

Crotoxin B is a basic phospholipase A2 found in the venom of C. durissus collilineatus and is one of the subunits which constitute the crotoxin. Here, we present the crystallization, X-ray diffraction data collection and molecular-replacement solution of the dimeric complex formed by two crotoxin B isoforms.
Keywords: crotoxin B, snake venoms, Crotalus durissus collilineatus, phospholipases A2
Abstract
Crotoxin B is a basic phospholipase A2 found in the venom of several Crotalus durissus ssp. rattlesnakes and is one of the subunits that constitute crotoxin, the main component of the venom of these snakes. This heterodimeric toxin is related to important envenomation effects such as neurological disorders, myotoxicity and renal failure. Although crotoxin was first crystallized in 1938, the first structural data only became available in 2007 (for crotoxin B from C. durissus terrificus) and showed an ambiguous result for the biological assembly, which could be either dimeric or tetrameric. In this work, the crystallization, X-ray diffraction data collection at 2.2 Å resolution and molecular-replacement solution of a dimeric complex formed by two crotoxin B isoforms from C. durissus collilineatus venom is presented.
1. Introduction
Crotoxin (Cro) is the main toxin found in the venoms of several South American Crotalus durissus ssp. rattlesnakes (Santoro et al., 1999 ▶). Cro is a β-neurotoxic phospholipase A2 (PLA2) with potent neuromuscular activity that blocks neuromuscular transmission (Vital-Brazil & Excell, 1970 ▶; Faure et al., 1993 ▶) by modifications of the neurotransmitter release at the motor nerve terminals (Hawgood & Smith, 1977 ▶). It is a heterodimeric complex composed of a noncovalent association between two subunits: an acidic nontoxic and non-enzymatic component (crotoxin A, CA or crotapotin; Aird et al., 1985 ▶, 1990 ▶) and a basic and toxic Asp49-PLA2 (crotoxin B, CB or basic Asp49-PLA2; Hendon & Fraenkel-Conrat, 1971 ▶; Aird et al., 1986 ▶). Different studies have indicated a large diversity between isoforms isolated not only from a pool containing venoms from different C. durissus ssp. specimens, but also from a unique snake specimen (Faure & Bon, 1987 ▶, 1988 ▶; Faure et al., 1991 ▶). This variety may arise from post-translational modifications occurring in a single precursor of crotoxin and/or be related to the expression of different mRNAs by individual snakes (Faure et al., 1994 ▶). Two of these isoforms (CB1 and CB2) were isolated from a cDNA library prepared from a single venom gland of C. durissus terrificus and their amino-acid compositions showed that they differed at eight positions (Faure et al., 1994 ▶). These substitutions resulted in slight modifications in the enzymatic and pharmacological properties of the toxin: the CB1 isoform has higher toxic and lower enzymatic activities compared with the CB2 isoform. This effect probably arises from the higher affinity of the CB1 isoform for the CA subunit compared with the CB2 isoform (Faure et al., 1993 ▶, 1994 ▶).
The Cro complex constitutes an example of molecular potentiation, since the CA subunit hinders the formation of nonspecific interactions between the CB subunit and its substrates or other molecules. Indeed, the CA or CB subunits have been reported to be pharmacologically inactive when they are isolated and CA is thought to act as a chaperone protein for CB, increasing its neurotoxic activity (Bon et al., 1979 ▶). Dissociation of the CA–CB complex (Cro) occurs by its interaction with specific inhibitors that are present in the pre-synaptic nerve endings and possibly in the sarcoplasma and other molecular membranes, leading to the induction of potent neurotoxic, cytotoxic and myotoxic effects by the CB subunit (Bon, 1997 ▶). Despite its harmful effects, Cro has been successfully tested as an inhibitor of tumour growth and an efficient analgesic substance (Corin et al., 1993 ▶; Rudd et al., 1994 ▶; Cura et al., 2002 ▶; Zhang et al., 2006 ▶). This fact enhances interest not only in its mechanism of action but also in the possible practical applications of Cro as a drug or even as a model for the development of biotechnological products.
Recently, crystals of CB from C. durissus terrificus were obtained and its structure was solved (Marchi-Salvador et al., 2007 ▶). These were the first structural data to be archived for Cro, CA or CB, despite the fact that Cro was the first animal toxin to be crystallized in 1938 (Slotta & Fraenkel-Conrat, 1938 ▶; Faure & Bon, 1988 ▶). The structure presents a novel tetrameric complex formed by two dimers of crotoxin B isoforms (CB1 and CB2; Marchi-Salvador et al., 2008 ▶). Additionally, the use of various techniques to study the biological oligomeric assembly was described, including dynamic light scattering, electrophoresis and bioinformatics tools. Based on these experiments, both dimeric and tetrameric conformations were found to be possible assemblies for CB (Marchi-Salvador et al., 2008 ▶).
C. durissus collilineatus is a clinically important rattlesnake species found in central Brazil, but few structural–functional studies have been performed using its venom. The absence of crotamine from its venom is particularly interesting since this toxin has myotoxic and neurotoxic activities and accounts for 20% of the dry venom weight in C. durissus terrificus. These findings suggested that the crotoxin isoforms from C. durissus collilineatus probably play a very important role in the action of the venom (Ponce-Soto et al., 2002 ▶). Recently, several studies of the isolation and characterization of crotoxin and phospholipases from C. durissus collilineatus venom have been reported (Ponce-Soto et al., 2002 ▶; Rangel-Santos et al., 2004 ▶; Toyama et al., 2005 ▶), but no structural studies are available to date.
In order to clarify the correct biological assembly of CB and to gain insights into its biological function, in the present article we describe the crystallization, X-ray diffraction data collection and molecular-replacement solution of a dimeric arrangement of two isoforms of crotoxin B (CB1 and CB2) isolated from C. durissus collilineatus venom.
2. Materials and methods
2.1. Purification and amino-acid sequence
Crotoxin B (CB) was isolated from C. durissus collilineatus snake venom by ion-exchange chromatography on a CM-Sepharose column (2 × 20 cm) previously equilibrated with 0.05 M ammonium bicarbonate buffer pH 8.0 (Soares et al., 1998 ▶). The dissociation of crotoxin and the isolation of the subunits were performed according to Hendon & Fraenkel-Conrat (1971 ▶). The amino-acid sequences of the two CB isoforms (CB1 and CB2) which form the asymmetric unit of the crystal used in the X-ray diffraction experiments were the same as those determined by Faure et al. (1994 ▶) as checked by the electron densities of the model. The homogeneity of the toxin was analyzed by polyacrylamide gel electrophoresis (PAGE) for basic proteins and SDS–PAGE according to Reisfeld et al. (1962 ▶) and Laemmli (1970 ▶), respectively.
2.2. Crystallization
A lyophilized sample of CB was dissolved in ultrapure water to a concentration of 10 mg ml−1. The sparse-matrix method (Jancarik & Kim, 1991 ▶) was used to perform initial screening of the crystallization conditions (Crystal Screens I and II, Hampton Research). Crystals of CB were obtained using the conventional hanging-drop vapour-diffusion method (McPherson, 1982 ▶), in which 1 µl protein solution and 1 µl reservoir solution were mixed and equilibrated against a reservoir (500 µl) containing 2.0 M ammonium sulfate and 0.1 M Tris–HCl pH 9.0. Crystals measured approximately 0.12 × 0.12 × 0.06 mm after two weeks at 291 K (Fig. 1 ▶).
Figure 1.

Crystals of crotoxin B from C. durissus collilineatus.
2.3. X-ray data collection and processing
X-ray diffraction data were collected from a single CB crystal at a wavelength of 1.4586 Å (at 100 K) using a synchrotron-radiation source (MX2 station, Laboratório Nacional de Luz Sincroton, LNLS, Campinas, Brazil) and a MAR CCD imaging-plate detector (MAR Research). A crystal was mounted in a nylon loop and flash-cooled in a stream of nitrogen gas at 100 K using no cryoprotectant. The crystal-to-detector distance was 105 mm and an oscillation range of 1° was used, resulting in the collection of a total of 91 images. Data processing was carried out using the HKL program suite (Otwinowski & Minor, 1997 ▶) at 2.2 Å resolution.
3. Results and discussion
The data-collection statistics are shown in Table 1 ▶. The crystals belonged to the tetragonal space group P43212, with unit-cell parameters a = b = 84.86, c = 65.67 Å. The data set is 93.6% complete at 2.2 Å resolution, with an R merge of 10.3%.
Table 1. X-ray diffraction data-collection and processing statistics.
Values in parentheses are for the highest resolution shell.
| Unit-cell parameters (Å) | a = b = 84.9, c = 65.7 |
| Space group | P43212 |
| Resolution (Å) | 50–2.2 (2.3–2.2) |
| Unique reflections | 16135 (1509) |
| Completeness (%) | 92.2 (93.6) |
| Rmerge† (%) | 10.3 (53.7) |
| Radiation source | Synchrotron (MX2 station, LNLS) |
| Data-collection temperature (K) | 100 |
| 〈I/σ(I)〉 | 10.2 (1.97) |
| Redundancy | 2.0 (2.0) |
| Matthews coefficient VM (Å3 Da−1) | 2.1 |
| Molecules in the ASU | 2 |
| Solvent content (%) | 41.8 |
R
merge = 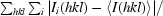
 , where Ii(hkl) is the intensity of an individual measurement of the reflection with Miller indices hkl and 〈I(hkl)〉 is the mean intensity of that reflection. Calculated for I > −3σ(I). Data were processed using the HKL suite (Otwinowski & Minor, 1997 ▶).
, where Ii(hkl) is the intensity of an individual measurement of the reflection with Miller indices hkl and 〈I(hkl)〉 is the mean intensity of that reflection. Calculated for I > −3σ(I). Data were processed using the HKL suite (Otwinowski & Minor, 1997 ▶).
Calculations based on the protein molecular weight indicate the presence of two molecules in the asymmetric unit. This corresponds to a Matthews coefficient (Matthews, 1968 ▶) V M of 2.11 Å3 Da−1 with a calculated solvent content of 41.8%. These values are within the expected range for typical protein crystals, assuming a value of 0.74 cm3 g−1 for the protein partial specific volume.
The crystal structure of CB from C. durissus collilineatus was determined using molecular-replacement techniques implemented in the program MOLREP (Vagin & Teplyakov, 1997 ▶) from the CCP4i program package (Potterton et al., 2003 ▶), using the coordinates of the monomer B of crotoxin B from C. durissus terrificus (PDB code 2qog; Marchi-Salvador et al., 2008 ▶) as a search model. The amino-acid sequences of the two CB isoforms (CB1 and CB2) which formed the asymmetric unit of the crystal used in the X-ray diffraction experiments were the same as those determined by Faure et al. (1994 ▶), as checked by inspection of the electron densities of the model. The structure presented a dimeric arrangement formed by a heterodimer of the CB1 and CB2 isoforms (Fig. 2 ▶) similar to one dimer of the CB structure from C. durissus terrificus (Marchi-Salvador et al., 2008 ▶). Initial analysis with the PISA program (Krissinel & Henrick, 2007 ▶) showed an interface area of 539.6 Å2, Δi G = −35.6 kJ mol−1, Δi G P-value = 0.347 and a CSS (complexation significance score) of 0.437, indicating that the dimeric structure is stable in solution.
Figure 2.

Dimeric structure of crotoxin B from C. durissus collilineatus shown as a ribbon diagram. The monomers of isoforms CB2 and CB1 are shown in blue and red, respectively. This figure was drawn using the PyMOL program (DeLano, 2002 ▶).
In conclusion, crotoxin B isolated from C. durissus collilineatus venom was crystallized and X-ray diffraction data were collected to 2.2 Å resolution. The structure presented a dimeric arrangement formed by the CB1 and CB2 isoforms similar to one dimer of the structure of CB from C. durissus terrificus. Initial analyses indicated that the dimeric structure was stable in solution. Consequently, these data indicate this dimer is a feasible biological assembly, indicating that the interface contacts are not formed as a consequence of crystal packing. Detailed studies of this structure and its structural comparison with crotoxin B from C. durissus terrificus may provide insights into the toxic mechanisms of crotoxin and other PLA2s.
Acknowledgments
The authors gratefully acknowledge financial support from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Laboratório Nacional de Luz Síncroton (LNLS, Campinas-SP).
References
- Aird, S. D., Kaiser, I. I., Lewis, R. V. & Kruggel, W. G. (1985). Biochemistry, 24, 7054–7058. [DOI] [PubMed]
- Aird, S. D., Kaiser, I. I., Lewis, R. V. & Kruggel, W. G. (1986). Arch. Biochem. Biophys.249, 296–300. [DOI] [PubMed]
- Aird, S. D., Yates, J. R., Martino, P. A., Shabanowitz, J., Hunt, D. F. & Kaiser, I. I. (1990). Biochim. Biophys. Acta, 1040, 217–224. [DOI] [PubMed]
- Bon, C. (1997). Venom Phospholipase A2 Enzymes: Structure, Function and Mechanism, edited by R. M. Kini, pp. 269–285. Chichester: John Wiley & Sons.
- Bon, C., Changeux, J. P., Jeng, T. W. & Fraenkel-Conrat, H. (1979). Eur. J. Biochem.99, 471–481. [DOI] [PubMed]
- Corin, R. E., Viskatis, L. J., Vidal, J. C. & Etcheverry, M. A. (1993). Invest. New Drugs, 11, 11–15. [DOI] [PubMed]
- Cura, J. E., Blanzaco, D. P., Brisson, C., Cura, M. A., Cabrol, R., Larrateguy, L., Mendez, C., Sechi, J. C., Silveira, J. S., Theiller, E., de Roodt, A. R. & Vidal, J. C. (2002). Clin. Cancer Res.8, 1033–1041. [PubMed]
- DeLano, W. L. (2002). The PyMOL Molecular Graphics System. DeLano Scientific LLC, San Carlos, California, USA.
- Faure, G. & Bon, C. (1987). Toxicon, 25, 229–234. [DOI] [PubMed]
- Faure, G. & Bon, C. (1988). Biochemistry, 27, 730–738. [DOI] [PubMed]
- Faure, G., Choumet, V., Bouchier, C., Camoin, L., Guillaume, J. L., Monegier, B., Vuilhorgne, M. & Bon, C. (1994). Eur. J. Biochem.223, 161–164. [DOI] [PubMed]
- Faure, G., Guillaume, J. L., Camoin, L., Saliou, B. & Bon, C. (1991). Biochemistry, 30, 8074–8083. [DOI] [PubMed]
- Faure, G., Harvey, A. L., Thomson, E., Saliou, B., Radvanyi, F. & Bon, C. (1993). Eur. J. Biochem.214, 491–496. [DOI] [PubMed]
- Hawgood, B. J. & Smith, J. W. (1977). Br. J. Pharmacol.61, 597–606. [DOI] [PMC free article] [PubMed]
- Hendon, R. A. & Fraenkel-Conrat, H. (1971). Proc. Natl Acad. Sci. USA, 68, 1560–1563. [DOI] [PMC free article] [PubMed]
- Jancarik, J. & Kim, S.-H. (1991). J. Appl. Cryst.24, 409–411.
- Krissinel, E. & Henrick, K. (2007). J. Mol. Biol.372, 774–797. [DOI] [PubMed]
- Laemmli, U. K. (1970). Nature (London), 227, 680–685. [DOI] [PubMed]
- Marchi-Salvador, D. P., Corrêa, L. C., Magro, A. J., Oliveira, C. Z., Soares, A. M. & Fontes, M. R. M. (2008). Proteins, 72, 883–891. [DOI] [PubMed]
- Marchi-Salvador, D. P., Corrêa, L. C., Salvador, G. H. M., Magro, A. J., Oliveira, C. Z., Iulek, J., Soares, A. M. & Fontes, M. R. M. (2007). Acta Cryst. F63, 1067–1069. [DOI] [PMC free article] [PubMed]
- Matthews, B. W. (1968). J. Mol. Biol.33, 491–497. [DOI] [PubMed]
- McPherson, A. (1982). Preparation and Analysis of Protein Crystals. New York: Wiley.
- Otwinowski, Z. & Minor, W. (1997). Methods Enzymol.276, 307–326. [DOI] [PubMed]
- Ponce-Soto, L. A., Toyama, M. H., Hyslop, S., Novello, J. C. & Marangoni, S. (2002). J. Protein Chem.21, 131–136. [DOI] [PubMed]
- Potterton, E., Briggs, P., Turkenburg, M. & Dodson, E. (2003). Acta Cryst. D59, 1131–1137. [DOI] [PubMed]
- Rangel-Santos, A., Dos-Santos, E. C., Lopes-Ferreira, M., Lima, C., Cardoso, D. F. & Mota, I. (2004). Toxicon, 43, 801–810. [DOI] [PubMed]
- Reisfeld, R. A., Lewis, U. J. & Williams, D. E. (1962). Nature (London), 195, 281–283. [DOI] [PubMed]
- Rudd, C. L., Viskatis, L. J., Vidal, J. C. & Etcheverry, M. A. (1994). Invest. New Drugs, 12, 183–184. [DOI] [PubMed]
- Santoro, M. L., Sousa-e-Silva, M. C., Gonçalves, L. R., Almeida-Santos, S. M., Cardoso, D. F., Laporta-Ferreira, I. L., Saiki, M., Peres, C. A. & Sano-Martins, I. S. (1999). Comp. Biochem. Physiol. C, 122, 61–73. [DOI] [PubMed]
- Slotta, K. H. & Fraenkel-Conrat, H. (1938). Ber. Dtsch. Chem. Ges.71, 1076–1081.
- Soares, A. M., Rodrigues, V. M., Homsi-Brandeburgo, M. I., Toyama, M. H., Lombardi, F. R., Arni, R. K. & Giglio, J. R. (1998). Toxicon, 36, 503–514. [DOI] [PubMed]
- Toyama, M. H., Toyama, D. O., Joazeiro, P. P., Carneiro, E. M., Beriam, L. O., Marangoni, L. S. & Boschero, A. C. (2005). Protein J.24, 103–112. [DOI] [PubMed]
- Vagin, A. & Teplyakov, A. (1997). J. Appl. Cryst.30, 1022–1025.
- Vital-Brazil, O. & Excell, B. J. (1970). J. Physiol. (London), 212, 34–35.
- Zhang, H. L., Han, R., Chen, Z. X., Chen, B. W., Gu, Z. L., Reid, P. F., Raymond, L. N. & Oin, Z. H. (2006). Toxicon, 48, 175–182. [DOI] [PubMed]