

Abstract
Objective
Increasing evidence suggests that chronic inflammation contributes to atherogenesis, and that acute inflammatory events cause plaque rupture, thrombosis, and myocardial infarction. The present studies examined how inflammatory factors, such as interferon-γ (IFNγ), cause increased sensitivity to apoptosis in vascular lesion cells.
Methods and Results
Cells from the fibrous cap of human atherosclerotic lesions were sensitized by interferon-γ (IFNγ) to Fas-induced apoptosis, in a Bcl-XL reversible manner. Microarray profiling identified 72 INFγ-induced transcripts with potential relevance to apoptosis. Half could be excluded because they were induced by IRF-1 overexpression, which did not sensitize to apoptosis. IFNγ treatment strongly reduced Mcl-1, phospho-Bcl-2 (ser70), and phospho-Bcl-XL (ser62) protein levels. Candidate transcripts were modulated by siRNA, overexpression, or inhibitors to assess the effect on IFNγ-induced Fas sensitivity. Surprisingly, siRNA knockdown of PSMB8 (LMP7), an ‘immunoproteasome’ component, reversed IFNγ-induced sensitivity to Fas ligation and prevented Fas/IFNγ-induced degradation of Mcl-1, but did not protect p-Bcl-2 or p-Bcl-XL. Proteasome inhibition markedly increased Mcl-1, p-Bcl-2, and p-Bcl-XL levels after IFNγ treatment.
Conclusions
While critical for antigen presentation, the immunoproteasome appears to be a key link between inflammatory factors and the control of vascular cell apoptosis, and thus may be an important factor in plaque rupture and myocardial infarction.
Keywords: myocardial infarction, apoptosis, stroke, restenosis, angioplasty, inflammation, Bcl-XL, Bcl-2, Mcl-1, interferon-γ, STAT1, LMP7, PSMB8, Fas, immunoproteasome, microarray
Advancing age, the single strongest risk factor for cardiovascular disease, favors lesion formation in response to injuries resulting from hyperlipidemia, hypertension, and smoking. Particularly in early lesions, apoptosis of monocyte/macrophages contributes to the accumulation of extracellular lipid and proteins1. In later stages, smooth muscle cell-like (SMC-like)/myofibroblasts encapsulate the lipid-rich regions, and their persistence contributes to matrix accumulation and negative remodeling2. However, when apoptosis occurs in the cap of advanced lesions, the plaque is prone to rupture, thereby triggering thrombosis and myocardial infarction. Increased apoptosis is frequently associated with clinically unstable coronary artery lesions3, and there is strong evidence that apoptosis is a major contributing factor to plaque instability and sudden coronary death4, 5.
Inflammation is intimately linked to the Fas/Fas ligand system, which can either drive proliferation or induce apoptosis of lymphocytes based on the particular context6. Cells within human lesions express Fas (CD95), the receptor for Fas ligand, a potent pro-apoptotic factor7. Inflammatory regions, which contain lymphocyte-derived interferon-γ (IFNγ) and Fas ligand, exhibit the highest levels of SMC-like apoptosis8. Activated macrophages induce apoptosis in LDC9 and SMC10 by Fas/Fas ligand interactions. Fas-ligand deficient mice show excessive vascular repair11, and Fas ligand-based gene therapy reduces intimal hyperplasia in animal models12, 13. Transcript profiling of human restenotic lesions observed activation of IFNγ signaling, and IFNγ receptor knockout mice have an attenuated neointimal response to injury14, consistent with an evolving theory that vascular diseases have a chronic inflammatory component15.
Cells grown from fibrous regions of human vascular lesions contain both apoptosis-sensitive and resistant cells, but apoptosis-resistant cells quickly dominate the culture16. Transcript profiling of lesion cells revealed that increased expression of Bcl-XL is a major determinant of resistance to Fas-mediated apoptosis16, 17. Inflammatory cytokines, especially interferons, can increase the apoptotic response to Fas ligation in SMCs7, 18, 19, endothelial cells20, and other cell types21. The present studies employed a microarray-based approach to identify factors that mediate the pro-apoptotic effect of IFNγ on human LDC.
Methods
Cells
Human atherosclerotic lesions were acquired by carotid endarterectomy by IRB-approved protocols. Lesion-derived cells (LDC) were isolated by explant of the fibrous cap, and cultured in M199 with 10% fetal bovine serum (FBS) and gentamicin (50 μg/mL). Microarray analysis16, 22 and α-actin positivity23 indicate that LDC are SMC-like cells.
Apoptosis assay: Small molecule and siRNA inhibition
The sensitivity to apoptosis was examined by determining survival after challenge with a Fas-activating IgM (clone CH11, Upstate Biotechnology). LDC were plated at 50 K/24-well in M199 + 10% FBS for 24 hours prior to overnight serum reduction to 1% FBS. Small molecule inhibitors, such as compstatin (Tocris Bioscience), pefabloc (BioChemika), cathepsin S inhibitor (Z-FL-COCHO, Calbiochem), MG132 (Z-Leu-Leu-Leu-al, Sigma), or epoxomycin (Sigma) were added 4-6 hours prior to IFNγ. LDC were pretreated with siRNA pools (Dharmacon OnTargetPlus) containing 4 siRNAs in Dharmafect for 24-48 hours prior to IFNγ treatment. IFNγ (human recombinant, R&D Systems, 5 ng/50 IU/mL) was added for 20-24 hours before Fas-activating antibody (50-100 ng/mL). After 24 hours, MTT was added for 4 hours, the stained cells were dissolved in DMSO, and the level of reduced MTT was measured by O.D. at 570 nm in a plate reader24.
Transcript profiling
Genome-wide microarray profiling was conducted of IFNγ-responsive transcripts in human LDC in vitro. Total RNA was prepared using RNAzol B and Qiagen RNeasy Mini columns. RNA quantity was assessed by spectrometry and RNA quality was assessed by Agilent Bioanalyzer. Total RNA (10 μg) was reverse transcribed, and biotin-labeled cRNA was produced by T7 in vitro transcription (IVT). Labeled cRNA was fragmented and hybridized to U133A GeneChips (Affymetrix, 22,282 transcripts). The IFNγ response was evaluated in LDC from three different patients.
Data analysis
The raw data was summarized and normalized using GC-RMA in GeneSpring GX7. A paired t-test identified 1,500 transcripts, which were further filtered by a >1.5 fold-change and potential relevance to apoptosis. Gene lists were compared to published interferon-inducible gene lists using LOLA25. Pathway analysis was conducted using Ingenuity Pathway Analysis (Ingenuity Systems).
Retroviral overexpression
Full-length cDNA clones were obtained from Open Biosystems, or RT-PCR amplified from LDC mRNA. Inserts were subcloned into MSCV-IRES-eGFP retroviral vector, upstream of an encephalomyocarditis virus internal ribosome entry site (IRES) cassette linked to the enhanced green fluorescent protein (eGFP) reporter gene.
Quantitative RT-PCR (qRT-PCR)
qRT-PCR of candidate genes was conducted using SYBR Green fluorescence detected with an ABI 7300 real-time PCR. Standard dilutions of the cDNA were used to interpolate relative transcript abundance. HBOA and ZNF were used as control genes to compensate for minor variations in mRNA quality or quantity. Primer sequences are contained in Supplementary Table I.
Western blot analysis
Protein lysates from untreated or IFNγ treated cells were prepared with a protease inhibitor cocktail (Roche) and phosphatase inhibitor cocktail (Sigma). Protein was quantified by BCA (Pierce), and 50 μg of protein was run on a 12% SDS/PAGE gel and transferred to PVDF. Primary antibodies (Supplementary Table III) were detected by peroxidase-labeled secondary antibody and SuperSignal WestDura substrate (Pierce) on a Kodak 2000MM chemiluminescent detection system.
Results
Interferonγ sensitizes lesion-derived cells (LDC) to apoptosis in a Bcl-XL reversible manner
Like SMC17, 18, 26, human LDC are resistant to apoptosis induced by the Fas-activating CH11 IgM (50 ng/mL), causing <10% apoptosis (Figures 1, 3 and 4). However, 24 hour pretreatment with IFNγ (5 ng/mL) synergistically increases the apoptotic response to Fas ligation to 70% apoptosis. When the mitochondrial apoptosis pathway was suppressed with retroviral expression of Bcl-XL17, the small pro-apoptotic effect of IFNγ alone was reduced, and the synergistic effect of IFNγ and Fas ligation was eliminated (Figure 1).
Figure 1. Bcl-XL overexpression overrides IFNγ sensitization.

Lesion cells were stably infected with eGFP or Bcl-XL with eGFP and sensitivity to Fas ligation was examined. IFNγ (5 ng/mL) was preincubated for 24 hours prior to activating anti-Fas CH11 monoclonal IgM (FAS) for 24 hours. Viable cells were quantitated by MTT (O.D. 570 nm). Staurosporine (STAUR, 20 μM) was added in place of IFNγ as a positive control. Data is expressed as percent of control survival (n=4 per group±s.e.m.). No Rx is no treatment. Asterisks are p<0.001 by t-test.
Figure 3. The effect of siRNA-mediated knockdown of PSMB8 on IFNγ-induced apoptotic sensitivity.
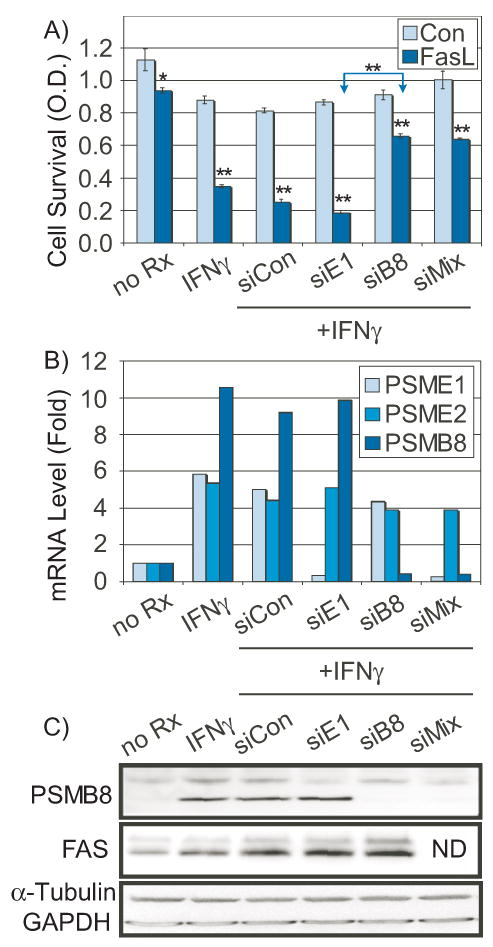
LDC were pretreated either with non-targeted siRNA (siCon), or with siRNA to PSME1 (siE1, 50 nM), PSMB8 (siB8, 50 nM), or a mixture of both (siMIX, 25 nM each). Panel A) IFNγ-induced apoptotic sensitivity to Fas ligation was then determined by MTT survival. Bars are mean ± s.e.m. (n=4, *=p<0.05, **=p<0.001). Panel B) mRNAs were analyzed for effective knockdown of the target transcripts, as well as related transcripts as controls for non-specific effects. Transcript levels of PSME1, PSME2, and PSMB8 were quantitated by qRT-PCR. Panel C) Western blot of PSMB8/LMP7, FAS, and control proteins: α-tubulin (α-Tubu) and glyceraldehyde 3-phosphate dehydrogenase (GAPDH). ND=not determined.
Figure 4. Mcl-1 is regulated by PSMB8 and modulates apoptosis in LDC.
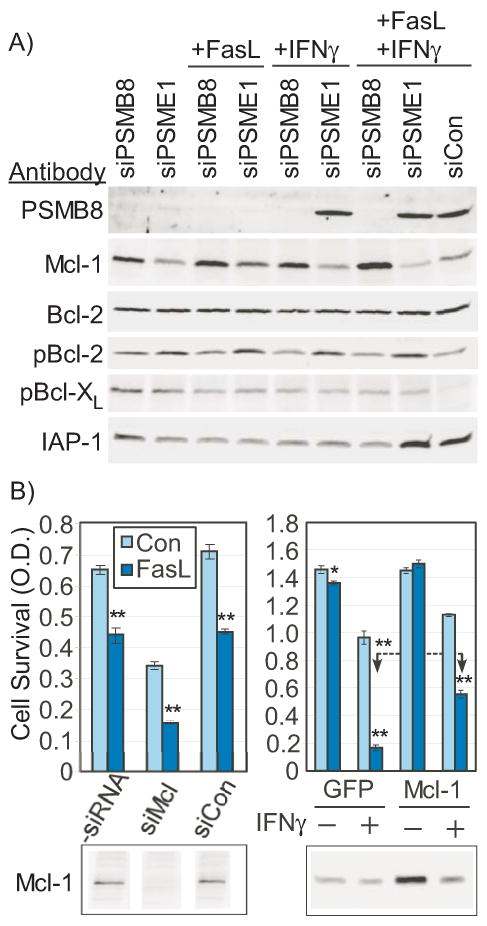
Panel A) LDC were pretreated with siRNA to PSMB8, PSME1, or non-targeted siRNA (siCon) for 48 hours prior to protein harvest and Western blot for candidate apoptosis regulators. Mcl-1 antibody is mouse monoclonal (Santa Cruz). Other antibodies are specified in Supplementary Table II. Panel B) Left panel: Mcl-1 was knocked down by siRNA (siMcl, 50 nM) 48 hours prior to examining sensitivity to apoptosis induced by Fas ligation. Bars are mean optical density (O.D.) of MTT compared to untreated control (Con) or cells pretreated with non-targeted siRNA (siCon, 50 nM). Right panel: Mcl-1 was over-expressed and compared to GFP-only expressing cells for sensitivity to apoptosis triggered by Fas ligation in the presence or absence of 24 hour pretreatment with IFNγ (5 ng/mL). Lower panels demonstrate parallel Western blots for Mcl-1 (mouse monoclonal). Bars are mean O.D. (n=4, *=p<0.05, **=p<0.001).
Transcript profiling of IFNγ-regulated messages in LDC
A full genome microarray profiling was conducted of IFNγ-responsive transcripts in human LDC in vitro. Using CRL1999 (ATCC) and radial artery smooth muscle as reference, the array data indicates that LDC are transcript-positive for 5 consensus smooth muscle markers: smooth muscle α-actin (ACTA2), smooth muscle myosin heavy chain (MYH11), calponin 1 (CNN1), aortic preferentially expressed protein 1 (APEG1), and SM22α/transgelin (TAGLN). Upon IFNγ treatment, the overwhelming majority of affected transcripts were induced (Supplementary Figure I), with some increases as much as 100-fold (INDO). A paired t-test identified almost 1,500 possible genes, which were further filtered by a >1.5 fold-change (135 transcripts), and potential relevance to apoptosis (72 transcripts, Supplementary Table III). These 72 transcripts were prioritized by the strength of their association to apoptosis, and their occurrence in other IFNγ studies, using LOLA25.
qRT-PCR confirmation of microarray results
Many of the key IFNγ-regulated transcripts identified by microarray profiling were confirmed by qRT-PCR of LDC treated with 5 ng/mL IFNγ for 24 hours prior to total RNA isolation (Supp. Table 3). Essentially all transcripts with a greater than 1.5-fold change by microarray were confirmed by qRT-PCR. Marked changes in caspases 1 and 7, IRF-1, STAT1, and proteasome component mRNAs were confirmed, with smaller changes (1.6 fold) in Fas (CD95), caspase 3 and 4 levels, and essentially unchanged levels of caspase 8.
Identification of IRF-1 inducible messages and IRF-1 effect on apoptosis by stable overexpression
Many of the transcriptional effects of IFNγ are mediated directly by STAT family members, and indirectly by induction of IRF-1, a potent transcription factor. IRF-1 was overexpressed in resistant LDC to partially mimic the effect of IFNγ. The majority of IFNγ-inducible messages were induced to a similar degree by IRF-1 overexpression (Supp. Table III). Consistent with prior reports, caspase 1 (interleukin converting enzyme, ICE)27 was strongly induced by IRF-1 expression. However, several transcripts were not elevated by IRF-1, such as Fas (CD95), which is regulated by IRF-828. Notably, PSMB8 (LMP7) was not induced in IRF-1 expressing cells, while other members of the proteasome group were slightly increased in IRF-1 expressers (2-fold), but markedly less than that induced by IFNγ (3.5 to 55-fold). Surprisingly, IRF-1 overexpression did not sensitize cells to apoptosis (not shown), and thus, the IRF-1-induced transcripts can be excluded as candidate factors.
Western blot confirmation of selected IFNγ-inducible proteins
Western blot analysis of the key proteins confirmed that antigen levels were elevated after IFNγ treatment (Figure 2). STAT1 protein was strongly elevated by IFNγ. Although Bcl-XL and Bcl-2 protein, potent anti-apoptotic molecules, were only slightly decreased by IFNγ, their phosphorylated forms, p-Bcl-XL (ser62) and p-Bcl-2 (ser70), were more markedly decreased. P-Bcl-2 (ser70) is a more active anti-apoptotic molecule29, while p-Bcl-XL (ser62) is less active30.
Figure 2. IFNγ effects on candidate apoptosis-related proteins.
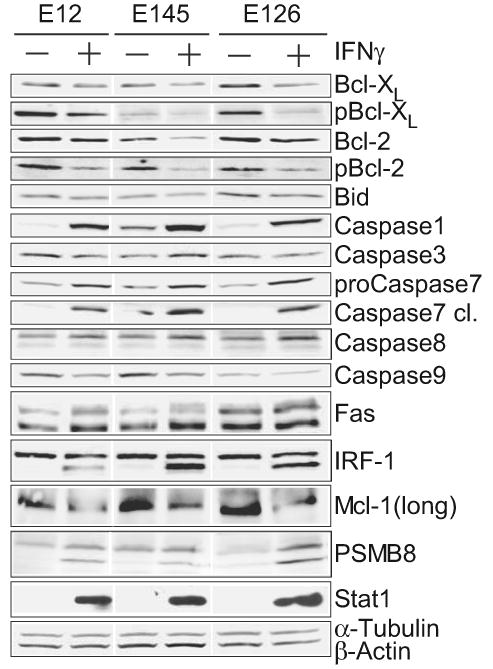
Transcripts identified from microarray analysis, and additional apoptosis-related proteins were confirmed by Western blot in three separate LDC lines (E12, E145, E126) in the absence (-) or presence (+) of IFNγ (5 ng/mL, 24 hours). The specific antibodies used are listed in Supplementary Table II. Mcl-1 antibody is polyclonal rabbit (Cell Signaling), which identifies the 37 kD anti-apoptotic long form.
Consistent with the strong elevation of its mRNA, caspase 1 (ICE) protein was strongly and consistently elevated by IFNγ. Both pro- and cleaved caspase 7 forms were elevated by IFNγ treatment. Caspases 8, but not caspases 3 and 9, was slightly elevated by IFNγ. Fas levels were increased by IFNγ although the levels in some untreated lines (E126) was greater than IFNγ-treated levels in other lines, suggesting that Fas levels alone do not determine sensitivity. Confirming prior work19, the cell surface expression of Fas, determined by flow cytometry, was increased two-fold in IFNγ-treated cells, although among different cell lines Fas surface expression was poorly correlated with apoptotic sensitivity (not shown).
Despite a two-fold increase in Mcl-1 mRNA levels, there was a marked decrease in Mcl-1 protein levels, which could be explained either by translational repression or increased ubiquitin-mediated proteosomal degradation31. Consistent with its mRNA levels, there was a marked increase in PSMB8/LMP7 protein antigen in IFNγ-treated cells.
Retroviral overexpression of candidate apoptosis modulators
A smaller set of candidate genes were examined by stable retroviral overexpression to evaluate their impact on apoptotic sensitivity. Surprisingly, caspase 1 (ICE), which has been reported to mediate apoptotic effects of interferon and IRFs27 in other cell types32, did not alter sensitivity to Fas ligation when overexpressed in LDC17. Other candidate transcripts: butrophylin (BTN3A3), caspase 7, ceramide kinase (CERK), G1P2, GBP1, IFITM1, phospholipid scramblase (PLSCR1), TRIM22, and VAMP4, were likewise ineffective.
Small-molecule inhibition
Certain IFNγ-induced targets were amenable to small-molecule inhibition. The complement pathway was strongly modulated by IFNγ, with induction of C1QR1, C1R, C1S, C3, C4A, and H Factor 1 (HF1). However, compstatin (10-50 μM), a complement inhibitor, was ineffective at blocking the IFNγ effect. Cathepsin S, which has known pro-apoptotic effects, was blocked with Z-FL-COCHO (0.02-2 μM, Calbiochem), but no effect on the IFNγ response was observed. NK4/IL32 is a ligand and activator for PR3, a proapoptotic serine protease. PR3 was blocked by the serine protease inhibitor Pefabloc (1-100 μM), but apoptotic sensitivity was unaltered.
siRNA-mediated knockdown of candidate apoptotic modulators
Using 4 siRNAs per target transcript, 14 candidate transcripts were targeted, including ADAR, ApoL6, caspase 7, caspase 9, CD47, CRADD/RAIDD, IL32/NK4, PKR, RIPK2, UBE1L, without abrogating the IFNγ-induced sensitization to apoptosis. However, as a positive control, siRNA-mediated knockdown of Bid completely blocked the IFNγ effect (not shown).
siRNA knockdown of PSMB8 blocks IFNγ-induced sensitivity
Unlike the prior 14 candidates, which serve as excellent controls for non-specific effects of siRNA, PSMB8/LMP7 knockdown clearly reversed IFNγ-induced sensitivity. As shown in Figure 3A, LDC treated with 5 ng/mL of IFNγ prior to Fas ligation (CH11, 50 ng/mL) show an increased sensitivity to apoptosis (83% survival reduced to 39% survival by INFγ) which is almost fully reversed (restored to 72% survival) by knockdown of PSMB8/LMP7 (50 nM siRNA), but not by knockdown of PSME1 (PA28). A 1:1 mix of siRNA to PSMB8/LMP7 and PSME1 (25 nM each) was equally effective at reversing the IFNγ effect. Non-targeted siRNA did not alter the response to IFNγ. Further, knockdown of PSMB9 (LMP2) or PSMB10 (MECL1), or combined knockdown, was ineffective in blocking IFNγ-induced sensitivity to Fas ligation (not shown). These experiments were repeated five times with similar results.
The specificity of the siRNA effect was evaluated by qRT-PCR for the target transcripts and closely related members of the immunoproteasome. As shown in Figure 3B, the siRNA was very specific, reducing the IFNγ-induced target transcripts to below their basal levels with only minimal effects on other members of the immunoproteasome. For further confirmation, siRNA to PSMB8/LMP7 reduced IFNγ-induced PSMB8/LMP7 antigen to unstimulated levels, as shown in Figure 3C. However, neither PSME1, nor non-targeted siRNA blocked IFNγ-induced PSMB8/LMP7 antigen levels. While PSMB8/LMP7 was essentially completely blocked, the induction of Fas antigen was not blocked by any of the siRNAs, and despite the elevated Fas levels, the PSMB8/LMP7 knockdown blocked sensitivity to apoptosis.
Mcl-1 is anti-apoptotic for LDC and regulated by PSMB8/LMP7
To identify the specific apoptotic regulators that were modulated by PSMB8/LMP7, candidate factors with a known modulation by the proteasome were analyzed by Western blot. SiRNA knockdown of PSMB8 consistently elevated Mcl-1 protein levels, which was most pronounced with combined IFNγ/Fas challenge, compared to PSME1 or non-targeted siRNA (Fig. 4A). The inhibitor of apoptosis proteins (IAP1, IAP2, XIAP), which have been previously described to be proteasome-regulated33, were not elevated by PSMB8 knockdown (IAP1 shown, Fig 4A). Likewise, Bcl-2, pBcl-2, and pBcl-XL were not elevated by PSMB8/LMP7 knockdown (Fig. 4A).
To determine whether regulation of Mcl-1 levels was a viable mode of apoptotic regulation in LDC, Mcl-1 was knocked down by siRNA and the effect on Fas-induced apoptosis was determined. Mcl-1 knockdown reduced survival by approximately 50% and increased the apoptotic response to Fas ligation from 32% to 55%, yielding a combined increase in killing from 32% to 77% (Fig. 4B, Left). Western blot confirmed essentially complete knockdown of Mcl-1 antigen without an effect of a non-targeted control siRNA (siCon). Conversely, retroviral overexpression of Mcl-1 markedly reduced the sensitivity to apoptosis (72% to 46% death) in the presence of IFNγFig. 4B, Right). However, the IFNγ-induced degradation of Mcl-1 severely limited the degree to which Mcl-1 could be overexpressed, as measured by Western blot (Fig. 4B). Thus, Mcl-1 is a potent survival factor, and Mcl-1 levels are increased by PSMB8 knockdown.
Mcl-1 levels are protected by proteasome inhibition
To confirm that Mcl-1 is proteasomally degraded, LDC were IFNγ-treated in the presence or absence of proteasome inhibitors for 24 hours prior to protein harvest. Co-treatment with IFNγ and MG132 or epoxomycin strikingly increased levels of Mcl-1 above untreated levels, and modestly increased the levels of p-Bcl-2 and p-Bcl-XL (Fig. 5). After proteasome inhibition, p-Bcl-2 antigen accumulates in higher mass forms consistent with ubiquitinated protein.
Figure 5. Effect of proteasome inhibition on Mcl-1, p-Bcl-2, and p-Bcl-XL levels.
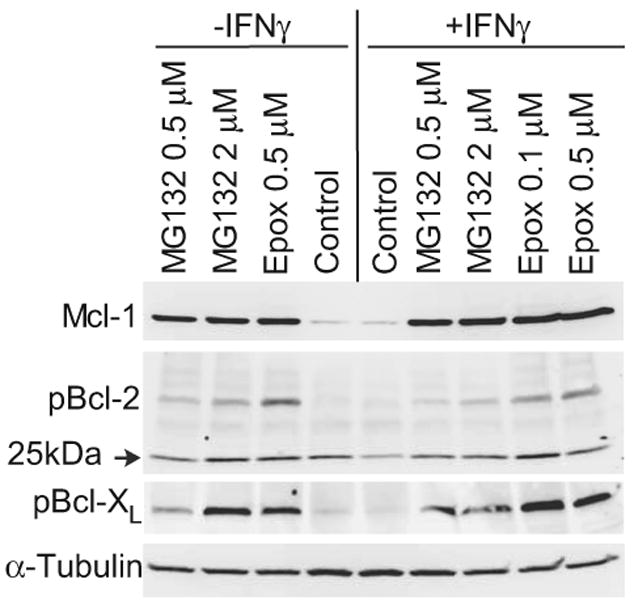
LDC were treated with proteasome inhibitors MG132 or epoxomycin alone (left lanes), or in combination with IFNγ (5 ng/ml) for 24 prior to protein collection and Western blot for Mcl-1 (mouse monoclonal), p-Bcl-2(ser70), or p-Bcl-XL(ser62). Equal protein loading of the lanes is confirmed by antibody to α-tubulin.
Discussion
The control of apoptosis during vascular repair may be a key determinant of both lesion progression and plaque rupture. Thus, understanding the immune and inflammatory influences on apoptosis in vascular cells may facilitate preventative and therapeutic strategies. A systematic examination of potential IFNγ targets revealed the unexpected role of the immunoproteasome component PSMB8/LMP7 as a major component of IFNγ-induced sensitivity. The immunoproteasome has been almost exclusively studied in the context of its role in antigen presentation, although recent studies in double MECL1 and LMP7 deficient T cells suggests that T cell proliferation is also affected by immunoproteasome activity34.
PSMB8/LMP7 is a well-known stress35 and IFNγ-inducible component of the immunoproteasome. The PSMB8 protein product, LMP7, replaces the constitutive β5 subunit, hence its alternate designation as β5i. PSMB8 has been reported to be induced by IFNγ through IRF-136, although our promoter analysis indicates there is only a single IRF-1 site (-2580 to TSS), but five STAT sites (-4574, -4563, -3406, -2160, -1148)37, which is consistent with the present data that IRF-1 overexpression did not induce PSMB8, while IFNγ strongly induced it (>10×). As shown in Figure 6, the three catalytic units of the constitutive proteasome are replaced after IFNγ treatment, as well as key changes to the regulatory subunits (PA28 α and β). The result is an ‘immuno’proteasome which cleaves proteins into short peptide antigens for presentation by MHC I38. The immunoproteasome, however, can activate NFκB and degrade IκB39.
Figure 6. Schematic representation of the interferon-induced immunoproteasome conversion.
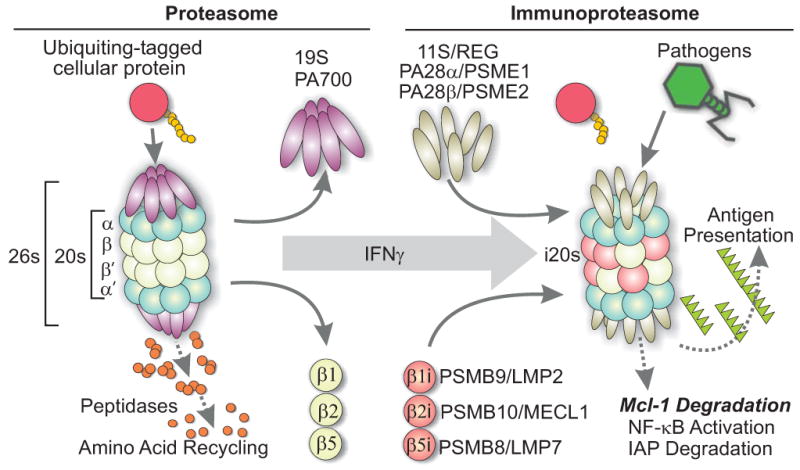
The constitutive 26S proteasome is composed of the 19S regulatory and the 20S catalytic subunits, each having several protein units. In immune activated cells, interferons and other activators trigger transcriptional increases in at least 5 subunits which cooperatively form an immunoproteasome with entirely new catalytic subunits (β1i, β2i, β5i), and new regulatory subunits PA28 α/β/γ. The immunoproteasome has a different substrate specificity and catalytically produces larger peptide fragments, which are better, suited to presentation by MHC class I. Its role in apoptosis could be via degradation of apoptotic inhibitors such as Mcl-1.
The present results suggest a straightforward mechanism for the apoptotic sensitivity associated with immunoproteasome induction. Mcl-1, a potent prosurvival factor, is ubiquitinated by the MULE ubiquitin ligase and degraded by the proteasome31, 40. While other substrates could be affected, it is clear that induction of immunoproteasome activity accelerates Mcl-1 degradation, thereby reducing the survival ability of the cell. Evidence from siRNA knockdown of PSMB8, Mcl-1, and overexpression of Mcl-1 strongly suggests a significant role for this mechanism in mediating the pro-apoptotic effects of IFNγ via the immunoproteasome. Additional effects of IFNγ on p-Bcl-2 and Fas receptor levels may also contribute importantly to the overall sensitivity to apoptosis.
Consistent with a possible in vivo relevance for the present results, important changes in the ubiquitin-proteasome system are observed in age-related atherosclerosis41. Stroke-prone, unstable carotid artery lesions exhibit elevated inflammatory markers and increased proteasome activity42. There are well-known changes in proteasome and immunoproteasome activities during the aging process43, 44, which might result from inflammatory stimuli, interferon activity, and result in altered apoptotic sensitivity. Likewise, changes in the immunoproteasome response to interferon is a feature of senescent cells45. While a general connection between inflammation, atherosclerosis, and myocardial infarction is well established, the precise molecular connections are only beginning to be elucidated. For instance, epidemiological evidence suggests that influenza infection is a strong risk for myocardial infarction46. Likewise, influenza47 and other viral infections48 are potent activators of the immunoproteasome. Combined, the present results identify a novel, and potentially important connection between immune activation and the control of vascular apoptosis.
Supplementary Material
Acknowledgments
a) Sources of Funding: The present studies were supported in part by a MERIT Award from the National Institutes on Aging (AG12712 to TM), a generous endowment to The Catherine Birch McCormick Genomics Center (TM), as well as generous financial support from the St. Laurent Institute (TM, GSL). b) Acknowledgements: The authors are grateful to Teresa Hawley for assistance with cell sorting, and to Robert Hawley and Ali Ramnani (all at GW Medical Center) for assistance with retroviral expression vectors.
The abbreviations used are
- DISC
death-inducing signaling complex
- HEPES
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid
- IFNγ
interferon–gamma
- LDC
lesion-derived cells
- MTT
3-(4,5-dimethylthiazol-2yl)-2,5-diphenyltetrazolium bromide
- eGFP
enhanced Green Fluorescent Promoter
- FACS
fluorescence-activated cell sorting
- GAPDH
glyceraldehyde 3-phosphate dehydrogenase
- PAGE
polyacrylamide gel electrophoresis
- qRT-PCR
quantitative reverse transcriptase-polymerase chain reaction
- SDS
sodium dodecyl sulfate
- TBS
Tris-buffered saline
- TBST
TBS-Tween-20
Footnotes
Disclosure: The authors have no competing financial interests.
References
- 1.Geng YJ, Libby P. Progression of atheroma: a struggle between death and procreation. Arterioscler Thromb Vasc Biol. 2002;22:1370–1380. doi: 10.1161/01.atv.0000031341.84618.a4. [DOI] [PubMed] [Google Scholar]
- 2.Rembold C. Could atherosclerosis originate from defective smooth muscle cell death? Persp Biol Med. 1996;39(3):405–408. doi: 10.1353/pbm.1996.0000. [DOI] [PubMed] [Google Scholar]
- 3.Bauriedel G, Hutter R, Welsch U, Bach R, Sievert H, Luderitz B. Role of smooth muscle cell death in advanced coronary primary lesions: implications for plaque instability. Cardiovasc Res. 1999;41:480–488. doi: 10.1016/s0008-6363(98)00318-6. [DOI] [PubMed] [Google Scholar]
- 4.Kolodgie FD, Narula J, Haider N, Virmani R. Apoptosis in atherosclerosis. Does it contribute to plaque instability? Cardiol Clin. 2001;19:127–139. doi: 10.1016/s0733-8651(05)70199-5. ix. [DOI] [PubMed] [Google Scholar]
- 5.Kolodgie FD, Narula J, Burke AP, Haider N, Farb A, Hui-Liang Y, Smialek J, Virmani R. Localization of apoptotic macrophages at the site of plaque rupture in sudden coronary death. Am J Pathol. 2000;157:1259–1268. doi: 10.1016/S0002-9440(10)64641-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Budd RC. Death receptors couple to both cell proliferation and apoptosis. J Clin Invest. 2002;109:437–441. doi: 10.1172/JCI15077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Geng YJ, Henderson LE, Levesque EB, Muszynski M, Libby P. Fas is expressed in human atherosclerotic intima and promotes apoptosis of cytokine-primed human vascular smooth muscle cells. Arterioscler Thromb Vasc Biol. 1997;17:2200–2208. doi: 10.1161/01.atv.17.10.2200. [DOI] [PubMed] [Google Scholar]
- 8.Kockx MM. Apoptosis in the atherosclerotic plaque: quantitative and qualitative aspects. Arterioscler Thromb Vasc Biol. 1998;18:1519–1522. doi: 10.1161/01.atv.18.10.1519. [DOI] [PubMed] [Google Scholar]
- 9.Boyle J, Bowyer D, Weissberg P, Bennett M. Human blood-derived macrophages induce apoptosis in human plaque-derived vascular smooth muscle cells fas-ligand/fas interactions. Arterioscler Thromb Vasc Biol. 2001;21:1402–1407. doi: 10.1161/hq0901.094279. [DOI] [PubMed] [Google Scholar]
- 10.Imanishi T, Han DK, Hofstra L, Hano T, Nishio I, Liles WC, Gown AM, Schwartz SM. Apoptosis of vascular smooth muscle cells is induced by Fas ligand derived from monocytes/macrophage. Atherosclerosis. 2002;161:143–151. doi: 10.1016/s0021-9150(01)00631-1. [DOI] [PubMed] [Google Scholar]
- 11.Matter CM, Chadjichristos CE, Meier P, von Lukowicz T, Lohmann C, Schuler PK, Zhang D, Odermatt B, Hofmann E, Brunner T, Kwak BR, Luscher TF. Role of endogenous Fas (CD95/Apo-1) ligand in balloon-induced apoptosis, inflammation, and neointima formation. Circulation. 2006;113:1879–1887. doi: 10.1161/CIRCULATIONAHA.106.611731. [DOI] [PubMed] [Google Scholar]
- 12.Jiang C, Yang YF, Cheng SH. Fas ligand gene therapy for vascular intimal hyperplasia. Curr Gene Ther. 2004;4:33–39. doi: 10.2174/1566523044578022. [DOI] [PubMed] [Google Scholar]
- 13.Luo Z, Garron T, Palasis M, Lu H, Belanger AJ, Scaria A, Vincent KA, Date T, Akita GY, Cheng SH, Barry J, Gregory RJ, Jiang C. Enhancement of Fas ligand-induced inhibition of neointimal formation in rabbit femoral and iliac arteries by coexpression of p35. Hum Gene Ther. 2001;12:2191–2202. doi: 10.1089/10430340152710531. [DOI] [PubMed] [Google Scholar]
- 14.Zohlnhofer D, Richter T, Neumann F, Nuhrenberg T, Wessely R, Brandl R, Murr A, Klein C, Baeuerle P. Transcriptome analysis reveals a role of interferon-γ in human neointima formation. Molecular Cell. 2001;7:1059–1069. doi: 10.1016/s1097-2765(01)00239-8. [DOI] [PubMed] [Google Scholar]
- 15.Hansson GK. Inflammation, atherosclerosis, and coronary artery disease. N Engl J Med. 2005;352:1685–1695. doi: 10.1056/NEJMra043430. [DOI] [PubMed] [Google Scholar]
- 16.Gagarin D, Yang Z, Butler J, Wimmer M, Du B, Cahan P, McCaffrey TA. Genomic profiling of acquired resistance to apoptosis in cells derived from human atherosclerotic lesions: potential role of STATs, cyclinD1, BAD, and Bcl-XL. J Mol Cell Cardiol. 2005;39:453–465. doi: 10.1016/j.yjmcc.2005.01.015. [DOI] [PubMed] [Google Scholar]
- 17.Yang Z, Gagarin D, Ramezani A, Hawley RG, McCaffrey T. Resistance to fas-induced apoptosis in cells from human atherosclerotic lesions: Elevated Bcl-Xl inhibits apoptosis and caspase 8 activation. J Vasc Res. 2007;44:483–494. doi: 10.1159/000106466. [DOI] [PubMed] [Google Scholar]
- 18.Geng YJ, Wu Q, Muszynski M, Hansson GK, Libby P. Apoptosis of vascular smooth muscle cells induced by in vitro stimulation with interferon-gamma, tumor necrosis factor-alpha, and interleukin-1 beta. Arterioscler Thromb Vasc Biol. 1996;16:19–27. doi: 10.1161/01.atv.16.1.19. [DOI] [PubMed] [Google Scholar]
- 19.Rosner D, Stoneman V, Littlewood T, McCarthy N, Figg N, Wang Y, Tellides G, Bennett M. Interferon-gamma induces Fas trafficking and sensitization to apoptosis in vascular smooth muscle cells via a PI3K- and Akt-dependent mechanism. Am J Pathol. 2006;168:2054–2063. doi: 10.2353/ajpath.2006.050473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Li JH, Kluger MS, Madge LA, Zheng L, Bothwell AL, Pober JS. Interferon-gamma augments CD95(APO-1/Fas) and pro-caspase-8 expression and sensitizes human vascular endothelial cells to CD95-mediated apoptosis. Am J Pathol. 2002;161:1485–1495. doi: 10.1016/s0002-9440(10)64424-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chawla-Sarkar M, Lindner DJ, Liu YF, Williams BR, Sen GC, Silverman RH, Borden EC. Apoptosis and interferons: role of interferon-stimulated genes as mediators of apoptosis. Apoptosis. 2003;8:237–249. doi: 10.1023/a:1023668705040. [DOI] [PubMed] [Google Scholar]
- 22.Du B, Fu C, Kent KC, Bush H, Jr, Schulick AH, Kreiger K, Collins T, McCaffrey TA. Elevated Egr-1 in human atherosclerotic cells transcriptionally represses the transforming growth factor-beta type II receptor. J Biol Chem. 2000;275:39039–39047. doi: 10.1074/jbc.M005159200. [DOI] [PubMed] [Google Scholar]
- 23.McCaffrey TA, Consigli S, Du B, Falcone DJ, Sanborn TA, Spokojny AM, Bush HL., Jr Decreased type II/type I TGF-β receptor ratio in cells derived from human atherosclerotic lesions. Conversion from an antiproliferative to profibrotic response to TGF-β1. J Clin Invest. 1995;96:2667–2675. doi: 10.1172/JCI118333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Heo DS, Park JG, Hata K, Day R, Herberman RB, Whiteside TL. Evaluation of tetrazolium-based semiautomatic colorimetric assay for measurement of human antitumor cytotoxicity. Cancer Res. 1990;50:3681–3690. [PubMed] [Google Scholar]
- 25.Cahan P, Ahmad AM, Burke H, Fu S, Lai Y, Florea L, Dharker N, Kobrinski T, Kale P, McCaffrey TA. List of lists-annotated (LOLA): a database for annotation and comparison of published microarray gene lists. Gene. 2005;360:78–82. doi: 10.1016/j.gene.2005.07.008. [DOI] [PubMed] [Google Scholar]
- 26.Chan SW, Hegyi L, Scott S, Cary NR, Weissberg PL, Bennett MR. Sensitivity to Fas-mediated apoptosis is determined below receptor level in human vascular smooth muscle cells. Circ Res. 2000;86:1038–1046. doi: 10.1161/01.res.86.10.1038. [DOI] [PubMed] [Google Scholar]
- 27.Horiuchi M, Yamada H, Akishita M, Ito M, Tamura K, Dzau VJ. Interferon regulatory factors regulate interleukin-1beta-converting enzyme expression and apoptosis in vascular smooth muscle cells. Hypertension. 1999;33:162–166. doi: 10.1161/01.hyp.33.1.162. [DOI] [PubMed] [Google Scholar]
- 28.Yang D, Thangaraju M, Browning DD, Dong Z, Korchin B, Lev DC, Ganapathy V, Liu K. IFN regulatory factor 8 mediates apoptosis in nonhemopoietic tumor cells via regulation of Fas expression. J Immunol. 2007;179:4775–4782. doi: 10.4049/jimmunol.179.7.4775. [DOI] [PubMed] [Google Scholar]
- 29.Ito T, Deng X, Carr B, May WS. Bcl-2 phosphorylation required for anti-apoptosis function. J Biol Chem. 1997;272:11671–11673. doi: 10.1074/jbc.272.18.11671. [DOI] [PubMed] [Google Scholar]
- 30.Upreti M, Galitovskaya EN, Chu R, Tackett AJ, Terrano DT, Granell S, Chambers TC. Identification of the major phosphorylation site in Bcl-xL induced by microtubule inhibitors and analysis of its functional significance. J Biol Chem. 2008;283:35517–35525. doi: 10.1074/jbc.M805019200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhong Q, Gao W, Du F, Wang X. Mule/ARF-BP1, a BH3-only E3 ubiquitin ligase, catalyzes the polyubiquitination of Mcl-1 and regulates apoptosis. Cell. 2005;121:1085–1095. doi: 10.1016/j.cell.2005.06.009. [DOI] [PubMed] [Google Scholar]
- 32.Chin YE, Kitagawa M, Kuida K, Flavell RA, Fu XY. Activation of the STAT signaling pathway can cause expression of caspase 1 and apoptosis. Mol Cell Biol. 1997;17:5328–5337. doi: 10.1128/mcb.17.9.5328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yang Y, Fang S, Jensen JP, Weissman AM, Ashwell JD. Ubiquitin protein ligase activity of IAPs and their degradation in proteasomes in response to apoptotic stimuli. Science. 2000;288:874–877. doi: 10.1126/science.288.5467.874. [DOI] [PubMed] [Google Scholar]
- 34.Caudill CM, Jayarapu K, Elenich L, Monaco JJ, Colbert RA, Griffin TA. T cells lacking immunoproteasome subunits MECL-1 and LMP7 hyperproliferate in response to polyclonal mitogens. J Immunol. 2006;176:4075–4082. doi: 10.4049/jimmunol.176.7.4075. [DOI] [PubMed] [Google Scholar]
- 35.Callahan MK, Wohlfert EA, Menoret A, Srivastava PK. Heat shock up-regulates lmp2 and lmp7 and enhances presentation of immunoproteasome-dependent epitopes. J Immunol. 2006;177:8393–8399. doi: 10.4049/jimmunol.177.12.8393. [DOI] [PubMed] [Google Scholar]
- 36.Namiki S, Nakamura T, Oshima S, Yamazaki M, Sekine Y, Tsuchiya K, Okamoto R, Kanai T, Watanabe M. IRF-1 mediates upregulation of LMP7 by IFN-gamma and concerted expression of immunosubunits of the proteasome. FEBS Lett. 2005;579:2781–2787. doi: 10.1016/j.febslet.2005.04.012. [DOI] [PubMed] [Google Scholar]
- 37.Cheung TH, Kwan YL, Hamady M, Liu X. Unraveling transcriptional control and cis-regulatory codes using the software suite GeneACT. Genome Biol. 2006;7:R97. doi: 10.1186/gb-2006-7-10-r97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Cardozo C, Kohanski RA. Altered properties of the branched chain amino acid-preferring activity contribute to increased cleavages after branched chain residues by the “immunoproteasome”. J Biol Chem. 1998;273:16764–16770. doi: 10.1074/jbc.273.27.16764. [DOI] [PubMed] [Google Scholar]
- 39.Visekruna A, Joeris T, Seidel D, Kroesen A, Loddenkemper C, Zeitz M, Kaufmann SH, Schmidt-Ullrich R, Steinhoff U. Proteasome-mediated degradation of IkappaBalpha and processing of p105 in Crohn disease and ulcerative colitis. J Clin Invest. 2006;116:3195–3203. doi: 10.1172/JCI28804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yang C, Kaushal V, Shah SV, Kaushal GP. Mcl-1 is downregulated in cisplatin-induced apoptosis, and proteasome inhibitors restore Mcl-1 and promote survival in renal tubular epithelial cells. Am J Physiol Renal Physiol. 2007;292:F1710–1717. doi: 10.1152/ajprenal.00505.2006. [DOI] [PubMed] [Google Scholar]
- 41.Marfella R, Di Filippo C, Laieta MT, Vestini R, Barbieri M, Sangiulo P, Crescenzi B, Ferraraccio F, Rossi F, D'Amico M, Paolisso G. Effects of ubiquitin-proteasome system deregulation on the vascular senescence and atherosclerosis process in elderly patients. J Gerontol A Biol Sci Med Sci. 2008;63:200–203. doi: 10.1093/gerona/63.2.200. [DOI] [PubMed] [Google Scholar]
- 42.Marfella R, D'Amico M, Di Filippo C, Baldi A, Siniscalchi M, Sasso FC, Portoghese M, Carbonara O, Crescenzi B, Sangiuolo P, Nicoletti GF, Rossiello R, Ferraraccio F, Cacciapuoti F, Verza M, Coppola L, Rossi F, Paolisso G. Increased activity of the ubiquitin-proteasome system in patients with symptomatic carotid disease is associated with enhanced inflammation and may destabilize the atherosclerotic plaque: effects of rosiglitazone treatment. J Am Coll Cardiol. 2006;47:2444–2455. doi: 10.1016/j.jacc.2006.01.073. [DOI] [PubMed] [Google Scholar]
- 43.Husom AD, Peters EA, Kolling EA, Fugere NA, Thompson LV, Ferrington DA. Altered proteasome function and subunit composition in aged muscle. Arch Biochem Biophys. 2004;421:67–76. doi: 10.1016/j.abb.2003.10.010. [DOI] [PubMed] [Google Scholar]
- 44.Ponnappan S, Ovaa H, Ponnappan U. Lower expression of catalytic and structural subunits of the proteasome contributes to decreased proteolysis in peripheral blood T lymphocytes during aging. Int J Biochem Cell Biol. 2007;39:799–809. doi: 10.1016/j.biocel.2007.01.002. [DOI] [PubMed] [Google Scholar]
- 45.Stratford FL, Chondrogianni N, Trougakos IP, Gonos ES, Rivett AJ. Proteasome response to interferon-gamma is altered in senescent human fibroblasts. FEBS Lett. 2006;580:3989–3994. doi: 10.1016/j.febslet.2006.06.029. [DOI] [PubMed] [Google Scholar]
- 46.Madjid M, Miller CC, Zarubaev VV, Marinich IG, Kiselev OI, Lobzin YV, Filippov AE, Casscells SW., 3rd Influenza epidemics and acute respiratory disease activity are associated with a surge in autopsy-confirmed coronary heart disease death: results from 8 years of autopsies in 34,892 subjects. Eur Heart J. 2007;28:1205–1210. doi: 10.1093/eurheartj/ehm035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chen W, Norbury CC, Cho Y, Yewdell JW, Bennink JR. Immunoproteasomes shape immunodominance hierarchies of antiviral CD8(+) T cells at the levels of T cell repertoire and presentation of viral antigens. J Exp Med. 2001;193:1319–1326. doi: 10.1084/jem.193.11.1319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Shin EC, Seifert U, Kato T, Rice CM, Feinstone SM, Kloetzel PM, Rehermann B. Virus-induced type I IFN stimulates generation of immunoproteasomes at the site of infection. J Clin Invest. 2006;116:3006–3014. doi: 10.1172/JCI29832. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.