests the development of glucocorticoid resistance in chronically lonely individuals. We, therefore, focus the current review on the association between perceived social isolation and cognition.
Perceived social isolation, known more colloquially as loneliness, was characterized in early scientific investigations as “a chronic distress without redeeming features” (9, p. 15). Recent research suggests that the social pain of loneliness evolved as a signal that one's connections to others are weakening and to motivate the repair and maintenance of the connections to others that are needed for our health and well being and for the survival of our genes (see Box 2;10). Experimental, cross-sectional, and longitudinal studies are beginning to elucidate the various ways in which loneliness is related to, and in some cases affects, human information processing.
Box 2. A Biological Signal to Renew the Connections Needed to Survive and Prosper.
The species Homo sapiens is fundamentally social, and in ontogeny and phylogeny humans need others to survive and prosper. Physical pain is an aversive signal that evolved to motivate one to take action to minimize damage to one's body. Loneliness, a social pain, is an aversive signal that evolved to motivate one to take action that minimizes threats or damage to one's social body. Research on social rejection, for instance, suggests that social pain co-opted the physical pain to extend its protective function to include those with whom we form connections. In Eisenberger, Lieberman, and Williams68, participants were excluded from or included in a social situation (i.e., a ball tossing game). Results revealed neural activation localized in a dorsal portion of the anterior cingulate cortex (dACC) that is implicated in the affective component of the physical pain response. Eisenberger and her colleagues suggest that “Because of the adaptive value of mammalian social bonds, the social attachment system… may have piggybacked onto the physical pain system to promote survival” (p. 291).
Loneliness also appears to modulate the rudimentary reward system to extend its protective function to social cognition and emotion. The ventral striatum, a key component of the mesolimbic dopamine system, is rich in dopaminergic neurons and is critical in reward processing and learning69, 70. The ventral striatum is activated by primary rewards such as stimulant drugs71, abstinence-induced cravings for primary rewards72, and secondary rewards such as money73. Evidence that social reward also activate the ventral striatum has begun to accumulate in studies of romantic love74, social cooperation75, social comparison76, and punitive altruism77. Cacioppo and colleagues78 investigated how an individual's loneliness was related to the differential activation of the ventral striatum to pleasant social versus matched nonsocial images. As depicted in Figure 2, lonely individuals showed weaker activation of the ventral striatum to pleasant pictures of people than of equally pleasant pictures of objects, whereas nonlonely individuals showed stronger activation of the ventral striatum to pleasant pictures of people than of objects. Experience sampling studies confirm that lonely individuals regard pleasant interpersonal interactions to be less pleasant but less so than do nonlonely individuals.31
Cognitive Capacities
Human social processes were once thought to have been incidental to learning and cognition, whereas the social complexities and demands of primate species are now thought to have contributed to the evolution of the neocortex and various aspects of human cognition. Consistent with this reasoning, human toddlers and chimpanzees have similar cognitive skills for engaging the physical world but toddlers have more sophisticated cognitive skills than chimpanzees for engaging the social world11; cross-species comparisons have revealed that the evolution of large and metabolically expensive brains is more closely associated with social than ecological complexity12; and a composite index of sociality in troops of baboons has been found to be highly correlated with infant survival.13
Evidence that social isolation might be related to fundamental aspects of cognition comes from animal research showing that isolation impairs learning that requires the inhibition of previously learned responses (e.g., reversal learning, extinction;14) and from human research showing that loneliness is a risk factor for cognitive decline15-17 and AD17. Gow and colleagues17 investigated the correlates of changes in mental ability of 488 individuals from the Lothian Birth Cohort Study who were tested at age 11 and 79. Among the variables tested were loneliness, social support, and social network (e.g., presence of significant others, number of significant others). After controlling for age-11 IQ, gender, years of education, and social class, only loneliness was associated significantly with changes in IQ. Although loneliness is temporally stable and heritable18, 19, the study by Gow and colleagues does not address the possibility that loneliness is a consequence rather than a predictor of cognitive decline. Two recent longitudinal studies do speak to this question.
Tilvis and colleagues15 measured cognition by the mini-mental state examination and the Clinical Dementia Rating at baseline and at one, five and ten year assessments of a population-based sample of 75-85 year old individuals. Results at the 10-year follow-up assessment revealed APOE4, elevated serum (ionized) calcium, and loneliness independently predicted cognitive decline. In a larger prospective study, Wilson et al.16 assessed 823 older adults free of dementia at enrollment. Participants completed an extensive battery of cognitive measures to assess global cognition, episodic memory, semantic memory, working memory, perceptual speed, and visuospatial ability. The lonelier were the participants, the poorer the cognitive performance within each of these domains at baseline, and loneliness was associated with greater cognitive declines in every domain except working memory and episodic performance. Furthermore, 76 individuals developed dementia during the 65 month study period. Cox proportional hazards models that controlled for age, sex, and education indicated that loneliness significantly increased the risk of clinical AD, and this association was unchanged when objective social isolation and other demographic and health-related factors served as covariates. Depression may also contribute to cognitive declines, so Wilson et al.16 also examined the possible role of depressive symptoms. The loneliness at baseline predicted cognitive decline and the onset of AD even when depressive symptoms served as a covariate. For instance, the association between loneliness and AD risk was reduced by about 16% when controlling for depressive symptoms and remained significant. Depressive symptomatology, on the other hand, was marginally related to the risk of AD, and this association was reduced by more than half when controlling for loneliness (see, also, Hertzog, Kramer, Wilson, & Lindenberger,20).
Brain autopsies were available for 67% of the participants who died during the study. Of these, 30% had a clinical diagnosis of AD. Loneliness and the neuropathological measures derived from the brain autopsy were each inversely related to global cognition at the last assessment prior to death, but loneliness was unrelated to the neuropathological measures. Although the mechanism underlying the association between loneliness and cognitive decline has not yet been identified, social isolation was recently shown to decrease central anti-inflammatory responses and survival rate, and increase the infarct size and edema development, following the induction of stroke in mice.3 The deficits in reversal learning associated with isolation in animals have also been associated with diminished prefrontal-cortico-striatal functioning21, a neural mechanism involved in the inhibition of previously learned responses. Socially isolated animals also show less dendritic arborization in the hippocampus and prefrontal cortex22 and decreased brain-derived neurotrophic factor23. To what extent loneliness produces similar neurophysiological changes and how such differences contribute to cognitive decline in humans are open questions. Consistent with the animal research, however, human studies indicate that loneliness impairs executive functioning, specifically the inhibition of prepotent responses. We turn to that evidence next.
Executive Functioning
Executive functioning includes the capacity to control one's attention, cognition, emotion, and/or behavior to better meet social standards or personal goals, that is, to self-regulate. Early evidence from young adults who performed a dichotic listening task suggested that attentional regulation was poorer in lonely than nonlonely individuals.24 Participants were asked to identify the consonant-vowel pair presented in the left or right ear. Typically, performance shows a right-ear advantage and performance is better for the ear to which participants have been instructed to attend. Lonely and nonlonely individuals showed an equivalent right ear advantage under the no-instruction condition and an equivalent attentional shift to the right ear when so instructed, but lonely participants showed a weaker left-ear advantage (the non-prepotent response) when instructed to attend to this ear.
Poorer self regulation when feeling isolated is not limited to attentional control. In cross-sectional and longitudinal research, lonely individuals have been found to have lower odds of engaging in regular exercise than nonlonely individuals, and the poorer emotional regulation of individuals when they felt lonely mediated the effect.25 Experimental manipulations that lead people to believe they face a future of social isolation also decrease self-regulation. In an illustrative study, Baumeister and colleagues26 had the participants complete two questionnaires: an introversion/extraversion test, and a personality inventory. Participants then were randomly assigned to receive no feedback (Control Group) or to receive feedback to induce feelings of a future of social isolation (Future Alone), social connection (Future Belonging), or general misfortune (Misfortune Control Group). Results revealed that the Future Alone group performed significantly worse than the other groups on the General Mental Ability Test of the Graduate Record Exam. Bad news itself was not enough to cause the disruption, only bad news about social connection.
In subsequent variations on this experimental paradigm, randomly assigned participants to the Future Alone Group, relative to the other groups, performed similarly on a rote memorization task but attempted the fewest problems and made the most mistakes on a logical reasoning task27, consumed more delicious but unhealthy foods27, and were more aggressive toward others28. A perceived future of social isolation, then, did not impair routine mental ability, only the higher order cognitive and self-regulatory processes that are characteristic of executive functioning. A brain scan conducted while participants performed moderately difficult math problems revealed that the brains of the future socially isolated participants were less active in the areas involved in the “executive control” of attention.29
Colorations of Cognition
Experimental manipulations of loneliness not only impair executive functioning, they produce higher negative mood, anxiety, anger, and depressive symptomatology.30 An experience sampling study, in which participants were beeped randomly nine times per day for seven days, confirmed that the social interactions of lonely, in contrast to nonlonely, individuals were more negative and less satisfying, and such interactions contributed subsequently to more negative moods and interactions.31 Evidence from behavioral and functional magnetic resonance imaging (fMRI) studies further suggests that loneliness increases attention to negative social stimuli (e.g., social threats). Using a modified emotional Stroop task, lonely participants, relative to nonlonely participants, showed greater Stroop interference specifically for negative social relative to negative non-social words.32 No differences between lonely and nonlonely participants were found in Stroop interference for positive social relative to positive non-social words. Stroop interference is used to gauge the implicit processing of stimuli, so these results suggest that loneliness is associated with a heightened accessibility of negative social information. Similarly, Yamada and Decety33 investigated the effects of subliminal priming on the detection of painful facial expressions. Using signal detection analyses, they found that lonely individuals were more sensitive (d′) to the presence of pain in dislikable faces than were nonlonely individuals.
The patterns of regional brain activation found when lonely and nonlonely individuals think about people differ, as well.34 A region associated with reward and appetitive behavior (i.e., ventral striatum) was more strongly activated in nonlonely than lonely individuals when exposed to pleasant social pictures in contrast to pleasant nonsocial pictures (see Figure 2). On the other hand, activation of the visual cortex to the presentation of unpleasant social, in contrast to nonsocial, pictures was directly related to the loneliness of the participant, indicative of greater visual attention to the negative social stimuli., These results are consistent with the behavioral data indicating that loneliness is related to the attention elicited by negative social stimuli.
Figure 2.
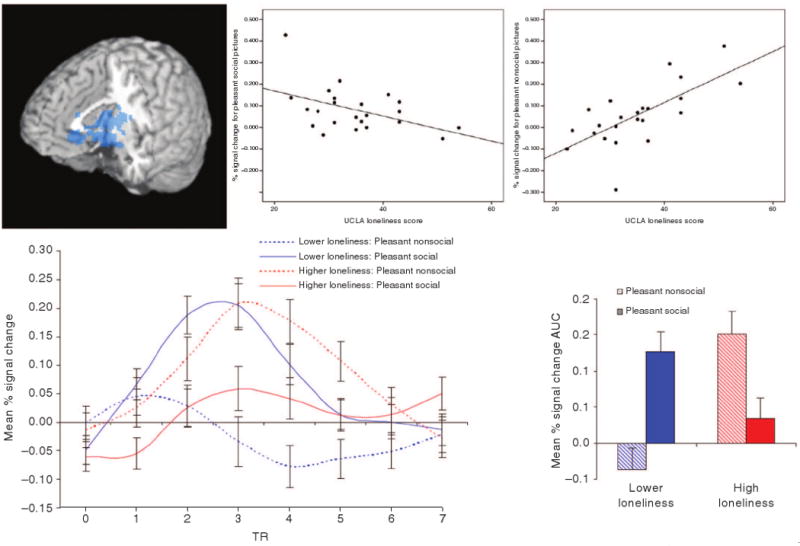
A cluster of voxels centered in the ventral striatum, but extending to the amygdala and portions of the anterior thalamus, showed an inverse relationship between loneliness and activation in the pleasant social–pleasant nonsocial contrast. The scatterplots demonstrate the association between loneliness and activity in this cluster in response to pleasant social pictures [r(21) = _.46, p < .05], and in response to pleasant nonsocial pictures [r(21) = .69, p < .001]. Estimated impulse response functions and mean percent signal change AUC for participants lower and higher in loneliness (estimates at 1 SD above and below the mean UCLA score in our sample are presented) show a crossover interaction for the relationship between loneliness and brain responses to pleasant social and pleasant nonsocial stimuli, such that nonlonely participants exhibit greater activation to pleasant pictures that contain social content and lonely participants exhibit greater activation to pleasant nonsocial pictures. (From Cacioppo, J. T., Norris, C. J., Decety, J., Monteleone, G., & Nusbaum, H. (2009). In the eye of the beholder: Individual differences in perceived social isolation predict regional brain activation to social stimuli. Journal of Cognitive Neuroscience, 2009 78.)
A possible casualty of loneliness and the priming of social threats is that lonely individuals may be more likely to focus on themselves, their needs, and their preservation in negative circumstances. To examine this possibility, activation in the temporoparietal junction (TPJ) -- a region that has been found previously to be activated in theory of mind tasks and in tasks in which individuals take the perspective of another, was also examined.34 Consistent with this reasoning, TPJ activation was observed when participants viewed unpleasant pictures of people versus objects, and loneliness was inversely related to amount of activation observed.
The hypersensitivity to negative social information and the diminished pleasure derived from positive social stimuli might be expected to shape social expectations and motivations and contribute to a downward spiraling of negative affect and depressive symptomatology. Indeed, loneliness is related to stronger expectations of and motivations to avoid bad social outcomes and weaker expectations of and motivations to approach good social outcomes.35 Furthermore, loneliness and depressive symptoms are distinct states both by measures of statistical30 and functional independence,36-38 and longitudinal studies have shown that loneliness predicts increases in depressive symptomatology above and beyond what can be explained by basal levels of depressive symptomatology.39-41 Results are mixed in terms of whether depressive symptoms have a reciprocal influence on loneliness.
Social Cognition
The brains of lonely, in contrast to nonlonely, individuals are on high alert for social threats, so lonely individuals tend to view their social world as threatening and punitive (see Box 3). Experimental manipulations of loneliness not only cause people to feel more anxious, fear negative evaluation, and act more coldly toward others30, it causes them to feel colder.42 Lonely individuals also tend to form more negative social impressions of others, and their expectations, attributional reasoning, and actions toward others tend to be less charitable than shown by nonlonely individuals.43 When an individual's negative social expectations elicit behaviors from others that validate these expectations, the expectations are buttressed and increase the likelihood of the individual behaving in ways that pushes away the very people to whom he or she most wants to be close to better fulfill their social needs.44, 45 Consequently, lonely individuals may view themselves to be passive victims in their social world, but they are active contributors through their self-protective and paradoxically self-defeating interactions with others.43
Box 3. Regulatory Loop and Remaining Questions.
Given our evolutionary heritage, the human brain and biology have been sculpted to seek meaningful connections with others. In evolutionary time, social groupings were relatively small and stable, and the pain of loneliness may have served both to promote the social connection necessary for the survival of the genes and as a deterrent to selfish actions that were detrimental to the group.58 The evidence reviewed here, however, suggests that loneliness in contemporary society may affect human cognition in sometimes maladaptive ways (see Figure 3). Specifically, feeling socially isolated can trigger implicit hypervigilance for social threats, which in turn produces attentional, confirmatory, and memorial biases. Accordingly, lonely individuals are more likely to attend to and construe their social world as threatening, hold more negative social expectations, and remember more negative social events than are nonlonely individuals. These cognitions increase the likelihood that individuals engage in behavioral confirmation processes, through which they produce more negative social interactions and elicit evidence confirming that they have little personal control or social value. These dispositions, in turn, alter the nature and likelihood of social engagement and activate neurobiological mechanisms that increase activation of the hypothalamic pituitary adrenal (HPA) axis and diminish sleep quality. Repeated or chronic activation of threat surveillance in a social context, coupled with diminished anabolic processes, may contribute to heightened cognitive load, diminished executive functioning, dysregulated brain and physiological systems, and broad based morbidity and mortality. Although this theoretical model is consistent with the evidence reviewed here, many details remain to be tested and refined. The following questions may be of special import.
To what extent is loneliness a consequence rather than an antecedent of incipient dementia? Longitudinal and experimental studies suggest loneliness may play a causal role in at least some of the observed associations between loneliness and cognitive functioning, but sensory loss, functional impairments, and cognitive impairments may also lessen social contact and increase loneliness. Questions remain about the extent to which, and conditions under which, the associations between loneliness and cognitive functioning reflect the effects of loneliness, the effects of cognition, and the effects of a third variable.
By what mechanism might loneliness contribute to cognitive decline as people age? Among the possible mechanistic pathways that warrant investigation are the effects of loneliness on: (a) elevated activation of the HPA axis and/or increased inflammatory responses in the brain, each of which may impact cognitive functioning; (b) behavioral or neural plasticity such that older individuals are less able to compensate for age-related degenerative changes in neural systems involved in cognitive functioning; (c) the simplification of social cognition and reduction in social stimulation and engagement that are tantamount to lower cognitive stimulation; (d) cognitive load (and reduction in available cognitive resources available for creative adaptations) produced by chronic surveillance for and protection from threats; (e) elevations in depression and/or reductions in physical activity; (f) the reduction in the number and quality of social interactions; and (g) impairments in the consolidation of learning that may result from diminished sleep quality.
Does the detrimental effect of loneliness occur later in life or is it cumulative across the lifespan? If cumulative, are the deleterious effects of loneliness on cognition (if causal) reversible? Increasing social contact and social support appear not to be sufficient to lower loneliness or to explain the effects of loneliness on cognition. How does one intervene to reduce perceived social isolation and promote healthy social connections? Does the nature of effective interventions for loneliness differ across age, gender, or ethnicity?
What are the brain mechanisms underlying the association between loneliness and cognition? Diminished prefrontal-cortico-striatal functioning, dendritic arborization, brain-derived neurotrophic factors, and central anti-inflammatory responses are just a few of the possibilities that are suggested by the extant evidence.
Although loneliness may interfere with a person's attempts to form stable and trusting social connections with other people, loneliness has been found to promote attempts to form social connections though memorial or inferential means, such as through nostalgic reminiscences,46 imputed parasocial relationships (relationships with imaginary television characters;47), and digital connections.48 In addition, loneliness promotes social connection through the anthropomorphism of pets,49 technological gadgets,50 celestial bodies,51 and supernatural entities.51 Together, this work suggests that when people feel socially isolated, they become more likely to use their cognitive capacities to try to fill the social void.
Contagion
Loneliness is typically investigated as an individual factor, but because perceived and objective isolation can be differentiated, loneliness can also vary within and across groups. Network linkage data from the population-based Framingham Heart Study were used to trace the topography of loneliness in social networks, and the path through which loneliness spreads through these networks.52 Results indicated that loneliness occurs in clusters within social networks, is disproportionately represented at the periphery of social networks, extends up to three degrees of separation, and is stronger for women than men.
Several features of the Framingham study pointed to loneliness spreading through a contagious process and moving lonely individuals closer to the edge of social networks over time. Contagion is defined as the transmission of a state by direct or indirect contact, and virulence is determined, in part, by exposure (i.e., dose). Longitudinal analyses indicated that loneliness in one individual at Time 1 was followed by others in the social network becoming lonelier by Time 2. Second, the closer the friend or contact was physically to this individual at Time 1, the lonelier the friend or contact became by Time 2. Third, loneliness was transmitted from the individual at Time 1 through friends and contacts to others beyond the individual's circle of contacts such that these other individuals became lonelier by Time 2. Fourth, the transmission of loneliness was stronger when the friendship between the individual who was lonely at Time 1 and others in the social network was reciprocal than not. Importantly, these results were unchanged when controlling for depressive symptomatology, indicating that the contagion of loneliness was not secondary to depression.52
If loneliness is contagious, driving away those who are lonely functions to keep the contagion in check, leading people who feel socially isolated to become objectively more isolated. Loneliness not only spreads from person to person within a social network, but it reduces the ties of these individuals to others within the network. The collective rejection of isolates observed in humans and other primates may therefore serve to protect the structural integrity of the social entities necessary for humans to survive and prosper.
Data from the Framingham study do not permit detailed investigation of the means by which loneliness was transmitted, but this contagion may occur through three different mechanisms: automatic emotional contagion,53 coextensive self-other overlap and the attendant susceptibility of shared states,54 and quality of social interactions.31 For instance, in an experience sampling study of everyday behavior, loneliness was associated with more negative affect and more negative social interactions, the quality of social interactions predicted subsequent affective states and vice versa, and more negative social interactions had longer lasting effects on affect than positive social interactions.31 These data are consistent with the notion that the contagion of loneliness can occur through the more negative social cognition and interpersonal interactions it engenders.
Conclusion
Cognition has been regarded as the quintessential individual activity. Mental representations and processes were rendered testable in the dawn of the cognitive sciences by virtue of reverse engineering: mathematical and computer models were created that specified stimulus inputs, information processing operations that acted on and transformed these inputs to produce and change representational structures, and information processing operations that led to observable responses. Computers today are no longer solitary devices, but rather they operate as a connected collective resulting in power, capacities, representations, and processes that were unforeseen. Social species create emergent organizations beyond the individual– structures ranging from dyads and families to institutions and cultures. These emergent structures evolved hand in hand with neural, hormonal, and genetic mechanisms to support them because the consequent social behaviors helped these organisms survive, reproduce, and care for offspring sufficiently long that they too reproduced. These emergent levels of organization have long been apparent, but identifying their biological and cognitive bases and consequences is one of the major problems for the cognitive sciences to address this century.
Figure 1.
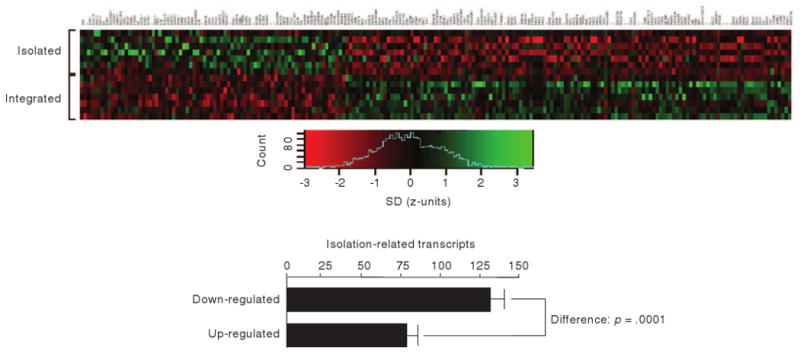
Differential gene expression in individuals high versus low in loneliness. Genome-wide transcriptional profiles were assessed in peripheral blood leukocyte RNA samples collected from individuals in the top and bottom 15% of the distribution of subjective social isolation. Analysis by Affymetrix U133A high-density oligonucleotide arrays identified 209 transcripts showing >30% difference in mean expression levels across groups (green = over-expression in high-lonely, red = under-expression). High subjective social isolation is associated with a statistically significant net reduction in the number of expressed genes (131 down-regulated versus 78 up-regulated, p value by exact binomial test). (From Cole, Hawkley, Arevalo, Sung, Rose, & Cacioppo, 2007 37.)
Figure 3.
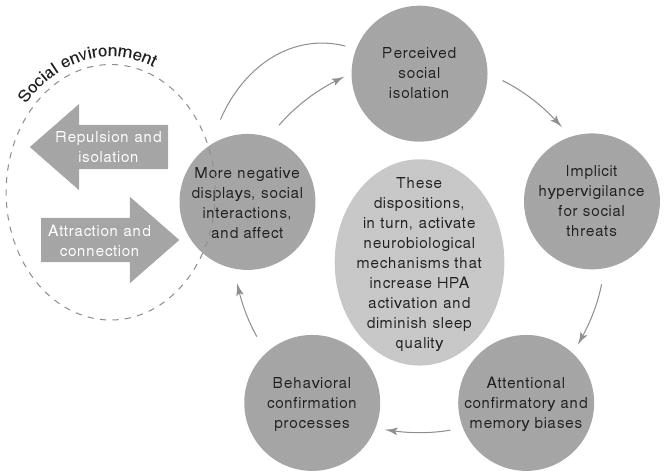
The effects of loneliness on human cognition.
Acknowledgments
This research was supported by National Institute of Aging Program Project Grant No. PO1 AG18911 & RO1 AG034052-01 and by a grant from the John Templeton Foundation.
Footnotes
Disclosure Statement: These funding sources had no role or influence in the preparation of this review.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Ruan H, Wu CF. Social interaction-mediated lifespan extension of Drosophila Cu/Zn superoxide dismutase mutants. Proc Natl Acad Sci U S A. 2008;105:7506–7510. doi: 10.1073/pnas.0711127105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Nonogaki K, et al. Social Isolation Affects the Development of Obesity and Type 2 Diabetes in Mice. Endocrinology. 2007;148:4658–4666. doi: 10.1210/en.2007-0296. [DOI] [PubMed] [Google Scholar]
- 3.Karelina D, et al. Social isolation alters neuroinflammatory response to stroke. Proceedings of the National Academy of Sciences. doi: 10.1073/pnas.0810737106. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dronjak S, et al. Immobilization and cold stress affect sympatho-adrenomedullary system and pituitary-adrenocortical axis of rats exposed to long-term isolation and crowding. Physiol Behav. 2004;81:409–415. doi: 10.1016/j.physbeh.2004.01.011. [DOI] [PubMed] [Google Scholar]
- 5.Stranahan AM, et al. Social isolation delays the positive effects of running on adult neurogenesis. Nat Neurosci. 2006;9:526–533. doi: 10.1038/nn1668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kanitz E, et al. Consequences of repeated early isolation in domestic piglets (Sus scrofa) on their behavioural, neuroendocrine, and immunological responses. Brain Behav Immun. 2004;18:35–45. doi: 10.1016/s0889-1591(03)00085-0. [DOI] [PubMed] [Google Scholar]
- 7.Nation DA, et al. The effect of social environment on markers of vascular oxidative stress and inflammation in the Watanabe heritable hyperlipidemic rabbit. Psychosom Med. 2008;70:269–275. doi: 10.1097/PSY.0b013e3181646753. [DOI] [PubMed] [Google Scholar]
- 8.Poletto R, et al. Effects of early weaning and social isolation on the expression of glucocorticoid and mineralocorticoid receptor and 11beta-hydroxysteroid dehydrogenase 1 and 2 mRNAs in the frontal cortex and hippocampus of piglets. Brain Res. 2006;1067:36–42. doi: 10.1016/j.brainres.2005.10.001. [DOI] [PubMed] [Google Scholar]
- 9.Weiss RS. Loneliness: The experience of emotional and social isolation. MIT Press; 1973. [Google Scholar]
- 10.Cacioppo JT, Patrick B. Loneliness: Human nature and the need for social connection. W. W. Norton & Company; 2008. [Google Scholar]
- 11.Herrmann E, et al. Humans Have Evolved Specialized Skills of Social Cognition: The Cultural Intelligence Hypothesis. Science. 2007;317:1360–1366. doi: 10.1126/science.1146282. [DOI] [PubMed] [Google Scholar]
- 12.Dunbar RIM, Shultz S. Evolution in the Social Brain. Science. 2007;317:1344–1347. doi: 10.1126/science.1145463. [DOI] [PubMed] [Google Scholar]
- 13.Silk JB, et al. Social bonds of female baboons enhance infant survival. Science. 2003;302:1231–1234. doi: 10.1126/science.1088580. [DOI] [PubMed] [Google Scholar]
- 14.Schrijver NC, et al. Double dissociation of social and environmental stimulation on spatial learning and reversal learning in rats. Behav Brain Res. 2004;152:307–314. doi: 10.1016/j.bbr.2003.10.016. [DOI] [PubMed] [Google Scholar]
- 15.Tilvis RS, et al. Predictors of Cognitive Decline and Mortality of Aged People Over a 10-Year Period. J Gerontol A Biol Sci Med Sci. 2004;59:M268–274. doi: 10.1093/gerona/59.3.m268. [DOI] [PubMed] [Google Scholar]
- 16.Wilson RS, et al. Loneliness and risk of Alzheimer disease. Arch Gen Psychiatry. 2007;64:234–240. doi: 10.1001/archpsyc.64.2.234. [DOI] [PubMed] [Google Scholar]
- 17.Gow AJ, et al. Social support and successful aging: Investigating the relationships between lifetime cognitive change and life satisfaction. Journal of Individual Differences. 2007;28:103–115. [Google Scholar]
- 18.Bartels M, et al. Genetic and environmental contributions to stability in loneliness throughout childhood. Am J Med Genet B Neuropsychiatr Genet. 2008;147:385–391. doi: 10.1002/ajmg.b.30608. [DOI] [PubMed] [Google Scholar]
- 19.Boomsma DI, et al. Genetic and environmental contributions to loneliness in adults: The Netherlands twin register study. Behav Genet. 2005;35:745–752. doi: 10.1007/s10519-005-6040-8. [DOI] [PubMed] [Google Scholar]
- 20.Hertzog C, et al. Enrichment Effects on Adult Cognitive Development: Can the Functional Capacity of Older Adults Be Preserved and Enhanced? Psychological Science in the Public Interest. 2008;9:1–65. doi: 10.1111/j.1539-6053.2009.01034.x. [DOI] [PubMed] [Google Scholar]
- 21.Heidbreder CA, Weiss IC, Domeney AM, Pryce C, Homberg J, Hedou G, Feldon J, Moran MC, Nelson P. Behavioral, neurochemical and endocrinological characterization of the early social isolation syndrome. Neuroscience. 2000;100:749–768. doi: 10.1016/s0306-4522(00)00336-5. [DOI] [PubMed] [Google Scholar]
- 22.Silva-Gomez AB, et al. Decreased dendritic spine density on prefrontal cortical and hippocampal pyramidal neurons in postweaning social isolation rats. Brain Res. 2003;983:128–136. doi: 10.1016/s0006-8993(03)03042-7. [DOI] [PubMed] [Google Scholar]
- 23.Barrientos RM, et al. Brain-derived neurotrophic factor mRNA downregulation produced by social isolation is blocked by intrahippocampal interleukin-1 receptor antagonist. Neuroscience. 2003;121:847–853. doi: 10.1016/s0306-4522(03)00564-5. [DOI] [PubMed] [Google Scholar]
- 24.Cacioppo JT, et al. Lonely traits and concomitant physiological processes: the MacArthur social neuroscience studies. Int J Psychophysiol. 2000;35:143–154. doi: 10.1016/s0167-8760(99)00049-5. [DOI] [PubMed] [Google Scholar]
- 25.Hawkley LC, et al. Loneliness predicts reduced physical activity: Cross-sectional and longitudinal analyses. Health Psychology. doi: 10.1037/a0014400. In Press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Baumeister R, DeWall N. The inner dimension of social exclusion: Intelligent thought and self-regulation among rejected persons. In: Williams KD, F JP, von Hippel W, editors. The social outcast: Ostracism, social exclusion, rejection, and bullying. Psychology Press; 2005. pp. 53–76. [Google Scholar]
- 27.Baumeister RF, et al. Social exclusion impairs self-regulation. J Pers Soc Psychol. 2005;88:589–604. doi: 10.1037/0022-3514.88.4.589. [DOI] [PubMed] [Google Scholar]
- 28.Twenge JM, et al. If you can't join them, beat them: Effects of social exclusion on aggressive behavior. J Pers Soc Psychol. 2001;81:1058–1069. doi: 10.1037//0022-3514.81.6.1058. [DOI] [PubMed] [Google Scholar]
- 29.Campbell WK, et al. A magnetoencephalography investigation of neural correlates for social exclusion and self-control. Soc Neurosci. 2006;1:124–134. doi: 10.1080/17470910601035160. [DOI] [PubMed] [Google Scholar]
- 30.Cacioppo JT, et al. Loneliness within a nomological net: An evolutionary perspective. Journal of Research in Personality. 2006;40:1054–1085. [Google Scholar]
- 31.Hawkley LC, et al. Multilevel modeling of social interactions and mood in lonely and socially connected individuals: The MacArthur social neuroscience studies. In: Ong AD, van Dulmen M, editors. Oxford Handbook of Methods in Positive Psychology. Oxford University Press; 2007. pp. 559–575. [Google Scholar]
- 32.Shintel H, et al. Accentuate the negative, eliminate the positive? Individual differences in attentional bias to positive and negative information. 47th Annual Meeting of the Psychonomic Society 2006 [Google Scholar]
- 33.Yamada M, Decety J. Unconscious affective processing and empathy: An investigation of subliminal priming on the detection of painful facial expressions. Pain. 2009;143:71–75. doi: 10.1016/j.pain.2009.01.028. [DOI] [PubMed] [Google Scholar]
- 34.Cacioppo JT, et al. In the Eye of the Beholder: Individual Differences in Perceived Social Isolation Predict Regional Brain Activation to Social Stimuli. J Cogn Neurosci. 2008;21:83–92. doi: 10.1162/jocn.2009.21007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gable SL. Approach and avoidance social motives and goals. J Pers. 2006;74:175–222. doi: 10.1111/j.1467-6494.2005.00373.x. [DOI] [PubMed] [Google Scholar]
- 36.Adam EK, et al. Day-to-day dynamics of experience--cortisol associations in a population-based sample of older adults. Proceedings of the National Academy of Science USA. 2006;103:17058–17063. doi: 10.1073/pnas.0605053103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Cole SW, et al. Social regulation of gene expression in human leukocytes. Genome Biology. 2007;8:R189.181–R189.113. doi: 10.1186/gb-2007-8-9-r189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hawkley LC, et al. Loneliness is a unique predictor of age-related differences in systolic blood pressure. Psychol Aging. 2006;21:152–164. doi: 10.1037/0882-7974.21.1.152. [DOI] [PubMed] [Google Scholar]
- 39.Cacioppo JT, et al. Loneliness as a specific risk factor for depressive symptoms: cross-sectional and longitudinal analyses. Psychol Aging. 2006;21:140–151. doi: 10.1037/0882-7974.21.1.140. [DOI] [PubMed] [Google Scholar]
- 40.Heikkinen RL, Kauppinen M. Depressive symptoms in late life: a 10-year follow-up. Arch Gerontol Geriatr. 2004;38:239–250. doi: 10.1016/j.archger.2003.10.004. [DOI] [PubMed] [Google Scholar]
- 41.Wei M, et al. Adult Attachment, Social Self-Efficacy, Self-Disclosure, Loneliness, and Subsequent Depression for Freshman College Students: A Longitudinal Study. Journal of Counseling Psychology. 2005;52:602–614. [Google Scholar]
- 42.Zhong CB, Leonardelli GJ. Cold and lonely: does social exclusion literally feel cold? Psychol Sci. 2008;19:838–842. doi: 10.1111/j.1467-9280.2008.02165.x. [DOI] [PubMed] [Google Scholar]
- 43.Cacioppo JT, Hawkley LC. People thinking about people: The vicious cycle of being a social outcast in one's own mind. In: Williams KD, et al., editors. The social outcast: Ostracism, social exclusion, rejection, and bullying. Psychology Press; 2005. pp. 91–108. [Google Scholar]
- 44.Murray SL, et al. Once hurt, twice hurtful: How perceived regard regulates daily marital interactions. J Pers Soc Psychol. 2003;84:126–147. [PubMed] [Google Scholar]
- 45.Downey GD, Romero-Canyas R. Rejection sensitivity as a predictor of affective and behavioral responses to interpersonal stress: A defensive motivational system. In: Williams KD, et al., editors. The social outcast: Ostracism, social exclusion, rejection, and bullying. The Psychology Press; 2005. [Google Scholar]
- 46.Zhou X, et al. Counteracting loneliness: on the restorative function of nostalgia. Psychol Sci. 2008;19:1023–1029. doi: 10.1111/j.1467-9280.2008.02194.x. [DOI] [PubMed] [Google Scholar]
- 47.Gardner WL, Knowles ML. Love makes you real: Favorite television characters are perceived as “real” in a social facilitation paradigm. Social Cognition. 2008;26:156–168. [Google Scholar]
- 48.Ozcan NK, Buzlu S. Internet use and its relation with the psychosocial situation for a sample of university students. Cyberpsychol Behav. 2007;10:767–772. doi: 10.1089/cpb.2007.9953. [DOI] [PubMed] [Google Scholar]
- 49.Epley N, et al. When we need a human: Motivational determinants of anthropomorphism. Social Cognition. 2008;26:143–155. [Google Scholar]
- 50.Epley N, et al. Creating social connection through inferential reproduction -Loneliness and perceived agency in gadgets, gods, and greyhounds. Psychological Science. 2008;19:114–120. doi: 10.1111/j.1467-9280.2008.02056.x. [DOI] [PubMed] [Google Scholar]
- 51.Epley N, et al. On seeing human: A three-factor theory of anthropomorphism. Psychol Rev. 2007;114:864–886. doi: 10.1037/0033-295X.114.4.864. [DOI] [PubMed] [Google Scholar]
- 52.Cacioppo JT, et al. Alone in the crowd: The structure and spread of loneliness in a large social network. J Pers Soc Psychol. doi: 10.1037/a0016076. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Hatfield E, et al. Emotional Contagion. Cambridge University Press; 1994. [Google Scholar]
- 54.Slotter EB, Gardner WL. Where do you end and I begin? Evidence for anticipatory, motivated self-other integration between relationship partners. J Pers Soc Psychol. 2009;96:1137–1151. doi: 10.1037/a0013882. [DOI] [PubMed] [Google Scholar]
- 55.Hawkley LC, et al. From social structure factors to perceptions of relationship quality and loneliness: The Chicago Health, Aging, and Social Relations Study. Journal of Gerontology: Social Sciences. 2008 doi: 10.1093/geronb/63.6.s375. In Press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ruiz JI. Emotional Climate in Organizations: Applications in Latin American Prisons. Journal of Social Issues. 2007;63:289–306. [Google Scholar]
- 57.Boomsma D, et al. Genetic and Environmental Contributions to Loneliness in Adults: The Netherlands Twin Register Study. Behav Genet. 2005;35:745–752. doi: 10.1007/s10519-005-6040-8. [DOI] [PubMed] [Google Scholar]
- 58.Cacioppo JT, Patrick B. Loneliness: Human nature and the need for social connection. W. W. Norton & Company; 2008. [Google Scholar]
- 59.Dykstra PA, Fokkema T. Social and Emotional Loneliness Among Divorced and Married Men and Women: Comparing the Deficit and Cognitive Perspectives. Basic & Applied Social Psychology. 2007;29:1–12. [Google Scholar]
- 60.Cacioppo JT, Hawkley LC. Loneliness. In: Leary MR, Hoyle RH, editors. Handbook of individual differences in social behavior. Guilford; In press. [Google Scholar]
- 61.Hawkey LC, et al. Loneliness is a unique predictor of age-related differences in systolic blood pressure. Psychol Aging. 2006;21:152–164. doi: 10.1037/0882-7974.21.1.152. [DOI] [PubMed] [Google Scholar]
- 62.Hawkley LC, et al. Loneliness predicts reduced physical activity: Cross-sectional & longitudinal analyses. Health Psychol. 2009;28:354–363. doi: 10.1037/a0014400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wen M, et al. Objective and perceived neighborhood environment, individual SES and psychosocial factors, and self-rated health: An analysis of older adults in Cook County, Illinois. Soc Sci Med. 2006;63:2575–2590. doi: 10.1016/j.socscimed.2006.06.025. [DOI] [PubMed] [Google Scholar]
- 64.Baumeister RF, et al. Effects of social exclusion on cognitive processes: Anticipated aloneness reduces intelligent thought. J Pers Soc Psychol. 2002;83:817–827. doi: 10.1037//0022-3514.83.4.817. [DOI] [PubMed] [Google Scholar]
- 65.Cole SW. Social regulation of leukocyte homeostasis: The role of glucocorticoid sensitivity. Brain Behav Immun. 2008;22:1049–1055. doi: 10.1016/j.bbi.2008.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Adam EK, et al. Day-to-day dynamics of experience-cortisol associations in a population-based sample of older adults. Proc Natl Acad Sci U S A. 2006;103:17058–17063. doi: 10.1073/pnas.0605053103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Steptoe A, et al. Loneliness and neuroendocrine, cardiovascular, and inflammatory stress responses in middle-aged men and women. Psychoneuroendocrinology. 2004;29:593–611. doi: 10.1016/S0306-4530(03)00086-6. [DOI] [PubMed] [Google Scholar]
- 68.Eisenberger NI, et al. Does Rejection Hurt? An fMRI Study of Social Exclusion. Science. 2003;302:290–292. doi: 10.1126/science.1089134. [DOI] [PubMed] [Google Scholar]
- 69.Delgado MR, et al. An fMRI study of reward-related probability learning. Neuroimage. 2005;24:862–873. doi: 10.1016/j.neuroimage.2004.10.002. [DOI] [PubMed] [Google Scholar]
- 70.O'Doherty JP. Reward representations and reward-related learning in the human brain: insights from neuroimaging. Curr Opin Neurobiol. 2004;14:769–776. doi: 10.1016/j.conb.2004.10.016. [DOI] [PubMed] [Google Scholar]
- 71.Leyton M. Conditioned and sensitized responses to stimulant drugs in humans. Prog Neuropsychopharmacol Biol Psychiatry. 2007;31:1601–1613. doi: 10.1016/j.pnpbp.2007.08.027. [DOI] [PubMed] [Google Scholar]
- 72.Wang Z, et al. Neural Substrates of Abstinence-Induced Cigarette Cravings in Chronic Smokers. J Neurosci. 2007;27:14035–14040. doi: 10.1523/JNEUROSCI.2966-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Seymour B, et al. Differential encoding of losses and gains in the human striatum. J Neurosci. 2007;27:4826–4831. doi: 10.1523/JNEUROSCI.0400-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Aron A, et al. Reward, motivation, and emotion systems associated with early-stage intense romantic love. J Neurophysiol. 2005;94:327–337. doi: 10.1152/jn.00838.2004. [DOI] [PubMed] [Google Scholar]
- 75.Rilling J, et al. A Neural Basis for Social Cooperation. Neuron. 2002;35:395–504. doi: 10.1016/s0896-6273(02)00755-9. [DOI] [PubMed] [Google Scholar]
- 76.Fliessbach K, et al. Social Comparison Affects Reward-Related Brain Activity in the Human Ventral Striatum. Science. 2007;318:1305–1308. doi: 10.1126/science.1145876. [DOI] [PubMed] [Google Scholar]
- 77.De Quervain DJF, et al. The Neural Basis of Altruistic Punishment. Science. 2004;305:1254–1258. doi: 10.1126/science.1100735. [DOI] [PubMed] [Google Scholar]
- 78.Cacioppo JT, et al. In the Eye of the Beholder: Individual Differences in Perceived Social Isolation Predict Regional Brain Activation to Social Stimuli. J Cogn Neurosci. 2009;21:1–10. doi: 10.1162/jocn.2009.21007. [DOI] [PMC free article] [PubMed] [Google Scholar]