

Abstract
Background
Endocannabinoids are novel lipid mediators with hypotensive and cardiodepressor activity. Here, we examined the possible role of the endocannabinergic system in cardiovascular regulation in hypertension.
Methods and Results
In spontaneously hypertensive rats (SHR), cannabinoid-1 receptor (CB1) antagonists increase blood pressure and left ventricular contractile performance. Conversely, preventing the degradation of the endocannabinoid anandamide by an inhibitor of fatty acid amidohydrolase reduces blood pressure, cardiac contractility, and vascular resistance to levels in normotensive rats, and these effects are prevented by CB1 antagonists. Similar changes are observed in 2 additional models of hypertension, whereas in normotensive control rats, the same parameters remain unaffected by any of these treatments. CB1 agonists lower blood pressure much more in SHR than in normotensive Wistar-Kyoto rats, and the expression of CB1 is increased in heart and aortic endothelium of SHR compared with Wistar-Kyoto rats.
Conclusions
We conclude that endocannabinoids tonically suppress cardiac contractility in hypertension and that enhancing the CB1-mediated cardiodepressor and vasodilator effects of endogenous anandamide by blocking its hydrolysis can normalize blood pressure. Targeting the endocannabinoid system offers novel therapeutic strategies in the treatment of hypertension.
Keywords: hypertension, blood pressure, contractility, endocannabinoids, pharmacology
Hypertension is a major health problem and, when untreated predisposes to cardiovascular morbidity and premature death.1 Effective treatment of hypertension reduces complications and improves life expectancy; however, side effects of available antihypertensive medications and differences in their efficacy in preventing end-organ damage justify the search for antihypertensive agents that act through novel mechanisms of action.
Δ9-Tetrahydrocannabinol, the psychoactive ingredient of marijuana, lowers blood pressure and heart rate in experimental animals,2,3 and hypotension has been reported after chronic marijuana use in humans.4 The endogenous cannabinoid ligands arachidonoyl ethanolamide (anandamide) and 2-arachidonoylglycerol (2-AG) also lower blood pressure and heart rate in rodents.5,6 Cannabinoids interact with G protein–coupled receptors to produce their effects. To date, 2 such receptors have been identified: cannabinoid-1 receptors (CB1), expressed at high levels in the brain7 but also present in peripheral tissues, including the heart8 and the vasculature,9,10 and CB2, expressed by immune and hematopoietic cells.11 Cannabinoids fail to lower blood pressure after selective blockade of CB15 or in CB1-knockout mice,12,13 which implicates CB1 in this effect.
Treatment of normotensive rats and mice with CB1 antagonists alone does not affect blood pressure,3,5 and baseline blood pressure is similar in CB1-knockout mice and their wild-type littermates,12,13 which indicates that CB1 receptors are not tonically active. This is also suggested by the lack of hypotension after inhibition of anandamide transport,14 in agreement with the relatively modest hypotensive effect of anandamide in normotensive animals.3,5 However, an increase in the hypotensive efficacy of cannabinoids has been noted in spontaneously hypertensive rats (SHR).15,16 Therefore, we examined whether endogenous cannabinoids may have a cardiovascular regulatory function in hypertension. Three different models of experimental hypertension were used to reduce the possibility of detecting changes limited to a given model rather than to hypertension itself. The results document tonic activation of cardiac and vascular CB1 in hypertension that limits increases in blood pressure and cardiac contractility. They also indicate that upregulation of CB1 is responsible for this tone and that increasing it by inhibiting the inactivation of endogenous anandamide can normalize blood pressure and cardiac contractile performance in hypertension.
Methods
Materials
Anandamide, the CB1 receptor antagonist AM251, and the anandamide transport inhibitors AM404 and OMDM-2 were from Tocris; the fatty acid amidohydrolase inhibitor URB597 was from Cayman Chemicals; and the CB1 antagonist SR141716 and the CB2 antagonist SR144528 were from the National Institute on Drug Abuse drug supply program. The synthetic CB agonist HU-2103 was a gift from R. Mechoulam (Hebrew University, Jerusalem, Israel). Drugs were mixed in corn oil and sonicated for 5 minutes at 4°C. The mixture was added to 4 parts of Pluronic F68 (Sigma-Aldrich) solution (40 mg/mL) dissolved in water and sonicated to obtain a stable suspension for bolus intravenous injections.
Animals
Rats were obtained from Harlan (Indianapolis, Ind). Male, 8- to 10-month-old SHR, age-matched male Wistar Kyoto rats (WKY), and 8- to 10-week-old male Sprague-Dawley rats were maintained on standard rat chow and water ad libitum. Dahl salt-sensitive and salt-resistant rats (male, Rapp strain, 6 weeks old) were maintained for 4 weeks on rat chow containing either 0.12% or 8% NaCl. Systolic blood pressure monitored daily by the tail-cuff technique was 120±11 mm Hg (salt-sensitive, 0.12% NaCl), 180±14 mm Hg (salt-sensitive, 8% NaCl), and 118±9 mm Hg (salt-resistant, 8% NaCl). Hypertension was induced in Sprague-Dawley rats by chronic infusion of angiotensin II (60 ng/min) via an osmotic minipump, as described previously.17 Rats were used 10 to 12 days after implantation of the minipump. The Institutional Animal Care and Use Committee had approved all procedures.
Hemodynamic Measurements
Rats were anesthetized with pentobarbital sodium (60 mg/kg IP) and tracheotomized to facilitate breathing. Animals were placed on controlled heating pads, and core temperature, measured via a rectal probe, was maintained at 37°C. A microtip pressure-volume catheter (SPR-838; Millar Instruments) was inserted into the right carotid artery and advanced into the left ventricle (LV) under pressure control as described previously.18,19 Polyethylene cannulas (P50) were inserted into the right femoral artery and vein for measurement of mean arterial pressure (MAP) and administration of drugs, respectively. After stabilization for 20 minutes, signals were continuously recorded with an ARIA pressure-volume conductance system (Millar) coupled to a Powerlab/4SP A/D converter (AD Instruments), stored, and displayed on a computer. Heart rate, maximal LV systolic pressure, MAP, and maximal slope of systolic pressure increment (+dP/dt) were computed with a cardiac pressure-volume analysis program (PVAN2.9; Millar). Cardiac output calculated and corrected according to in vitro and in vivo volume calibrations with PVAN2.918 was normalized to body weight (cardiac index [CI]). Total peripheral resistance index (TPRI) was calculated by the equation TPRI=MAP/CI. In 3 experiments, drugs were microinjected into the fourth cerebral ventricle as described previously.20
Immunohistochemistry
Affinity purified CB1 polyclonal antibody was raised in rabbit against the first 100 amino acids of human CB1.21 Formalin-fixed, paraffin-embedded aortic sections deparaffinized and rehydrated in PBS were preincubated in blocking buffer (1% BSA, 0.6% Triton-X-100 in PBS, pH 7.4) for 1 hour, then incubated with the primary antibody at 24°C for 1 hour and then at 4°C for 24 hours. Sections were washed, and endogenous peroxidase activity was suppressed with 3% H2O2 in PBS for 10 minutes. Sections were washed 3 times in PBS and incubated in biotinylated secondary antibody (goat-anti-rabbit IgG 1:1000; Vector) and in ABC reagents according to the manufacturer’s instructions. Color was developed in 3,3′-diaminobenzidine and staining visualized by light microscopy and quantified by densitometry. Specificity of the reaction was controlled by preabsorption of the antibodies with 1 to 10 μg/mL of the immunizing peptide.
Western Blotting
Frozen myocardial tissue from SHR and WKY was minced and homogenized in ice-cold lysis buffer (50 mmol/L Tris, pH 7.5, 1% Nonidet P-40, 0.25% sodium deoxycholate, 150 mmol/L NaCl, 1 mmol/L each of EDTA, phenylmethylsulfonyl fluoride, and sodium orthovanadate, and 1 μg/mL each of aprotinin, leupeptin, and pepstatin). One hundred micrograms of lysate protein was size-fractionated by 10% SDS-PAGE and transblotted to a nitrocellulose membrane. Western blotting, with rabbit anti-human CB1 polyclonal antibody21 at 5 μg/mL or a rabbit antibody against human fatty acid amidohydrolase (FAAH; Alpha Diagnostics), was done as described previously.22 Immunoreactive bands were visualized with an enhanced chemiluminescence reaction (Amersham Pharmacia) and quantified by densitometry.
Measurement of Endocannabinoid Levels
Tissue levels of anandamide and 2-AG were quantified by liquid chromatography/in-line mass spectrometry, as described in detail elsewhere.23 Values are expressed as femtomoles or picomoles per milligram of wet tissue.
Statistical Analyses
Time-dependent variables were analyzed by ANOVA followed by the Dunnett post hoc test. In other cases, the Student t test was used, as appropriate. Values with P<0.05 were considered statistically significant.
Results
CB1 Blockade Unmasks Cannabinoid-Mediated Cardiovascular Depression in Hypertensive Rats
In pentobarbital-anesthetized normotensive WKY, intravenous injection of the CB1 antagonist SR141716 (3 mg/kg) had no effect on blood pressure or on any of the other hemodynamic parameters measured (Figure 1, left). In contrast, in SHR, the same treatment elicited marked and sustained further increase in blood pressure with no change in heart rate, and there was a parallel increase in myocardial contractility as deduced from the increases in dP/dt and LV systolic pressure. CI was also increased. These effects of SR141716 were mediated peripherally, because intracerebroventricular microinjection of 100 μg of SR141716 had no effect on blood pressure (from 165±7 to 173±15 mm Hg) or the other hemodynamic parameters (data not shown). The dramatic difference in the cardiac response to CB1 blockade is also evident in the LV pressure/volume relationship, which was unaffected by SR141716 in WKY (Figure 2a), whereas in SHR, a leftward displacement indicating increased contractile performance was elicited by SR141716 (Figure 2b) or by another CB1 antagonist, AM251 (Figure 2c). In contrast, treatment of SHR with the CB2 antagonist SR144528 (3 mg/kg) failed to influence any hemodynamic parameters (not shown).
Figure 1.
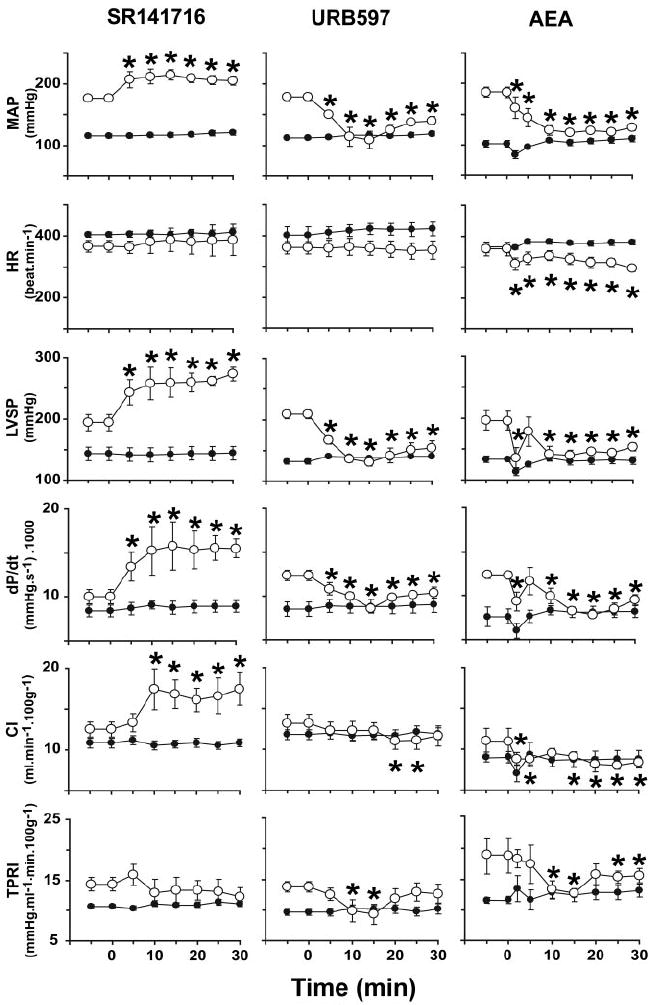
Hemodynamic effects of CB1 antagonist SR141716 (3 mg/kg), FAAH antagonist URB597 (10 mg/kg), and anandamide (AEA; 10 mg/kg) in WKY (●) and SHR (○). Drugs were injected at 0 minutes. Values are mean±SEM. *P<0.05 of corresponding baseline values (n=4 to 10 for each condition). HR indicates heart rate.
Figure 2.
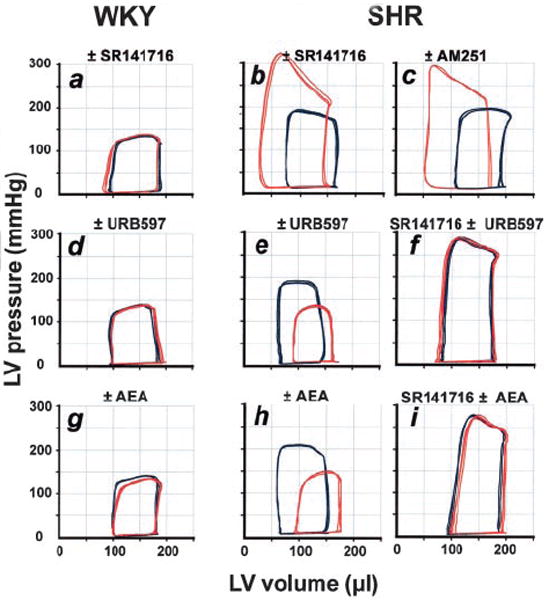
Representative LV pressure-volume loops from WKY (a, d, g) and SHR (b, c, e, f, h, i) before (blue) and after (red) treatment with indicated agents or their combinations (for doses, see text). Experiments were repeated in 3 more animals in each treatment group with similar results. AEA indicates anandamide.
A pressor response to CB1 blockade was also evident in 2 other models of hypertension. AM251 did not affect blood pressure in normotensive Sprague-Dawley rats but caused a prolonged pressor response in rats with angiotensin II–induced hypertension (Figure 3b). In salt-sensitive Dahl rats, SR141716 had no effect on mean blood pressure in normotensive animals kept on a low-salt diet but elicited a pressor response in animals made hypertensive by an 8% NaCl-containing diet. Salt-resistant Dahl rats kept on the same 8% NaCl diet remained normotensive and unresponsive to SR141716 (Figure 4).
Figure 3.
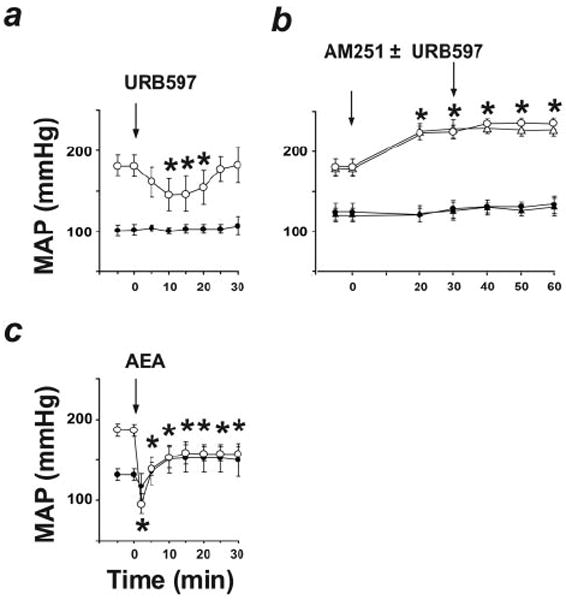
Effects of URB597 (a); AM251 followed by vehicle (circles) or URB597 (triangles; b), and anandamide (AEA; c) on MAP in normotensive rats (solid symbols) and angiotensin II–induced hypertensive rats (open symbols). Values are mean±SEM. *P<0.05 of baseline values (n=4 to 7 for each condition). Note that AM251 blocks the hypotensive effect of subsequently administered URB597 in SHR (b).
Figure 4.
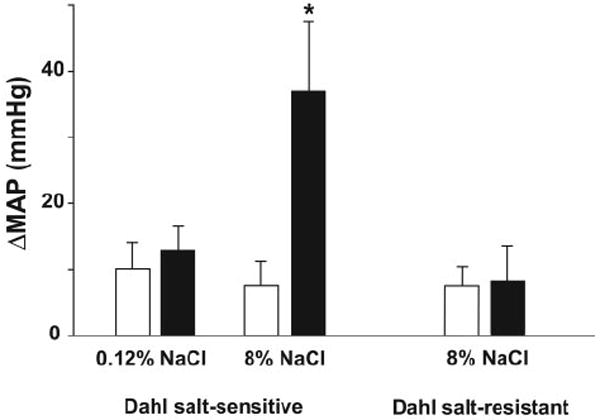
Change in MAP after vehicle (open columns) or 3 mg/kg SR141716 (solid columns) in Dahl salt-sensitive rats fed on 0.12% and 8% salt diet and in Dahl salt-resistant rats on 8% salt diet. Values are mean±SEM. *P<0.05 of baseline values (n=4 to 5 for each condition). Basal MAP after anesthesia was 97±5, 118±6, and 98±4 mm Hg in the 3 groups, respectively.
Potentiation of CB1 Activity by Inhibition of Endocannabinoid Inactivation
URB597, an inhibitor of FAAH that degrades anandamide in vivo,24 increases brain levels of anandamide but not 2-AG and unmasks tonic analgesia mediated by CB1.25 Treatment of WKY with URB597 (10 mg/kg IV) had no detectable hemodynamic effects, whereas in SHR, URB597 decreased arterial pressure to normotensive levels for >30 minutes and also decreased LV systolic pressure, dP/dt, and TPRI (Figure 1, middle panels; Figure 2d). We used a maximally effective dose of URB597 based on dose-response studies (n=3), which established its hypotensive ED50 at 1.7 mg/kg.
A URB597-induced decrease in cardiac contractility in SHR was also indicated by the change in pressure/volume relationship (Figure 2e). URB597 similarly lowered blood pressure in angiotensin II–treated hypertensive rats but not in their normotensive controls (Figure 3a). The effects of URB597 in the hypertensive animals could be prevented by CB1 antagonists (Figure 2f and Figure 3b). MAP of SHR was also decreased by treatment with the anandamide transport inhibitors AM404 (10 mg/kg, −63±19 mm Hg, n=4) or OMDM-2 (5 mg/kg, −39±10 mm Hg, n=4),26 which increase anandamide levels at the receptor by blocking its cellular uptake, whereas no significant change was observed in WKY.
Increased Sensitivity to CB1 Agonists in Hypertensive Rats
The effects of URB597 were remarkably similar to those of exogenous anandamide. In WKY, anandamide (10 mg/kg) caused a modest and short-lasting (<5 minutes) decrease in blood pressure and cardiac contractility, without other hemodynamic changes (Figure 1, right panels). In SHR, the same dose of anandamide caused a sustained decrease in blood pressure to near normotensive levels, accompanied by decreases in heart rate, cardiac contractility (dP/dt, LV systolic pressure), CI, and TPRI (Figure 1). The cardiac pressure-volume effects of anandamide were similar to those of URB597 (Figure 2g through 2i), and anandamide-induced hypotension was potentiated in rats with angiotensin II–induced hypertension (Figure 3c). The effects of the synthetic CB agonist HU210 were similarly potentiated, with its hypotensive EC50 reduced from 5.9 to 1.5 μg/kg and its maximal hypotensive effect increased from −39±14 to −108±11 mm Hg in WKY (n=6) versus SHR (n=6, P<0.01), respectively.
Because anandamide is a known ligand for vanilloid receptors (VR1),27 we tested the ability of the VR1 antagonist capsazepine to inhibit the hypotensive response to URB597 or anandamide in SHR. As illustrated in Figure 5, the hypotensive response to either agent was not affected by capsazepine pretreatment.
Figure 5.
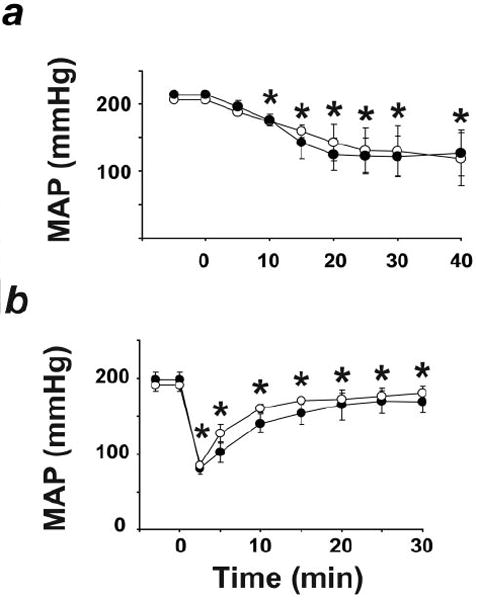
Hypotensive effect of URB597 (a) or anandamide (b) in SHR is unaffected by VR1 antagonist capsazepine. URB597 or anandamide (10 mg/kg) was injected at 0 minutes and 10 minutes after injection of vehicle (●) or 5 mg/kg capsazepine (○). *P<0.05 of corresponding baseline values. Values are mean±SEM, n=5 to 6 in each group.
Upregulation of Cardiac and Vascular Endothelial CB1 in SHR
The expression of CB1 in the myocardium and aorta of SHR and WKY was analyzed by immunohistochemistry and Western blotting with a CB1 antibody. CB1 expression was significantly greater in both cardiac tissue and the aortic endothelium of SHR than in WKY (Figure 6).
Figure 6.
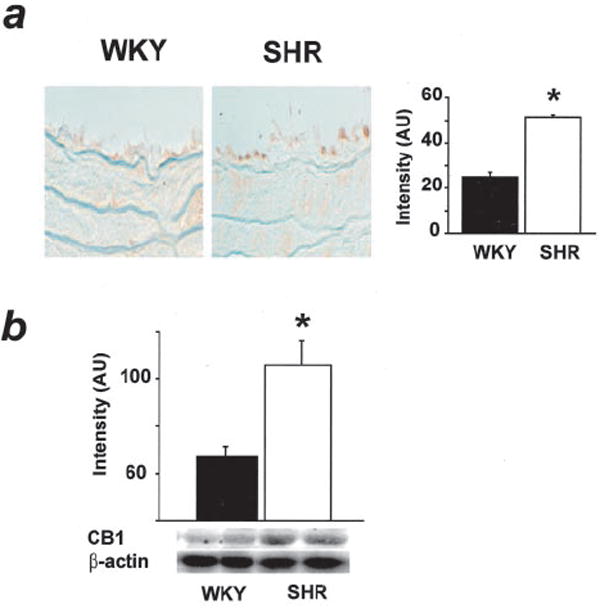
CB1 receptor immunohistochemistry in aorta (a) and Western blot of cardiac CB1 (b) in WKY and SHR. Values are mean±SE from 4 separate experiments. *P<0.05, WKY vs SHR.
Endocannabinoid Levels and FAAH Expression in WKY and SHR
In myocardium, FAAH expression was unexpectedly increased and anandamide levels correspondingly decreased in SHR compared with WKY (Figure 7), with no difference in 2-AG levels. However, URB597 treatment caused a greater increase in myocardial anandamide levels in SHR than in WKY(Figure 7). Interestingly, a similar increase was observed when myocardial anandamide levels in SHR myocardium were determined under control conditions (10.8±0.4 fmol/mg, n=3) and 10 minutes after the intravenous injection of 10 mg/kg anandamide, ie, at the time of the peak hypotensive response (14.9±1.3 fmol/mg, n=4, P<0.05). Plasma levels of anandamide were not different at the same time points (1.43±0.05 versus 1.53±0.1 pmol/mL), and the URB597-induced increase in plasma anandamide (1.78±0.20 versus 2.44±0.20 pmol/mL, or 1.3-fold) was also smaller than the increase observed in the myocardium (2.2-fold; Figure 7b).
Figure 7.
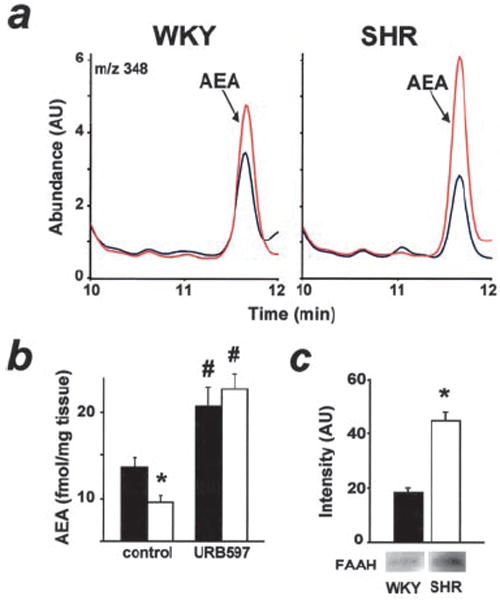
a, Representative ion chromatogram of untreated (blue) and URB597-treated (red) hearts from WKY rats (left) and SHR (right). b, Myocardial anandamide content of untreated and URB597-treated WKY (solid bar) and SHR (open bar; n=4 each group). *P<0.05, WKY vs SHR; #P<0.05 untreated vs URB597-treated rats. c, Western blot of cardiac FAAH (n=3).
Discussion
The present findings provide evidence for a novel cardiovascular regulatory mechanism mediated by endocannabinoids acting at CB1, stimulation of which lowers blood pressure through reductions in both cardiac contractility and vascular resistance. In various forms of hypertension, this endocannabinoid system becomes tonically active, probably owing to an upregulation of cardiac and vascular endothelial CB1. Furthermore, increased activation of CB1 through blocking of the metabolic degradation or intracellular uptake of the endocannabinoid anandamide normalizes blood pressure, offering a novel approach for the pharmacotherapy of hypertension.
CB1 antagonists did not affect blood pressure in normotensive rats, and inhibitors of FAAH or anandamide transport were similarly ineffective, which indicates that the cannabinoid system is inactive under normotensive conditions. In contrast, CB1 antagonists elicited sustained further increases in blood pressure in rats with 3 different forms of hypertension, which suggests that the elevated basal blood pressure itself is responsible for the tonic activation of CB1. Interestingly, CB1 antagonists increased cardiac contractile performance without significantly affecting peripheral vascular resistance, which suggests that the primary target of endocannabinoids in hypertension is the heart. The lack of change in heart rate further suggests that they decrease contractility directly rather than through inhibition of sympathetic tone. Correspondingly, inhibition of FAAH by URB597 decreased cardiac contractility without affecting heart rate. However, URB597 reduced both cardiac contractility and peripheral vascular resistance, and these effects are remarkably similar to those of exogenous anandamide, except that anandamide also caused bradycardia. This reduction in vascular resistance suggests that CB1 effects on vascular tone require higher levels of endocannabinoids than does suppression of cardiac contractile performance, which could result from differences in the relative concentration of endogenous ligand and receptor in the 2 tissues. URB597 preferentially increases anandamide rather than 2-AG levels in the brain,25 and the same was shown here for the heart. Thus, anandamide or a related fatty acid amide, rather than 2-AG, may be responsible for the activation of CB1 in hypertension.
CB1 receptors in the vasculature9,10 mediate vasodilation,9,28 and in the myocardium, they mediate negative inotropy,8 and both of these sites may be implicated in the hypotensive effect of anandamide.29 Additionally, presynaptic CB1 receptors on sympathetic nerve terminals inhibit norepinephrine release30 and may mediate anandamide-induced bradycardia.29 The lack of bradycardia after URB597 may be related to low tissue levels of anandamide, FAAH, or both at cardiac sympathetic nerve terminals. Central administration of SR141716 at a dose that blocks cannabinoid-induced behavioral effects31 caused no hemodynamic changes in SHR, which rules out a central site of action.
Anandamide can elicit vasodilation13,32-34 and decreased cardiac contractility35 through an additional Gi/Go-coupled receptor distinct from CB1 or CB2, which is inhibited by SR14171613 but not by other CB1 antagonists such as AM251.35,36 In the present experiments, AM251 and SR141716 were equally effective in eliciting pressor and cardiostimulatory responses and in blocking the depressor effects of URB597 in hypertensive rats. This indicates that their likely target is CB1 rather than the non-CB1/non-CB2 receptor described above. The vanilloid VR1 receptor can also mediate anandamide-induced vasodilation27,37; however, it is not involved in the effects described here, because SR141716, which does not affect VR1-mediated vasodilation by capsaicin,13 completely blocked the effect of URB597 (Figure 2f), and the VR1 antagonist capsazepine failed to influence the hypotensive response to URB597 or anandamide (Figure 5). Furthermore, we found no difference in the hypotensive response to anandamide in VR1−/− mice and their controls, and SR141716 completely blocked the effect in both.38
Responses to CB1 agonists and URB597 were greatly potentiated in hypertensive rats, which suggests increased target-organ sensitivity rather than increased endocannabinoid levels as the underlying mechanism. Indeed, an increase in CB1 expression was evident in both the myocardium and the aortic endothelium of SHR versus WKY, although receptor expression in the aorta may not reflect that in resistance vessels. Additional changes in the coupling of CB1 to signaling pathways, such as may result from an upregulation of inhibitory G proteins,39 are also possible. Tissue levels of endocannabinoids were unchanged or even reduced in SHR compared with WKY, although the greater rise in myocardial anandamide after blockade of FAAH in SHR may also contribute to their increased cardiovascular response to the FAAH antagonist.
The increased reactivity of CB1 in hypertension can be exploited therapeutically. Direct activation of CB1 by plantderived or synthetic agonists is unacceptable because of psychotropic effects. However, by blocking the inactivation of endocannabinoids, a more selective action may be achieved due to the distinct tissue distribution of FAAH. Indeed, in a recent study, URB597 elicited CB1-mediated anxiolytic and analgesic responses without causing other cannabinoid-like effects, such as hypothermia and catalepsy.25 Although a higher dose of URB597 was required to reduce blood pressure in SHR, the effect was completely blocked by CB1 antagonists, and URB597 was ineffective in normotensive rats. This suggests that the action of URB597 is fully accounted for by endocannabinoid-mediated stimulation of CB1.
The elevated systolic performance of the hypertensive ventricle has been attributed to a hypertrophic response that compensates for the increased wall stress.40 Blocking endocannabinoid inactivation reduces both the elevated arterial pressure and the increased LV contractile performance without affecting the same parameters in normotensive animals. This offers an innovative approach to the pharmacotherapy of hypertension by simultaneously targeting and normalizing the inappropriately increased LV contractile performance and vascular tone.
References
- 1.Lew EA. High blood pressure, other risk factors and longevity: the insurance viewpoint. Am J Med. 1973;55:281–294. doi: 10.1016/0002-9343(73)90130-7. [DOI] [PubMed] [Google Scholar]
- 2.Vollmer RR, Cavero I, Ertel RJ, et al. Role of the central autonomic nervous system in the hypotension and bradycardia induced by (-)-delta 9-trans-tetrahydrocannabinol. J Pharm Pharmacol. 1974;26:186–198. doi: 10.1111/j.2042-7158.1974.tb09252.x. [DOI] [PubMed] [Google Scholar]
- 3.Lake KD, Compton DR, Varga K, et al. Cannabinoid-induced hypotension and bradycardia in rats mediated by CB1-like cannabinoid receptors. J Pharmacol Exp Ther. 1997;281:1030–1037. [PubMed] [Google Scholar]
- 4.Benowitz NL, Jones RT. Cardiovascular effects of prolonged delta-9-tetrahydrocannabinol ingestion. Clin Pharmacol Ther. 1975;18:287–297. doi: 10.1002/cpt1975183287. [DOI] [PubMed] [Google Scholar]
- 5.Varga K, Lake K, Martin BR, et al. Novel antagonist implicates the CB1 cannabinoid receptor in the hypotensive action of anandamide. Eur J Pharmacol. 1995;278:279–283. doi: 10.1016/0014-2999(95)00181-j. [DOI] [PubMed] [Google Scholar]
- 6.Járai Z, Wagner JA, Goparaju SK, et al. Cardiovascular effects of 2-arachidonoyl glycerol in anesthetized mice. Hypertension. 2000;35:679–684. doi: 10.1161/01.hyp.35.2.679. [DOI] [PubMed] [Google Scholar]
- 7.Matsuda LA, Lolait SJ, Brownstein MJ, et al. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature. 1990;346:561–564. doi: 10.1038/346561a0. [DOI] [PubMed] [Google Scholar]
- 8.Bonz A, Laser M, Kullmer S, et al. Cannabinoids acting on CB1 receptors decrease contractile performance in human atrial muscle. J Cardiovasc Pharmacol. 2003;41:657–664. doi: 10.1097/00005344-200304000-00020. [DOI] [PubMed] [Google Scholar]
- 9.Gebremedhin D, Lange AR, Campbell WB, et al. Cannabinoid CB1 receptor of cat cerebral arterial muscle functions to inhibit L-type Ca2+ channel current. Am J Physiol. 1999;266:H2085–H2093. doi: 10.1152/ajpheart.1999.276.6.H2085. [DOI] [PubMed] [Google Scholar]
- 10.Liu J, Gao B, Mirshahi F, et al. Functional CB1 cannabinoid receptors in human vascular endothelial cells. Biochem J. 2000;346:835–840. [PMC free article] [PubMed] [Google Scholar]
- 11.Munro S, Thomas KL, Abu-Shaar M. Molecular characterization of a peripheral receptor for cannabinoids. Nature. 1993;365:61–65. doi: 10.1038/365061a0. [DOI] [PubMed] [Google Scholar]
- 12.Ledent C, Valverde O, Cossu G, et al. Unresponsiveness to cannabinoids and reduced addictive effects of opiates in CB1 receptor knockout mice. Science. 1999;283:401–404. doi: 10.1126/science.283.5400.401. [DOI] [PubMed] [Google Scholar]
- 13.Járai Z, Wagner JA, Varga K, et al. Cannabinoid-induced mesenteric vasodilation through an endothelial site distinct from CB1 or CB2 receptors. Proc Natl Acad Sci U S A. 1999;96:14136–14141. doi: 10.1073/pnas.96.24.14136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Calignano A, La Rana G, Beltramo M, et al. Potentiation of anandamide hypotension by the transport inhibitor, AM404. Eur J Pharmacol. 1997;337:R1–R2. doi: 10.1016/s0014-2999(97)01297-1. [DOI] [PubMed] [Google Scholar]
- 15.Kosersky DS. Antihypertensive effects of delta9-tetrahydrocannabinol. Arch Int Pharmacodyn. 1978;233:76–81. [PubMed] [Google Scholar]
- 16.Lake KD, Martin BR, Kunos G, et al. Cardiovascular effects of anandamide in anesthetized and conscious normotensive and hypertensive rats. Hypertension. 1997;29:1204–1210. doi: 10.1161/01.hyp.29.5.1204. [DOI] [PubMed] [Google Scholar]
- 17.Imig JD, Zhao X, Capdevila JH, et al. Soluble epoxide hydrolase inhibition lowers arterial blood pressure in angiotensin II hypertension. Hypertension. 2002;39:690–694. doi: 10.1161/hy0202.103788. [DOI] [PubMed] [Google Scholar]
- 18.Pacher P, Liaudet L, Bai P, et al. Potent metalloporphyrin peroxynitrite decomposition catalyst protects against the development of doxorubicin-induced cardiac dysfunction. Circulation. 2003;107:896–904. doi: 10.1161/01.cir.0000048192.52098.dd. [DOI] [PubMed] [Google Scholar]
- 19.Pacher P, Liaudet L, Mabley J, et al. Pharmacologic inhibition of poly-(adenosine diphosphate-ribose) polymerase may represent a novel therapeutic approach in chronic heart failure. J Am Coll Cardiol. 2002;40:1006–1016. doi: 10.1016/s0735-1097(02)02062-4. [DOI] [PubMed] [Google Scholar]
- 20.Ramirez-Gonzalez MD, Tchakarov L, Mosqueda Garcia R, et al. Beta-endorphin acting on the brainstem is involved in the antihypertensive action of clonidine and alpha-methyldopa in rats. Circ Res. 1983;53:150–157. doi: 10.1161/01.res.53.2.150. [DOI] [PubMed] [Google Scholar]
- 21.Wager-Miller J, Westenbroek R, Mackie K. Dimerization of G protein-coupled receptors: CB1 cannabinoid receptors as an example. Chem Phys Lipids. 2002;121:83–89. doi: 10.1016/s0009-3084(02)00151-2. [DOI] [PubMed] [Google Scholar]
- 22.Liu J, Tian Z, Gao B, et al. Dose-dependent activation of antiapoptotic and proapoptotic pathways by ethanol treatment in human vascular endothelial cells: differential involvement of adenosine. J Biol Chem. 2002;277:20927–20933. doi: 10.1074/jbc.M110712200. [DOI] [PubMed] [Google Scholar]
- 23.Wang L, Liu J, Harvey-White J, et al. Endocannabinoid signaling via cannabinoid receptor 1 is involved in ethanol preference and its age-dependent decline in mice. Proc Natl Acad Sci U S A. 2003;100:1393–1398. doi: 10.1073/pnas.0336351100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cravatt BF, Demarest K, Patricelli MP, et al. Supersensitivity to anandamide and enhanced endogenous cannabinoid signaling in mice lacking fatty acid amide hydrolase. Proc Natl Acad Sci U S A. 2001;98:9371–9376. doi: 10.1073/pnas.161191698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kathuria S, Gaetani S, Fegley D, et al. Modulation of anxiety through blockade of anandamide hydrolysis. Nat Med. 2003;9:76–81. doi: 10.1038/nm803. [DOI] [PubMed] [Google Scholar]
- 26.De Lago E, Ligresti A, Ortar G, et al. In vivo pharmacological actions of two novel inhibitors of anandamide cellular uptake. Eur J Pharmacol. 2004;484:249–257. doi: 10.1016/j.ejphar.2003.11.027. [DOI] [PubMed] [Google Scholar]
- 27.Zygmunt PM, Petersson J, Andersson DA, et al. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature. 1999;400:452–457. doi: 10.1038/22761. [DOI] [PubMed] [Google Scholar]
- 28.Deutsch DG, Goligorsky MS, Schmid PC, et al. Production and physiological actions of anandamide in the vasculature of the rat kidney. J Clin Invest. 1997;100:1538–1546. doi: 10.1172/JCI119677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wagner JA, Járai Z, Bátkai S, et al. Hemodynamic effects of cannabinoids: coronary and cerebral vasodilation mediated by cannabinoid CB(1) receptors. Eur J Pharmacol. 2001;423:203–210. doi: 10.1016/s0014-2999(01)01112-8. [DOI] [PubMed] [Google Scholar]
- 30.Ishac EJ, Jiang L, Lake KD, et al. Inhibition of exocytotic noradrenaline release by presynaptic cannabinoid CB1 receptors on peripheral sympathetic nerves. Br J Pharmacol. 1996;118:2023–2028. doi: 10.1111/j.1476-5381.1996.tb15639.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gomez R, Navarro M, Ferrer B, et al. A peripheral mechanism for CB1 cannabinoid receptor-dependent modulation of feeding. J Neurosci. 2002;22:9612–9617. doi: 10.1523/JNEUROSCI.22-21-09612.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mukhopadhyay S, Chapnick BM, Howlett AC, et al. Anandamide-induced vasorelaxation in rabbit aortic rings has two components: G protein dependent and independent. Am J Physiol. 2002;282:H2046–H2054. doi: 10.1152/ajpheart.00497.2001. [DOI] [PubMed] [Google Scholar]
- 33.Offertáler L, Mo FM, Batkai S, et al. Selective ligands and cellular effectors of a G protein-coupled endothelial cannabinoid receptor. Mol Pharmacol. 2003;63:699–705. doi: 10.1124/mol.63.3.699. [DOI] [PubMed] [Google Scholar]
- 34.Bukoski RD, Batkai S, Jarai Z, et al. CB(1) receptor antagonist SR141716A inhibits Ca(2+)-induced relaxation in CB(1) receptor-deficient mice. Hypertension. 2002;39:251–257. doi: 10.1161/hy0202.102702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ford WR, Honan SA, White R, et al. Evidence of a novel site mediating anandamide-induced negative inotropic and coronary vasodilatator responses in rat isolated hearts. Br J Pharmacol. 2002;135:1191–1198. doi: 10.1038/sj.bjp.0704565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ho WS, Hiley V. Vasodilator actions of abnormal-cannabidiol in rat isolated small mesenteric artery. Br J Pharmacol. 2003;138:1320–1332. doi: 10.1038/sj.bjp.0705160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Li J, Kaminski NE, Wang DH. Anandamide-induced depressor effect in spontaneously hypertensive rats: role of the vanilloid receptor. Hypertension. 2003;41:757–762. doi: 10.1161/01.HYP.0000051641.58674.F7. [DOI] [PubMed] [Google Scholar]
- 38.Pacher P, Bátkai S, Kunos G. Haemodynamic profile and responsiveness to anandamide of TRPV1 receptor knock-out mice. J Physiol (London) 2004;558:647–657. doi: 10.1113/jphysiol.2004.064824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Anand-Srivastava MB. Enhanced expression of inhibitory guanine nucleotide regulatory protein in spontaneously hypertensive rats: relationship to adenylate cyclase inhibition. Biochem J. 1992;288:79–85. doi: 10.1042/bj2880079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Cingolani OH, Yang XP, Cavasin MA, et al. Increased systolic performance with diastolic dysfunction in adult spontaneously hypertensive rats. Hypertension. 2003;41:249–254. doi: 10.1161/01.hyp.0000052832.96564.0b. [DOI] [PubMed] [Google Scholar]