

Abstract
In mammals, ten Toll like receptors (TLR) recognize conserved pathogen associated molecular patterns (PAMP), resulting in the induction of inflammatory innate immune responses. One of these, TLR9, is activated intracellularly by bacterial DNA and synthetic oligodeoxynucleotides (ODN) containing unmethylated CpG dinucleotides. Following treatment with CpG ODN, TLR9 is found in LAMP-1 positive lysosomes and we asked which intracellular compartment contains TLR9 prior to CpG exposure. Surprisingly, we found by microscopy and supporting biochemical evidence that both transfected and endogenously expressed human TLR9 is retained in the endoplasmic reticulum (ER). By contrast, human TLR4 trafficked to the cell surface, indicating that ER retention is not a property common to all TLRs. Since TLR9 is observed in endocytic vesicles following exposure to CpG ODN, our data indicate that a special mechanism must exist for translocating TLR9 to the signaling compartments that contain the CpG DNA.
Keywords: Human, B lymphocytes, Toll-Like Receptors
Introduction
The Toll like receptors (TLRs) comprise a family of at least 10 proteins that recognize conserved patterns on microorganisms known as pathogen associated molecular patterns (PAMPs) (1). Exposure of TLRs to PAMPs triggers inflammatory processes that result in enhanced uptake and the direct killing of microorganisms as well as the generation of primary adaptive immune responses (2,3). Mice deficient in MyD88 and humans lacking IRAK4, two components immediately downstream of TLRs in their signaling pathways, are highly susceptible to infection, demonstrating the critical role that TLRs play in host responses to pathogens (4,5). Genetic deletion or reconstitution experiments have demonstrated that individual TLRs are both essential and sufficient for specific responsiveness to a wide variety of PAMPs; for example TLR2 is the receptor for lipopeptides, TLR3 for dsRNA, TLR4 for LPS, and TLR5 for flagellin. TLR9 recognizes bacterial and viral DNA and studies using synthetic single stranded oligodeoxynucleotides (ODNs) have defined the immunogenic sequences of bacterial DNA as consisting of unmethylated CpG motifs in the context of species-dependent surrounding sequences (6,7). TLR9 is expressed primarily on antigen presenting cells such as B cells and dendritic cells (DC)2. In human DC, TLR9 is restricted to a subset of DC, plasmacytoid DC, responsible for production of high levels of type I interferon (8). Phagocytes endocytose microorganisms and lyse them in phagolysosomes, where their DNA is released and presumably interacts with TLR9, initiating an inflammatory response. In systemic autoimmune diseases, the stimulation of TLR9 in B cells by endocytosed immune complexes containing hypomethylated CpG DNA has been hypothesized to play a deleterious role in the induction of autoantibodies (9,10). Thus, the recognition of DNA by TLR9 plays important roles in both host defense and autoimmunity.
Several lines of evidence suggest that the molecular recognition of CpG DNA occurs inside the cell, perhaps in lysosomes. First, in contrast to other TLRs, TLR9 has not been detected on the surface of cells (11,12). In addition, chloroquine and other inhibitors of endosome maturation inhibit signaling by CpG DNA (11,13), and MyD88 is recruited to vesicles containing the lysosomal marker, LAMP-1 in cells stimulated with CpG DNA (11). Finally, TLR9 and CpG DNA localize to a common intracellular compartment (12). Studies using fluorescently labeled ODNs indicate that both stimulatory and non-stimulatory DNA are internalized non-specifically by surface receptors, but only the stimulatory DNA activates TLR9 in the intracellular compartment where both co-localize (11,12). In this paper we address the important question of where TLR9 is located within the cell prior to CpG exposure. We show that transfected and endogenous TLR9 are retained intracellularly in a reticular compartment that we identify as the endoplasmic reticulum (ER). We also propose mechanisms that may explain how TLR9 gets from the ER to the signaling compartment.
Materials and Methods
Cell Culture
HeLa and HEK293T cells (ATCC, Manassas, VA) were cultured in DMEM containing 2 mM L-glutamine, 50 U/ml penicillin, 50 μg/ml streptomycin, and 10% low endotoxin fetal bovine serum. B cell lines (gifts from Dr. Art Shaffer, NCI, Bethesda, MD) were cultured in RPMI 1640 containing the same supplements.
Antibodies
Anti-Grp94 and anti-KDEL antibodies were from Stratagene (San Diego, CA), anti-calnexin from Stressgen (Victoria, B.C.), anti-hemagglutinin (HA) epitope (clone 3F10) and anti-HA HRP (clone 12CA5) were from Roche (Indianapolis IN), and anti-HA-Alexa 488 (clone HA1.1) was from Covance (Berkeley, CA). PE-conjugated anti-TLR4 (HTA125) and unconjugated and PE-anti-TLR9 (eB72-1665) were from eBioscience (San Diego, CA). Alexa 488 and Alexa 546 conjugated secondary antibodies (for microscopy) were from Molecular Probes (Eugene, OR) and APC secondary antibodies (for flow cytometry) were from Caltag (Burlingame, CA).
Transfections and constructs
HeLa and HEK293T cells were transfected using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) and calcium phosphate precipitation, respectively. Human TLR9 and TLR4 tagged at their C-termini with enhanced green fluorescent protein (TLR9-GFP, TLR4-GFP), yellow fluorescent protein (TLR9-YFP, TLR4-YFP), or cyan fluorescent protein (TLR9-CFP) were generated by inserting the TLR in frame into pEGFP-N1, pEYFP-N1, or pECFP-N1, respectively (BD Clontech, Palo Alto, CA). HA-tagged TLR9 was generated by inserting TLR9, lacking its leader sequence but retaining its stop codon, in frame into pDisplay (Invitrogen). All TLR fusion proteins were shown to activate NF-κB in response to CpG (for TLR9) or LPS (for TLR4/MD-2) in HEK293 cells. CFP-tagged constructs used to identify organelles were pECFP-ER and pECFP-Golgi (BD Clontech), pECFP-C1 Rab5, 7, 9, and 11 (gifts from Dr. Juan Bonifacino, NICHD, Bethesda, MD). Monomeric red fluorescent protein (RFP)-tagged CD63 was generated by excising CD63 from CD63-GFP (a gift from Dr. Paul Roche, NCI, Bethesda, MD) and ligating it into a monomeric RFP vector (14) modified by Dr. Larry Samelson (NCI, Bethesda, MD).
Flow Cytometry
HEK293T cells were transfected in 6 well plates and after 48 hours stained directly for surface expression, or fixed, permeabilized (using CalTag Fix and Perm) and stained for total expression, using a directly conjugated fluorescent antibody. For human B cell lines, cells were fixed, permeabilized and labeled with anti-TLR9 followed by an Alexa 488 secondary antibody. Data were collected with a FACSCalibur cytometer (BD Immunocytometry Systems, San Jose, CA), and analyzed using FlowJo (Tree Star, Ashland, OR).
Confocal Microscopy
HEK293T cells were transfected in 6 well plates and 48 hours later were plated in HBSS on LabTek II (Campbell, CA) coverglass chambers precoated with poly-L-lysine. After 1 hour, the medium was changed to growth medium containing 10mM HEPES and 100 μg/ml cycloheximide for 1 hour before visualization. HeLa cells were transfected on coverslips, fixed, permeabilized and stained. BJAB cells were fixed, permeabilized, and stained for calnexin and TLR9 using anti-calnexin and anti-TLR9 Abs and counter-stained with Alexa 546 anti-rabbit and Alexa-488 anti-rat secondary Abs, respectively. In parallel, cells were stained with isotype control antibodies. These control cells were used to set the background levels for image collection. All cells were visualized using a Zeiss LSM510 laser scanning confocal microscope. Images were processed for presentation in figures using Adobe Photoshop 6.0 (Adobe Systems, San Jose, Ca).
Glycosidase Treatment
HEK293 cells stably expressing HA-TLR9 or BJAB cells were lysed as described (11) and immunoprecipitated with anti-HA or anti-TLR9 mAbs, respectively. The immunoprecipitates were treated with Endoglycosidase H (EndoH) or Peptide: N-Glycosidase F (PNGaseF) according to manufacturer instructions (New England Biolabs, Beverly, MA). Treated samples were separated by SDS-PAGE, transferred to nitrocellulose and probed with anti-HA or anti-TLR9.
Pulse Chase
After transfection with untagged TLR9, 5×105 HeLa cells were pulsed with 0.2 mCi 35S-methionine for 30 minutes in methionine free medium. The cells were washed twice with culture medium supplemented with 300 mg/L methionine and chased for various times in culture medium with 75 mg/L methionine. Cells were harvested in PBS with 20 mM EDTA, lysed, immunoprecipitated with anti-TLR9 mAb, and treated with glycosidase before separation by SDS-PAGE. Gels were fixed, treated with Enlightening (NEN Life Science, Boston, MA) and exposed to film for 48 hours.
Results
TLR9 is retained intracellularly
Although most TLRs are expressed at the cell surface, recent studies have shown that TLR9 is located in intracellular compartments. To determine the extent of intracellular localization of endogenously expressed TLR9, permeabilized or non-permeabilized Ramos B cells were stained with an anti-TLR9 mAb and examined by flow cytometry. As seen in Fig. 1A, the permeabilized cells (“Total”) stained positive for TLR9, but no TLR9 was detected on the cell surface (non-permeabilized cells). We then analyzed the expression of TLR4-GFP and TLR9-GFP in transiently transfected HEK293T cells. Figure 1B shows that only TLR4 was surface labeled with a PE-conjugated mAb, whereas both TLR4 and TLR9 were labeled with mAbs in permeabilized cells. Finally, HEK293T cells were transfected with TLR4-YFP or TLR9-YFP and examined live by laser scanning confocal (LSC) microscopy. After treatment with cycloheximide to block protein synthesis and allow newly synthesized protein to reach its final destination, TLR4-YFP was observed on the cell surface and in a perinuclear area, most likely the Golgi. In contrast, TLR9-YFP remained in an intracellular compartment (Figure 1C) even after a 24-hour incubation with cycloheximide (data not shown). Thus, TLR9 is retained intracellularly and is not detected on the cell surface in either TLR9 transfectants or in cells endogenously expressing TLR9.
Figure 1.

TLR9 is retained intracellularly, TLR4 traffics to the cell surface.
A and B) Flow cytometric analyses. A) Ramos B cells were stained directly with an anti-TLR9 mAb (Surface) or were fixed and permeabilized before staining (Total). Shaded histograms represent cells stained with an isotype-matched control mAb. B) HEK293T cells transfected with TLR4-GFP (upper panels) or TLR9-GFP (lower panels) were stained for surface or total expression as in panel A using PE-conjugated anti-TLR4 or anti-TLR9 mAbs. C) Laser scanning confocal (LSC) microscopy analysis. Live HEK293T cells transfected with TLR4-YFP (left panels) or TLR9-YFP (right panels) were treated with 100 μg/ml cycloheximide for 1 hour prior to visualization to block protein synthesis. A single section (0.8 μm) is shown for YFP (upper panels) and as an overlay of YFP with the differential interference contrast image (lower panels). Scalebar = 4.5 μm
TLR9 is located in the ER
To identify the compartment(s) to which TLR9 localizes, HeLa cells, co-transfected with fluorescently tagged TLR9 and markers for selected vesicles and organelles, were examined by LSC microscopy. TLR9 localized to a reticular compartment that did not contain Rab5, an early endosome marker, Rab7 or Rab9, late endosome markers, Rab11, a recycling endosome marker, CD63, a lysosome marker, or a commercial Golgi marker (Figure 2 and data not shown). In stark contrast, TLR9 co-localized with a commercial ER marker (Figure 2). In additional experiments, HEK293 cells stably expressing HA-TLR9 also showed reticular staining for TLR9. In these cells the TLR9 localized to a compartment that was stained with antibodies to the ER-resident KDEL motif, the ER-retained Hsp90 family member, Grp94, and MHC Class II invariant chain, which is retained in the ER in the absence of the αand βchains (Figure 3A and data not shown).
Figure 2.

TLR9 co-localizes with ER markers
HeLa cells were co-transfected with TLR9-YFP and either Rab5-CFP (early endosome), Rab7-CFP (late endosome), or Rab11-CFP (recycling endosome); with TLR9-GFP and CD63-mRFP (lysosome); and with TLR9-CFP and pEYFP-ER (a commercial marker for ER). Cells were fixed, mounted and examined by LSC microscopy. Single 0.8 μm sections pseudocolored green (TLR9, left panels) and red (organelle marker, middle panels) are shown. An overlay of the two channels is shown in the right panels. Scalebar = 9 μm
Figure 3.
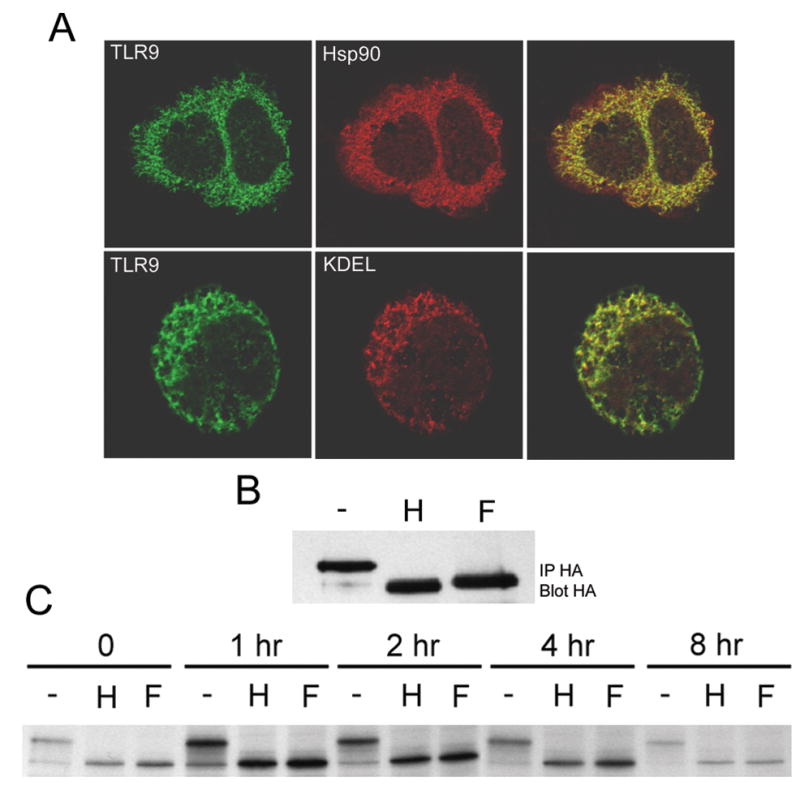
Stably expressed TLR9 resides in the ER.
A) HEK293 cells stably expressing HA-TLR9 were stained for TLR9 (with anti-HA Alexa 488) and the ER markers Hsp90 (upper panels, two connected cells) and KDEL (lower panels) using Alexa 546 secondary antibodies. Single 0.8 μm sections from LSC microscopy are shown for TLR9 (left panels) and ER markers (middle panels). Overlays are shown in the right panels. Scalebar = 9μm. B and C) TLR9 does not enter the Golgi.
B) HA immunoprecipitates (from the same cells as in A) were incubated with medium alone (−), EndoH (H) or PNGase F (F). The proteins were analyzed by SDS-PAGE and immunoblotting using an anti-HA-HRP mAb. C) Untagged TLR9, labeled with 35S-methionine and chased for the indicated times, was immunoprecipiated and treated with glycosidase as in B. Shown is the autoradiograph of the SDS-PAGE gel.
After leaving the ER, most newly synthesized proteins enter the Golgi stack where their N-linked carbohydrates are processed and become resistant to digestion with EndoH (15). We have previously shown that TLR4 contains an EndoH resistant form that is located on the cell surface (16). In contrast, HA-TLR9 is sensitive to both EndoH and PNGaseF (an enzyme that digests both immature and mature N-linked carbohydrates), at steady state (Figure 3B) and for at least eight hours following synthesis (Figure 3C). Although previous studies have indicated that TLR9 localizes in vesicles after treatment with CpG ODN, our current data show that TLR9 resides in the ER prior to treatment.
ER localization of endogenously expressed TLR9
Previous studies have shown that B cells express TLR9 and respond to TLR9-dependent ligands (6,17). To determine whether endogenously expressed TLR9 also localizes to the ER, we first analyzed a panel of B cell lines for TLR9 expression. By flow cytometry, BJAB expressed the highest level of TLR9 followed by Ramos, and Raji. JY expressed barely detectable levels of TLR9, and no TLR9 was detected in H929 or WIL2 (Figures 1A, 4A). Consistent with our earlier results, permeabilization was required for staining, indicating that TLR9 was located intracellularly as expected. Immunoprecipitation and blotting of TLR9 from the B cell lines showed a 160 kDa protein whose expression level correlated with flow cytometry results and was the correct molecular weight of the glycosylated form (Figure 4B). The anti-TLR9 antibody also detected a faint, slower migrating band that was also seen in the isotype control (not shown) and was therefore not specific. Because BJAB cells expressed the highest amount of TLR9, we used these cells to examine the intracellular location of TLR9. B cells contain a large nucleus and very little cytoplasm; therefore the different organelles are not resolved with high precision in these cells. However, by LSC microscopy, TLR9 gave a clearly discernable staining that colocalized with the ER marker calnexin (Fig 5A). In addition, TLR9 was sensitive to both EndoH and PNGaseF digestion (Figure 5B), resulting in the expected 113 kDa unglycosylated form, indicating that it did not reach the Golgi. Taken together, these data provide strong evidence that endogenously expressed TLR9, like transfected TLR9, resides solely in the ER in unstimulated cells.
Figure 4.

Endogenous TLR9 is retained intracellularly in B cell lines.
A) Four B cell lines (BJAB, Raji, WIL2 and JY) were permeabilized, labeled with anti-TLR9 antibody and stained with an Alexa 488 secondary antibody. Shaded profiles represent isotype controls. B) Endogenous TLR9 from the four B cell lines was immunoprecipated and analyzed by SDS-PAGE. After transferring to nitrocellulose, TLR9 was detected with the same antibody. In addition to the TLR9 band (arrow), a faint non-specific band migrating immediately above TLR9 is evident in all samples.
Figure 5.

Endogenous TLR9 is in the ER.
A) BJAB cells were stained for calnexin, an ER marker (red), and TLR9 (green). Shown is a 0.7 μm section of TLR9 (upper left), calnexin (upper right), an overlay of the two (lower left) and the differential contrast image (lower right). Scalebar =4.5 μm. B) TLR9 was immunoprecipitated from BJAB cells and digested with EndoH (H) or PNGaseF (F) as in 3B. The trace upper band in the EndoH digest corresponds to the non-specific band seen in Figure 4B.
Discussion
Recognition of bacterial DNA induces a potent innate immune response and affects the development of adaptive responses. It has been demonstrated that bacterial DNA or synthetic ODNs containing unmethylated CpG motifs are recognized only after endocytosis and not at the cell surface (10, 12). Recently, several studies have shown that TLR9 colocalizes with CpG ODN in a vesicular compartment, identified as a LAMP-1 positive lysosome, where signaling is initiated (11,12). What remained unclear was where TLR9 resides before stimulation. Here we show that both endogenously expressed and transfected TLR9 are retained in the ER.
In agreement with previous results, we were unable to detect TLR9 on the surface of transfected cells or B cell lines, although these same cells clearly expressed TLR9 intracellularly. When TLR9 was either labeled with a fluorescent mAb or fused to a fluorescent protein, it was located in a perinuclear reticular compartment that co-localized with ER markers, but not with markers specific for endocytic vesicles or, more importantly, lysosomes where it has been detected after stimulation with CpG ODN. Endogenously expressed TLR9 in B cells or ectopically expressed TLR9 in transfectants was sensitive to digestion with EndoH, an enzyme that digests immature N-linked sugars that are added in the ER but are modified to an EndoH resistant form in the Golgi. In fact, newly synthesized TLR9 remained sensitive to EndoH digestion even after 8 hours. Since most proteins traffic directly to the Golgi from the ER, the EndoH sensitivity of TLR9 in unstimulated cells suggests that the TLR9 has never left the ER. Although we have observed that TLR9 is located in the ER in several cell types, we cannot rule out that in other cells TLR9 may reside in a different cellular compartment. The precedence for this is TLR4, which has been shown to reside in a distinct compartment depending upon the cell type examined (18–20).
It is unclear how TLR9 reaches the lysosome where stimulation by CpG DNA occurs. One possibility is suggested by recent studies of Desjardins and colleagues (21,22) who demonstrated that the ER supplies membrane and ER proteins to phagosomes during particle engulfment. Since bacterial DNA is released in phagosomes following lysis, TLR9 might enter the phagosome directly from the ER. According to this hypothesis the ER is the predicted location of TLR9 in resting cells. In contrast to phagocytosis, soluble CpG DNA is taken up in non-phagocytic cells such as B cells, by receptor mediated and fluid phase endocytosis. This process also has important biological consequences; it stimulates responses that protect against pathogens (23) but may also contribute to systemic autoimmune diseases (9,10). Endocytosis of soluble molecules such as CpG DNA leads to their entry into early and late endosomes and lysosomes. Unlike phagosomes, these vesicles are formed from membranes that bud off from the plasma membrane or Golgi, and not by direct fusion with the ER. Nevertheless, the direct fusion between endosomes and the ER may occur by a previously unknown pathway. In favor of the direct pathway of ER to endosomes, we have not detected an EndoH resistant form of TLR9 following stimulation of cells with CpG ODN (C.A.L. unpublished result). Indeed, Latz et al (24) suggest that an EndoH sensitive form of TLR9 reaches early endosomes, where CpG DNA is localized soon after endocytosis. Biotinylation studies demonstrated that upon CpG DNA stimulation, a small amount of TLR9 becomes accessible to the extracellular environment, possibly by ER to endosome fusion. However, if a small fraction of TLR9 molecules trafficked to vesicles by the conventional pathway (via the Golgi), then our current studies, and those of Latz (24), may not be sensitive enough to detect the presence of mature sugars on TLR9. Thus, we cannot rule out the possibility that TLR9 may pass through the Golgi en route to the CpG containing vesicles by a conventional mechanism. Studies are currently underway to identify which mechanism(s) are involved.
Footnotes
Abbreviations used in this manuscript: TLR, Toll-like receptor; PAMP, Pathogen associated molecular pattern; ODN, oligodeoxynucleotide; ER, Endoplasmic reticulum; DC, Dendritic cell; LAMP-1, Lysosome-associated membrane protein type 1; GFP, Green fluorescent protein; EndoH, endoglycosidase H; PNGaseF, Peptide: N-Glycosidase F; LSC, Laser Scanning Microscopy.
Reference List
- 1.Takeda K, Kaisho T, Akira S. Toll-like Receptors. Annu Rev Immunol. 2003;21:335–376. doi: 10.1146/annurev.immunol.21.120601.141126. [DOI] [PubMed] [Google Scholar]
- 2.Kaisho T, Akira S. Regulation of dendritic cell function through Toll-like receptors. Curr Mol Med. 2003;3:373–385. doi: 10.2174/1566524033479726. [DOI] [PubMed] [Google Scholar]
- 3.Doyle SE, O’Connell RM, Miranda GA, Vaidya SA, Chow EK, Liu PT, Suzuki S, Suzuki N, Modlin RL, Yeh WC, Lane TF, Cheng G. Toll-like Receptors induce a phagocytic gene program through p38. J Exp Med. 2004;199:81–90. doi: 10.1084/jem.20031237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kawai T, Adachi O, Ogawa T, Takeda K, Akira S. Unresponsiveness of MyD88-deficient mice to endotoxin. Immunity. 1999;11:115–122. doi: 10.1016/s1074-7613(00)80086-2. [DOI] [PubMed] [Google Scholar]
- 5.Medvedev AE, Lentschat A, Kuhns DB, Blanco JC, Salkowski C, Zhang S, Arditi M, Gallin JI, Vogel SN. Distinct mutations in IRAK-4 confer hyporesponsiveness to lipopolysaccharide and interleukin-1 in a patient with recurrent bacterial infections. J Exp Med. 2003;198:521–531. doi: 10.1084/jem.20030701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Krieg AM, Yi AK, Matson S, Waldschmidt TJ, Bishop GA, Teasdale R, Koretzky GA, Klinman DM. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature. 1995;374:546–549. doi: 10.1038/374546a0. [DOI] [PubMed] [Google Scholar]
- 7.Lund J, Sato A, Akira S, Medzhitov R, Iwasaki A. Toll-like receptor 9-mediated recognition of Herpes simplex virus-2 by plasmacytoid dendritic cells. J Exp Med. 2003;198:513–520. doi: 10.1084/jem.20030162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hornung V, Rothenfusser S, Britsch S, Krug A, Jahrsdorfer B, Giese T, Endres S, Hartmann G. Quantitative expression of toll-like receptor 1–10 mRNA in cellular subsets of human peripheral blood mononuclear cells and sensitivity to CpG oligodeoxynucleotides. J Immunol. 2002;168:4531–4537. doi: 10.4049/jimmunol.168.9.4531. [DOI] [PubMed] [Google Scholar]
- 9.Leadbetter EA, I, Rifkin R, Hohlbaum AM, Beaudette BC, Shlomchik MJ, Marshak-Rothstein A. Chromatin-IgG complexes activate B cells by dual engagement of IgM and Toll-like receptors. Nature. 2002;416:603–607. doi: 10.1038/416603a. [DOI] [PubMed] [Google Scholar]
- 10.Viglianti GA, Lau CM, Hanley TM, Miko BA, Shlomchik MJ, Marshak-Rothstein A. Activation of autoreactive B cells by CpG dsDNA. Immunity. 2003;19:837–847. doi: 10.1016/s1074-7613(03)00323-6. [DOI] [PubMed] [Google Scholar]
- 11.Ahmad-Nejad P, Hacker H, Rutz M, Bauer S, Vabulas RM, Wagner H. Bacterial CpG-DNA and lipopolysaccharides activate Toll-like receptors at distinct cellular compartments. Eur J Immunol. 2002;32:1958–1968. doi: 10.1002/1521-4141(200207)32:7<1958::AID-IMMU1958>3.0.CO;2-U. [DOI] [PubMed] [Google Scholar]
- 12.Takeshita F, Leifer CA, Gursel I, Ishii KJ, Takeshita S, Gursel M, Klinman DM. Cutting edge: Role of Toll-like receptor 9 in CpG DNA-induced activation of human cells. J Immunol. 2001;167:3555–3558. doi: 10.4049/jimmunol.167.7.3555. [DOI] [PubMed] [Google Scholar]
- 13.Krieg AM. CpG motifs in bacterial DNA and their immune effects. Ann Rev Immunol. 2002;20:709–760. doi: 10.1146/annurev.immunol.20.100301.064842. [DOI] [PubMed] [Google Scholar]
- 14.Campbell RE, Tour O, Palmer AE, Steinbach PA, Baird GS, Zacharias DA, Tsien RY. A monomeric red fluorescent protein. Proc Natl Acad Sci USA. 2002;99:7877–7882. doi: 10.1073/pnas.082243699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Helenius A, Aebi M. Intracellular functions of N-linked glycans. Science. 2001;291:2364–2369. doi: 10.1126/science.291.5512.2364. [DOI] [PubMed] [Google Scholar]
- 16.Visintin A, Mazzoni A, Spitzer JA, Segal DM. Secreted MD-2 is a large polymeric protein that efficiently confers lipopolysaccharide sensitivity to Toll-like receptor 4. Proc Natl Acad Sci USA. 2001;98:12156–12161. doi: 10.1073/pnas.211445098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hemmi H, Takeuchi O, Kawai T, Kaisho T, Sato S, Sanjo H, Matsumoto M, Hoshino K, Wagner H, Takeda K, Akira S. A Toll-like receptor recognizes bacterial DNA. Nature. 2000;408:740–745. doi: 10.1038/35047123. [DOI] [PubMed] [Google Scholar]
- 18.Latz E, Visintin A, Lien E, Fitzgerald KA, Monks BG, Kurt-Jones EA, Golenbock DT, Espevik T. Lipopolysaccharide rapidly traffics to and from the Golgi apparatus with the toll-like receptor 4-MD-2-CD14 complex in a process that is distinct from the initiation of signal transduction. J Biol Chem. 2002;277:47834–47843. doi: 10.1074/jbc.M207873200. [DOI] [PubMed] [Google Scholar]
- 19.Hornef MW, Normark BH, Vandewalle A, Normark S. Intracellular recognition of lipopolysaccharide by toll-like receptor 4 in intestinal epithelial cells. J Exp Med. 2003;198:1225–1235. doi: 10.1084/jem.20022194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hornef MW, Frisan T, Vandewalle A, Normark S, Richter-Dahlfors A. Toll-like receptor 4 resides in the Golgi apparatus and colocalizes with internalized lipopolysaccharide in intestinal epithelial cells. J Exp Med. 2002;195:559–570. doi: 10.1084/jem.20011788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Desjardins M. ER-mediated phagocytosis: a new membrane for new functions. Nat Rev Immunol. 2003;3:280–291. doi: 10.1038/nri1053. [DOI] [PubMed] [Google Scholar]
- 22.Gagnon E, Duclos S, Rondeau C, Chevet E, Cameron PH, Steele-Mortimer O, Paiement J, Bergeron JJ, Desjardins M. Endoplasmic reticulum-mediated phagocytosis is a mechanism of entry into macrophages. Cell. 2002;110:119–131. doi: 10.1016/s0092-8674(02)00797-3. [DOI] [PubMed] [Google Scholar]
- 23.Krieg AM. CpG DNA: trigger of sepsis, mediator of protection, or both? Scand J Infect Dis. 2003;35:653–659. doi: 10.1080/00365540310015999. [DOI] [PubMed] [Google Scholar]
- 24.Latz E, Schoenemeyer A, Visintin A, Fitzgerald KA, Monks BG, Knetter CF, Lien E, Nilsen NJ, Espevik T, Golenbock DT. TLR9 signals after translocating from the ER to CpG DNA in the lysosome. Nat Immunol. 2004;5:190–198. doi: 10.1038/ni1028. [DOI] [PubMed] [Google Scholar]