

Abstract
Adults, preschool children, and nonhuman primates detect and categorize food objects according to substance information, conveyed primarily by color and texture. In contrast, they perceive and categorize artifacts primarily by shape and rigidity. The present experiments investigated the origins of this distinction. Using a looking time procedure, Experiment 1 extended previous findings that rhesus macaques (Macaca mulatta) generalize learning about novel food objects by color over changes in shape. Six additional experiments then investigated whether human infants show the same signature patterns of perception and generalization. Nine-month-old infants failed to detect food objects in accord with their intrinsic properties, in contrast to rhesus monkeys tested in previous research with identical displays. Eight-month-old infants did not privilege substance information over other features when categorizing foods, even though they detected and remembered this information. Moreover, infants showed the same property generalization patterns when presented with foods and tools. The category-specific patterns of perception and categorization shown by human adults, children, and adult monkeys therefore were not found in human infants, providing evidence for limits to infants’ domains of knowledge.
1. Introduction
Infant animals, including humans, must come to recognize individual members of their social group and predict those individuals’ behaviors, distinguish potential predators and prey, learn the layout of their territory so as to navigate efficiently between significant locations, select safe and nutritious foods, and identify, categorize, and reason about a plethora of objects. Their learning task is made more challenging by the diversity of the world in which they live. Some entities, like other people and animals, move autonomously and change posture, whereas others, like trees and rocks, are relatively rigid and stable. Some entities, like tools, have functional properties that depend largely on their shape and rigidity; others, like foods, have functional properties that depend largely on their substances. How do young animals and children come to master this diversity?
Many psychologists have proposed that humans and other animals are endowed with special-purpose systems for learning about entities of particular kinds such as inanimate manipulable objects, goal-directed agents, animals, foods, social partners, and competitors (e.g., Barkow, Cosmides, & Tooby, 1992; Hirschfeld & Gelman, 1994). On this view, each of a multitude of “core knowledge” systems emerges early in development, serves to identify the entities in its domain by analyzing their distinctive characteristics, and supports the acquisition of further knowledge about those entities by focusing on the critical features that distinguish different members of the domain. Evidence for such systems comes from convergent studies of the distinctive signature limits on perceptual, cognitive, and neural processing of entities from different domains, as shown by infant humans and animals, behaviorally competent animals reared under controlled conditions, and adult humans living in diverse cultures and environments (for reviews, see Spelke, 2004; Spelke & Kinzler, 2007). When diverse experiments find signature limits that are specific to processing within a given domain, but that are general across ages, species, rearing conditions, and cultures, those findings provide evidence for a distinct, domain-specific system of core knowledge.
In the present work, we ask whether one domain of evolutionary and ecological significance - the domain of food - qualifies as a domain of core knowledge (see also Rozin, 1990; Rozin & Kalat, 1971; Rozin & Schulkin, 1990). In particular, we focus on previous findings that human adults, children, and animals attend to substance information - conveyed by color, texture, and odor - when identifying and classifying foods, but attend to other properties such as shape when identifying and classifying artifacts. This research is consistent with the thesis that food forms a distinctive core domain. Nevertheless, the evidence is inconclusive because these distinctive signatures of processing in the food domain have not been studied in infants and have received little study in animals reared under appropriately controlled conditions. Thus, we report evidence from adult nonhuman primates reared under naturalistic but limited conditions, and from human infants. Across seven experiments, we test whether the distinctive patterns of detecting and categorizing food and nonfood objects shown by human adults and children are present in these populations.
1.1. Problems and solutions for generalist animals in the domain of food
Food identification and selection are challenging tasks for humans and other generalist animals. Confronted with a great diversity of potential foods, they must avoid inedible substances and choose beneficial and varied sources of nutrition (Rozin, 1976, 1977, 1990; Rozin & Pelchat, 1988; Rozin & Schulkin, 1990). Accordingly, a number of mechanisms serve to guide generalist animals’ learning and behavior in the food domain (for reviews, see Barker, Best, & Domjan, 1977; Rozin & Kalat, 1971; Rozin & Schulkin, 1990; Shepherd & Raats, 2006). For example, humans and other animals eat foods that satisfy innate taste biases (e.g., for salt; Rozin & Schulkin, 1990), tend to avoid unfamiliar foods (e.g., Domjan, 1977; Pliner & Salvy, 2006), readily learn and store associations between nausea and ingested substances (e.g., Garb & Stunkard, 1974; Garcia, Ervin, & Koelling, 1966; Garcia & Koelling, 1966; Gustavson, 1977; Richter, 1953; Rozin & Kalat, 1971), and model their food choices after those of conspecifics (Galef, 1996; Galef & Beck, 1990; Rozin, 1988, 2007).
Many of these food selection strategies require organisms to attend to relevant intrinsic properties of foods - including taste, color, texture, and odor. For instance, in order to avoid a kind of food that has made it sick in the past, an animal must be able to generalize learning about a particular food to new foods with similar properties. Below we summarize previous research investigating attention to food-relevant properties in human adults, children, and nonhuman animals. We focus in particular on studies that distinguish organisms’ perception and reasoning about foods vs. artifacts, as this is the literature that is the most relevant to the present research questions and findings.
1.2. Properties that guide classification of food and nonfood objects in human adults and children
Adults attend to color, texture, odor, and taste information when discriminating between edible and inedible entities and when categorizing different kinds of foods (Lavin & Hall, 2002; Ross & Murphy, 1999; Rozin & Fallon, 1987). They reject foods based on bad tastes, smells, and unappealing textures (Rozin & Fallon, 1987), and avoid foods that smell and taste similar to things that made them sick as children or as adults (Bernstein, 1999; Logue, Ophir, & Strauss, 1981; Pelchat & Rozin, 1982). When presented with laboratory tasks in which they must reason about novel entities, adults generalize learning about unfamiliar foods according to color, texture, and odor information, but generalize learning about unfamiliar artifacts according to shape (Lavin & Hall, 2002).
As young as three years of age, children show the same patterns of learning and generalization for foods vs. artifacts as adults (e.g., Lavin & Hall, 2002; Macario, 1991; Santos, Hauser, & Spelke, 2002). In one experiment, Macario (1991) presented three- and four-year-old children with novel objects described as things to eat. Children were introduced to a target object (e.g., pink wax in the shape of a wishbone) and then were shown two other objects: a “color-match” (e.g., pink wax in the shape of a kidney) and a “shape-match” (e.g., green wax in the shape of a wishbone). When asked which one tasted like the target, children were more likely to choose the color match than the shape match. In contrast, children tested with the same novel objects described as toys showed the opposite pattern of reasoning: they were more likely to choose the shape-match than the color-match (see also Brown, 1990).
Preschool-age children also distinguish between properties relevant for classifying foods and artifacts when generalizing novel words for unfamiliar entities. Lavin and Hall (2002) taught three-year-old children novel words applied to novel objects and substances presented either as foods or as toys. Like adults, children were more likely to extend novel words by shape than by color, texture, and smell when the stimuli were described as toys than when they were described as foods. In addition to an effect of domain (food vs. artifact), there was an effect of solidity (object vs. substance): participants were more likely to extend words by shape when the stimuli were solid objects than when they were substances (see also Soja, Carey, & Spelke, 1991).
Adults and children therefore show different patterns of learning and generalization for foods and artifacts. These findings in isolation do not reveal, however, whether learning about foods depends on a distinctive core system of knowledge. Children and adults’ performance could be supported by a dedicated system for analyzing foods that is present in infancy and that serves to guide attention to substance information throughout the lifespan. Alternatively, children may learn about the relevant properties of foods through experiences with different kinds of entities. Studies of animals with limited exposure to different foods, and studies of minimally experienced human infants, can begin to shed light on these possibilities.
1.3. Properties that guide detection and classification of foods and nonfoods in nonhuman animals with controlled or limited exposure to foods
Animals possess a broad ability to learn associations between stimulus properties and outcomes in several domains. Classic studies by Garcia and others on poison avoidance learning provided evidence for specificity, however, in animals’ learning about foods: when presented with both a bright light and a food paired with nausea, laboratory-raised rats subsequently avoided the food, but not the light; when the light and food were paired with an electrical shock, rats avoided the light instead of the food (Garcia & Koelling, 1966). Moreover, when rats became ill after ingesting both a familiar food that had not sickened them in the past and a novel food, they subsequently avoided only the latter substance (Revusky & Bedarf, 1967). These findings and others (see Barker, Best, & Domjan, 1977 and Stricker, 1990, for thorough reviews) suggest that animals are capable of attending to relevant properties when learning about novel foods. Does animals’ learning about foods depend on a system that privileges information about the substance properties?
In recent years, research on a semi-free-ranging population of rhesus monkeys living on the island of Cayo Santiago has begun to address this question. This population of monkeys lives freely in social groups on a small island colony established in the 1930s (Rawlins & Kessler, 1987). Half of their diet consists of soil, flowers, leaves, and small berries available on the island; the remainder is comprised of monkey chow provided at feeding stations. These monkeys therefore have less of an opportunity to learn about different types of foods and the properties by which they are detected and classified than animals who live and forage in the wild.
Two previous lines of work provide evidence that adult monkeys who have lived their entire lives in this colony detect and categorize food objects appropriately. One line of research (Munakata, Santos, Spelke, Hauser & O’Reilly 2001) tested whether this population of monkeys can parse natural yet novel food objects (e.g., lemons, fresh ginger root) in visual displays in which food objects were presented either alone or in contact with one another. Monkeys were tested in a looking time procedure modeled after studies of object perception with human infants (Spelke, Breinlinger, Jacobson, & Phillips, 1993). In one experiment, monkeys first viewed two stationary, adjacent food objects, one on top of the other. Then, a human hand grasped the top of the upper object and lifted either that object, or both objects together, into the air. The outcome displays remained at rest for 10 s, while monkeys’ looking times were recorded. Looking times in the experimental conditions were compared to looking times of monkeys in a baseline condition, who viewed only the two outcome displays with each object held by a hand. Monkeys in the experimental condition looked longer at the event in which the two food objects moved together, relative to baseline, providing evidence that they perceived the boundary between the objects in the display. A second experiment used the same method but presented a single stationary object, followed by outcome events in which the object moved as a whole or only its top half moved into the air. In this experiment, monkeys in the experimental condition looked equally at events in which the object moved as a whole vs. broke apart, relative to baseline. This second finding suggests that monkeys saw each food object as a potentially breakable entity, perhaps because food objects are meant to be taken apart and eaten.
The studies by Munakata et al. (2001) indicate that rhesus monkeys use property information in (initially) static displays to reason about likely boundaries of food objects, and they suggest that monkeys do so in a manner that is particularly appropriate to the parsing and tracking of food objects. These studies do not, however, provide clear evidence for domain specificity in monkeys’ apprehension of food objects. First, multiple properties were available to guide monkeys’ parsing of foods in these experiments, including color, texture, and shape. Additionally, monkeys were never presented with analogous displays containing inedible objects, so it is unclear whether the patterns observed are unique to perception of foods.
A second line of research - focused on categorization - has directly assessed monkeys’ attention to color vs. shape when generalizing learning about unfamiliar foods and artifacts (Santos, Hauser, & Spelke, 2001; Santos, Miller, & Hauser, 2003). In one food categorization study, for example, a human experimenter first pretended to eat a novel object (e.g., a piece of pink Play-Doh in the shape of a sphere), so as to provide information that the object was edible. Following this familiarization event, monkeys were allowed to choose between an object of the same color as the target, but in a new shape (e.g., pink Play-Doh in the shape of a donut) vs. with an object of the same shape as the target, but in a new color (e.g., green Play-Doh in the shape of a sphere). Monkeys selectively approached the food that matched the target’s color, demonstrating that they generalized learning about food objects by color over shape (Santos et al., 2001).
In contrast to their reasoning in the food domain, monkeys from this colony were found to use shape information when learning about novel tools (Santos et al., 2003). Santos et al. (2003) tested adult monkeys using a looking time procedure in which participants were habituated to scenes of a novel tool (e.g., an “L” made from purple clay) pushing a small grape down a ramp. At test, monkeys watched trials in which the pushing action was performed by a tool with the same shape, but new color (e.g., an “L” made from pink clay) vs. trials in which the event was performed by a tool with the same color but new (nonfunctional) shape (e.g., a stubby stick made from purple clay). Monkeys looked longer at the latter type of trial, providing evidence that they see shape as a more critical property than color for classifying objects (see also Hauser, 1997).
Although these findings suggest that monkeys identify food objects by their substance properties, all the above studies have a critical shortcoming: The particular objects used to test for sensitivity to shape and substance properties differed for the food objects vs. the tools. In particular, the food objects presented to monkeys had relatively simple shapes and relatively intricate and interesting textures, whereas the tool objects presented to monkeys had a more interesting and prominent shape. A critical question, therefore, is whether monkeys would show differing patterns of learning and generalization if the same objects, with the same shape and substance properties, were used either as foods or as tools. Experiment 1 addresses this question with the methods and objects from Santos et al. (2003) presented in a food context.
1.4. Properties that guide parsing and classification of foods and nonfoods in human infants
There is a large literature devoted to understanding food acceptance and selection in children after the weaning period - including the development of taste preferences, neophobia, and notions of disgust (e.g., Birch & Marlin, 1982; Birch, McPhee, Steinberg, & Sullivan, 1990; Cooke, Wardle, & Gibson, 2003; Fallon, Rozin, & Pliner, 1984; Pliner & Stallberg-White, 2000; for reviews see Birch, 1990; Birch & Fisher, 1996; Rozin, 1990; and Rozin, 1996) - but little research has directly investigated young infants’ perception and categorization of foods. Nevertheless, a wealth of research has investigated human infants’ detection and categorization of nonfood objects, and the findings of this research contrast with the above findings on monkeys’ individuation and categorization of foods.
Infants often fail to perceive object boundaries in stationary displays by analyzing the colors, textures, and forms of surfaces when they are presented either with simple geometric shapes (Kestenbaum, Termine & Spelke, 1987; Spelke et al., 1993) or with familiar, meaningful objects such as toy animals and vehicles (Xu, Carey & Welch, 1999; although see Needham & Baillargeon, 2000, for exceptions). Instead, studies suggest that infants perceive the unity and boundaries of objects primarily by analyzing the spatial relationships and motions of surfaces, grouping together surfaces that are in contact and that undergo common motion (e.g., Jusczyk, Johnson, Spelke & Kennedy, 1999; Kellman & Spelke, 1983; Spelke, von Hofsten & Kestenbaum, 1989; see Kellman & Arterberry, 1998, and Condry, Smith, & Spelke, 2001, for reviews). Although surface colors and forms do influence object perception under certain conditions (Johnson & Aslin, 1996; Needham, 1997; Smith, Johnson, & Spelke, 2003), the findings suggest the primacy of spatio-temporal properties in infants’ perception of objects (Carey & Xu, 2001; Spelke, 1990). None of these experiments, however, presented food objects with shapes and textures like those tested by Munakata et al. (2001) with monkeys. It is possible, therefore, that human infants would show the same distinctive processing of food objects shown by adult monkeys, if they were tested with the same displays.
There is a similar gap in studies of infants’ object categorization. Numerous experiments provide evidence that infants use shape information, but not substance properties, when generalizing learning about artifact objects (Graham, Kilbreath, & Welder, 2004; Welder & Graham, 2001). For example, infants as young as 13 months of age generalize learning about hidden properties (e.g., rattling, squeaking) of artifacts by shape (across changes in texture), but not by texture (over changes in shape) (Graham et al., 2004). Further studies have investigated infants’ learning about animals, providing evidence that 7-month-old infants generalize learning about animals and their motions by shape (over changes in color), but not by color (Shutts, Markson, & Spelke, 2009). To our knowledge, no studies to date have investigated infants’ use of shape and substance properties in learning about foods. Do infants, like adult monkeys, categorize foods according to substance information, and therefore show distinctive learning patterns in the food domain?
1.5. Overview
The current paper is comprised of three parts, each addressing one of the above three open questions. In Part 1, we use a looking time method to investigate adult monkeys’ learning and generalization about foods and tools, using the same objects and object properties to test learning in the two domains (Experiment 1). In Part 2, we use the method of Munakata et al. (2001) to investigate 9-month-old infants’ perception of the unity and boundaries of food objects, using the same object displays as those used in Munakata et al’s studies of monkeys (Experiments 2 and 3). In Part 3, we use a looking time procedure to investigate whether 8-month-old infants show specific patterns of learning and generalization for foods (Experiments 4-6) and artifacts (Experiment 7).
2. PART 1: rhesus monkeys’ selective generalization of learning about objects
The subjects for this experiment were adult rhesus monkeys (Macaca mulatta) living in the Cayo Santiago field site (Rawlins & Kessler, 1987). Approximately half of the monkeys’ diet consists of Purina monkey chow provided at feeding stations; the remainder consists of leaves, flowers, small berries, and soil found on the island. Although subjects in this population are semi-free-ranging, they have less experience with natural food objects than animals living in the wild, and no experience with the kinds of food objects presented in Experiment 1.
Experiment 1 investigated whether monkeys show different patterns of generalization for food objects than for artifacts. To address this question, the experiment was conducted using the objects, events, and procedures of previous studies of this population that focused on monkeys’ artifact categories (Santos et al., 2003). The procedure was a familiarization paradigm like that of Experiment 1 of Santos et al. (2003), except that instead of using an object as a tool, the experimenter pretended to eat it. As in the Santos et al. (2003) experiment, monkeys were presented with an L-shaped object of a distinctive color, held by an experimenter on three trials. Whereas the monkeys in the past research saw the experimenter use the object as a tool, those in the present study saw the experimenter taste the object. Then, the monkeys viewed test trials with two new objects: one of a different color and the other of a different, truncated shape. In the experiments of Santos et al. (2003), monkeys looked longer when the artifact object appeared with a changed shape than when it appeared with a changed color. If they did so because they generally prefer to look at shape changes, then they should show a similar pattern in this experiment; namely, they should look longer at the shape change test condition than at the color change condition. However, we predicted that subjects would show a different pattern of looking in this experiment, responding more to a change in the food object’s color than a change in its shape. In contrast to their performance with tool objects, rhesus monkeys should look longer at a color change test trial than a shape change test trial when the objects they view are treated as foods.
2.1. Experiment 1
2.1.1 Method
2.1.1.1 Subjects
Twenty-two monkeys were successfully tested in this experiment. Only monkeys found to be relatively isolated from other group members were chosen to participate. Thirteen additional subjects were tested but did not complete testing due to subject inattention, interference from other animals, previous testing, and/or experimental error. Two other subjects’ data were eliminated during coding by the coder (who was blind to condition) because the videotape was judged too blurry to code.
2.1.1.2. Apparatus
Figure 1 presents the events and objects in this experiment. The objects were two different stick shapes (one L-shaped stick, and one stick with a stubby base) made from purple or pink clay. The objects were identical to the ones used as tool stimuli in Santos et al. (2003), and were placed on a stage identical to that used in Santos et al. (2003).
Figure 1.

Displays from Experiment 1. After familiarization with a single object (e.g., left), the monkey viewed test displays with an object of the same shape but novel color (center) and an object of the same color but novel shape (right).
2.1.1.3. Procedure
Monkeys were tested in the field. One experimenter presented the displays to the subject, while another recorded the subject’s looking behavior using a video camera. Each subject viewed three familiarization trials and two test trials. In each of these trials, subjects watched the following series of events: The experimenter picked up the object from the stage and then showed the subject the object (e.g., a purple L-shaped object). As the subject watched, the experimenter placed the object in his mouth, mouthed the object for 3 s, and said “Mmm.” The experimenter then placed the object on the upper platform and called “Start” and the subject’s looking time was recorded for the next 10 s. This familiarization trial was repeated three times. Other than the eating actions, this procedure was the same as in Santos et al. (2003, Exp. 1).
After viewing all three familiarization trials, subjects were given two test trials: a new shape test trial and a new color test trial. Each of these tests was identical to the familiarization trial, except that subjects watched the experimenter eat a novel object. In the new color condition, the experimenter pretended to eat an object of the same shape but new color (e.g., pink L-shaped object). In the new shape condition, the experimenter pretended to eat an object of the same color but new shape (e.g., purple stubby stick). After pretending to eat the object, the experimenter placed the novel object on the upper platform and called “Start” and the subject’s looking time was recorded for the next 10 s. As in previous experiments, each subject received one trial of each test condition.
2.1.1.4. Coding
As in the previous experiments (Santos et al., 2003), videotapes were acquired onto a Macintosh computer and were analyzed with Adobe Premiere software. These digitized sequences were scored by one coder who was blind to the experimental condition. The coder examined looking during each frame (30 frames = 1 s) of the 10 s looking period that followed each trial. A look for the purposes of these experiments refers to a period of 5 frames or longer during which the subject’s head was oriented towards the stage. A second coder then scored a subset of these trials to establish reliability (r = .80).
2.2. Results
All subjects appeared to habituate across the first three trials: They looked reliably less on the third familiarization trial than on the first (t(21) = 3.75, p < .001). We then examined whether or not they recovered looking on the test trials. Monkeys looked longer to both the new shape test trial (t(21) = 2.32, p < .05) and the new color test trial (t(21) = 3.91, p < .001). However, although monkeys looked longer in both test trials than in the previous familiarization trial, they showed a significant difference in looking between the two test trials (t(21) = 3.17, p < .005). In contrast to monkeys tested by Santos et al. (2003), those in the present study looked significantly longer at the color change trial than at the shape change trial (see Figure 2).
Figure 2.
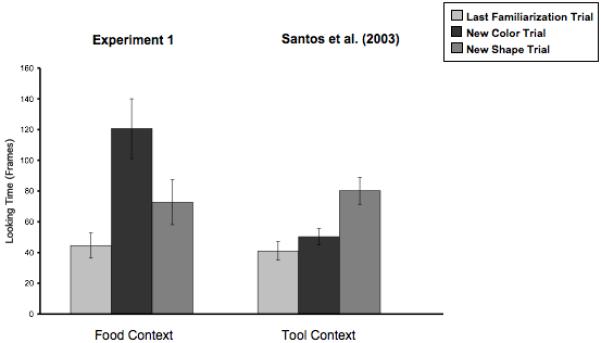
Results from Experiment 1 of the present paper and Experiment 1 of Santos et al. (2003).
A repeated-measures ANOVA with experiment as a between-subject factor, and test condition (shape change test or color change test) as a within-subject factor was conducted to compare results from Santos et al.’s (2003; Experiment 1) tool experiment to the present findings. This analysis revealed only a marginal main effect of experiment (F(1,43)= 3.8, p = .06) and no effect of test condition (F(1,43) = 1.01, n.s.), suggesting that subjects do not differ in their overall duration of looking in the two experiments or at the two test displays. There was, however, a significant interaction between experiment and test condition (F(1,43) = 19.05, p < .001). Subjects demonstrated a different pattern of looking across the two experiments, looking longer at the shape change in Santos et al. (2003) and looking longer at the color change in the current Experiment 1.
3. General Discussion of Part 1
In Experiment 1, monkeys were presented with events in which a human experimenter ate a novel object and then later ate a new object with either a new shape or a new color. We found that subjects noticed both featural changes; subjects recovered looking to a change in the eaten object’s color and to a change in the eaten object’s shape. However, the magnitude of looking differed reliably across the two featural changes. Monkeys looked almost twice as long when the experimenter ate a differently-colored object than when the experimenter ate a differently-shaped object. This finding suggests that although monkeys detected both changes they found a change in a food’s color to be more salient than a change in the food’s shape. This result is consistent with previous findings with this population using an object choice task (see Santos et al., 2001).
This pattern of looking longer at changes in an object’s color stands in contrast to the pattern observed by Santos et al. (2003) where the same object was used as a tool. In those experiments, monkeys attended more to a change in the object’s shape and showed no increase in looking time to a change in its color. The statistical interaction between the experiments suggests that monkeys do not have a general bias to attend to changes in an object’s shape. Instead, the featural changes that monkeys find salient seem to depend on the context in which the object is introduced. When monkeys observe an object acting as a tool, they attend more to its shape; when they observe the very same object being eaten, they attend more to its color. Because the same objects and a highly similar method, were used across these experiments, monkeys’ differential generalization provides evidence for domain-specific learning about foods and nonfoods.
Since the participants in these experiments had no experience with the food objects in Experiment 1 and relatively impoverished experience with diverse foods in their environment, the findings may be explained by the existence of distinct core systems for representing and learning about food vs. nonfood objects. Nevertheless, monkeys on Cayo Santiago had had opportunities to learn about food-relevant properties over the course of their lives. Therefore, the remaining experiments tested for core knowledge of foods in a population with even less relevant experience, namely human infants.
4. PART 2: human infants’ individuation of food objects
The experiments in this section test whether 9-month-old human infants, like adult rhesus monkeys, parse food objects according to intrinsic features available in static displays. Although many experiments have investigated infants’ parsing of objects, all studies to date have used artifact objects - either simple geometrical solids or familiar artifacts such as cups, books, and toy cars (Kestenbaum et al., 1987; Spelke et al., 1993; Xu et al., 1999). These experiments provide evidence that infants often fail to parse objects at color, texture, and shape boundaries in stationary displays. One possibility is that young infants are generally unable to use static properties to parse objects of any kind. Another possibility is that infants would be able to parse objects at boundaries when viewing objects for which substance properties are particularly relevant (i.e., foods), and/or objects with more complex and natural shapes and textures.
A series of studies using a preferential looking method illustrate the methods, findings, and limitations of previous research on infants’ object parsing. In these studies (Spelke et al., 1993), infants aged 3-9 months were presented repeatedly either with a single, homogeneous object or with two objects of contrasting shapes and colors, one on top of the other on a supporting surface. While the object(s) remained at rest, a hand entered the display and grasped its top, and then looking time was measured until the infant looked away from the display. Following habituation, infants were given a series of test trials presenting two alternating events: The object array appeared as before, the hand grasped and lifted the top of the display, and either (1) the top half of the display moved alone while the bottom half remained at rest on the surface (a natural motion in the case of two separate objects, but unnatural for one object) or (2) the entire object display rose together into the air (a natural motion in the case of a single object, but unnatural for two separate objects). Looking times to the outcomes of these events were measured and compared to those of a separate group of infants in a baseline condition, who viewed the same outcome displays with no prior exposure to the original arrangement of objects.
Infants as young as 3 months looked longer at the event in which the single object broke apart, providing evidence that they perceived the homogeneous object as a single, cohesive body. In contrast, infants as old as 9 months showed no differential looking at the events in which the two stationary objects moved separately or together. Because infants look longer at events in which two distinct objects move together when the objects are separated in space or undergo relative motion (Kestenbaum et al., 1987; Spelke et al., 1989; von Hofsten & Spelke, 1985), this finding suggests that infants perceive object boundaries specified by surface arrangements and motions, but sometimes fail to perceive object boundaries specified by surface colors, textures, or forms.
This conclusion contrasts markedly with the findings of Munakata et al. (2001), who used a variation of Spelke et al.’s (1993) method to test rhesus monkeys’ parsing of food objects. Because the displays from past studies of infants differ from those used with monkeys, however, the source of the differing results is not clear. In particular, the above studies, like most studies of object parsing in infancy, presented artifact objects with geometrically regular shapes and uniform textures. In contrast, the objects presented to monkeys were natural food objects with natural shapes and complex textures. It is possible that infants and monkeys alike will parse natural objects differently from uniform geometrical solids.
The present experiments therefore investigate human infants’ perception of the unity and boundaries of natural food objects using the displays of Munakata et al. (2001). Experiment 2 investigated infants’ perception of two adjacent food objects of contrasting color, texture, and shape. If human infants perceive natural food objects in the same manner as adult monkeys, then they should parse these objects as separate units and look longer when they move rigidly together. In contrast, if infants perceive natural food objects in the same manner as they perceive manufactured objects, then they may fail to perceive these objects as separate units. In Experiment 3, we investigated infants’ perception of a single food object of a single, natural color and texture. If human infants perceive single food objects as do adult monkeys, then they should fail to perceive a single food object as an unbreakable whole and should look equally at events in which the food object moves as a rigid unit vs. breaks apart. In contrast, if infants perceive food objects in the same manner as manufactured objects, then they should perceive the single object as a unit and look longer when its unity is broken by separate motion of the top half of the object, as in past research (Spelke et al., 1993).
In order to maximize the comparability of the infant and monkey experiments, the infants in Experiments 2 and 3 were tested with the same types of food objects in the same arrangements as were the monkeys in the studies of Munakata et al. (2001). A more difficult decision concerned the testing procedure, which differed in one respect in the past experiments with monkeys vs. infants: monkeys were given far briefer exposure to the initial array of objects than infants because, as unrestrained adults, they were expected to form a representation of the object array more rapidly than infants and to walk away from the display, ending the experiment, if shown the same events over many repetitions. Because pre-locomotor infants might need longer exposures in order to form object representations, we decided to maximize infants’ chances of perceiving all the objects correctly by using the full habituation method of Spelke et al. (1993).
4.1. Experiment 2
Nine-month-old infants were tested in two conditions: experimental and baseline. Infants in the experimental condition were habituated to two stationary food objects arranged one on top of the other on a puppet stage. Then the infants were tested with events in which a hand grasped the top object and either just that object, or both objects, rose rigidly into the air and then remained stationary. Looking time was recorded, beginning at the end of the motion and ending when the infant looked away from the display. Infants in the baseline condition were presented with the same event outcomes with no prior motion: a stationary display in which the hand held either the top object or both objects in the air. If infants perceived the two food objects as two separately movable bodies, then the infants in the experimental condition were expected to look longer at the outcome of the event in which the objects moved rigidly together, and this preference should have exceeded any baseline preference for that outcome display.
As in past research with monkeys, infants were tested with a miniature pumpkin on top of a piece of ginger root (see Fig. 3, left). These displays were motionless during the habituation events. For the test events of the experimental condition, a single hand grasped the pumpkin and lifted it, and either the grasped object moved by itself (relative motion) or the two objects moved together (common motion). The former event appears natural to adults and the latter event appears unnatural. In the test events of the baseline condition, either a hand held the top object, or a hand held both of the objects by grasping the pair at the boundary and supporting both objects. Both these displays appear natural to adults.
Figure 3.
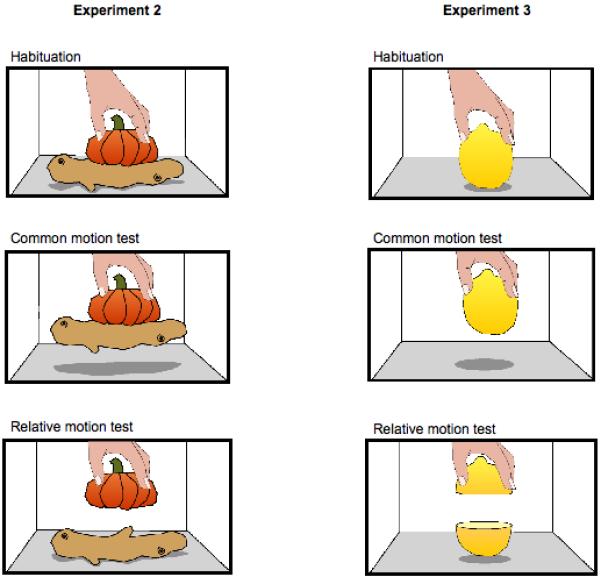
Displays from the experimental conditions of Experiments 2 and 3.
4.1.1. Participants
Participants were 20 9-month-old infants (15 males; M=9 months, 1 day; range = 8 months, 15 days to 10 months, 0 days). One additional infant was tested but not included in data analyses because of experimenter error.
4.1.1.2. Displays
Displays were presented on a stage composed of white foam-core measuring 75-cm (wide) × 30-cm (tall) × 38-cm (deep). Above the stage was a 75-cm × 30-cm screen that could be lowered to occlude the display stage between trials. The objects were a miniature orange pumpkin (6-cm tall × 8-cm diameter) and a segment of tan ginger root (4-cm tall × 4-cm diameter × 12-cm long). In the display, the pumpkin rested on top of the ginger root. During test trials, the experimenter’s hand reached down into the display and lifted the pumpkin 10 cm above the floor of the stage, then paused there for the duration of the trial. The infant was seated in a high chair positioned 80 cm from the front of the stage. The display objects subtended visual angles of 5.8° × 5° (pumpkin) and 8.6° × 2.2° (ginger root).
4.1.1.3. Design
Ten infants participated in the experimental condition, and 10 participated in the baseline condition. Half the participants in each condition were tested with each order of test trials (common motion first vs. relative motion first). The 10 infants in the baseline condition of this experiment also participated in the baseline condition of Experiment 3, separated by a break of about 3 minutes; half the infants received the present baseline condition first.
4.1.1.4. Procedure
Infants were seated in a high chair facing the display stage and were accompanied by a parent who was seated behind the infant. The study began with calibration in which the experimenter used a squeaking toy to direct the baby’s attention to all parts of the display area, signaling to the coders the target area of the infant’s eye movements. Coders were seated in a separate room viewing a video feed showing only the infant’s face, and were thus unaware of the infant’s condition. Coders depressed buttons connected to a Gateway 2000 computer to indicate when the infant was attending to the display. The computer recorded the infant’s looking time on each trial and calculated inter-observer agreement. The start of each trial was controlled by the experimenter, who initiated computer recording. Trials ended when the infant looked away from the display for 2 s consecutively or once 60 s had elapsed. Once calibration was complete, the occluder screen was lowered to occlude the stage and the experiment began.
On each habituation trial of the experimental condition, the screen was raised to reveal the pair of objects situated one on top of the other. The experimenter’s hand reached down into the display from behind a curtain, tapped on the top object with one finger, and then came to rest on the top object. Recording of the infant’s looking time began when the hand came to rest on the top object, and was controlled by the experimenter who pressed a key connected to the recording computer. Habituation trials continued in this manner until criterion of a 50% decline in looking was reached. The criterion for habituation was a defined as three consecutive trials whose average time was less than half of the average of the first three habituation trials. Once the infant reached criterion, test trials began. On each test trial, the screen was raised to reveal the same two objects as in the habituation trials. The experimenter’s hand reached down into the display and grasped the top object, then lifted it 10 cm above the stage floor and paused there for the duration of the trial. On the Relative Motion trial, the top object was lifted alone. On the Common Motion trial, both objects rose when the top one was lifted because they had been surreptitiously connected. Recording of the infant’s looking time began when the hand and object(s) came to rest, and was controlled by the experimenter who pressed a key at the end of the motion.
Infants in the baseline condition were presented with the two test trial events without habituation. These test trials presented the same pair of objects (pumpkin/ginger) as in the experimental condition, except the objects were stationary throughout the trial. In each trial of the baseline, the screen was raised to reveal the two objects and the experimenter’s hand holding one or both of the objects 10 cm above the stage. This display mimicked the final outcome in the experimental test trials with the experimenter’s hand holding the objects in mid-air, but did not show the objects being lifted. In the Relative Motion outcome, the experimenter held only the top object. In the Common Motion outcome, the experimenter held both objects by grasping them at their juncture, thus supporting both objects with one hand. Half of the infants in each condition saw each outcome first.
4.1.5. Dependent measures and analyses
Looking time was recorded during the outcome displays by two condition-blind independent observers. Inter-observer agreement, assessed as the correlation between button presses by the two observers, was calculated by the computer 10 times/s during each trial and averaged over the experiment. Inter-observer agreement averaged 91% in Experiment 2.
Paired-sample t tests were used to compare infants’ looking on the last habituation trial to looking on each of the two test trials in the experimental condition, as well as to compare looking on separated vs. connected test outcomes in each condition. Test trial data were also subjected to a 2 (condition: experimental vs. baseline) by 2 (test motion: relative vs. common) ANOVA.
4.2. Results
Figure 4 (left) presents looking time on the last habituation trial for the infants in the experimental condition, as well as looking time to each of the test outcomes for infants in both conditions. Infants in the experimental condition showed marginally increased looking from the last habituation trial to each of the two test trials (relative motion: t(9) = 2.26, p = .05; common motion: t(9) = 2.08, p = .067). There was no difference between infants’ looking at the relative vs. common motion test outcomes in the experimental condition (t<1) or in the baseline condition (t(9) = 1.65, n.s.). The ANOVA revealed no effect of condition (F<1), no effect of test outcome (F<1), and no interaction between the two variables (F(1,18) = 1.13, n.s.).
Figure 4.
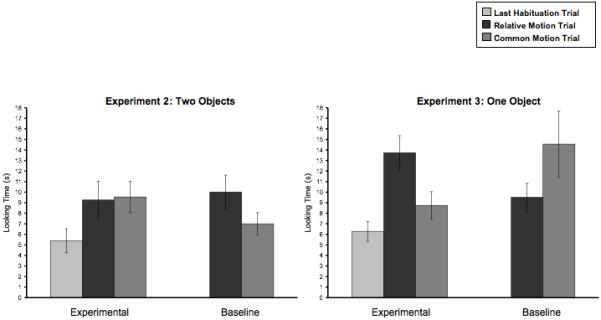
Results from Experiments 2 and 3.
4.3. Discussion
When human infants were presented with two stationary, adjacent food objects, they looked equally at an event in which the top object was lifted and moved on its own and one in which the top object was lifted and both objects rose together. Thus, Experiment 2 provides no evidence that infants perceived the two food objects as distinct, separately movable bodies.
Because the principal findings of this experiment were negative, we conducted a replication with a new sample of 20 infants (10 in the experimental condition, and 10 in the baseline condition). For the new experiment, we used a second pair of food objects that Munakata et al. (2001) had presented to monkeys, and that monkeys had parsed as two objects: a green pepper on top of a potato. The findings of this replication experiment were the same as in Experiment 2. Moreover, when the data from all 40 infants were considered together, the findings were still negative: Infants looked equally long at the common motion and relative motion trial in both the experimental (common motion M = 9.98 s; relative motion M = 10.16 s; t<1) and the baseline condition (common motion M = 10.42; relative motion M = 10.78; t<1), and there was still no interaction of test outcome by condition (F<1). However, considering all 40 infants, dishabituation to the relative motion and common motion trials in the experimental condition moved from marginal to significant (t(19) = 2.87, p < .01; t(19) = 3.12, p < .01, respectively), providing evidence that infants dishabituated to both events.
The present findings contrast with those of Munakata et al.’s (2001) study of adult rhesus monkeys. When monkeys were tested with the same displays and a similar (though briefer) method, they looked reliably longer at the outcome of the unnatural event in which the two objects moved together. This contrast suggests that the ability to perceive the boundaries of food objects varies either by species (rhesus monkeys vs. humans) or by age (adults vs. 9-month-old infants). We consider these two possibilities in the discussion following Experiment 3.
In Experiment 3, we investigated a second aspect of object perception in infancy: perception of a single object as a unitary, commonly moving body. Recall that adult monkeys presented with a single food object appeared not to perceive the object as a commonly movable unit, because they looked equally long at events in which the object moved as a whole or broke in two. In contrast, human infants presented with a single geometrical solid have been shown through this and other methods to perceive such an object as a unit (e.g., Spelke et al., 1993). If a single system of representation underlies infants’ perception of artifact objects and natural food objects, then infants who are presented with a single food object also should perceive its unity and should look longer at the outcome of an event in which the object breaks apart than at one in which it moves as a whole.
4.4. Experiment 3
Nine-month-old infants in the experimental condition were habituated to a single stationary food object on the same stage as in Experiment 2. Then the infants were tested with events in which a hand grasped the top of the object and either the whole object, or just its top half, rose into the air. Infants in the baseline condition were presented with the same event outcomes with no prior motion, and looking times to the outcome displays were compared across conditions as in Experiment 2.
As in past research with monkeys, infants were tested with a lemon (see Fig. 3, right). These displays were motionless during the habituation events. For the test events of the experimental condition, a single hand grasped the top of the lemon and lifted it. This event appears natural to adults when the object moves as a whole and unnatural when the top half of the object moves separately from the bottom. In the test events of the baseline condition, a single hand held either the whole object or the top half. Because the object was never previously presented as a whole, both these displays appear natural to adults.
4.5. Method
Participants were 20 infants (M = 9 months, 0 days; range = 8 months, 13 days to 9 months, 17 days), half males and half females, drawn from the same population as in Experiment 2. No additional infants were removed from the sample. The object was an 8-cm high × 6-cm diameter ripe lemon. For relative motion outcome trials the lemon was cut in half horizontally. The cut in the display object was made with a very sharp knife such that the cut line was nearly invisible when the two halves were placed together. When the halved object was presented in the outcome trials, the infant was able to see a part of the inside of the half lemon that was resting on the stage. The visual angles subtended by the display objects measured 4.3° × 5.7° (whole lemon) and 4.3° × 2.9° (half lemon). All other aspects of the displays, events, design, procedure, and analyses were the same as in Experiment 2. Inter-observer agreement averaged 90% in Experiment 3.
4.6. Results
Fig. 4 (right) presents the data from Experiment 3. Infants in the experimental condition maintained low levels of looking (relative to habituation) at the test outcome in which the object moved as a whole (t(9) = 1.49, n.s.), but showed increased looking at the test outcome in which the object broke apart (t(9) = 4.13, p < .01). Infants in the experimental condition looked significantly longer at the test outcome in which the object broke apart, compared to the outcome in which it moved as a whole (t(9) = 2.34, p < .05), while infants in the baseline condition tended to look longer at the test outcome in which the object moved as a whole (t(9) = 2.21, p =.054). A 2 (condition) by 2 (test outcome) ANOVA revealed no effect of condition (F<1) or test outcome (F<1). Most important, there was a significant interaction of condition by test outcome (F(1,18) = 10.36, p < .005): Infants in the experimental condition showed a greater preference for the outcome of the event in which the object broke apart than did those in the baseline condition.
4.7. Discussion
Presented with a whole lemon that was grasped and lifted, infants looked longer when the object broke apart and only its top half rose into the air than when the object moved as a whole. This tendency reliably exceeded baseline looking toward the former outcome display. Taken together, the results provide evidence that infants perceived the single lemon as a separately movable whole. These findings provide a further contrast with the findings of studies of adult monkeys (Munakata et al., 2001) and a further convergence with the findings of studies of human infants presented with simple artifacts (Spelke et al., 1993). Just as in past research with simple artifacts, infants perceived a natural food object as unitary, commonly movable whole.
4.8. General Discussion of Part 2
The results from Part 2 provide no evidence that human infants parse food objects in accord with their substance or shape properties. In this respect, the findings contrast markedly with results from studies of adult rhesus monkeys with minimal food experience. Infants’ performance with food objects in Experiments 2 and 3 accords, however, with the performance of infants in many previous experiments using artifacts (e.g., Spelke et al., 1993). The results of Experiment 3 indicate that infants attend to food objects and are able to make some predictions about them, but the predictions they make are the same as those made for artifacts. Therefore, Experiments 2 and 3 provide no evidence for food as a domain in human infants’ knowledge.
Since the objects in Experiments 2 and 3 were not presented in an eating context, however, it is possible that infants were not aware they were foods. To address this concern, the experiments of Part 3 provide infants with information about the category membership of different substances and objects (e.g., by demonstrating eating for foods). In Part 3 we ask whether human infants also differ from adult monkeys in their patterns of generalizing learning about foods and artifacts. Given that the rhesus monkeys in Experiment 1 generalized learning about food by its color over its shape, we ask whether human infants show the same patterns of domain-specific learning about foods.
5. PART 3: Human infants’ categorization of foods and artifacts
The present experiments used a habituation of looking time method to test for specific patterns of learning and generalization about foods and nonfoods at 8 months of age. We tested 8-month-old infants because this is the age at which most American infants have started to consume solid foods as a part of their regular diet. Food is therefore a relevant domain for infants, but a domain about which they might just be beginning to learn. Before investigating infants’ generalization of learning about foods, however, we asked whether infants perceive and remember both substance and shape properties of food.
5.1. Experiment 4
Experiment 4 investigated whether infants perceive and remember properties of food substances and containers. Separate groups of infants were tested in two different conditions. In the “Color/Texture Discrimination Condition”, infants were habituated to an actor eating one type of food substance from one type of container (e.g., green sugar in a champagne glass). At test, they were shown a trial in which the actor ate the familiar substance from the familiar container (e.g., green sugar in a champagne glass) vs. a trial in which the actor ate a novel food from the familiar container (e.g., orange juice in a champagne glass). If infants discriminate between foods on the basis of substance color and texture differences, they were expected to look longer at the test trial in which the actor ate the novel food. In the “Container Discrimination Condition”, infants were habituated to an actor eating one type of food from a particular container (e.g., green sugar in a champagne glass). At test, they viewed one trial in which the actor ate from a familiar container that held the familiar substance (e.g., green sugar in a champagne glass) vs. a trial in which the actor ate from a novel container that held the familiar substance (e.g., green sugar in a bowl). If infants discriminate between food containers on the basis of shape, they were expected to look longer at the test trial in which the actor ate from the novel container.
5.2. Method
5.2.1. Participants
The participants were 32 full-term 8-month-old infants (16 females; M=8 months, 6 days; range = 7 months, 22 days - 8 months, 16 days) drawn from the same population as in Experiments 2 and 3. Eleven additional infants were tested, but not included in the final sample due to experimental error (n=1), failure to meet the habituation criterion (n=6), equipment failure (n=2), or fussiness (n=1).
5.2.2. Materials
The substances were pulpy orange juice, pulpy orange juice dyed green with food coloring, orange sugar crystals (same color as the orange juice), and green sugar crystals (same color as the green juice). The containers were clear glass bowls measuring 6 cm in height and 10.5 cm in diameter and clear champagne glasses measuring 21 cm in height and 5.5 cm in diameter.
5.2.3. Apparatus
Fig. 5 displays some of the events used in this experiment. The events were presented in a well-lit stage with a viewing area that measured 76 cm (width) × 72 cm (height). The actor, a female wearing a white t-shirt and navy visor, kneeled at a white countertop and was visible to infants from the chest up. White foam core surrounded the viewing area and navy curtains were hung behind the actor to conceal equipment. A white shelf was placed on the countertop in front of the actor in order to elevate the containers so that they were approximately at infants’ eye-level. A navy curtain with the same dimensions as the viewing area could be raised and lowered to reveal and hide the actor and other contents of the stage area between trials. Soft music was played from a small stereo unit hidden behind the stage in order to mask any minor sounds made by the actor and experimenter.
Figure 5.
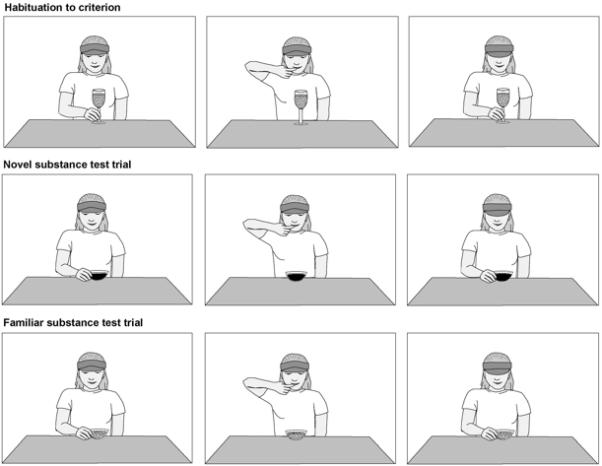
Example displays from the Generalization-by-Color/Texture Condition of Experiment 5. The same events were used in the other conditions of Experiments 4 and 5, arranged so that only one property varied during the test phase (Experiment 4) or so that the test trials presented a novel vs. familiar container with a novel substance (Generalization-by-Container Condition, Experiment 5).
A lipstick camera mounted below the countertop was used to capture infants’ looking, while another camera focused on the stage. Data were recorded and coded as in Part 2.
5.2.4. Design
Half of infants (n=16) were tested in the Color/Texture Discrimination Condition and half were tested in the Container Discrimination Condition. The type of substance and container used in the habituation phase were counterbalanced across infants in both conditions. During habituation, half of infants viewed juice and half viewed sugar, half saw the substance in orange and half saw it in green, and half were shown the glass bowl and half where shown the champagne glass. During the test trial phase, half of infants in the Color/Texture Discrimination Condition saw the familiar substance trial first, while half saw the novel substance trial first. For infants tested in the Container Discrimination Condition, half saw the familiar container test trial first, while half saw the novel container trial first.
5.2.5. Procedure
Infants sat on a parent’s lap in a chair approximately 1.5 m from the testing stage. Parents were asked to remain quiet for the duration of the experiment and were informed (and later reminded) that they should close their eyes during the test trials so that they were blind to the contents of the stage.
5.2.6. Familiarization Trial
The habituation phase was preceded by a brief (approximately 8 s) familiarization trial in which the actor picked up the container from the white shelf, said “what’s this?” to draw infants’ attention, and then stirred the contents of the container with her finger to emphasize that it was a substance. She then took a taste of the substance using her finger, said “yummy” to indicate that it was a food, placed the container back on the white shelf, and then looked down. The actor began the event as soon as the curtain was raised; the curtain was lowered as soon as she finished. Infants’ looking during the familiarization trial was not recorded and did not count toward the criterion for habituation.
5.2.7. Habituation Phase
On every habituation trial, the curtain was raised to reveal the actor looking down at a container resting on the shelf in front of her. The actor dipped her finger into the food, took a taste of it, rested her hand on the container, and then looked down so that infants could not see her face. Once the actor’s hand had come to rest on the container, an experimenter sitting behind the actor (but not visible to infants) pressed a key on the computer to initiate recording of infants’ looking behavior. When infants looked away from the stage for 2 s consecutively or once they had accumulated 120 s of looking, the computer beeped to signal the end of the trial and the experimenter lowered the screen. Habituation trials were administered until infants reached a criterion of 50% decline in looking time on three consecutive trials, relative to the first three consecutive trials that had totaled 12 s or more. The computer signaled when this criterion had been met and then test trials were administered.
5.2.8. Test Phase
Infants viewed two test trials that were identical in procedure to habituation trials. The test pair for the Color/Texture Discrimination Condition consisted of one trial where the actor ate a familiar food from a familiar container (i.e., same as habituation) and one trial where the actor ate a novel food from the familiar container. The test pair for the Container Discrimination Condition was comprised of one trial where the actor ate from a familiar container that contained a familiar substance (i.e., same as habituation) and one trial where the actor ate from a novel container that contained a familiar substance.
5.2.9. Dependent measures and analyses
Infants’ looking behavior was coded online by two independent observers who were blind to condition and trial type. The average inter-observer agreement (calculated as in Experiments 2 and 3) in was 94%.
Test trial looking times were analyzed by a repeated-measures ANOVA with trial type (novel vs. familiar) as a within-subject factor and condition (Color/Texture Discrimination vs. Container Discrimination) as a between-subject factor. Looking times within each condition were analyzed using paired-samples t tests.
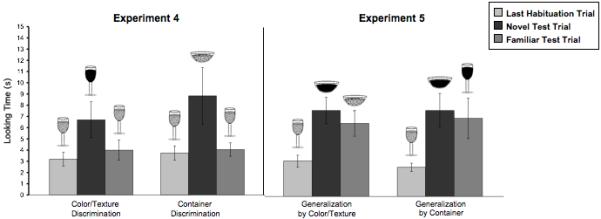
5.3. Results
At test, infants looked longer at the novel display in both conditions (see Fig. 6, left). An ANOVA revealed a main effect of trial type (F(1,30) = 7.88, p < .01), no effect of condition (F<1), and no interaction of condition by trial type (F<1). Infants in the Color/Texture Discrimination Condition looked longer at the test trial where the actor ate the new food than at the test trial where she ate the familiar food (t(15) = 2.52, p < .05). Infants’ average looking during the last habituation trial in the Color/Texture Discrimination Condition differed significantly from the novel test trial (t(15) = 2.26, p < .05), but not from the familiar test trial (t<1). Infants in the Container Discrimination Condition looked marginally longer at the test trial where the actor ate from the novel container than at the test trial where she ate from the familiar container (t(15) = 1.96, p = .069). Infants recovered looking from the last trial of habituation to the novel test trial (t(15) = 2.13, p <. 05) but not to the familiar test trial (t<1).
Figure 6.
Results from Experiments 4 and 5.
5.4. Discussion
The results from Experiment 4 provide evidence that 8-month-old infants perceive and remember properties of food substances when observing eating events. Additionally, the results indicate that our method effectively reveals this ability. The design of Experiment 4 does not reveal whether infants track the substance properties of foods over changes in containers, however, since each condition presented only one kind of property change (i.e., either a change in the color/texture of the substance or a change in container shape), holding all other properties constant. Experiment 5 therefore was undertaken to test whether infants, like adults, young children, and nonhuman primates, show category-specific learning and generalization in the food domain. In one condition, we investigated whether infants focus on information about the color and texture of foods by testing for generalization by those properties across a change in container. In another condition, we asked whether infants focus on information about containers when learning about foods, by testing for generalization by container shape across a change in food substance.
5.5. Experiment 5
Experiment 5 investigated whether infants recognize a familiar food when it is held in a novel container, and recognize a familiar container when it holds a novel food. In the “Generalization-by-Color/Texture Condition”, infants were habituated to scenes in which an actor ate one type of food substance from one type of container (e.g., green sugar in a champagne glass). Then, in test trials, infants were shown a scene in which the actor ate that same food from the new container (e.g., green sugar in a bowl) vs. a scene in which the actor ate a new food from a new container (e.g., orange juice in a bowl). If infants generalize learning about a food substance on the basis of color and texture, over a change in container shape, they were expected to look longer at the trial where the actor ate the new kind of food. In the “Generalization-by-Container Condition”, infants were habituated to an actor eating one type of food substance from one type of container (e.g., green sugar in a champagne glass). In test trials, they were shown a scene in which the actor ate from a familiar container that held a new substance (e.g., orange juice in a champagne glass) vs. a scene in which the actor ate from a new container that held the new substance (e.g., orange juice in a bowl). If infants generalize learning about food on the basis of container shape, over a change in food color and texture, they were expected to look longer at the trial where the actor ate from the novel container.
5.6. Method
The method was the same as Experiment 4, except as follows. The final sample consisted of 32 infants with a mean age of 8 months, 6 days (range = 7 months, 19 days - 8 months, 19 days). Eleven additional infants were tested, but not included in analyses because of experimental error (n=5), failure to meet the habituation criterion (n=5), or extremely lengthy (>3 SDs) looking during the test trial phase (n=1).
Sixteen infants were tested in the Generalization-by-Color/Texture Condition and 16 were tested in the Generalization-by-Container Condition. The test trials for the Generalization-by-Color/Texture Condition consisted of one trial where the actor ate a familiar food substance (i.e., same food as shown during habituation) from a novel container and one trial where the actor ate a novel food substance from a novel container (see Fig. 5). The test trials for the Generalization-by-Container Condition consisted of one trial where the actor ate from a familiar container (i.e., same container as shown during habituation) that held a novel substance and one trial where the actor ate from a novel container that held a novel substance. Average inter-observer agreement was 95%.
5.7. Results
Infants showed no tendency to generalize habituation across changes either in food substances or containers (Fig. 6, right). A repeated-measures ANOVA revealed no effect of test trial type (F<1), no effect of condition (F<1), and no interaction between the factors (F<1). After habituation, infants in the Generalization-by-Color/Texture Condition significantly increased their looking both at the test event with the novel substance and container (t(15) = 4.20, p < .001) and at the test event with the familiar substance and novel container (t(15) = 4.51, p < .001). A comparison of looking at the two test events revealed no preference for the novel substance (paired-sample t<1). Similarly, infants in the Generalization-by-Container Condition increased their looking from the last habituation trial to each kind of test trial (i.e., with the novel substances and the novel or familiar containers) (t(15)= 3.19, p < .01, t(15) = 2.53, p < .05, respectively). A comparison of looking at these two test events also revealed no preference for the novel container (paired-sample t<1).
5.8. Discussion
The results from Experiment 5 provide no evidence that infants track nonsolid foods by their substance properties when they appear in different containers. If infants, like adults, young children, and nonhuman primates, privileged color and texture when reasoning about foods, they should have looked longer in the Generalization-by-Color/Texture Condition at the test trial in which the actor ate the food with the novel color and texture. Instead, infants looked equally long at novel and familiar substance test trials. Results from the Generalization-by-Container Condition indicate that infants do not generalize learning by container shape either, as infants looked equally long at novel and familiar container test trials. Infants’ recovery of interest from habituation to the test phase in both conditions suggests that they noticed that the foods or containers had changed, but they showed no differential interest when both changes occurred at once.
We hypothesized that infants’ failure to generalize information appropriately in Experiment 5 might have stemmed from a general difficulty with representing nonsolid substances. Indeed, recent evidence suggests that even adult non-human primates fail to generalize across color when tested with non-solid foods (see Addessi & Visalberghi, 2001). We chose to use nonsolid substances in Experiment 5 because we thought infants would be most familiar with foods of that nature. However, some studies have shown that while infants are able to represent solid objects, quantifying and tracking them over space and time, infants of 8 months are unable to perform the same operations with nonsolid entities (e.g., Huntley-Fenner, Carey, & Solimando, 2002). Accordingly, Experiment 6 investigated infants’ learning and generalization about solid foods.
5.9. Experiment 6
Infants in Experiment 6 were shown novel, solid food objects, rather than substances, and were tested for generalization of property information as in Experiment 5. Participants in the “Generalization-by-Color/Texture Condition” (Fig. 5) were habituated to an actor eating a food object with a particular color/texture and shape (e.g., a green sugar-coated object in the shape of a champagne glass). At test, they were shown a trial in which the actor tasted an object with the same color/texture, but new shape (e.g., a green sugar-coated object in the shape of a bowl) vs. a trial in which she tasted an object with a new color/texture and new shape (e.g., an orange smooth object in the shape of a bowl). If infants generalize learning about food objects by color/texture, they were expected to look longer at the test trial in which the actor ate the food with the new color/texture. Infants in the “Generalization-by-Shape Condition” were habituated to an actor eating one kind of food object (e.g., a green sugar-coated object in the shape of a champagne glass). For the test phase, they viewed a trial in which the actor tasted an object with the same shape, but new color/texture (e.g., an orange smooth object in the shape of a champagne glass) vs. a trial in which she tasted an object with a new shape and new color/texture (e.g., an orange smooth object in the shape of a bowl). If infants generalize learning about food objects by shape, they were expected to look longer at the trial in which the actor ate the food with the new shape.
5.10. Method
The method was very similar to Experiment 5, with the following exceptions: The participants were 15 female and 17 male 8-month-old-infants (M = 8 months, 2 days; range = 7 months, 17 days - 8 months, 22 days). Data from 12 additional infants were excluded for experimental error (n=4), failure to habituate (n=4), equipment failure (n=2), parental interference (n=1), and extremely lengthy (>3 SDs) looking during test trials (n=1). The stimuli were solid “foods” made from lightweight modeling clay, sculpted and painted to look like the stimuli used in Experiments 4 and 5. The foods were either in the shape of the champagne glass or in the shape of the bowl and each shape came in four different color/texture combinations: orange paint with smooth finish, orange paint coated with orange sugar, green paint with smooth finish, and green paint coated with green sugar.
On every trial, the actor picked up the food object, pretended to take a bite from the back of it, returned it to the white shelf, and left her hand resting on it. Sixteen infants were tested in the Generalization-by-Color/Texture Condition. During test trials, they saw the actor eat an object with a familiar color/texture and novel shape vs. an object with a novel color/texture and novel shape. Another group of 16 infants were tested in the Generalization-by-Shape Condition. At test, they saw the actor eat an object with a familiar shape and novel color/texture vs. an object with a novel shape and novel color/texture. Reliability between coders was 95%.
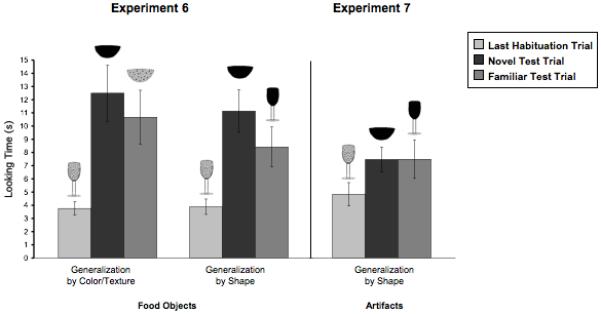
5.11. Results
Infants showed no consistent generalization of habituation to solid food objects, either by substance or by shape (Fig. 7, left). A repeated-measures ANOVA revealed no effect of test trial type (F(1,30) = 1.31, n.s.), no effect of condition (F(1,30) = 1.15, n.s.), and no interaction between the factors. Infants tested with novel shapes and either familiar or novel substances recovered interest from the last habituation trial to both the novel and familiar substance test trials (t(15) = 4.08, t(15) = 3.08, p < .01, respectively). Comparisons of looking at the two test trials revealed equal looking at the trial where the actor ate the food with the novel color/texture as at the trial where she ate the food with a familiar color/texture (paired-sample t<1). Infants tested with novel substances and either familiar or novel shapes also increased their looking from the last habituation trial to each of the test trials, both with the novel shape (t(15) = 4.05, p < .001) and with the familiar shape(t(15) = 2.80, p < .05). They too looked equally long at the trial where the actor ate the food with a novel shape and the trial where she ate the food with a familiar shape (t(15) = 1.25, n.s.).
Figure 7.
Results from Experiments 6 and 7.
5.12. Discussion
Infants in Experiment 6 failed to generalize information about either the shape or the substance properties of solid food objects. As in Experiment 5, infants looked equally long at the novel (color/texture) and familiar (color/texture) test trials in the Generalization-by-Color/Texture Condition. Participants in the Generalization-by-Shape Condition looked equally long at the novel (shape) and familiar (shape) test trials. The significant recovery of looking time toward test trials in both conditions indicates that infants were able to discriminate changes in color/texture and shape. The equal looking times during the two kinds of test trials indicates that they did not prioritize substance or shape information when generalizing learning these objects.
The results from Experiments 5 and 6 cast doubt on the hypothesis that infants show adult-like, domain-specific classification of food objects or food substances. One open question, however, is whether infants’ behavior in these experiments is specific to the domain of food, or whether the same patterns would be observed if infants were tested with artifact objects of similar appearance. Previous work has shown that infants generalize learning about nonobvious properties (e.g., rattling) of artifacts by shape, over changes in texture (Graham et al., 2004; Welder & Graham, 2001). It is not clear whether younger infants will show the same learning and generalization for artifact objects. Experiment 7 used the method and displays of Experiment 6 to ask whether 8-month-old infants attend to shape when generalizing learning about novel artifacts.
5.13. Experiment 7
Infants in Experiment 7 were shown the same stimuli as infants in Experiment 6, however the objects were presented as rattles rather than as foods. Infants were tested only for generalization by shape (across a change in color/texture). During habituation, infants watched trials in which an actor picked up an object with a particular color/texture and shape (e.g., a green sugar-coated object in the shape of a champagne glass) and rattled it twice. For the test phase, infants watched one trial in which the actor used an object with the same shape, but new color/texture (e.g., an orange smooth object in the shape of a champagne glass) vs. trials in which she used an object with a new shape and new color/texture (e.g., an orange smooth object in the shape of a bowl). If young infants generalize learning about artifacts by shape, they were expected to look longer at test trials in which the actor rattled the object with the new shape.
5.14. Method
The method was similar to Experiment 6, with the following exceptions: The participants were 16 8-month-old infants (7 females; M=8 months, 4 days; range = 7 months, 20 days - 8 months, 20 days). The data of 7 additional infants were excluded from analyses for failure to habituate (n=4), fussiness (n=2), and parental interference (n=1).
Instead of tasting the objects, the actor used them as rattles. To accomplish the rattling noise, an experimenter (standing behind the actor, not visible to infants), shook a cup filled with beads in synchrony with the actor’s shaking motions. For the familiarization trial, the actor picked up the object, said “what’s this?” to draw infants’ attention, raised the object to the side of her face, shook the object twice, said “wow”, placed the object on the white shelf, and then looked down. On habituation and test trials, the actor simply picked up the object, raised it to the side of her face, shook it twice, placed the object on the white shelf, and then looked down. As in the previous studies, infants’ looking was recorded once the actor’s hand came to rest on the object.
All 16 infants were tested for generalization by shape (across a change in color/texture). On the familiar test trial, the actor shook an object with a familiar shape and novel color/texture. On novel test trial, the actor shook an object with a novel shape and novel color/texture. Both of the test objects made the same rattling noise as the habituation object. Reliability between coders was 94%.
5.15. Results
Infants showed no consistent generalization by object shape (Fig. 7, right). After habituation to a rattle of one shape and color, infants looked equally at two test rattles that both had novel substance properties, even though one had the same shape as the habituation rattle (t<1). Infants looked significantly longer at the novel test trial than at the last habituation trial (t(15) = 3.46, p < .05). The means for the last habituation trial and the first familiar test trial were in the predicted direction, but the difference was not significant (t(15) = 1.51, n.s.).
To compare the results of Experiment 7 to the analogous condition of Experiment 6 (i.e., Generalization-by-Shape with foods), a repeated-measures ANOVA was conducted with experiment (6 vs. 7) as a between-subject factor and test trial type (novel vs. familiar) as a within-subject factor. The analysis revealed no effect of experiment (F(1,30) = 2.73, n.s.), no effect of trial type (F<1), and no interaction between experiment and trial type (F<1).
5.16. Discussion
Infants in Experiment 7 failed to generalize learning about artifacts across a change in color/texture: They looked equally long at the familiar (shape) and the novel (shape) trial. This result contrasts with previous findings that older infants (13-24 months) classify artifact objects according to shape, over changes in color and texture (Graham et al., 2004; Welder & Graham, 2001). The divergent findings may be due to differences in age, as infants in the present study were younger than participants in the previous studies (e.g., Baldwin, Markman, & Melartin, 1993; Graham et al., 2004; Welder & Graham, 2001). Alternatively, the difference may be the result of a change in method, as earlier studies tested infants in the context of reaching or exploratory play.
Infants showed virtually identical patterns of looking in Experiment 7 and the Generalization-by-Shape Condition of Experiment 6. Thus, infants responded to property changes in the same manner, regardless of whether the objects were eaten as foods or used as artifacts.
6. General Discussion of Part 3
The results from Experiments 4-6 provide evidence that 8-month-old infants detect and remember properties of food substances, objects, and containers when observing eating events. We found no evidence, however, that infants use these properties to track food substances over changes in their containers or shapes. Likewise, we found no evidence that infants track solid objects - whether food containers, food objects, or rattles - by their shapes, over changes in their contents or substance properties. Unlike adults, young children, and nonhuman primates, the infants in our experiments did not privilege changes in food color/texture over changes in food (container or object) shape. Moreover, infants in Experiment 7 did not generalize learning about artifacts by shape, over changes in color/texture.
The results of Experiment 7 are especially surprising in light of previous studies that have demonstrated shape-based learning by infants. Older infants generalize learning about novel artifacts (e.g., rattles) by shape, over changes in texture (e.g., Graham et al., 2004; Welder & Graham, 2001). Thus, one question raised by the current work is why infants in Experiment 7 did not demonstrate shape-based learning about novel artifacts. One possibility is that young infants privilege shape information only when an object’s shape is saliently related to its function. An object’s capacity to rattle, however, is often unrelated to its shape or external features. Perhaps young infants would demonstrate shape-based learning if they were taught about objects whose functions and forms were related (e.g., Brown, 1990; Imai & Genter, 1997).
Whatever the reasons for the negative findings of Experiment 7, the experiments in this section cast doubt on the hypothesis that infants show selective learning and generalization about foods and nonfoods, privileging substance properties for food. In this respect, the findings converge with the findings of the experiments in Part 2, in which infants failed to show selective individuation of food objects by privileging substance properties. Beyond the findings in Part 2, the present studies show that infants fail to show domain-specific processing of food objects even when they have observed another person tasting the objects, providing information that the objects are edible. Together, Experiments 2-7 provide evidence for striking commonalities in infants’ processing of objects across the food and artifact domains.
The findings of Experiments 4-7 contrast markedly with those of Experiment 1. Whereas adult rhesus monkeys showed distinctive patterns of generalization of learning for foods and artifacts, human infants did not. We now turn to this contrast, and its implications for the core knowledge hypothesis.
7. General Discussion
Do humans and nonhuman primates have a dedicated system for detecting and categorizing foods? The present experiments began to address this question through studies of monkeys tested with unfamiliar food objects, and through studies of human infants. On the one hand, Experiment 1 provides evidence that adult monkeys show domain-specific learning about food objects, privileging color over shape. Because they live on an island that provides limited natural food objects, and receive half of their nourishment from monkey chow, these findings provide suggestive evidence for such a core system.
Nevertheless, evidence from these monkeys is not sufficient, in itself, to provide evidence for a system of core knowledge. Though adult monkeys on Cayo Santiago have had less experience with diverse foods than monkeys living in the wild, they have had opportunities to engage in food selection and learning. Studies of infants are therefore still necessary to shed further light on whether perception and categorization of foods is guided by a core system of knowledge in monkeys or humans.
Although the relevant studies have not been conducted for infant monkeys, the experiments in Part 3 tested for core knowledge of food in human infants. In contrast to adult monkeys, human infants showed no evidence for domain-specific parsing and learning about foods. The contrasting findings from infants and nonhuman primates are particularly striking because very similar methods were used with the two populations. The studies of object individuation in monkeys and human infants used identical displays and similar procedures. The studies of property generalization employed the same design strategy of presenting the same objects, and using them in different ways (i.e., either as foods or as artifacts). Additionally, in the present work, both monkeys and human infants were tested with preferential-looking methods. Moreover, some of the present findings accord with previous results from other studies of rhesus monkeys and human infants: Santos et al. (2001) found domain-specific property generalization for foods and tools using a forced-choice method. Additionally, Spelke et al. (1993) found that infants tested with artifacts showed very similar findings to the parsing results of Experiments 2 and 3. These reasons make it unlikely that differences in methods account for the differences between the observed capacities of monkeys vs. human infants.
How can we account for these contrasting findings? One possibility is that human infants possess a core system for reasoning about foods according to relevant visual properties, but that our methods did not detect this system. For example, Experiments 2 and 3 presented infants with food objects but, consistent with the monkey studies on which they were based, they did not show infants that the objects were edible either by feeding the infant or by allowing the infant to observe the eating of another person. Although the subsequent experiments did provide such information, and yielded similarly negative findings, future research could repeat the methods of Experiments 2 and 3, but provide infants with richer cues regarding the edibility of the objects.
As a second possibility, 8-month-old infants may know that color and texture are more relevant properties than shape in the food domain, but the methods of Experiments 4-6 may not have been sensitive enough to detect this knowledge because the test events present changes in both shape and substance. Numerous studies of infants cast doubt on this possibility: Research in which multiple properties of displays change from the habituation or familiarization phase to the test phase and infants’ looking toward one test stimulus vs. another is compared are quite common in research with infants, and these studies often reveal significant looking time differences and positive effects (e.g., Eimas & Quinn, 1994; Hespos, Ferry, & Ripps, in press; Hespos, Saylor, & Grossman, 2009; Kellman & Spelke, 1983; Quinn & Eimas, 1996; Woodward, 1998). Nevertheless, perhaps our displays were too complex or perhaps the property differences we presented were too extreme to show successful generalization by infants. Additional research on infants’ categorization of food objects and substances could present more dramatic changes in color/texture and less dramatic changes in object and container shape, in order to ask whether infants are capable of generalizing learning about foods under simpler or more salient conditions. Finally, future studies could employ methods such as reaching and crawling to assess the development of infants’ and toddlers’ perception and categorization in the food domain.
A third possibility is that there is a core system for representing food in accord with substance information and other natural properties, but it emerges later in human development. This explanation seems especially plausible since human infants, like other mammals, have a long period of nursing (Rozin & Pelchat, 1988) and therefore are not actively engaged in their own food selection. Because parents are largely responsible for their young children’s diets, infants and toddlers may not need mechanisms for determining whether a food is familiar or novel, safe or hazardous, or nutritious or nonnutritive in early infancy. In support of this idea, studies by Rozin and colleagues have shown that infants and toddlers are willing to put nearly anything in their mouths, including entities that are inedible, disgusting, and dangerous (Rozin, Fallon, & Augustoni-Ziskind, 1986; Rozin, Hammer, Oster, Horowitz, & Marmora, 1986). In fact, though children begin to use sensory features to guide food selection in the preschool years, a complete mature food rejection taxonomy does not seem to emerge until middle childhood (Fallon et al., 1984; Rozin, Fallon, & Augustoni-Ziskind, 1985).
Cashdan (1994, 1998) has proposed that the first two to three years of life is a sensitive period for learning about food. In support of this assertion, Cashdan notes that while children aged one to two years are very willing to try new foods, their receptiveness declines significantly between two and four years of age. In order to display neophobia toward foods, of course, children must be able to use perceptible properties such as color and texture to determine which foods are novel and which are familiar. Interestingly, the children who participated in the food categorization studies conducted by Lavin and Hall (2002), Macario (1991), and Santos et al. (2002) were all around the age at which children typically begin to show neophobic reactions to foods.
A final possibility is that there is no core system dedicated to the identification and categorization of foods. Instead, humans and monkeys may learn which properties are relevant to individuating and categorizing food objects. For example, human infants may learn that color and textural properties are correlated with how food objects taste, and that shape properties are correlated with how artifact objects function. Similarly, monkeys in the present population may learn about substance-taste correlations by eating dirt vs. leaves, and they may learn about object functions perhaps by opening food bins and manipulating water dispensers on the island.
An additional mechanism - available both to human infants and to monkeys - allows learning about different kinds of objects by observation of the behaviors of others (e.g., Csibra & Gergely, 2006; Mumme & Fernald, 2003; Tomasello, Kruger, & Ratner, 1993). In the domain of food, for example, the choices and preferences of others are available to guide infants’ early learning and food selection. Many animals, including rats (e.g., Galef & Whiskin, 1995), lambs (e.g., Mirza & Provenza, 1990), and chimpanzees (e.g., Ueno & Matsuzawa, 2005), are influenced by the behaviors of conspecifics when making their own food selections (see Galef, 1996 and Galef & Beck, 1990, for reviews).
Studies of young children have also shown an effect of social modeling on children’s food acceptance (e.g., Addessi, Galloway, Visalberghi, & Birch, 2005; Birch, 1980; Duncker, 1938; Harper & Sanders, 1975; Hendy & Raudenbush, 2000; Shutts, Kinzler, McKee, & Spelke, in press). For example, 12-month-old infants preferentially reach for a food endorsed by an adult speaker of their own language over an adult speaker of a foreign language, even when infants know that both foods are highly palatable (Shutts, Kinzler et al., in press). Thus, even infants may learn food preferences from observing choices of those in their culture (Rozin, 1988; 2007; c.f. Rozin, 1991).
Although the present research does not decide among these accounts, its results make two contributions. First, the findings limit the space of hypotheses concerning the origins of domain-specific reasoning about foods. If there is a core system for learning about foods, then it differs from other systems of core knowledge in humans by emerging considerably later. If general-purpose mechanisms support learning about foods, then this learning can proceed under somewhat impoverished experiences in monkeys. Second, the findings illustrate a research strategy that can be pursued to address these questions, by studying primates raised under more tightly controlled conditions, and by studying developmental changes in human infants, paced both to changes in experience and in maturational state. We hope that the present research, and the contrasting evidence it has yielded, serves as an impetus in this direction.
Acknowledgements
The rhesus macaque studies were supported by an NIH (NCRR grant CM-5-P40RR003640-13) award to the Caribbean Primate Research Center and the University of Puerto Rico-Medical Sciences Campus, as well as by Harvard University. The authors would like to thank Cory Miller and Marc Hauser for their help in designing and running these studies. The authors are also grateful to James Ayala and Melissa Gerald for her help in securing the Cayo Santiago field station. This work was approved by the Cayo Santiago IACUC committee and conforms to guidelines for the use of animals in research.
The infant studies were supported by NIH grant HD23103 to Elizabeth Spelke. The authors thank Ariel Grace and Elizabet Spaepen for assistance with testing.
References
- Addessi E, Galloway AT, Visalberghi E, Birch LL. Specific social influences on the acceptance of novel foods in 2-5-year-old children. Appetite. 2005;45:264–271. doi: 10.1016/j.appet.2005.07.007. [DOI] [PubMed] [Google Scholar]
- Addessi E, Visalberghi E. Social facilitation of eating novel food in tufted capuchin monkeys (Cebus apella): input provided, responses affected, and cognitive implications. Animal Cognition. 2001;4:297–303. doi: 10.1007/s100710100113. [DOI] [PubMed] [Google Scholar]
- Baldwin D, Markman EM, Melartin RL. Infants’ ability to draw inferences about nonobvious object properties: Evidence from exploratory play. Child Development. 1993;64:711–728. [PubMed] [Google Scholar]
- Barker LM, Best MR, Domjan M, editors. Learning mechanisms in food selection. Baylor University Press; 1977. [Google Scholar]
- Barkow J, Cosmides L, Tooby J, editors. The adapted mind: Evolutionary psychology and the generation of culture. Oxford University Press; New York: 1992. [Google Scholar]
- Bernstein IL. Taste aversion learning: A contemporary perspective. Nutrition. 1999;15:229–234. doi: 10.1016/s0899-9007(98)00192-0. [DOI] [PubMed] [Google Scholar]
- Birch LL. Effects of peer models’ food choices and eating behaviors on preschoolers’ food preferences. Child Development. 1980;51:489–496. [Google Scholar]
- Birch LL. Development of food acceptance patterns. Developmental Psychology. 1990;26:515–519. [Google Scholar]
- Birch LL, Fisher JA. The role of experience in the development of children’s eating behavior. In: Capaldi ED, editor. Why We Eat What We Eat: The Psychology of Eating. APA; Washington, D.C.: 1996. pp. 113–141. [Google Scholar]
- Birch LL, Marlin DW. I don’t like it; I never tried it: Effects of exposure to food on two-year-old children’s food preferences. Appetite. 1982;4:353–360. doi: 10.1016/s0195-6663(82)80053-6. [DOI] [PubMed] [Google Scholar]
- Birch LL, McPhee L, Steinberg L, Sullivan SS. Conditioned flavor preferences in young children. Physiology & Behavior. 1990;47:501–505. doi: 10.1016/0031-9384(90)90116-l. [DOI] [PubMed] [Google Scholar]
- Brown AL. Domain-specific principles affect learning and transfer in children. Cognitive Science. 1990;14:107–133. [Google Scholar]
- Carey S, Xu F. Infants’ knowledge of objects: Beyond object files and object tracking. Cognition. 2001;80:179–213. doi: 10.1016/s0010-0277(00)00154-2. [DOI] [PubMed] [Google Scholar]
- Cashdan E. A sensitive period for learning about food. Human Nature. 1994;5:279–291. doi: 10.1007/BF02692155. [DOI] [PubMed] [Google Scholar]
- Cashdan E. Adaptiveness of food learning and food aversions in children. Social Science Information. 1998;37:613–632. [Google Scholar]
- Condry K, Smith WC, Spelke ES. Development of perceptual organization. In: Lacerda F, von Hofsten C, Heiman M, editors. Emerging cognitive abilities in early infancy. Erlbaum; Hillsdale, NJ: 2001. [Google Scholar]
- Cooke L, Wardle J, Gibson EL. Relationship between parental report of food neophobia and everyday food consumption in 2-6-year-old children. Appetite. 2003;41:205–206. doi: 10.1016/s0195-6663(03)00048-5. [DOI] [PubMed] [Google Scholar]
- Csibra G, Gergely G. Social learning and social cognition: the case for pedagogy. In: Munakata Y, Johnson MH, editors. Processes of change in brain and cognitive development. Attention and performance XXI. Oxford University Press; Oxford: 2006. pp. 249–274. [Google Scholar]
- Domjan M. Attenuation and enhancement of neophobia for edible substances. In: Barker LM, Best MR, Domjam M, editors. Learning mechanisms in food selection. Baylor University Press; Waco, TX: 1977. pp. 151–179. [Google Scholar]
- Duncker K. Experimental modification of children’s food preferences through social suggestion. The Journal of Abnormal and Social Psychology. 1938;33:489–507. [Google Scholar]
- Eimas PD, Quinn PC. Studies on the formation of perceptually based basic-level categories in young infants. Child Development. 1994;65:903–917. [PubMed] [Google Scholar]
- Fallon AE, Rozin P, Pliner P. The child’s conception of food: The development of food rejections with special reference to disgust and contamination sensitivity. Child Development. 1984;55:566–575. [PubMed] [Google Scholar]
- Galef BG. Social influences on food preferences and feeding behaviors of vertebrates. In: Capaldi ED, editor. Why we eat what we eat: The psychology of eating. American Psychological Association; Washington, DC: 1996. pp. 207–231. [Google Scholar]
- Galef BG, Beck M. Diet selection and poison avoidance by mammals individually and in social groups. In: Stricker EM, editor. Handbook of Neurobiology: Neurobiology of Food and Fluid Intake. Vol. 11. Plenum Press; New York: 1990. pp. 329–349. [Google Scholar]
- Galef BG, Whiskin EE. Learning socially to eat more of one food than of another. Journal of Comparative Psychology. 1995;109:99–101. doi: 10.1037/0735-7036.109.1.99. [DOI] [PubMed] [Google Scholar]
- Garb JL, Stunkard AJ. Taste aversion in man. American Journal of Psychiatry. 1974;131:1204–1207. doi: 10.1176/ajp.131.11.1204. [DOI] [PubMed] [Google Scholar]
- Garcia J, Koelling RA. Relation of cue to consequence in avoidance learning. Psychonomic Science. 1966;4:123–124. [Google Scholar]
- Garcia J, Ervin FR, Koelling RA. Learning with prolonged delay of reinforcement. Psychonomic Science. 1966;5:121–122. [Google Scholar]
- Graham SA, Kilbreath CS, Welder AN. 13-month-olds rely on shared labels and shape similarity for inductive inferences. Child Development. 2004;75:409–427. doi: 10.1111/j.1467-8624.2004.00683.x. [DOI] [PubMed] [Google Scholar]
- Gustavson CR. Comparative and field aspects of learned food aversions. In: Barker LM, Best MR, Domjam M, editors. Learning mechanisms in food selection. Baylor University Press; Waco, TX: 1977. pp. 23–43. [Google Scholar]
- Harper LV, Sanders KM. The effect of adults’ eating on young children’s acceptance of unfamiliar foods. Journal of Experimental Child Psychology. 1975;20:206–214. [Google Scholar]
- Hauser MD. Artifactual kinds and functional design features: What a primate understands without language. Cognition. 1997;64:285–308. doi: 10.1016/s0010-0277(97)00028-0. [DOI] [PubMed] [Google Scholar]
- Hendy HM, Raudenbush B. Effectiveness of teacher modeling to encourage food acceptance in preschool children. Appetite. 2000;34:61–76. doi: 10.1006/appe.1999.0286. [DOI] [PubMed] [Google Scholar]
- Hespos SJ, Ferry AL, Rips LJ. Infants have different expectations for solids and liquids. in press. [DOI] [PubMed] [Google Scholar]
- Hespos SJ, Saylor MM, Grossman SR. Developmental Psychology. Infants’ ability to parse continuous actions. in press. [DOI] [PubMed] [Google Scholar]
- Hirschfeld LA, Gelman SA, editors. Mapping the mind: Domain specificity in cognition and culture. Cambridge University Press; New York: 1994. [Google Scholar]
- Huntley-Fenner G, Carey S, Solimando A. Objects are individuals but stuff doesn’t count: Perceived rigidity and cohesiveness influence infants’ representation of small groups of discrete enties. Cognition. 2002;85:203–221. doi: 10.1016/s0010-0277(02)00088-4. [DOI] [PubMed] [Google Scholar]
- Imai M, Gentner D. A crosslinguistic study of early word meaning: Universal ontology and linguistic influence. Cognition. 1997;62:169–200. doi: 10.1016/s0010-0277(96)00784-6. [DOI] [PubMed] [Google Scholar]
- Johnson SP, Aslin RN. Perception of object unity in young infants: The roles of motion, depth and orientation. Cognitive Development. 1996;11:161–180. [Google Scholar]
- Jusczyk PW, Johnson SP, Spelke ES, Kennedy LJ. Synchronous change and perception of object unity: Evidence from adults and infants. Cognition. 1999;71:257–288. doi: 10.1016/s0010-0277(99)00026-8. [DOI] [PubMed] [Google Scholar]
- Kellman PJ, Arterberry ME. The cradle of knowledge: Development of perception in infancy. MIT Press; Cambridge MA: 1998. [Google Scholar]
- Kellman PJ, Spelke ES. Perception of partly occluded objects in infancy. Cognitive Psychology. 1983;15:483–524. doi: 10.1016/0010-0285(83)90017-8. [DOI] [PubMed] [Google Scholar]
- Kestenbaum R, Termine N, Spelke ES. Perception of objects and object boundaries by three-month-old infants. British Journal of Developmental Psychology. 1987;5:367–383. [Google Scholar]
- Lavin TA, Hall DG. Domain effects in lexical development: learning words for foods and toys. Cognitive Development. 2002;16:929–950. [Google Scholar]
- Logue AW, Ophir I, Strauss KE. The acquisition of taste aversions in humans. Behaviour Research and Therapy. 1981;19:319–333. doi: 10.1016/0005-7967(81)90053-x. [DOI] [PubMed] [Google Scholar]
- Macario JF. Young children’s use of color in classification: Foods and canonically colored objects. Cognitive Development. 1991;6:17–46. [Google Scholar]
- Mirza SN, Provenza FD. Preference of the mother affects selection and avoidance of foods by lambs differing in age. Applied Animal Behaviour Science. 1990;28:255–263. [Google Scholar]
- Mumme DL, Fernald A. The infant as onlooker: Learning from emotional reactions observed in a television scenario. Child Development. 2003;74:221–237. doi: 10.1111/1467-8624.00532. [DOI] [PubMed] [Google Scholar]
- Munakata Y, Santos L,R, Spelke ES, Hauser MD, O’Reilly RC. Visual representation in the wild: How rhesus monkeys parse objects. Journal of Cognitive Neuroscience. 2001;13:44–58. doi: 10.1162/089892901564162. [DOI] [PubMed] [Google Scholar]
- Needham A. Factors affecting infants’ use of featural information in object segregation. Current Directions in Psychological Science. 1997;6:26–33. [Google Scholar]
- Needham A, Baillargeon R. Infants’ use of featural and experiential information in segregating and individuating objects: A reply to Xu, Carey, & Welsh. Cognition. 2000;74:255–284. doi: 10.1016/s0010-0277(99)00075-x. [DOI] [PubMed] [Google Scholar]
- Pelchat ML, Rozin P. The special role of nausea in the acquisition of food dislikes by humans. Appetite. 1982;3:341–351. doi: 10.1016/s0195-6663(82)80052-4. [DOI] [PubMed] [Google Scholar]
- Pliner P, Salvy SJ. Food neophobia in humans. In: Shepherd R, Raats M, editors. The psychology of food choice. CABI; Wallingford, UK: 2006. pp. 75–92. [Google Scholar]
- Pliner P, Stallberg-White C. “Pass the ketchup, please”: familiar flavors increase children’s willingness to taste novel foods. Appetite. 2000;34:95–103. doi: 10.1006/appe.1999.0290. [DOI] [PubMed] [Google Scholar]
- Quinn PC, Eimas PD. Perceptual cues that permit categorical differentiation of animal species by infants. Journal of Experimental Child Psychology. 1996;63:189–211. doi: 10.1006/jecp.1996.0047. [DOI] [PubMed] [Google Scholar]
- Rawlins RG, Kessler MG. The Cayo Santiago macaques: History, behavior, and biology. SUNY Press; Albany, NY: 1987. [Google Scholar]
- Revusky SH, Bedarf EW. Association of illness with prior ingestion of novel foods. Science. 1967;155:219–220. doi: 10.1126/science.155.3759.219. [DOI] [PubMed] [Google Scholar]
- Richter CP. Experimentally produced behavior reactions to food poisoning in wild and domesticated rats. Annals of the New York Academy of Sciences. 1953;56:225–239. doi: 10.1111/j.1749-6632.1953.tb30219.x. [DOI] [PubMed] [Google Scholar]
- Ross BH, Murphy GL. Food for thought: Cross-classification and category organization in a complex real-world domain. Cognitive Psychology. 1999;38:495–553. doi: 10.1006/cogp.1998.0712. [DOI] [PubMed] [Google Scholar]
- Rozin P. The selection of food by rats, humans and other animals. In: Rosenblatt J, Hinde RA, Beer C, Shaw E, editors. Advances in the study of behavior. Vol. 6. Academic Press; New York: 1976. pp. 21–76. [Google Scholar]
- Rozin P. The significance of learning mechanisms in food selection: Some biology, psychology, and sociology of science. In: Barker LM, Best MR, Domjam M, editors. Learning mechanisms in food selection. Baylor University Press; Waco, TX: 1977. pp. 557–581. [Google Scholar]
- Rozin P. Social learning about food by humans. In: Zentall TR, Galef BG, editors. Social learning: Psychological and biological perspectives. Lawrence Erlbaum Associates; Hillsdale, NJ: 1988. pp. 165–187. [Google Scholar]
- Rozin P. Development in the food domain. Developmental Psychology. 1990;26:555–562. [Google Scholar]
- Rozin P. Sociocultural influences on human food selection. In: Capaldi ED, editor. Why We Eat What We Eat: The Psychology of Eating. APA; Washington, D.C.: 1996. pp. 113–141. [Google Scholar]
- Rozin P. Food and eating. In: Kitayama S, Cohen D, editors. Handbook of cultural psychology. The Guilford Press; New York, NY: 2007. pp. 391–416. [Google Scholar]
- Rozin P, Fallon AE. A perspective on disgust. Psychological Review. 1987;94:23–41. [PubMed] [Google Scholar]
- Rozin P, Fallon A, Augustoni-Ziskind M. The child’s conception of food: The development of contamination sensitivity to “disgusting” substances. Developmental Psychology. 1985;21:1075–1079. [Google Scholar]
- Rozin P, Fallon A, Augustoni-Ziskind M. The child’s conception of food: The development of categories of acceptable and rejected substances. Journal of Nutrition Education. 1986 a;18:75–81. [Google Scholar]
- Rozin P, Hammer L, Oster H, Horowitz T, Marmora V. The child’s conception of food: Differentiation of categories of rejected substances in the 16 months to 5 year age range. Appetite. 1986 b;7:141–151. doi: 10.1016/s0195-6663(86)80014-9. [DOI] [PubMed] [Google Scholar]
- Rozin P, Kalat JW. Specific hungers and poison avoidance as adaptive specializations of learning. Psychological Review. 1971;78:459–486. doi: 10.1037/h0031878. [DOI] [PubMed] [Google Scholar]
- Rozin P, Pelchat ML. Memories of mammaries: Adaptations to weaning from milk. In: Epstein AN, Morrison AR, editors. Progress in Psychobiology and Physiological Psychology. Vol. 13. Academic Press; New York, NY: 1988. pp. 1–29. [Google Scholar]
- Rozin PN, Schulkin J. Food selection. In: Stricker EM, editor. Handbook of Behavioral Neurobiology: Neurobiology of Food and Fluid Intake. Vol. 10. Plenum Press; New York, NY: 1990. pp. 297–328. [Google Scholar]
- Santos LR, Hauser MD, Spelke ES. Recognition and categorization of biologically significant objects by rhesus monkeys (macaca mulatta): The domain of food. Cognition. 2001;82:127–155. doi: 10.1016/s0010-0277(01)00149-4. [DOI] [PubMed] [Google Scholar]
- Santos LR, Hauser MD, Spelke ES. Domain-specific knowledge in human children and non-human primates: Artifact and food kinds. In: Bekoff M, Allen C, Burghardt G, editors. The Cognitive Animal. MIT Press; Cambridge, MA: 2002. [Google Scholar]
- Santos LR, Miller CT, Hauser MD. Representing tools: How two nonhuman primate species distinguish between the functionally relevant and irrelevant features of a tool. Animal Cognition. 2003;6:269–281. doi: 10.1007/s10071-003-0171-1. [DOI] [PubMed] [Google Scholar]
- Shepherd R, Raats M. The psychology of food choice. CABI; Wallingford, UK: 2006. [Google Scholar]
- Shutts K, Kinzler KD, McKee C, Spelke ES. The effect of social information on 12-month-old infants’ food preferences. Journal of Cognition and Development. doi: 10.1080/15248370902966636. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shutts K, Markson L, Spelke ES. The developmental origins of animal and artifact concepts. In: Hood B, Santos L, editors. The origins of object knowledge. Oxford University Press; Oxford: 2009. [Google Scholar]
- Smith WC, Johnson SP, Spelke ES. Motion and edge sensitivity in perception of object unity. Cognitive Psychology. 2003;46:31–64. doi: 10.1016/s0010-0285(02)00501-7. [DOI] [PubMed] [Google Scholar]
- Spelke ES. Principles of object perception. Cognitive Science. 1990;14:29–56. [Google Scholar]
- Spelke ES. Core knowledge. In: Kanwisher N, Duncan J, editors. Attention and performance: Functional neuroimaging of visual cognition. Vol. 20. Oxford University Press; Oxford: 2004. pp. 29–56. [Google Scholar]
- Spelke ES, Breinlinger K, Jacobson K, Phillips A. Gestalt relations and object perception: A developmental study. Perception. 1993;22:1483–1501. doi: 10.1068/p221483. [DOI] [PubMed] [Google Scholar]
- Spelke ES, Kinzler KD. Core knowledge. Developmental Science. 2007;10:89–96. doi: 10.1111/j.1467-7687.2007.00569.x. [DOI] [PubMed] [Google Scholar]
- Spelke ES, von Hofsten C, Kestenbaum R. Object perception and object-directed reaching in infancy: Interaction of spatial and kinetic information for object boundaries. Developmental Psychology. 1989;25:185–196. [Google Scholar]
- Soja N, Carey S, Spelke ES. Ontological categories guide young children’s inductions of word meaning: Object terms and substance terms. Cognition. 1991;38:179–211. doi: 10.1016/0010-0277(91)90051-5. [DOI] [PubMed] [Google Scholar]
- Stricker EM, editor. Handbook of Behavioral Neurobiology: Neurobiology of food and fluid intake. Vol. 10. Plenum Press; New York, N.Y.: 1990. [Google Scholar]
- Tomasello M, Kruger A, Ratner H. Cultural learning. Behavioral and Brain Sciences. 1993;16:495–552. [Google Scholar]
- Ueno A, Matsuzawa T. Response to novel food in infant chimpanzees: Do infants refer to mothers before ingesting food on their own? Behavioural Processes. 2005;68:85–90. doi: 10.1016/j.beproc.2004.09.002. [DOI] [PubMed] [Google Scholar]
- von Hofsten C, Spelke ES. Object perception and object-directed reaching in infancy. Journal of Experimental Psychology: General. 1985;114:198–212. doi: 10.1037//0096-3445.114.2.198. [DOI] [PubMed] [Google Scholar]
- Welder AN, Graham A. The influence of shape similarity and shared labels on infants’ inductive inferences about nonobvious object properties. Child Development. 2001;72:1653–1673. doi: 10.1111/1467-8624.00371. [DOI] [PubMed] [Google Scholar]
- Woodward AL. Infants selectively encode the goal object of an actor’s reach. Cognition. 1998;69:1–34. doi: 10.1016/s0010-0277(98)00058-4. [DOI] [PubMed] [Google Scholar]
- Xu F, Carey S, Welch J. Infants’ ability to use object kind information for object individuation. Cognition. 1999;70:137–166. doi: 10.1016/s0010-0277(99)00007-4. [DOI] [PubMed] [Google Scholar]