

Abstract
We have developed lipid–polycation–DNA (LPD) nanoparticles containing DOTAP and targeted with polyethylene glycol (PEG) tethered with anisamide (AA) to specifically deliver siRNA to H460 human lung carcinoma cells which express the sigma receptor. A novel non-glycerol based cationic lipid which contains both a guanidinium and a lysine residue as the cationic headgroup, i.e. DSGLA, downregulated pERK more efficiently in H460 cells than DOTAP. As demonstrated by using fluorescently labeled siRNA, LPD-PEG-AA prepared with DSGLA efficiently delivered siRNA to the cytoplasm of the H460 cells. Although the siRNA delivered by LPD-PEG-AA containing either DOTAP or DSGLA could effectively silence EGFR expression, a synergistic cell killing effect in promoting cellular apoptosis was only observed with DSGLA. The fluorescently labeled siRNA was efficiently delivered into the cytoplasm of H460 xenograft tumor by the LPD-PEG-AA containing either DOTAP or DSGLA 4 h after intravenous injection. Three daily injections (0.6 mg/kg) of siRNA formulated in the LPD-PEG-AA containing either DOTAP or DSGLA could effectively silence the epidermal growth factor receptor (EGFR) in the tumor, but the formulation containing DSGLA could induce more cellular apoptosis. A significant improvement in tumor growth inhibition was observed after dosing with LPD-PEG-AA containing DSGLA. Thus, DSGLA served as both a formulation component as well as a therapeutic agent which synergistically enhanced the activity of siRNA.
Keywords: Gene delivery, cationic lipid, siRNA, apoptosis, nanoparticles, sigma receptor
Introduction
Non-small cell lung cancer (NSCLC) is the most common lung cancer which is the most common cancer and the major killer throughout the world according to the World Health Organization (WHO) cancer report.1 Because of their propensity to metastasize early and develop resistance to a wide range of anticancer drugs, the prognosis of lung cancer patients has had limited improvement and innovative strategies that effectively treat lung cancer are urgently needed.
Recently, new approaches such as immunotherapy, targeted agents and gene therapy are developed for cancer therapy. The success of gene therapy critically depends on the safety and efficacy of the transfection vector used in delivering the therapeutic gene.2,3 Small interfering RNA (siRNA) that can induce sequence-specific gene silencing has been developed as a potential cancer therapy agent.4-6 Cancer growth inhibition was observed after siRNA mediated knockdown of the overexpressed oncogenes such as EGFR that are essential to NSCLC proliferation.7 Cationic liposomes as well as viral vectors have been shown to be powerful tools to deliver siRNA.8
To enhance gene transfection effect, various cationic liposomes have been synthesized to deliver plasmid DNA, antisense or siRNA to the cytoplasm or nucleus.9-13 A cationic lipid generally contains two parts: a cationic headgroup and a hydrophobic moiety such as hydrocarbon chains.14-16 One of the critical factors that influences nucleic acid delivery is the composition of the cationic headgroup.17 For example, a spacer between the headgroup of cholesterol-based gemini lipids increases the serum compatibility of the lipoplex.18,19 Spermine or spermidine containing headgroup has the ability to condense nucleic acid. Kim et al. demonstrated that a lysine headgroup can enhance gene expression efficiency and decrease cytotoxicity.20 Obata et al. reported that cationic lipids bearing lysine or arginine as a cationic headgroup showed higher gene transfection activity, more serum compatibility, and lower cytotoxicity compared with Lipofectamine2000.17 Furthermore, cationic transfection amphiphiles containing guanidinium functionality, which mimics the arginyl residues in DNA binding protein such as histones and protamine, were first reported by Vigneron et al.21 The guanidinium group remains protonated over a much wider range of pH than other basic groups due to its remarkably high pKa value. It also forms characteristic parallel zwitterionic hydrogen bonds N−H+⋯O− with phosphate ions. The guanidinium groups are also capable of forming hydrogen bonds with nucleic acid bases, thus further enhancing the capacity to deliver plasmid DNA or siRNA.
Our laboratory has developed LPD nanoparticles which are composed of cationic liposomes and polycation-condensed DNA to deliver plasmid DNA or siRNA.7,22,23 We have demonstrated that siRNA can be formulated in LPD and causes gene silencing activity in the treated cells. The LPD formulation contained a commercially available cationic lipid DOTAP. However, DOTAP activates ERK in the dendritic cells.24 ERK is a member of the mitogen activated protein kinase (MAPK) family, activation of which could lead to an antiapoptosis effect.
In order to combat this potential problem in cancer therapy and improve siRNA delivery efficacy, we developed a novel non-glycerol based cationic lipid DSGLA which contains both guanidinium and lysine residues as a cationic headgroup. We substituted the DOTAP with DSGLA to form LPD nanoparticles for siRNA delivery in vitro and in vivo. Here we report the studies of formulating EGFR siRNA in LPD prepared with DSGLA. We have shown enhanced cellular uptake of siRNA, pronounced downregulation of the target gene, increased apoptosis of the tumor cells and improved antitumor activity of the novel formulation as compared with the formulation containing DOTAP.
Experimental Section
Materials
DOTAP and cholesterol were purchased from Avanti Polar Lipids, Inc. (Alabaster, AL). Protamine sulfate (fraction X from salmon) and calf thymus DNA (for hybridization, phenol–chloroform extracted and ethanol precipitated) were from Sigma-Aldrich (St. Louis, MO). The EGFR and control siRNA sequences are adopted from the previous studies.7 Synthetic 19-nt RNAs with 3′ dTdT overhangs on both sequences were purchased from Dharmacon (Lafayette, CO). The sequence of EGFR siRNA was 5′-AACACAGTGGAGCGAATTCCT-3′, and high-purity control siRNA with sequence 5′-AATTCTCCGAACGTGTCACGT-3′ was also synthesized in Dharmacon. For quantitative studies, cy3 was conjugated to 5′ sense sequence. 5′ cy3 labeled siRNA sequence was also obtained from Dharmacon. NCI-H460 human lung cancer cells were obtained from American Type Culture Collection. Cells were maintained in RPMI-1640 medium supplemented with 10% fetal bovine serum (Invitrogen, Carlsbad, CA), 100 U/mL penicillin, and 100 μg/mL streptomycin (Invitrogen, Carlsbad, CA).
Experimental Animals
Female athymic nude mice of age 6–8 weeks were purchased from Charles River Laboratories (Wilmington, MA). All work performed on animals was in accordance with and permitted by the University of North Carolina Institutional Animal Care and Use committee.
Synthesis of DSGLA
Synthesis of DSGLA (N,N-distearyl-N-methyl-N-2[N′-(N2-guanidino-l-lysinyl)] aminoethyl ammonium chloride) achieved in 5 steps reacting the precursor mixed primary–tertiary amine (N-aminoethyl-N,N-di-n-octadecylamine) with Nα-tert-butyloxycarbonyl-Nz-benzyloxycarbonyl-l-lysine by conventional dicyclohexylcarbodiimide (DCC) coupling. Deportation of CBZ group using Pd/H2 followed by guanidinylation using bis-BOC thiourea generated the intermediate N-2-[N′-(Nε-BOC, Nα-(dibenzyloxycarbonyl guanidino-l-lysinyl)] aminoethyl-N,N-di-n-octadecylamine. Quaternizing the guanidinylated intermediate with excess methyl iodide followed by acid deprotection generated quaternized intermediate. The resulting quaternized intermediate was passed through chloride ion-exchange chromatography and yielded the final product DSGLA. We characterized the compound with 1H NMR spectra and LSIMS. Detailed synthetic procedures, spectral and purity data are delineated in another manuscript (Bathula et al., unpublished).
Preparation of Liposomes
A cationic lipid and cholesterol in 1:1 mol ratio were dissolved in a mixture of chloroform in a glass vial. The solvent was removed with a thin flow of moisture-free nitrogen gas, and the dried lipid film was then kept under high vacuum for 8 h. An amount of 5 mL of sterile deionized water was added to the vacuum-dried lipid film, and the mixture was allowed to swell overnight. The vial was then vortexed for 2–3 min at room temperature and sonicated in a bath type sonicator for 5 min followed by extrusion (Hamilton Co., Reno, NV) through 400, 200, and 100 nm membrane filters, followed by storage at 4 °C before use. The resulting clear aqueous liposomes were used in forming LPD.
Preparation of PEGylated LPD Formulations
LPD were prepared as previously described with slight modifications.25 Briefly, small unilamellar liposomes consisting of DOTAP (or DSGLA) and cholesterol (1:1 molar ratio) were prepared by thin film hydration followed by membrane extrusion. The total lipid concentration of the liposome was fixed at 10 mM. LPD was composed of DOTAP (or DSGLA)/cholesterol liposome, protamine, and the mixture of siRNA and calf thymus DNA (1:1 weight ratio). To prepare LPD, 6 μL of protamine (2 mg/mL), 47 μL of deionized water, and 8 μL of a mixture of siRNA and calf thymus DNA (2 mg/mL) were mixed in a 1.5 mL tube. The complex was allowed to stand at room temperature for 10 min before the addition of 40 μL of DOTAP (or DSGLA)/ cholesterol liposome (total lipid concentration = 10 mM). LPD nanoparticles were kept at room temperature for another 10 min before further application. PEGylated LPD formulations were prepared by the postinsertion method.26,27 Briefly, 100 μL of preformed LPD was mixed with 0.63–16 μL of DSPE-PEG or DSPE-PEG-AA (20 mg/mL) and then incubated at 50–60 °C for 10 min. The resulting formulations were allowed to cool to room temperature before use. The particle size of LPD and PEGylated LPD was measured by using a Coulter N4 Plus particle sizer (Beckman Coulter, San Francisco, CA). Particle sizes were reported as the mean ± standard deviation.
Cellular Uptake and Quantification Study
H460 cells (1 × 105 per well) were seeded in 12-well plates (Corning Inc., Corning, NY) 12 h before experiments. Cells were treated with different formulations at a concentration of 100 nM for cy3 labeled siRNA in serum containing medium at 37 °C for 4 h. Cells were washed twice with PBS. Cells were fixed with 3.8% paraformaldehyde in PBS at room temperature for 10 min, mounted onto a glass slide, and imaged by a Leica SP2 confocal microscope. Cy3 positive cells were detected and quantified by flow cytometry (Becton-Dickinson, Heidelberg, Germany). Results were processed using the Cellquest software (Becton-Dickinson).
Assessment of Apoptosis by TUNEL Staining
TUNEL assay was conducted using a TACS TdT Kit (R&D Systems, Minneapolis, MN). H460 cells (5 × 104 per well) were seeded into 24-well plates. Cells were treated with different formulations at a concentration of 500 nM for siRNA in serum containing medium at 37 °C for 72 h. Cells were washed once with PBS, and then fixed in 4% buffered paraformaldehyde–PBS (pH 7.4) for 30 min at room temperature. Endogenous peroxidase was inactivated with 0.3% H2O2 methanol for 15 min at room temperature. The plates were then rinsed with PBS, and after processing with permeabilization buffer, labeling buffer containing terminal deoxynucleotidyl transferase and fluorescein isothiocyanate-deoxyuridine 5-triphosphate was added to the plate. The plate was incubated in a humid atmosphere at 37 °C for 60 min. The reaction was terminated by stop solution and developed with DAB according to manufacturer’s instructions. Samples were imaged using a Nikon Microphot SA microscope. The number of apoptosis cells within the rectangular area of 300 cells was counted on three or four areas for each treatment.
Western Blot Analysis
Cells were lysed in lysis buffer for 20 min on ice, and the soluble extract was recovered by centrifugation. Extracts were separated on a 10% acrylamide gel and transferred to a PVDF membrane. Membranes were blocked for 1 h in 5% skim milk and then incubated for 1 h with monoclonal antibodies directed against pERK (Santa Cruz Biotechnology, Inc.) or polyclonal antibodies against EGFR (BD Transduction Laboratories), ERK 2 and Actin (Santa Cruz Biotechnology, Inc.) for standardization. Membranes were washed in PBST (PBS, 0.1% Tween-20) and then incubated for 1 h with appropriate secondary antibodies. Membranes were again washed and then developed by an enhanced chemiluminescence system according to the manufacturer’s instructions (PerkinElmer).
For in vitro p-ERK inhibition study, H460 cells were seeded in 12-well plates (1 × 105 per well) for 24 h. Cells were treated with different lipids at the concentration of 10 μM and were collected after 1 h, 24 h and 48 h for measuring p-ERK expression. For in vitro EGFR gene silencing study, H460 cells were seeded in 6-well plates (2 × 105 per well) for 24 h. Cells were treated with siRNA-containing different formulations (250 nM siRNA) and were collected after 72 h for measuring EGFR protein expression. For in vivo EGFR gene silencing and pERK inhibition study, tumor-bearing mice were given iv injections of siRNA with different formulations at the dose of 0.6 mg siRNA/kg. One day after the third injection, the mice were killed and the tumors were collected for Western blot analysis.
Immunofluorescence Microscopy
H460 cells were washed, fixed with methanol/acetone (1:1), and permeabilized with triton X100 (1%). Cells were incubated with rabbit polyclonal anti-apoptosis-inducing-factor (AIF) (Santa Cruz Biotechnology, Inc.) (1:100) for 1 h. After washed with PBS, the fluorescently labeled secondary antibody was added and incubated for 1 h. Nuclei were counterstained with Vectashield mounting solution (Vector Laboratories, Inc., Burlingame, CA) containing DAPI.
Tissue Distribution and siRNA Uptake
Mice with tumor size of ~1 cm2 were intravenously injected with cy3 labeled siRNA in different formulations (1.2 mg/kg or 1.8 nmol of siRNA per injection). Four hours later, mice were killed and tissues were collected, fixed in 10% formalin and embedded in paraffin. Tissues were sectioned (7 μm thick) and imaged using a Leica SP2 confocal microscope.
Tumor Growth Inhibition Study
H460 xenograft tumor-bearing mice (size 9–16 mm2) were intravenously injected with siRNA-containing formulations at the dose of 0.6 mg/ kg (one injection per day for 3 days). Tumor growth in the treated mice was monitored after treatment.
Statistical Analysis
All statistical analyses were performed by Student’s t test. Data were considered statistically significant when the p value was less than 0.05.
Results
Preparation and Characterization of the Nanoparticle Containing the Novel Cationic Lipid
We have developed a LPD nanoparticle formulation which is targeted with AA and contains the cationic lipid DOTAP to specifically deliver siRNA to H460 human lung carcinoma cells which express the sigma receptor. Our novel non-glycerol based cationic lipid (DSGLA) (Figure 1A) could be readily used to formulate siRNA in LPD-PEG or LPD-PEG-AA nanoparticles, similar to the original DOTAP lipid. The particle size of the nanoparticles is around 100 nm, and the zeta potential is 25 mV. To further characterize the new formulation, we used 10 mol % NBD-cholesterol labeled liposomes or 10 mol % DSPE-PEG2000-carboxyfluorescein (DSPE-PEG2000-CF) labeled DSPE-PEG2000 to make PEGylated LPD and the final formulation was separated by using a Sepharose CL 2B column. As shown in Figure S1 in the Supporting Information, two major particle populations were observed in the DSPE-PEG2000-CF or NBD-cholesterol labeled LPD. Figure S1 in the Supporting Information indicates that the nanoparticles containing DOTAP eluted in the first peak were composed of 47.3% of total lipids and 23.4% of the input DSPE-PEG2000. The nanoparticles containing DSGLA eluted in the first peak were composed of 40.6% of total lipids and 21.2% of the input DSPE-PEG2000. Based on our calculation, 11.4 mol % of the outer leaflet of the lipid bilayer containing DSGLA was modified with DSPE-PEG2000 and 10.8 mol % of the one containing DSGLA was modified with DSPE-PEG2000. The ratio of PEGylation to LPD containing DOTAP was similar to those containing DSGLA. The particle size and zeta potential of the PEGylated LPD containing either DSGLA or DOTAP collected from the first peak was around 160 nm and 3 mV.
Figure 1.

The structure of DSGLA and pERK inhibition induced by lipids. (A) The structure of DSGLA. (B) pERK and ERK expression in H460 cells after incubation with 10 μM DSGLA and DOTAP for various times.
Inhibition of ERK1/2 Activation by Novel Lipid Treatment
We first studied the activation of ERK in the H460 cells. The phosphorylation of ERK is often associated with the antiapoptosis phenotype.28,29 H460 cells were treated with DOTAP or DSGLA liposomes for different time periods, and ERK1/2 phosphorylation which leads to the activation of the MAP kinase activity was measured by Western blot analysis. As shown in Figure 1B, 10 μM DSGLA could decrease ERK1/2 activation. Total ERK1/2 expression remained unperturbed under all conditions. The lipids without guanidine group did not cause the inhibition of ERK1/2 activation (Figure S2 in the Supporting Information). However, DOTAP increased ERK1/2 activation, an effect which is consistent with our previous findings.24 Besides, we also found that DSGLA could more efficiently induce reactive oxygen species (ROS) in H460 cells than DOTAP (Figure S3 in the Supporting Information). Interestingly, there was no statistically significant difference between the cytotoxicities of DSGLA and DOTAP based on MTT assays and flow cytometry with propidium iodide (PI) staining (Bathula et al., unpublished). DSGLA alone is as safe as DOTAP. These observations suggest that inhibition of the ERK pathway by DSGLA may not induce cell death but can present a synergistic pro-apoptosis effect when combined with other treatment.
Intracellular Uptake of siRNA in Vitro
To achieve targeted delivery of siRNA in cancer gene therapy, we postinserted PEGylated lipids onto our LPD formulation to increase the serum stability.23 In addition, we also tethered anisamide, a compound specifically binding to the sigma receptor, to the distal end of PEG as a targeting ligand.30 As shown in Figure 2A, confocal microscopy showed that the uptake to the cellular cytoplasm of H460 cells, which express sigma receptor,7 of the fluorescently labeled siRNA formulated with LPD prepared with DSGLA was much greater than that prepared with DOTAP. Furthermore, for both lipids, the fluorescence signal in the cells treated with LPD-PEG-AA was much stronger than that of cells treated with LPD-PEG. Quantitatively (Figure 2B), the fluorescently labeled siRNA uptake by LPD-PEG-AA containing DSGLA was about 2-fold higher than that of LPD-PEG-AA containing DOTAP as measured by flow cytometry. Figure 2B also showed that ligand conjugation increased the delivery efficiency of PEGylated LPD prepared with DSGLA by 3-fold. Thus, the results indicate that the LPD-PEG-AA prepared with DSGLA could efficiently deliver siRNA to the tumor cells and the delivery was highly ligand dependent. The targeted LPD (AA+) did not show different siRNA uptake compared to nontargeted LPD (AA−) in the sigma receptor negative cell line CT26 (Figure S4 in the Supporting Information), suggesting that the cellular uptake of siRNA is related to sigma receptor expression.
Figure 2.
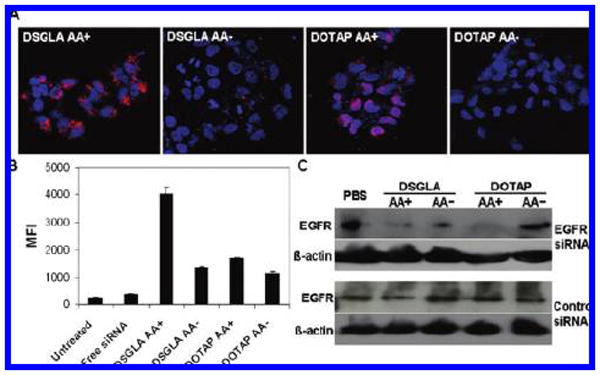
Intracellular uptake of siRNA and EGFR expression inhibited by siRNA formulation in H460 cells in vitro. Fluorescence photographs of cultured H460 cells after treatment with 5′-cy3 labeled siRNA against an irrelevant target in LPD-PEG or LPD-PEG-AA with DSGLA or DOTAP as the cationic lipid for 4 h (A). Quantitative measurement of mean fluorescence intensity (MFI) of cy3 siRNA uptake by flow cytometry. Data = mean ± SD, n = 3 (B). (C) Western blot analysis of EGFR and β-Actin in H460 cells treated with LPD-PEG with formulations containing anisamide ligand (AA+) or without (AA−). Formulations were prepared with either DOTAP or DSGLA.
Inhibition of EGFR Expression in H460 Cells
To further demonstrate the biological activity of the nanoparticle formulation, siRNA against EGFR was delivered by LPD formulations containing either DSGLA or DOTAP. The siRNA silencing effect on EGFR levels was determined by Western blot analysis. We compared different formulations in the presence or the absence of AA targeting ligand. Free anti-EGFR siRNA had little effect due to the poor cellular uptake of this negatively charged oligonucleotide (data not shown). Cultured H460 cells were treated with anti-EGFR siRNA-containing LPD-PEG-AA prepared with DSGLA or DOTAP, and EGFR protein expression was measured after 72 h (Figure 2C). LPD-PEG-AA formulations with either DSGLA or DOTAP knocked down EGFR expression with H460 cells in approximately equal efficiency. However, anti-EGFR siRNA in LPD-PEG (AA−) prepared with either DSGLA or DOTAP could only slightly downregulate EGFR (Figure 2C). Control siRNA did not show any silencing activity with any formulations. The data indicates that the siRNA could effectively suppress EGFR expression and the silencing activity was formulation dependent.
Synergistic Apoptosis Induction in Vitro
The effect of the combination of anti-EGFR siRNA and DSGLA on cancer cell killing effect was further studied. To determine whether depletion of EGFR could promote tumor cell death, TUNEL assays were performed at 72 h after treatment with either anti-EGFR or control siRNA formulations. Figure 3A indicates that about 15 ± 3% of H460 cells treated with EGFR siRNA-containing LPD-PEG-AA prepared with DS-GLA underwent apoptosis. This value was higher than the ones treated with EGFR siRNA-containing LPD-PEG prepared with DSGLA, EGFR siRNA-containing LPD-PEG-AA prepared with DOTAP, or control siRNA-containing LPD-PEG-AA prepared with DSGLA (Figure 3A). It was also observed that about 4 ± 1% of H460 cells treated with EGFR siRNA-containing LPD-PEG-AA prepared with DOTAP underwent apoptosis, compared to less than 1% in the control siRNA and EGFR siRNA-containing LPD-PEG (Figure 3A). In addition, about 2.5% of H460 cells treated with control siRNA-containing LPD-PEG-AA prepared with DSGLA underwent apoptosis as opposed to less than 1% in the control siRNA-containing LPD-PEG (Figure 3A). Thus, the data indicate that killing effect mediated by LPD nanoparticles was siRNA sequence specific, targeting ligands specific and formulation lipid dependent. Thus, a synergistic effect between siRNA against EGFR and DSGLA, but not DOTAP, in promoting cellular apoptosis was observed and the synergy was well controlled by AA.
Figure 3.
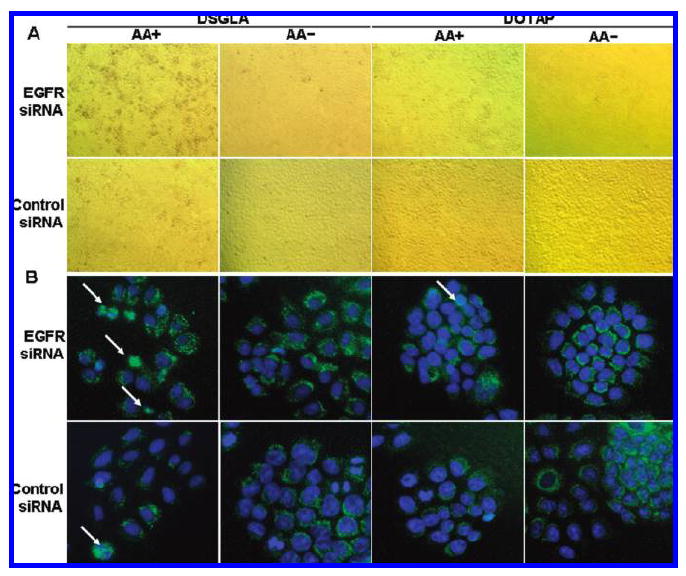
Apoptosis induced by siRNA formulation in vitro. Cells treated with different formulations for 72 h and analyzed for TUNEL staining (A), or AIF distribution (B). Cells with nuclear AIF are shown by arrows. Formulations contained anisamide ligand (AA+) or without (AA−). Formulations were prepared with either DOTAP or DSGLA.
Redistribution of cytochrome c and AIF is an early event in the cellular apoptotic process.31,32 To further evaluate the enhancement of H460 lung cancer cell death by the combination of EGFR siRNA and DSGLA, we examined the involvement of AIF by immunofluorescence microscopy (Figure 3B). Immunofluorescence detection of AIF in untreated control cells normally yields a punctate cytoplasmic staining pattern with some preference for the perinuclear area as a typical pattern for mitochondrial localization.33-35 Cells treated with EGFR siRNA-containing LPD-PEG-AA prepared with DSGLA showed an increased translocation of AIF from the cytoplasm into the nucleus (Figure 3B). No significant translocation was observed in other treatment groups. The results indicate that combined treatment with EGFR siRNA formulated with DSGLA interacted synergistically to promote cell death in H460, and that the synergistic effect was controlled by the targeting ligands.
To confirm that the selective synergistic cellular killing effect of the targeted nanoparticles, H460 cells were stained with annexin V-FITC and PI and analyzed by flow cytometry for apoptosis. As shown in Figure S4 in the Supporting Information, a ligand and lipid-dependent induction of apoptosis was observed. It indicates that about 25% of H460 cells treated with EGFR siRNA-containing LPD-PEG-AA prepared with DSGLA underwent apoptosis. The sigma receptor negative CT26 cells were also assessed by an annexin V-FITC and PI binding assay. As shown in Figure S4 in the Supporting Information, the treatment of EGFR siRNA-containing LPD-PEG-AA prepared with DSGLA induces a low level of apoptosis (6.3%) in CT26 cells compared with the similar rate of apoptosis induced by other formulations. It indicates that the synergistic cellular killing effect of DSGLA AA+ containing EGFR siRNA is sigma receptor specific.
Tissue Distribution and Intracellular Uptake of siRNA
We further studied the cy3-siRNA distribution and bioavailability in major tissues in the H460 xenograft model 4 h after intravenous (iv) injections using confocal microscopy. As shown in Figure 4, the intracellular fluorescence signals were hardly detected in the tumor tissues collected from the mice treated with LPD-PEG prepared with DSGLA and DOTAP. The LPD-PEG-AA prepared with DSGLA or DOTAP showed strong cytosolic delivery of cy3 siRNA in the tumor tissue, while other tissues showed lower uptake of siRNA. The distribution of cy3 siRNA in the tumor was heterogeneous. These results indicate that the LPD-PEG-AA prepared with the novel DSGLA can efficiently deliver siRNA to the tumor tissue and that the intracellular delivery is highly ligand dependent.
Figure 4.
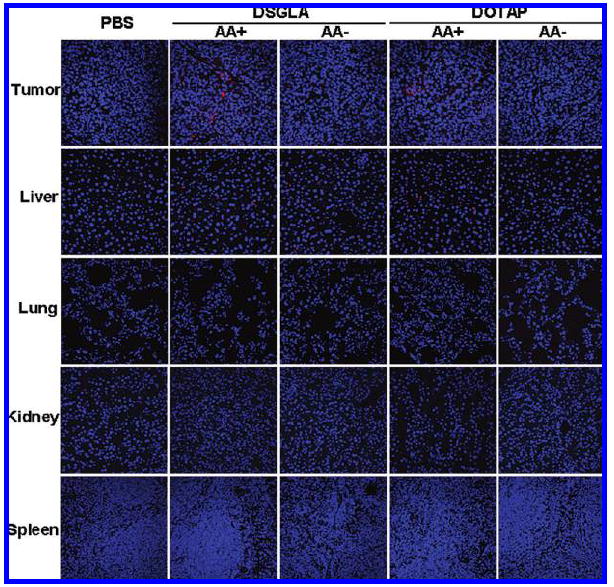
Tissue distribution and intracellular uptake of siRNA in different formulations. Fluorescence signal of cy3 labeled siRNA in different tissues observed by confocal microscopy.
EGFR Gene Silencing, Inhibition of ERK1/2 Activation and Apoptosis Induction
To examine the biological activities of siRNA in vivo, EGFR levels in the xenograft tumor were detected by Western blotting (Figure 5A). EGFR in H460 tumor was silenced by EGFR siRNA in LPD-PEG-AA prepared with DSGLA and DOTAP. The EGFR siRNA-containing LPD-PEG prepared with DSGLA and DOTAP showed only a partial effect, whereas the control siRNA showed no effect. The inhibition of ERK1/2 activation was only observed with LPD-PEG-AA prepared using DSGLA (Figure 5B). We also stained for apoptotic markers in the H460 tumor (Figure 6). Figure 6C indicates that about 8% of H460 cells treated with EGFR siRNA-containing LPD-PEG-AA prepared with DSGLA underwent apoptosis detected by TUNEL staining. This value was higher than that of tumors treated with EGFR siRNA-containing LPD-PEG prepared with DSGLA, EGFR siRNA-containing LPD-PEG-AA prepared with DOTAP, or control siRNA-containing LPD-PEG-AA prepared with DSGLA (Figures 6A and 6C). To further evaluate the enhancement of killing effect in the H460 tumor by the combination of EGFR siRNA and DSGLA, we examined the involvement of AIF in cellular apoptosis (Figure 6B). Cells treated with EGFR siRNA-containing LPD-PEG-AA prepared with DSGLA showed an increased translocation of AIF from the cytoplasm into the nucleus (Figures 6B and 6C). No significant translocation was observed in other treatment groups. The results indicate that combined treatment with EGFR siRNA formulated with DSGLA interacted synergistically to promote cell death in the H460 tumor and that the synergy effect was ligand dependent.
Figure 5.
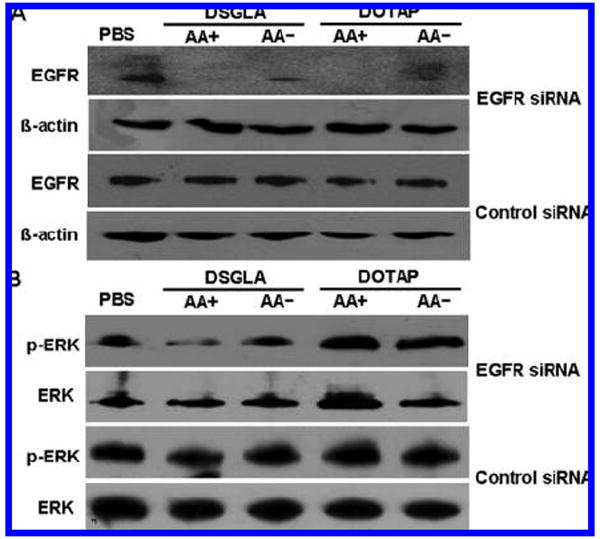
EGFR and p-ERK expression in H460 xenograft tumor. Western blot analysis of EGFR (A) and p-ERK (B) in the H460 xenograft tumor after treatment with different formulations.
Figure 6.
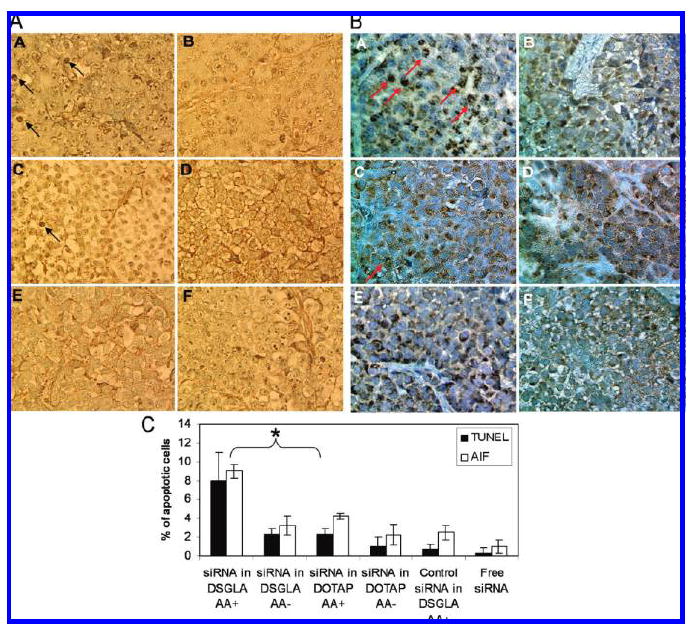
Synergistic apoptosis induction in H460 xenograft tumor. TUNEL staining (positive cells indicated by dark arrows) (A) and AIF expression and localization (cells with nuclear AIF indicated by red arrows) (B) in H460 tumor cells after treatment with EGFR siRNA with different formulations in vivo. (C) Quantitative analysis of TUNEL positive staining and nuclear translocation of AIF in the tumors treated with different formulations. a, EGFR siRNA in DSGLA AA+; b, EGFR siRNA in DSGLA AA−; c, EGFR siRNA in DOTAP AA+; d, EGFR siRNA in DOTAP AA−; e, control siRNA in DSGLA AA+; f, free siRNA. Data = mean ± SD, n = 3–4. * indicates P < 0.05.
Tumor Growth Inhibition
Three injections of EGFR siRNA in LPD-PEG-AA containing DOTAP showed a partial inhibition of tumor growth (P < 0.01 at day 11) similar to that of siRNA in LPD-PEG containing DSGLA (at day 8) (Figure 7). A significant improvement in tumor growth inhibition was observed when treated with combination of siRNA and DSGLA (LPD-PEG-AA containing DSGLA) (P < 0.001 at day 6). Other control treatments had no or much lower therapeutic effects.
Figure 7.
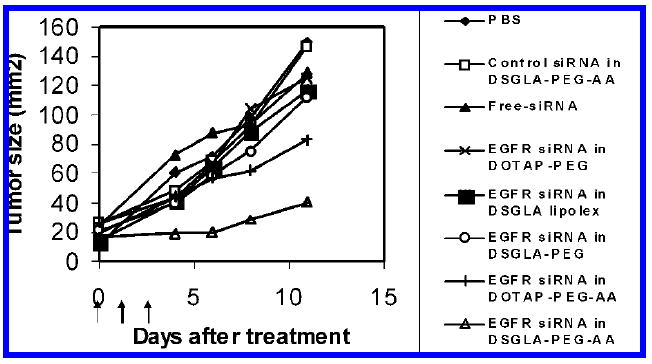
H460 xenograft tumor growth inhibition by siRNA in different formulations. Solid arrows indicate the intravenous administrations of siRNA (0.6 mg/kg). Data = mean, n = 5–7. SD of the data points is not shown for clarity.
Discussion
In this study, our objective was to develop a novel cationic lipid that could avoid the possible antiapoptotic effect of DOTAP, yet could still deliver siRNA with high efficiency. We have synthesized a lysine based cationic lipid containing a guanidine group and tested its ability to form LPD. Our studies demonstrate that siRNA formulated in LPD prepared with DSGLA showed enhanced cellular uptake, gene silencing activity and synergistic therapeutic activity with EGFR siRNA in H460 tumor cells in vitro and in vivo. This synergistic therapeutic effect is AA ligand dependent which is targeted to the sigma receptor overexpressed in many human cancer cells.36,37
The strategy that we used to achieve tumor targeted delivery is based on the enhanced permeability and retention (EPR) effect.38,39 Although the normal cells also express the sigma receptor,40 they are not accessible by the blood borne nanoparticles. Sigma receptor-targeting ligands such as AA play the major role to increase the intracellular uptake of the nanoparticles but do not enhance tumor localization.41
Obata et al. reported that cationic lipids bearing lysine or arginine as a cationic headgroup showed higher plasmid transfection efficacy and are more serum compatible than Lipofectamine2000.17 From our studies, we have demonstrated that DSGLA containing LPD showed higher cellular uptake of siRNA in H460 cells in vitro than DOTAP, although the enhanced siRNA uptake was not observed in vivo. We suggest that the enhanced transfection effect may be related to the guanidine containing headgroup, and this hypothesis will be further evaluated. DSGLA also showed a biological activity to downregulate pERK (Figure 1B), but the lipids without guanidine group did not cause this effect (Figure S2 in the Supporting Information). Clearly, the guanidine group plays an important role in determining the biological characteristics of the lipids.
We have found that DSGLA promoted apoptosis induced by EGFR siRNA synergistically. The mechanisms for promoting apoptosis of H460 cells induced by DSGLA could be complicated. A guanidine group, which easily donates an electron, may generate a superoxide forming a hydroxyl radical.42 We also found that DSGLA can more efficiently induce reactive oxygen species (ROS) in H460 cells than DOTAP (Figure S3 in the Supporting Information).43 It may involve different pathways in cancer cell killing. For example, in this study, inhibition of p-ERK, essential for cell survival, could be just one of the mechanisms.44 ROS mediates apoptosis in many different cell types. It plays an important role as a second messenger in many signaling pathways such as Akt, TNF and MAPK pathways.45 It also regulates the expression or stability of pro- or antiapoptosis protein such as Bcl-2, the key enzyme suppressing apoptosis.46 ROS may also cause membrane and DNA damage, which enhances poly(ADP-ribose) polymerase-1 (PARP-1) activation and triggers AIF translocation from mitochondria to the nucleus.47,48 The extent of the ROS regulation of signaling pathways is not fully understood. We will study further to understand the network of signaling pathways influenced by DSGLA. The information may provide new therapeutic targets for cancer therapy.
Cancer cells develop multiple mechanisms to ensure proliferation, metastasis and survival.49,50 Inhibiting a resistance or survival pathway is often an effective way to enhance the toxicity of a chemotherapeutic drug.51,52 It is often necessary to target multiple pathways to efficiently reduce the growth of cancer cells.53 For this purpose, we studied the effect of combined treatment of human lung cancer cells with siRNA targeting EGFR and DSGLA.
Our data (Figures 3 and 6) indicate that DSGLA significantly enhanced the killing effect of EGFR silencing in a formulation dependent manner both in vitro and in vivo, although the lipid itself is not cytotoxic (Bathula et al., unpublished). The synergistic pERK inactivation of EGFR siRNA delivered by LPD-PEG-AA containing DSGLA was demonstrated in a xenograft model of H460 cells (Figure 5B). However, DSGLA without the cooperation of EGFR siRNA (control siRNA delivered by LPD-PEG-AA containing DSGLA) or EGFR siRNA without the cooperation of DSGLA (EGFR siRNA delivered by LPD-PEG-AA containing DOTAP) did not show pERK inactivation (Figure 5B). EGFR promotes cell proliferation by both the Raf/MEK/ERK and the PI3K/PDK1/Akt pathways.54 Thus, inhibition of pathways other than Raf/MEK/ERK, which is already effectively inhibited by DSGLA, by EGFR siRNA should bring enhanced apoptosis in cells treated with both agents (Figure 6). The synergistic therapeutic effect of LPD-PEG-AA containing DSGLA was also demonstrated in a xenograft model of H460 cells (Figure 7).
In conclusion, we have described a target specific nanoparticle formulation that contains both siRNA and DSGLA, a new cationic lipid, and plays both roles of a delivery component and a therapeutic agent. As far as we know, this is the first demonstration of such an approach and it may serve as a safe and effective anticancer drug.
Supplementary Material
Acknowledgments
CT26 cell line was generously supplied by Dr. Moo J. Cho. We thank Michael Hooker Microscopy Facility at UNC for the microscopy images. This research was supported by NIH Grant CA129835 and by the University of North Carolina at Chapel Hill. Shyh-Dar Li is appreciated for the assistance in preparing the manuscript.
Footnotes
Supporting Information Available: Figures S1–S4 and additional experimental details. This material is available free of charge via the Internet at http://pubs.acs.org.
References
- 1.Payne S. The WHO Gender and Health Research Series. WHO; Geneva: 2005. Gender in lung cancer and smoking research; pp. 1–43. [Google Scholar]
- 2.Christopher ME, Wong JP. Recent developments in delivery of nucleic acid-based antiviral agents. Curr Pharm Des. 2006;12(16):1995–2006. doi: 10.2174/138161206777442146. [DOI] [PubMed] [Google Scholar]
- 3.Kawashita Y, Fujioka H, Ohtsuru A, Kaneda Y, Kamohara Y, Kawazoe Y, Yamashita S, Kanematsu T. The efficacy and safety of gene transfer into the porcine liver in vivo by HVJ (Sendai virus) liposome. Transplantation. 2005;80(11):1623–9. doi: 10.1097/01.tp.0000184447.88283.f3. [DOI] [PubMed] [Google Scholar]
- 4.Hou JQ, He J, Wang XL, Wen DG, Chen ZX. Effect of small interfering RNA targeting survivin gene on biological behaviour of bladder cancer. Chin Med J (Engl) 2006;119(20):1734–9. [PubMed] [Google Scholar]
- 5.Ohri SS, Vashishta A, Proctor M, Fusek M, Vetvicka V. Depletion of Procathepsin D Gene Expression by RNA Interference: A Potential Therapeutic Target for Breast Cancer. Cancer Biol Ther. 2007;6(7) doi: 10.4161/cbt.6.7.4325. [DOI] [PubMed] [Google Scholar]
- 6.Qian H, Yu J, Li Y, Wang H, Song C, Zhang X, Liang X, Fu M, Lin C. RNA interference against metastasis-associated gene 1 inhibited metastasis of B16F10 melanoma cell in a C57BL/6 model. Biol Cell. 2007;99:573–81. doi: 10.1042/bc20060130. [DOI] [PubMed] [Google Scholar]
- 7.Li SD, Chen YC, Hackett MJ, Huang L. Tumor-targeted delivery of siRNA by self-assembled nanoparticles. Mol Ther. 2008;16(1):163–9. doi: 10.1038/sj.mt.6300323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tong AW. Small RNAs and non-small cell lung cancer. Curr Mol Med. 2006;6(3):339–49. doi: 10.2174/156652406776894554. [DOI] [PubMed] [Google Scholar]
- 9.Nakanishi M, Noguchi A. Confocal and probe microscopy to study gene transfection mediated by cationic liposomes with a cationic cholesterol derivative. Adv Drug Delivery Rev. 2001;52(3):197–207. doi: 10.1016/s0169-409x(01)00207-1. [DOI] [PubMed] [Google Scholar]
- 10.Lechardeur D, Lukacs GL. Intracellular barriers to non-viral gene transfer. Curr Gene Ther. 2002;2(2):183–94. doi: 10.2174/1566523024605609. [DOI] [PubMed] [Google Scholar]
- 11.Zuhorn IS, Hoekstra D. On the mechanism of cationic amphiphile-mediated transfection. To fuse or not to fuse: is that the question? J Membr Biol. 2002;189(3):167–79. doi: 10.1007/s00232-002-1015-7. [DOI] [PubMed] [Google Scholar]
- 12.Geusens B, Lambert J, De Smedt SC, Buyens K, Sanders NN, Van Gele M. Ultradeformable cationic liposomes for delivery of small interfering RNA (siRNA) into human primary melanocytes. J Controlled Release. 2009;133(3):214–20. doi: 10.1016/j.jconrel.2008.10.003. [DOI] [PubMed] [Google Scholar]
- 13.Han SE, Kang H, Shim GY, Suh MS, Kim SJ, Kim JS, Oh YK. Novel cationic cholesterol derivative-based liposomes for serum-enhanced delivery of siRNA. Int J Pharm. 2008;353(1–2):260–9. doi: 10.1016/j.ijpharm.2007.11.026. [DOI] [PubMed] [Google Scholar]
- 14.Gaucheron J, Santaella C, Vierling P. Highly fluorinated lipospermines for gene transfer: synthesis and evaluation of their in vitro transfection efficiency. Bioconjugate Chem. 2001;12(1):114–28. doi: 10.1021/bc000089y. [DOI] [PubMed] [Google Scholar]
- 15.Sen J, Chaudhuri A. Design, syntheses, and transfection biology of novel non-cholesterol-based guanidinylated cationic lipids. J Med Chem. 2005;48(3):812–20. doi: 10.1021/jm049417w. [DOI] [PubMed] [Google Scholar]
- 16.Bombelli C, Bordi F, Ferro S, Giansanti L, Jori G, Mancini G, Mazzuca C, Monti D, Ricchelli F, Sennato S, Venanzi M. New Cationic Liposomes as Vehicles of m-Tetrahydroxyphenylchlorin in Photodynamic Therapy of Infectious Diseases. Mol Pharmaceutics. 2008;5(4):672–9. doi: 10.1021/mp800037d. [DOI] [PubMed] [Google Scholar]
- 17.Obata Y, Suzuki D, Takeoka S. Evaluation of cationic assemblies constructed with amino acid based lipids for plasmid DNA delivery. Bioconjugate Chem. 2008;19(5):1055–63. doi: 10.1021/bc700416u. [DOI] [PubMed] [Google Scholar]
- 18.Bajaj A, Kondaiah P, Bhattacharya S. Gene transfection efficacies of novel cationic gemini lipids possessing aromatic backbone and oxyethylene spacers. Biomacromolecules. 2008;9(3):991–9. doi: 10.1021/bm700930y. [DOI] [PubMed] [Google Scholar]
- 19.Bajaj A, Kondaiah P, Bhattacharya S. Synthesis and gene transfer activities of novel serum compatible cholesterol-based gemini lipids possessing oxyethylene-type spacers. Bioconjugate Chem. 2007;18(5):1537–46. doi: 10.1021/bc070010q. [DOI] [PubMed] [Google Scholar]
- 20.Kim HS, Moon J, Kim KS, Choi MM, Lee JE, Heo Y, Cho DH, Jang DO, Park YS. Gene-transferring efficiencies of novel diamino cationic lipids with varied hydrocarbon chains. Bioconjugate Chem. 2004;15(5):1095–101. doi: 10.1021/bc049934t. [DOI] [PubMed] [Google Scholar]
- 21.Vigneron JP, Oudrhiri N, Fauquet M, Vergely L, Bradley JC, Basseville M, Lehn P, Lehn JM. Guanidinium-cholesterol cationic lipids: efficient vectors for the transfection of eukaryotic cells. Proc Natl Acad Sci U S A. 1996;93(18):9682–6. doi: 10.1073/pnas.93.18.9682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Li S, Rizzo MA, Bhattacharya S, Huang L. Characterization of cationic lipid-protamine-DNA (LPD) complexes for intravenous gene delivery. Gene Ther. 1998;5(7):930–7. doi: 10.1038/sj.gt.3300683. [DOI] [PubMed] [Google Scholar]
- 23.Li SD, Huang L. Targeted delivery of antisense oligodeoxy-nucleotide and small interference RNA into lung cancer cells. Mol Pharmaceutics. 2006;3(5):579–88. doi: 10.1021/mp060039w. [DOI] [PubMed] [Google Scholar]
- 24.Yan W, Chen W, Huang L. Mechanism of adjuvant activity of cationic liposome: phosphorylation of a MAP kinase, ERK and induction of chemokines. Mol Immunol. 2007;44(15):3672–81. doi: 10.1016/j.molimm.2007.04.009. [DOI] [PubMed] [Google Scholar]
- 25.Cui Z, Han SJ, Vangasseri DP, Huang L. Immunostimulation mechanism of LPD nanoparticle as a vaccine carrier. Mol Pharmaceutics. 2005;2(1):22–8. doi: 10.1021/mp049907k. [DOI] [PubMed] [Google Scholar]
- 26.Ishida T, Iden DL, Allen TM. A combinatorial approach to producing sterically stabilized (Stealth) immunoliposomal drugs. FEBS Lett. 1999;460(1):129–33. doi: 10.1016/s0014-5793(99)01320-4. [DOI] [PubMed] [Google Scholar]
- 27.Perouzel E, Jorgensen MR, Keller M, Miller AD. Synthesis and formulation of neoglycolipids for the functionalization of liposomes and lipoplexes. Bioconjugate Chem. 2003;14(5):884–98. doi: 10.1021/bc034068q. [DOI] [PubMed] [Google Scholar]
- 28.Eckert A, Bock BC, Tagscherer KE, Haas TL, Grund K, Sykora J, Herold-Mende C, Ehemann V, Hollstein M, Chneiweiss H, Wiestler OD, Walczak H, Roth W. The PEA-15/PED protein protects glioblastoma cells from glucose deprivation-induced apoptosis via the ERK/MAP kinase pathway. Oncogene. 2008;27(8):1155–66. doi: 10.1038/sj.onc.1210732. [DOI] [PubMed] [Google Scholar]
- 29.McCubrey JA, Steelman LS, Chappell WH, Abrams SL, Wong EW, Chang F, Lehmann B, Terrian DM, Milella M, Tafuri A, Stivala F, Libra M, Basecke J, Evangelisti C, Martelli AM, Franklin RA. Roles of the Raf/MEK/ERK pathway in cell growth, malignant transformation and drug resistance. Biochim Biophys Acta. 2007;1773(8):1263–84. doi: 10.1016/j.bbamcr.2006.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Banerjee R, Tyagi P, Li S, Huang L. Anisamide-targeted stealth liposomes: a potent carrier for targeting doxorubicin to human prostate cancer cells. Int J Cancer. 2004;112(4):693–700. doi: 10.1002/ijc.20452. [DOI] [PubMed] [Google Scholar]
- 31.Daugas E, Susin SA, Zamzami N, Ferri KF, Irinopoulou T, Larochette N, Prevost MC, Leber B, Andrews D, Penninger J, Kroemer G. Mitochondrio-nuclear translocation of AIF in apoptosis and necrosis. FASEB J. 2000;14(5):729–39. [PubMed] [Google Scholar]
- 32.Fehlberg S, Gregel CM, Goke A, Goke R. Bisphenol A diglycidyl ether-induced apoptosis involves Bax/Bid-dependent mitochondrial release of apoptosis-inducing factor (AIF), cytochrome c and Smac/DIABLO. Br J Pharmacol. 2003;139(3):495–500. doi: 10.1038/sj.bjp.0705275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Loeffler M, Daugas E, Susin SA, Zamzami N, Metivier D, Nieminen AL, Brothers G, Penninger JM, Kroemer G. Dominant cell death induction by extramitochondrially targeted apoptosis-inducing factor. FASEB J. 2001;15(3):758–67. doi: 10.1096/fj.00-0388com. [DOI] [PubMed] [Google Scholar]
- 34.Lorenzo HK, Susin SA, Penninger J, Kroemer G. Apoptosis inducing factor (AIF): a phylogenetically old, caspase-independent effector of cell death. Cell Death Differ. 1999;6(6):516–24. doi: 10.1038/sj.cdd.4400527. [DOI] [PubMed] [Google Scholar]
- 35.Modjtahedi N, Giordanetto F, Madeo F, Kroemer G. Aposptosis-inducing factor: vital and lethal. Trends Cell Biol. 2006;16(5):264–72. doi: 10.1016/j.tcb.2006.03.008. [DOI] [PubMed] [Google Scholar]
- 36.John CS, Bowen WD, Varma VM, McAfee JG, Moody TW. Sigma receptors are expressed in human non-small cell lung carcinoma. Life Sci. 1995;56(26):2385–92. doi: 10.1016/0024-3205(95)00232-u. [DOI] [PubMed] [Google Scholar]
- 37.Vilner BJ, John CS, Bowen WD. Sigma-1 and sigma-2 receptors are expressed in a wide variety of human and rodent tumor cell lines. Cancer Res. 1995;55(2):408–13. [PubMed] [Google Scholar]
- 38.Matsumura Y, Maeda H. A new concept for macromolecular therapeutics in cancer chemotherapy: mechanism of tumoritropic accumulation of proteins and the antitumor agent smancs. Cancer Res. 1986;46(12 Part 1):6387–92. [PubMed] [Google Scholar]
- 39.Tanaka T, Shiramoto S, Miyashita M, Fujishima Y, Kaneo Y. Tumor targeting based on the effect of enhanced permeability and retention (EPR) and the mechanism of receptor-mediated endocytosis (RME) Int J Pharm. 2004;277(1–2):39–61. doi: 10.1016/j.ijpharm.2003.09.050. [DOI] [PubMed] [Google Scholar]
- 40.Klouz A, Said DB, Ferchichi H, Kourda N, Ouanes L, Lakhal M, Tillement JP, Morin D. Protection of cellular and mitochondrial functions against liver ischemia by N-benzyl-N′-(2-hydroxy-3,4-dimethoxybenzyl)-piperazine (BHDP), a sigma1 ligand. Eur J Pharmacol. 2008;578(2–3):292–9. doi: 10.1016/j.ejphar.2007.09.038. [DOI] [PubMed] [Google Scholar]
- 41.Pirollo KF, Chang EH. Does a targeting ligand influence nanoparticle tumor localization or uptake. Trends Biotechnol. 2008;26(10):552–8. doi: 10.1016/j.tibtech.2008.06.007. [DOI] [PubMed] [Google Scholar]
- 42.Hiramatsu M. A role for guanidino compounds in the brain. Mol Cell Biochem. 2003;244(1–2):57–62. [PubMed] [Google Scholar]
- 43.Yan W, Chen W, Huang L. Reactive oxygen species play a central role in the activity of cationic liposome based cancer vaccine. J Controlled Release. 2008;130(1):22–8. doi: 10.1016/j.jconrel.2008.05.005. [DOI] [PubMed] [Google Scholar]
- 44.Liu WH, Cheng YC, Chang LS. ROS-mediated p38alpha MAPK activation and ERK inactivation responsible for upregulation of Fas and FasL and autocrine Fas-mediated cell death in Taiwan cobra phospholipase A(2)-treated U937 cells. J Cell Physiol. doi: 10.1002/jcp.21713. Published online Jan 29, 2009. [DOI] [PubMed] [Google Scholar]
- 45.Giles GI. The redox regulation of thiol dependent signaling pathways in cancer. Curr Pharm Des. 2006;12(34):4427–43. doi: 10.2174/138161206779010549. [DOI] [PubMed] [Google Scholar]
- 46.Kim S, Lee TJ, Leem J, Choi KS, Park JW, Kwon TK. Sanguinarine-induced apoptosis: generation of ROS, down-regulation of Bcl-2, c-FLIP, and synergy with TRAIL. J Cell Biochem. 2008;104(3):895–907. doi: 10.1002/jcb.21672. [DOI] [PubMed] [Google Scholar]
- 47.Fleury C, Mignotte B, Vayssiere JL. Mitochondrial reactive oxygen species in cell death signaling. Biochimie. 2002;84(2–3):131–41. doi: 10.1016/s0300-9084(02)01369-x. [DOI] [PubMed] [Google Scholar]
- 48.Zorov DB, Juhaszova M, Sollott SJ. Mitochondrial ROS-induced ROS release: an update and review. Biochim Biophys Acta. 2006;1757(5–6):509–17. doi: 10.1016/j.bbabio.2006.04.029. [DOI] [PubMed] [Google Scholar]
- 49.Ivanov VN, Hei TK. Combined treatment with EGFR inhibitors and arsenite upregulated apoptosis in human EGFR-positive melanomas: a role of suppression of the PI3K-AKT pathway. Oncogene. 2005;24(4):616–26. doi: 10.1038/sj.onc.1208125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Strong TV. Gene therapy for carcinoma of the breast: Genetic immunotherapy. Breast Cancer Res. 2000;2(1):15–21. doi: 10.1186/bcr24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Olie RA, Simoes-Wust AP, Baumann B, Leech SH, Fabbro D, Stahel RA, Zangemeister-Wittke U. A novel antisense oligonucleotide targeting survivin expression induces apoptosis and sensitizes lung cancer cells to chemotherapy. Cancer Res. 2000;60(11):2805–9. [PubMed] [Google Scholar]
- 52.Pakunlu RI, Wang Y, Tsao W, Pozharov V, Cook TJ, Minko T. Enhancement of the efficacy of chemotherapy for lung cancer by simultaneous suppression of multidrug resistance and antiapoptotic cellular defense: novel multicomponent delivery system. Cancer Res. 2004;64(17):6214–24. doi: 10.1158/0008-5472.CAN-04-0001. [DOI] [PubMed] [Google Scholar]
- 53.Wu X, Deng Y, Wang G, Tao K. Combining siRNAs at two different sites in the EGFR to suppress its expression, induce apoptosis, and enhance 5-fluorouracil sensitivity of colon cancer cells. J Surg Res. 2007;138(1):56–63. doi: 10.1016/j.jss.2006.06.027. [DOI] [PubMed] [Google Scholar]
- 54.Navolanic PM, Steelman LS, McCubrey JA. EGFR family signaling and its association with breast cancer development and resistance to chemotherapy (Review) Int J Oncol. 2003;22(2):237–52. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.