

Abstract
The relationship between the blood-oxygenation-level dependent (BOLD) signal and its underlying neuronal activity is still inconclusive. The task of completely understanding this relationship has been encumbered not only by the complexity of neuronal responses, but also by nonlinear characteristics of the vascular response that are not correlated to neural activity. Repeated stimuli inducing replicable neural responses led to successively smaller BOLD amplitudes and delayed BOLD onset latencies when inter-stimulus intervals (ISIs) are shorter than 4-6 seconds, indicating significant nonlinearity between the BOLD signal and the underlying neuronal activity. We have provided evidence that large-vessel BOLD contributions could be the source of the nonlinearity (Zhang et al., Neuroimage 43: 204-12, 2008). By utilizing the spin-echo (SE) BOLD fMRI method at high magnetic fields to suppress large-vessel BOLD contributions, we found that the BOLD signal from the micro-vascular activity is replicable in response to replicated neuronal activities even at ISIs as short as ~1 second, suggesting that the micro-vascular BOLD activity is essentially a linear system. These results indicate that micro-vascular BOLD signals should provide an accurate estimate of the amplitude of neuronal activity changes. Consequently, the findings are important in understanding and resolving the controversy in the neurovascular coupling relationship. In addition, the difference in BOLD response times between macro- and micro-vascular activities demonstrated herein will have a significant impact on functional connectivity and causality studies. Moreover, SE fMRI at high fields, due to its capability of accurately representing the strength of neural activity as well as its previously shown high spatial specificity to activated brain regions, is an ideal choice for mapping brain function and quantifying the stimulus-evoked brain activity noninvasively.
Keywords: fMRI, neurovascular coupling, BOLD linearity, paired-stimulus paradigm, large vessel.
Introduction
Blood-oxygenation-level-dependent (BOLD) based functional magnetic resonance imaging (fMRI) has been widely used to study brain function (Bandettini et al., 1992; Kwong et al., 1992; Ogawa et al., 1992; Raichle and Mintun, 2006). This technique is based on locally coupled neuronal activity and hemodynamic changes (Ogawa et al., 1993). Although the fMRI technique shows promise for pinpointing and tracking brain activities, its potential is limited by the fact that the quantitative relationship between the BOLD signal and the underlying neural activity remains elusive (Logothetis et al., 2001; Smith et al., 2002). The task of completely understanding this relationship has been entangled not only by the complexity of various neuronal responses such as local field potential and spike activity, but also by various characteristics of vascular responses that are not directly correlated to neural activity (referred to non-neural vascular activity herein). This is because non-neural vascular activity can significantly vary the amplitude of the BOLD signal; albeit this variation does not represent a change of neural activity. For instance, identical neural activity may not be able to repeatedly trigger the same BOLD response due to a refractory effect of vascular response (Zhang et al., 2008b). As a result, different relationships regarding the neurovascular coupling can be observed based on the non-neural vascular components and whether or not they are accounted for and/or corrected when one tries to directly correlate the neuronal response to the BOLD signal.
One key issue of non-neural vascular response is the linearity of the BOLD signal which has remained an ongoing research topic for more than a decade (Boynton et al., 1996). This issue is vital in interpreting the amplitude of the BOLD signal since, if the BOLD response were a linear system, the fMRI signal could then simply be characterized by a hemodynamic-impulse-response function and this would yield a reliable measure of neuronal activity change. Nevertheless, the majority of studies have reported significant nonlinear effects in the BOLD signal (Birn and Bandettini, 2005; Birn et al., 2001; Boynton et al., 1996; Buxton et al., 1998; Cannestra et al., 1998; Dale and Buckner, 1997; Friston et al., 1998; Friston et al., 2000; Glover, 1999; Huettel and McCarthy, 2000; Inan et al., 2004; Janz et al., 2001; Pfeuffer et al., 2003; Robson et al., 1998; Soon et al., 2003; Vazquez and Noll, 1998; Wager et al., 2005; Zhang et al., 2008a; Zhang et al., 2008b). In these studies, repeated stimuli led to successively smaller BOLD amplitudes and delayed BOLD onset latencies when inter-stimulus intervals (ISIs) are shorter than a few seconds (Huettel and McCarthy, 2000; Zhang et al., 2008b). As a result, the strength of neural activity change evoked by brain stimulation cannot be accurately reflected by the corresponding BOLD amplitude. In addition, these nonlinear effects make it difficult to quantitatively compare fMRI responses among different states of brain activity and, thus, partially limit the applicability and potential in fMRI-related neuroscience research.
The mechanism underlying BOLD nonlinearity is poorly understood, partially because of its uncertain physiological origin. Birn and Bandettini have suggested that the observed BOLD nonlinearity could come from neuronal and/or vascular origins (Birn and Bandettini, 2005). By ensuring invariant neuronal activities in a paired-stimulus paradigm, we have selectively investigated vascular related (non)linear BOLD effects in the absence of any neuronal nonlinear effects based on the gradient-echo (GE) fMRI measurement (Zhang et al., 2008b). Under this circumstance, we demonstrated significant nonlinear BOLD effects including reduced BOLD amplitude and delayed BOLD onset latency in response to the second of paired stimuli when the ISI was shorter than 4-6 seconds. More importantly, we found BOLD nonlinearity was much less significant when the contributions from large vessels to the BOLD signal were partially reduced (Zhang et al., 2008b). These findings raise the question of whether or not the BOLD nonlinearity originates primarily from hemodynamic changes in large vessels which could be significantly distant from the actual neuronal activity site. The answer to this question is important because it would examine the possibility that the vascular response from the microvasculature, triggered by local neuronal activity, is a linear system and thus provides a precise measure of neural activity strength; whereas the nonlinear effects observed in the BOLD signal in previous studies are only artifacts from down-stream large vessels. Elucidation of this issue should have a profound impact on our understanding and modeling of the BOLD signal and its relationship with the underlying neuronal activity.
Unfortunately, our earlier study based on gradient-echo (GE) fMRI measurements is not sufficient to provide a definite answer to this question (Zhang et al., 2008b). This is because the criterion used to separate large vessel contributions to the BOLD signal was based on an index υ defined by the ratio between the standard deviation and the mean of the MRI signal intensity obtained from a series of fMRI images acquired during the resting condition. Since the MRI signal intensity at large vessels is relatively low, due to the short transverse relaxation time of blood water, and variation is high due to substantially large physiological noise, pixels located at large vessels tend to have larger υ values while pixels only containing microvasculature tend to have a smaller υ value. As a result, large-vessel contributions to the BOLD signal were reduced by eliminating activated pixels with large υ values. Although this criterion is sufficient to observe the effects of large vessels on the nonlinear characteristics of the BOLD signal, it is difficult to set a clear cut-off threshold to stringently separate pixels containing only large or small vessels because of the partial volume effect and thus it does not guarantee a strict segregation between macro- and micro-vascular activities. Consequently, residual nonlinearity was still observed in the BOLD signal after removing activated pixels with large υ values, though it became much less significant (Zhang et al., 2008b).
Spin-echo (SE) fMRI at high magnetic fields, on the other hand, is known to be primarily sensitive to micro-vascular activity (Boxerman et al., 1995; Cheng et al., 2001; Duong et al., 2003; Moon et al., 2007; Ogawa et al., 1993; Yacoub et al., 2003; Yacoub et al., 2008; Yacoub et al., 2007). This is true because the intravascular component of the BOLD signal is diminished at higher fields due to the dramatic shortening of blood T2 relative to tissue T2 (Thulborn et al., 1982). Furthermore, the extravascular BOLD signal from large vessels is eliminated using the SE fMRI method because the 180° radiofrequency (RF) pulse refocuses the static dephasing effects induced by magnetic field inhomogeneities around large vessels. Therefore, high-field SE fMRI signals originate largely from micro-vascular activities with high spatial specificity to the site of neuronal activation (Yacoub et al., 2008; Yacoub et al., 2007). Given this feature, in the present study we have investigated (non)linear characteristics of the BOLD signal from the microvasculature using a simple design of combining SE fMRI and a paired-stimulus paradigm composed of a pair of visual stimuli separated by an ISI between them. The ISIs selected were longer than 1 second to ensure invariant neuronal responses to all visual stimuli so that only the vascular effects were involved and investigated (Hoffken et al., 2008; Zhang et al., 2008a; Zhang et al., 2008b). Under these experimental settings we examined and compared the BOLD amplitude and onset latency in response to the second of paired stimuli to that of the single stimulus as a function of ISI.
To better illustrate our results and draw conclusions, we compared the data based on the SE BOLD signal to the data based on the GE BOLD signal adapted from our previous results (Zhang et al., 2008b), which were acquired under very similar experimental settings.
Materials and Methods
Subjects and Paradigm
Eight healthy subjects (four males, four females; 21-55 years old; mean± standard deviation: 32±14) participated in this study. The subjects provided informed consent which was approved by the Institutional Review Board of the University of Minnesota.
A paired-stimulus paradigm composed of a pair of visual stimuli with an ISI between them was used. Visual stimulation presented as 8 Hz flickering red/black light (100% contrast) was generated by a pair of red LED goggles (Grass Instruments, Quincy, MA). The duration of each stimulus was time locked to the repetition time (TR = 1.15 second) of echo-planar imaging (EPI) acquisition. Visual stimuli were displayed in the full visual field either singly or in pairs separated by an ISI of 1, 2, 4 or 8 TRs. Successive trials of single stimulus or paired stimuli were separated by sufficiently long inter-trial interval (ITI = 30 TRs, equivalent to 34.5 s) to allow the hemodynamic response to fully return to baseline and avoid any interference in vascular responses between consecutive stimulation trials. It also allowed us to individuate the BOLD response to the second of the paired stimuli by subtracting the BOLD response to the single stimulus from that to paired stimuli at each ISI. During the baseline condition, subjects were in uniform darkness.
Experiment procedure and data acquisition
All experiments were performed on a 4T/90 cm bore magnet (Oxford, UK) system interfaced with the Varian INOVA console (Varian Inc., Palo Alto, CA). A half-volume quadrature 1H RF surface coil was used to allow for sufficient RF field homogeneity in the visual cortex. Head motion was limited with a foam-padded holder. At the beginning of the experiment, axial, sagittal and coronal anatomic MR images were acquired by a conventional T1-weighted TurboFLASH method (Haase, 1990). On the basis of anatomic images, one coronal slice in the middle of the calcarine fissure was selected for acquiring fMRI data using SE EPI with the acquisition parameters: field of view (FOV) = 12.8×12.8 cm2, 64×64 in-plane matrix size, echo time (TE) = 67 ms, TR = 1150 ms, slice thickness = 5 mm, in-plane resolution = 2.0×2.0 mm2.
The fMRI experiment was conducted using an event-related task design. For each single or paired stimulation task, 10-12 trials were repeated in one run and two runs were repeated in each experiment. A total of ten fMRI runs corresponding to five tasks (1 single task and 4 paired tasks with different ISIs) were acquired in a pseudo-randomized order in each experiment. The subjects’ attention was maintained by asking them to count the number of trials, which was to be reported by the subject at the end of each run. The number of trials varied slightly and randomly across runs so that subjects could not predict the number for each run. The total trial number for each task across two runs was maintained to be approximately the same (n = 22).
Data analysis
The fMRI data analysis was performed using STIMULATE (Stimulate, Center for Magnetic Resonance Research, University of Minnesota) (Strupp, 1996) and Matlab (The Mathworks Inc., Natick, MA, USA). All fMRI time series were segmented from 5 time points before and 30 time points after the onset of stimulation. Epochs identified were selectively averaged for each task, resulting in five averaged time series for single- and paired-stimulus tasks.
Activation maps were generated using a time-shifted cross-correlation (CC) method (Bandettini et al., 1993; Xiong et al., 1995). A triangle function was used as the reference function for each stimulus. A relatively low threshold was used (CC coefficient > 0.3, p < 0.02) to create an fMRI activation map for each task. The final activation map was generated based on common activated pixels in all five fMRI maps obtained under the five tasks. This procedure yielded a very high threshold for the final map (p = 0.025 = 3×10-9) assuming acquisition was independent across different runs. A region of interest (ROI) was defined by all activated pixels at the proximity of the calcarine fissure in the final map for each subject. This ROI was later used for all tasks.
A ROI-based analysis was used to quantify BOLD (non)linearity. Time courses were averaged across all the activated pixels within the ROI for each task. The averaged time course was then calculated as a percentage change relative to the baseline condition. The time courses at paired-stimulus conditions in percentage change were subsequently normalized to the single-stimulus condition. This normalization method allows inter-subject averaging without needing to account for different BOLD amplitudes across subjects. After normalization, the time course at the single-stimulus condition was subtracted from the time course at each paired-stimulus condition, resulting in the BOLD response to the second stimulus for a given ISI. BOLD (non)linearity at each paired-stimulus condition was quantified by the ratio of BOLD integrals between the BOLD response to the second stimulus and the response to the single stimulus (Zhang et al., 2008b).
No temporal or spatial filter was applied in our analysis to avoid any type of bias that may affect the BOLD amplitude.
Results
Figure 1 shows an example of an SE fMRI activation map created using the procedure described in Materials and Methods from a representative subject. Activated pixels are primarily located along the cortical ribbon of gray matter (delineated with green lines) in the human visual cortex, consistent with the notion that the high-field SE fMRI signal mainly detects BOLD responses from micro-vascular activity without significant large-vessel contaminations.
Figure 1.
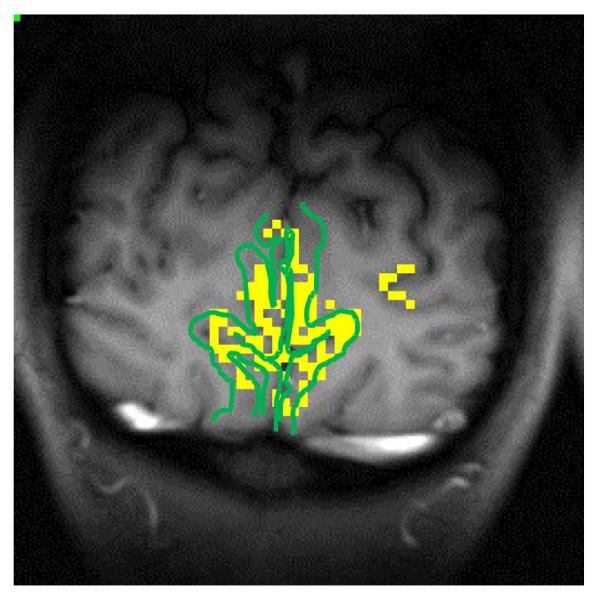
Spin-echo fMRI activation map created by the procedure described in Methods and Materials. Briefly, an fMRI activation map was created for each task using a time-shifted CC method and a relatively low CC coefficient. The final fMRI activation map was generated based on common activated pixels in all five fMRI maps obtained under varied tasks. The cortical ribbon of gray matter in the human visual cortex was delineated with green lines.
Figure 2a shows the time courses of SE BOLD responses to the single stimulus and the second of paired visual stimuli at different ISIs. The BOLD response to the second stimulus was individuated by subtracting the BOLD response to the single stimulus from that to paired stimuli at each ISI. To align the onset time of the second stimulus, the time course obtained after subtraction was shifted by the corresponding ISI. No obvious difference was observed between the time evolutions of BOLD responses to the single stimulus and second stimuli even at an ISI as short as ~1 s. This can be seen in the residual of the subtraction of the single BOLD response from the second BOLD response shown in Fig. 2b. After normalizing to the single-stimulus condition, the relative BOLD integral ratio between the responses to the second stimulus and the single stimulus as a function of ISI is shown in Fig. 3a. The dependency of the relative BOLD integral ratio on ISI based on the SE fMRI signal is in remarkable contrast to the GE BOLD signal shown in Fig. 3b (adapted from Figure 5a in (Zhang et al., 2008b)) and those reported in the literature (Huettel and McCarthy, 2000). The relative GE BOLD signal is characterized with reduced BOLD amplitude at short ISIs and a progressive recovery as ISI becomes longer, whereas the relative SE BOLD signal is essentially independent of ISI. Statistically, one-way ANOVA analysis grouping ISI shows that there is no significant difference in the relative BOLD integral ratio among ISIs for the SE BOLD signal (F = 1.79, p = 0.2), whereas the relative BOLD integral ratio based on GE BOLD signals strongly depends on ISI (F = 10.6, p < 0.001). Moreover, the relative BOLD integral ratios based on SE fMRI measurements are not statistically different from unity (or 1), indicating a negligible vascular refractory effect within the ISI range studied (p > 0.9, paired t-test). Figure 3b also demonstrates the effect of reducing large-vessel contributions to the GE BOLD signal on its nonlinear characteristics by setting different thresholds on the υ index. Clearly, BOLD nonlinearity is less significant (i.e. BOLD integral ratio is less dependent on ISI and closer to 1) when excluding pixels containing large vessels (υ < 0.005) compared to the case where there was no large vessel removal. This trend was more prominent at a threshold of υ < 0.003. Consistent with this trend, the BOLD nonlinearity virtually disappears when large-vessel contributions are removed by using SE fMRI at high magnetic field (Fig. 3a).
Figure 2.

(a) Time courses of the SE BOLD responses to the single stimulus and the second stimuli at all ISIs. The BOLD response to the second of paired stimuli was individuated by subtracting the BOLD response to the single stimulus from that to paired stimuli at each ISI. To align the onset of the second stimulus, the time course obtained after subtraction was shifted by the corresponding ISI. (b) The residue after subtracting the single BOLD response from the second BOLD responses at all ISIs.
Figure 3.

Dependency of relative BOLD integral ratio on ISI based on (a) SE fMRI signal (bars indicate one time standard deviation) and (b) GE fMRI signal (adapted from Fig. 5a in Zhang et al., Neuroimage 43: 204-12, 2008). In (b), it also shows the effect of reducing the contribution of large-vessel hemodynamic activity to the BOLD signal on its nonlinear characteristics by setting different thresholds on the υ index. BOLD nonlinearity is less significant (i.e. BOLD integral ratio is less dependent on ISI and closer to unity) after excluding pixels containing large vessels at a threshold of υ < 0.005. This trend is more prominent at a threshold of υ < 0.003
Besides the BOLD amplitudes at different ISIs, we also examined the onset latency delay between the second-stimulus evoked BOLD response during paired visual stimulation and the single-stimulus evoked BOLD response. All BOLD time courses were fitted to two Gamma functions used to account for the positive BOLD signal and undershoot, respectively. Figure 4a shows that the onset latencies of the GE BOLD signals in response to the second stimuli are significantly delayed at short ISIs; this latency delay is gradually shortened as ISI gets longer. By contrast, the onset latency of SE BOLD response is independent of ISI and not significantly different from zero (p = 0.60, paired two-tail t test). This result is also consistent with our earlier report that reducing the effect of the large-vessel component in the BOLD signal could weaken the dependency of BOLD latency delay on ISI and would thus result in less significant BOLD nonlinearity as shown in Figure 4b. Taken together, our results clearly demonstrate strong linear characteristics of the micro-vascular BOLD signal measured by SE fMRI at high field at ISIs as short as 1 second.
Figure 4.

The BOLD onset latency delay relative to the single-stimulus BOLD response based on the GE fMRI signal without vessel removal (red triangles) and SE fMRI signal (blue circles) in (a) and GE fMRI signal with the removal of large-vessel activities (black circles) in (b).
Discussion
In our previous work we investigated the source of vascular related (non)linear BOLD effects in the absence of any neuronal refractory effects (Zhang et al., 2008b). We found that excluding pixels containing large vessels significantly increased normalized BOLD amplitude and shortened BOLD onset latency when the vascular response was in the refractory period. In contrast, the procedure of reducing large vessel signals had little effect on BOLD activity at sufficiently long ISIs, where the vascular refractoriness disappears. These findings suggest the possibility that the vascular response from the microvasculature could be a linear system, whereas the nonlinear effects observed in the BOLD signal in previous studies might be simply artifacts from down-stream large vessels. If this is the case, the relative amplitude of local neuronal activity can be accurately estimated from the microvascular sensitive fMRI signal.
In order to explore this idea in the present study we examined (non)linear characteristics of the BOLD signal using SE fMRI acquisitions at a high magnetic field, where large-vessel contaminations in the BOLD signal are largely suppressed. Under this circumstance we observed completely different BOLD linearity characteristics. The SE BOLD responses to paired visual stimuli replicated each other even at a very short ISI of approximately 1s. Also as opposed to the case reported in the GE BOLD signal, SE BOLD responses to the second of paired stimuli did not demonstrate obvious signal onset latency delay with similar ISIs. Taken together, the SE BOLD signal, which is primarily sensitive to micro-vascular activity, reflects a highly linear system.
Significance in investigating the neurovascular coupling relationship
The remarkable contrast between GE and SE BOLD signals in terms of their linearity characteristics suggests significant difference between macro- and micro-vascular BOLD signals in response to brain stimulation. This difference might provide a clue for resolving the controversy in the quantitative relationship between the BOLD signal and the underlying neural activity (Logothetis et al., 2001; Smith et al., 2002). In order to compare the amplitudes of neurophysiologic (local field potential or spike activity) and BOLD signals, so as to elucidate the quantitative neurovascular coupling relationship, it would be ideal to use the BOLD component that more closely reflects variations in neural activity. Since the BOLD signal from macro-vascular hemodynamic activity contains prominent refractoriness that can completely be ascribed to vascular origin, correlating this BOLD component to the corresponding neural activity can yield arbitrary results and potentially misleading conclusions. On the other hand, micro-vascular BOLD signals show little or no vascular refractoriness, based on the results of the present and previous studies (Zhang et al., 2008b), and should provide a more reliable measure to study the neurovascular coupling relationship.
Influence on resting state functional connectivity and causality studies
In addition to the pronounced effects that large-vessel hemodynamic activities have on the nonlinear characteristics of the BOLD amplitude changes, there is also a significantly different temporal behavior between macro- and micro-vascular BOLD responses. This difference will have significant impact on functional connectivity and causality studies. Based on the results from this and earlier studies, micro-vascular activity seems to be prompt and varies proportionally as the status of the underlying neuronal activity changes. By contrast, macro-vascular activity is much more sluggish and demonstrates a substantial refractory period on the scale of 2-4 s. These results are consistent with several other studies demonstrating that large-vessel effects can cause the BOLD signal onset latency delays of up to 4 s relative to capillary effects (Lee et al., 1995). The difference between macro- and micro-vascular response times will affect the relative timing of the BOLD signals, thereby confounding the ability to infer neural connectivity and causality using the fMRI correlation method because of a fundamental assumption in most fMRI correlation studies that the relative timing of BOLD signals from different brain regions reflects the timing of their underlying neural activity (Chang et al., 2008). Therefore, it is critical to correct for vascular related BOLD onset latency differences when investigating connectivity and causality between different brain regions or within a brain region with distinct vascular architectures (Chang et al., 2008). In contrast, the SE BOLD signal at high magnetic fields should provide an ideal option to avoid this problem.
Possible Explanation
The empirical results observed in the present study manifest that the BOLD signal from the micro-vascular activities is virtually linear, whereas the nonlinearity primarily comes from macro-vascular responses. We postulate that this different BOLD characteristics in distinct vascular populations likely results from a mismatch of relative timing between micro-vascular and macro-vascular responses compared to the neuronal activity. The micro-vascular activity responds to the change of the underlying neuronal activity with consistent relative delay (i.e. micro-vascular BOLD onset latency). Consequently, micro-vascular BOLD signals elevated by repeated stimuli are coherent relative to the timing of the neuronal activity change. Since superposition of coherent signals maintains the relative amplitude variation as well as relative timing from each source, this feature makes the BOLD signal summed from micro-vascular activities contain the characteristics of linearity. On the other hand, macro-vascular activities are influenced by various up-stream micro-vascular responses with inconsistent relative delays owing to different transit times. As a result, the macro-vascular BOLD signal is not coherent relative to the neuronal activity. Particularly, at large vessels the second-stimulus BOLD response contributed by micro-vascular activities from certain sites can interfere with the first-stimulus BOLD responses from other micro-vascular sites if ISI is not sufficiently long. This notion can be supported by the coincidence that the refractory period of the macro-vascular activity is approximately 4 s (Zhang et al., 2008b) and that large-vessel effects can cause the BOLD signal onset latency delays of up to 4 s relative to capillary effects (Lee et al., 1995). This concept can also be applied to continuous stimulations. However, further investigation and evidences are needed to validate this notion.
Implications
The significance of the findings in this study is as follows. (i) The results indicate that the micro-vascular activity measured by the SE BOLD signal should provide an accurate estimate of the amplitude of neuronal activity changes. This characteristic has an impact on rapid event-related fMRI design, in which individual trials occur every few seconds before the BOLD signal returns to the baseline, and thus its application in numerous neuroscience fields. (ii) This study highlights the importance of separating the contributions from different vascular components when interpreting and modeling the BOLD signal, and investigating its relationship with the underlying neural activity. (iii) This study demonstrated differences in onset latency times between macro- and micro-vascular responses that will have an impact on functional connectivity and causality studies. (iv) Finally, SE fMRI at high fields, due to its capability to accurately represent both the amplitude of neuronal activity change as well as its spatial specificity to activated brain regions (Yacoub et al., 2008; Yacoub et al., 2007), should be an ideal choice for mapping brain function and quantifying the neuronal activity.
Although the results of the present study reveal that the micro-vascular BOLD signal is a highly linear system even at ISI as short as ~ 1 s, we have not tested the case when ISI is shorter than 1 s. Therefore it needs to be cautious when one intends to solve nonlinear BOLD problems in the short ISI range of < 1s. It also needs to be clarified that the linearity of micro-vascular BOLD signal does not necessarily imply a linear neurovascular coupling relationship (i.e. variation of neuronal activities is accompanied with a proportional change of hemodynamic activities). Instead, the data of the present study suggest that micro-vascular BOLD signals provide an accurate estimate of the amplitude of neuronal activity changes owing to the elimination of non-neural vascular component in the BOLD signal. In summary, the present study will help bridge the gap between the BOLD-based fMRI signal and the underlying neuronal activity; furthermore, they will advance the capability and accuracy of the fMRI approach for quantitatively studying brain function and dysfunction.
Acknowledgements
This work was supported in part by NIH grants of NS41262, EB00329, EB00513, MH70800, EB000331, P41 RR08079, and P30NS057091; the Keck foundation.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bandettini PA, Jesmanowicz A, Wong EC, Hyde JS. Processing strategies for time-course data sets in functional MRI of the human brain. Magn Reson Med. 1993;30:161–173. doi: 10.1002/mrm.1910300204. [DOI] [PubMed] [Google Scholar]
- Bandettini PA, Wong EC, Hinks RS, Tikofsky RS, Hyde JS. Time course EPI of human brain function during task activation. Magn Reson Med. 1992;25:390–397. doi: 10.1002/mrm.1910250220. [DOI] [PubMed] [Google Scholar]
- Birn RM, Bandettini PA. The effect of stimulus duty cycle and “off” duration on BOLD response linearity. Neuroimage. 2005;27:70–82. doi: 10.1016/j.neuroimage.2005.03.040. [DOI] [PubMed] [Google Scholar]
- Birn RM, Saad ZS, Bandettini PA. Spatial heterogeneity of the nonlinear dynamics in the FMRI BOLD response. Neuroimage. 2001;14:817–826. doi: 10.1006/nimg.2001.0873. [DOI] [PubMed] [Google Scholar]
- Boxerman JL, Bandettini PA, Kwong KK, Baker JR, Davis TL, Rosen BR, Weisskoff RM. The intravascular contribution to fMRI signal change: Monte Carlo modeling and diffusion-weighted studies in vivo. Magn Reson Med. 1995;34:4–10. doi: 10.1002/mrm.1910340103. [DOI] [PubMed] [Google Scholar]
- Boynton GM, Engel SA, Glover GH, Heeger DJ. Linear systems analysis of functional magnetic resonance imaging in human V1. J Neurosci. 1996;16:4207–4221. doi: 10.1523/JNEUROSCI.16-13-04207.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buxton RB, Wong EC, Frank LR. Dynamics of blood flow and oxygenation changes during brain activation: the balloon model. Magn Reson Med. 1998;39:855–864. doi: 10.1002/mrm.1910390602. [DOI] [PubMed] [Google Scholar]
- Cannestra AF, Pouratian N, Shomer MH, Toga AW. Refractory periods observed by intrinsic signal and fluorescent dye imaging. J Neurophysiol. 1998;80:1522–1532. doi: 10.1152/jn.1998.80.3.1522. [DOI] [PubMed] [Google Scholar]
- Chang C, Thomason ME, Glover GH. Mapping and correction of vascular hemodynamic latency in the BOLD signal. Neuroimage. 2008;43:90–102. doi: 10.1016/j.neuroimage.2008.06.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng K, Waggoner RA, Tanaka K. Human ocular dominance columns as revealed by high-field functional magnetic resonance imaging. Neuron. 2001;32:359–374. doi: 10.1016/s0896-6273(01)00477-9. [DOI] [PubMed] [Google Scholar]
- Dale AM, Buckner RL. Selective averaging of rapidly presented individual trials using fMRI. Hum Brain Mapp. 1997;5:329–340. doi: 10.1002/(SICI)1097-0193(1997)5:5<329::AID-HBM1>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- Duong TQ, Yacoub E, Adriany G, Hu X, Ugurbil K, Kim SG. Microvascular BOLD contribution at 4 and 7 T in the human brain: gradient-echo and spin-echo fMRI with suppression of blood effects. Magn Reson Med. 2003;49:1019–1027. doi: 10.1002/mrm.10472. [DOI] [PubMed] [Google Scholar]
- Friston KJ, Josephs O, Rees G, Turner R. Nonlinear event-related responses in fMRI. Magn Reson Med. 1998;39:41–52. doi: 10.1002/mrm.1910390109. [DOI] [PubMed] [Google Scholar]
- Friston KJ, Mechelli A, Turner R, Price CJ. Nonlinear responses in fMRI: the Balloon model, Volterra kernels, and other hemodynamics. Neuroimage. 2000;12:466–477. doi: 10.1006/nimg.2000.0630. [DOI] [PubMed] [Google Scholar]
- Glover GH. Deconvolution of impulse response in event-related BOLD fMRI. Neuroimage. 1999;9:416–429. doi: 10.1006/nimg.1998.0419. [DOI] [PubMed] [Google Scholar]
- Haase A. Snapshot FLASH MRI. Applications to T1, T2, and chemical-shift imaging. Magn Reson Med. 1990;13:77–89. doi: 10.1002/mrm.1910130109. [DOI] [PubMed] [Google Scholar]
- Hoffken O, Grehl T, Dinse HR, Tegenthoff M, Bach M. Paired-pulse behavior of visually evoked potentials recorded in human visual cortex using patterned paired-pulse stimulation. Exp Brain Res. 2008 doi: 10.1007/s00221-008-1374-0. [DOI] [PubMed] [Google Scholar]
- Huettel SA, McCarthy G. Evidence for a refractory period in the hemodynamic response to visual stimuli as measured by MRI. Neuroimage. 2000;11:547–553. doi: 10.1006/nimg.2000.0553. [DOI] [PubMed] [Google Scholar]
- Inan S, Mitchell T, Song A, Bizzell J, Belger A. Hemodynamic correlates of stimulus repetition in the visual and auditory cortices: an fMRI study. Neuroimage. 2004;21:886–893. doi: 10.1016/j.neuroimage.2003.10.029. [DOI] [PubMed] [Google Scholar]
- Janz C, Heinrich SP, Kornmayer J, Bach M, Hennig J. Coupling of neural activity and BOLD fMRI response: new insights by combination of fMRI and VEP experiments in transition from single events to continuous stimulation. Magn Reson Med. 2001;46:482–486. doi: 10.1002/mrm.1217. [DOI] [PubMed] [Google Scholar]
- Kwong KK, Belliveau JW, Chesler DA, Goldberg IE, Weisskoff RM, Poncelet BP, Kennedy DN, Hoppel BE, Cohen MS, Turner R, et al. Dynamic magnetic resonance imaging of human brain activity during primary sensory stimulation. Proc Natl Acad Sci U S A. 1992;89:5675–5679. doi: 10.1073/pnas.89.12.5675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee AT, Glover GH, Meyer CH. Discrimination of large venous vessels in time-course spiral blood-oxygen-level-dependent magnetic-resonance functional neuroimaging. Magn Reson Med. 1995;33:745–754. doi: 10.1002/mrm.1910330602. [DOI] [PubMed] [Google Scholar]
- Logothetis NK, Pauls J, Augath M, Trinath T, Oeltermann A. Neurophysiological investigation of the basis of the fMRI signal. Nature. 2001;412:150–157. doi: 10.1038/35084005. [DOI] [PubMed] [Google Scholar]
- Moon CH, Fukuda M, Park SH, Kim SG. Neural interpretation of blood oxygenation level-dependent fMRI maps at submillimeter columnar resolution. J Neurosci. 2007;27:6892–6902. doi: 10.1523/JNEUROSCI.0445-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ogawa S, Menon RS, Tank DW, Kim SG, Merkle H, Ellermann JM, Ugurbil K. Functional brain mapping by blood oxygenation level-dependent contrast magnetic resonance imaging. A comparison of signal characteristics with a biophysical model. Biophys J. 1993;64:803–812. doi: 10.1016/S0006-3495(93)81441-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ogawa S, Tank DW, Menon R, Ellermann JM, Kim SG, Merkle H, Ugurbil K. Intrinsic signal changes accompanying sensory stimulation: functional brain mapping with magnetic resonance imaging. Proc Natl Acad Sci U S A. 1992;89:5951–5955. doi: 10.1073/pnas.89.13.5951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeuffer J, McCullough JC, Van de Moortele PF, Ugurbil K, Hu X. Spatial dependence of the nonlinear BOLD response at short stimulus duration. Neuroimage. 2003;18:990–1000. doi: 10.1016/s1053-8119(03)00035-1. [DOI] [PubMed] [Google Scholar]
- Raichle ME, Mintun MA. Brain work and brain imaging. Annu Rev Neurosci. 2006;29:449–476. doi: 10.1146/annurev.neuro.29.051605.112819. [DOI] [PubMed] [Google Scholar]
- Robson MD, Dorosz JL, Gore JC. Measurements of the temporal fMRI response of the human auditory cortex to trains of tones. Neuroimage. 1998;7:185–198. doi: 10.1006/nimg.1998.0322. [DOI] [PubMed] [Google Scholar]
- Smith AJ, Blumenfeld H, Behar KL, Rothman DL, Shulman RG, Hyder F. Cerebral energetics and spiking frequency: the neurophysiological basis of fMRI. Proc Natl Acad Sci U S A. 2002;99:10765–10770. doi: 10.1073/pnas.132272199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soon CS, Venkatraman V, Chee MW. Stimulus repetition and hemodynamic response refractoriness in event-related fMRI. Hum Brain Mapp. 2003;20:1–12. doi: 10.1002/hbm.10122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strupp JP. Stimulate: A GUI based fMRI analysis software package. Neuroimage. 1996;3:S607. [Google Scholar]
- Thulborn KR, Waterton JC, Matthews PM, Radda GK. Oxygenation dependence of the transverse relaxation time of water protons in whole blood at high field. Biochim Biophys Acta. 1982;714:265–270. doi: 10.1016/0304-4165(82)90333-6. [DOI] [PubMed] [Google Scholar]
- Vazquez AL, Noll DC. Nonlinear aspects of the BOLD response in functional MRI. Neuroimage. 1998;7:108–118. doi: 10.1006/nimg.1997.0316. [DOI] [PubMed] [Google Scholar]
- Wager TD, Vazquez A, Hernandez L, Noll DC. Accounting for nonlinear BOLD effects in fMRI: parameter estimates and a model for prediction in rapid event-related studies. Neuroimage. 2005;25:206–218. doi: 10.1016/j.neuroimage.2004.11.008. [DOI] [PubMed] [Google Scholar]
- Xiong J, Gao JH, Lancaster JL, Fox PH. Clustered pixels analysis for functional MRI activation studies of the human brain. Hum Brain Mapp. 1995;3:287–301. [Google Scholar]
- Yacoub E, Duong TQ, Van De Moortele PF, Lindquist M, Adriany G, Kim SG, Ugurbil K, Hu X. Spin-echo fMRI in humans using high spatial resolutions and high magnetic fields. Magn Reson Med. 2003;49:655–664. doi: 10.1002/mrm.10433. [DOI] [PubMed] [Google Scholar]
- Yacoub E, Harel N, Ugurbil K. High-field fMRI unveils orientation columns in humans. Proc Natl Acad Sci U S A. 2008;105:10607–10612. doi: 10.1073/pnas.0804110105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yacoub E, Shmuel A, Logothetis N, Ugurbil K. Robust detection of ocular dominance columns in humans using Hahn Spin Echo BOLD functional MRI at 7 Tesla. Neuroimage. 2007;37:1161–1177. doi: 10.1016/j.neuroimage.2007.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang N, Liu Z, He B, Chen W. Noninvasive study of neurovascular coupling during graded neuronal suppression. J Cereb Blood Flow Metab. 2008a;28:280–290. doi: 10.1038/sj.jcbfm.9600531. [DOI] [PubMed] [Google Scholar]
- Zhang N, Zhu XH, Chen W. Investigating the source of BOLD nonlinearity in human visual cortex in response to paired visual stimuli. Neuroimage. 2008b;43:204–212. doi: 10.1016/j.neuroimage.2008.06.033. [DOI] [PMC free article] [PubMed] [Google Scholar]