

Abstract
We investigated the influence of acute wounding on tumor growth in a syngeneic mouse breast cancer model. Metastatic mouse breast cancer cells (4T1) were orthotopically injected into the mammary fat pads of BALB/c mice, and animals were wounded locally by full thickness dermal incisions above the mammary fat pads or remotely above the scapula 9 days later. Local but not remote wounding increased tumor size when compared to sham treatment. Injection of wound fluid close to the tumor site increased tumor growth while in vitro wound fluid as compared to serum increased the proliferation rate of 4T1 cells. Our results show that wound stroma can unfavorably influence growth of nearby tumors. This effect is T-cell dependent as local wounding had no effect on tumor growth in nu/nu mice. The effect of wounding on tumor growth can be mimicked by acellular wound fluid suggesting that T-cells secrete or mediate secretion of cytokines or growth factors that then accelerate tumor growth. Here we define an experimental model of wound promoted tumor growth that will enable us to identify mechanisms and therapeutic targets in order to reduce the negative impact of tissue repair on residual tumors.
Keywords: breast cancer, wound, microenvironment
Introduction
During their lifetime, approximately 10% of American women will be afflicted by breast cancer. Surgical procedures are frequently performed to confirm diagnosis and to resect tumors. They leave behind injured tissue, and initiate a wound healing response that is a sequence of tightly controlled events (1). Initially, cytokines and other effector molecules are released from damaged tissue, initiating a rapid invasion of inflammatory cells into the wound that persists for several days. Inflammatory cells orchestrate the migration and proliferation of a variety of cells leading to remodeling of the injured tissue and tissue repair (1). Wound healing and tumor progression both involve processes of cell proliferation, inflammation, and angiogenesis. Hence, tumors have been described as “wounds that do not heal” (2); conversely “wounds may be regarded as a tumor which heals itself” (3). Clinically, chronic wounds are associated with increased risk of tumor formation (4), as with squamous or basal cell carcinomas that originate from chronic skin wounds (5), implying that wound healing and tumorigenesis may not only share common physiological features but may also influence each other.
The mechanisms of wound promoted tumorigenesis and tumor progression are not fully understood. The impact of acute wounding as occurs during biopsy or extirpative surgery on an existing tumor has not been addressed. Perhaps the greatest risk is the presence of unsuspected microdeposits of tumor left behind after tumor resection. Animal models have been used to define the influence of preexisting wounds on tumor cells, or the influence of wounding on initiated hosts. Tumor incidence and tumor volume are higher if melanoma or fibrosarcoma cells are injected into a wound as compared to unwounded tissue (6;7), indicating that a preexisting wound microenvironment facilitates the establishment of tumors from a tumor cell inoculate. Likewise, the coinjection of wound fluid and melanoma cells resulted in increased tumor volumes (7). It has also been shown that full thickness transcutaneous wounding is a sufficient event for tumor expression and growth in both Rous-sarcoma virus infected chickens and v-ras transgenic mice, demonstrating that acute wounding can promote tumorigenesis in a host that is already initiated by viral infection or by oncogene expression (8-10). These reports suggest a strong interaction between the wound and tumor microenvironments that can accelerate tumorigenesis and tumor progression.
Clinically, surgical procedures are typically performed in the proximity of a pre-existing tumor as a necessary component of tumor treatment. While these procedures attempt to eradicate the tumor for the benefit of the patient, local tumor recurrence and implantation of tumor cells along the wound or the needle tract have been described. This is often attributed to mechanical tumor spread, but the local wound environment itself may similarly influence residual tumor cells. Currently there are few adequate animal models to assess the effect of surgery on a pre-existing tumor and to investigate mechanisms of wound-promoted tumor growth. Here, we describe a syngeneic orthotopic mouse model of wound-promoted tumor growth in breast cancer to evaluate how acute wounds, such as occur during tumor surgery, affect tumor growth.
Materials and Methods
Tissue culture
4T1(11), 4T07, Met1, TSA, (provided by Dr. Fred Miller, Barbara Ann Karmanos Cancer Institute, Detroit, MI) and B16F1 cells (provided by Glenn Merlino, National Cancer Institute, Bethesda)) were grown in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS, Invitrogen, Carlsbad, CA) at 37°C in 5% CO2.
For proliferation assays, cells (2000/well) were plated in 96 well-plates. At 6h medium was changed to DMEM containing 3% mouse serum, or 3% mouse plasma, or 3% wound fluid, and cells were incubated for another 2 days, trypsinized, and counted using a hemocytometer.
Animal Model
We used a syngeneic, immunocompetent mouse model to study the effect of wounding on tumor progression (Fig. 1A). 1×104 4T1 mouse metastatic breast cancer cells (11) were suspended in 50μl of Dulbecco's phosphate buffered saline (Invitrogen, Carlsbad, CA), and injected through the skin using 28G needles into the mammary fat pads IV/V and IX/X of female BALB/c mice (age 8 weeks, Charles River, Frederick, MD). Nine days later, under anesthesia (isoflurane to effect, buprenorphine 0.75μg/g BW i.p), the animals were wounded locally by performing a 1cm long, full thickness dermal incision above the previously inoculated mammary fat pads. Care was taken not to injure the underlying mammary gland in order to avoid mechanical spread of tumor cells. Remote wounding was performed as a 1cm long, bilateral, dermal incision in the suprascapular region 9 days after tumor cell inoculation (Fig 1A). Turpentine (30ul) was injected s.c into the suprascapular pocket 9 days after tumor cell inoculation. Wound fluid (40μl) from 9 day old wounds, or serum, or plasma was injected s.c. daily into the proximity of tumor bearing sites starting 9 days after tumor cell inoculation. Control animals underwent only tumor cell inoculation and anesthesia, but were not wounded. Tumor sizes were evaluated by measuring the two main axes of the tumors with calipers, and tumor volumes estimated as long axis × (short axis)2 × 0.52 (12). Animals were euthanized by CO2 asphyxiation 23 days after tumor cell inoculation, and tumor tissue was collected for histological analyses using routine procedures.
Figure 1.
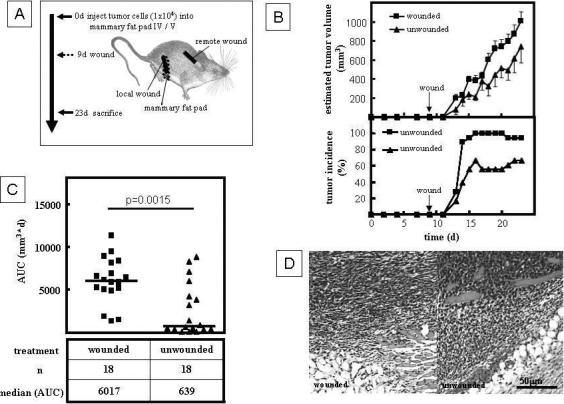
Wounding promotes tumor growth in a mouse model of breast cancer. A. Experimental design. 4T1 cells (104 in 50ul DPBS) were orthotopically injected into the mammary fat pads IV/V and IX/X of syngeneic BALB/c mice, followed by local or remote dermal incision 9 days later. Tumor growth was assessed using calipers. Animals were euthanized and autopsied 23 days after inoculation with tumor cells. B. Local wounding increased tumor volume and tumor incidence in the syngeneic model described in A. Data points present the mean±SEM of n=18 tumors. C. Wounding significantly increased the cumulative tumor mass as measured by AUC (Mann-Whitney Test). D. Histology of tumors dissected from wounded and unwounded animals 23 days after tumor cell inoculation.
Collection of Wound Fluid
Mice were anesthesized as described above, and 8mm polyvinylalcohol sponges (M-Pact, Eudora, KA) were implanted subcutaneously in the lower neck and flanks. To produce day 3−14 wound fluid, mice were euthanized at desired time intervals after implantation, and sponges were removed under aseptic conditions. Wound fluid was mechanically squeezed from the sponge into test tubes, and rendered acellular by centrifugation (800g, 4°C, 20 min). Cell-free wound fluid from 2 to 3 animals was pooled, and stored at −80°C until further use.
Statistical Analysis
Data are presented as average ± standard error of mean (SEM) as indicated in the figure legend. Cumulative tumor mass was calculated as area under the curve (AUC) by plotting tumor volumes and calculating the product of tumor volume at any given day and the time between measurements (days). Statistical analyses using Mann-Whitney Test or ANOVA with Dunn's Multiple Comparison Analysis or Dunnet's Multiple Comparison Test were performed in GraphPad Prism 2.
Results and Discussion
Local but not remote wounding accelerates tumor growth
To investigate the effect of a local wound microenvironment on breast cancer growth, we inoculated the mammary fat pad IV/V and IX/X with 4T1 cells (104/site), and then wounded the wounded the animals in the proximity of the inoculated fat pads by dermal incision (1cm) 9 days later (Fig. 1A). At this point, tumors were undetectable by palpation; however, tumor cell nests in the mammary fat pad were observed histologically (data not shown). Wounding significantly increased tumor incidence, average tumor volume (Fig 1B), and cumulative tumor mass (AUC) when compared to unwounded control animals (Fig.1C). The increased tumor volume was due to an increased tumor mass rather than massive inflammatory infiltration of the wound microenvironment as wounded and unwounded tumors had similar histology (Fig. 1D). Our findings confirm that the local wound microenvironment promotes tumor progression.
Wound promoted tumor growth may be due to local effects of wounding such as cell-cell interactions or local secretion of cytokines and growth factors(13), or to systemic effects such as acute phase response or perioperative stress (14). In order to distinguish these possibilities, we wounded animals either locally, in the proximity of the mammary fat pads IV/V and IX/X, or remotely, in the suprascapular region. Local wounding again significantly increased the tumor burden as compared to sham treatment while remote wounding did not alter tumor growth (Fig. 2A). To exclude the possibility that altered tumor growth is due to a non-specific inflammatory response, we induced an acute phase response by subcutaneous injection of turpentine (15) into suprascapular pockets 9 days after tumor cell inoculation. The induction of a robust acute phase response did not alter tumor growth as compared to unwounded animals (Fig. 2B).
Figure 2.
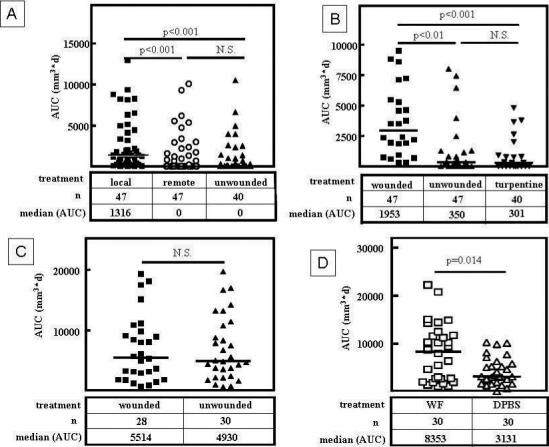
Local but not remote wounding promotes tumor growth in a T-cell dependent fashion. A. In contrast to local wounding, remote wounding by suprascapular dermal incision does not significantly alter the cumulative tumor mass as measured by AUC in an orthotopic syngeneic mouse model of breast cancer (Kruskal-Wallis Test / Dunn's Multiple Comparison Analysis). B. Local wounding but not induction of a systemic acute phase response by injection of turpentine (30μl) into the suprascapular pocket 9 days after tumor cell inoculation into the mammary fat pads IV/V and IX/X significantly increases the cumulative tumor mass (AUC, Kruskal-Wallis Test / Dunn's Multiple Comparison Analysis). C. In nude mice (BALB/c nu/nu) local wounding does not promote tumor growth (Mann-Whitney Test). D. Daily injection of wound fluid (40μl) into the proximity of tumor cell inoculation sites starting 9 days after tumor cell inoculation significantly increased the cumulative tumor mass (AUC) as compared to injection of DPBS (Mann-Whitney Test). Bars indicate medians.
These results indicate that the local wound environment accelerates growth of nearby preexisting tumors. The importance of the local wound microenvironment has also been highlighted in previous studies. For example, wounding is required for local induction of tumor growth in Rous sarcoma virus infected chickens (10); these tumors only originate in the wound bed of infected chickens and not elsewhere. It also has been shown that tumor take is higher and tumor growth is accelerated if melanoma cells (B16) or sarcoma cells (Meth A) are injected into a preexisting cutaneous wound bed (6;7). While these previous experiments indicate an interaction between wounds and tumor growth they are not analogous to the clinical situation as they deal with hosts that have been previously initiated or wounded before tumor inoculation. In contrast, our model more closely mimics the relationship between tumor and wound as it is clinically observed, as we introduce a wound into a pre-existing tumor microenvironment. Our results imply that surgical procedures may influence the progression of biopsied tumors or residual tumors in the proximity of the wound bed after apparently curative surgery.
Wound promoted tumor growth depends on a functional T-cell compartment
The initial inflammatory response during wound healing is mediated by neutrophils, macrophages, and T-lymphocytes(1). We chose to investigate the influence of the T-cell compartment on wound-promoted tumor growth. We inoculated nu/nu BALB/c animals which lack a functional T-cell compartment with 4T1 cells and wounded the animals 9 days later. In this model, wounding did not accelerate tumor growth (Fig. 2C), suggesting that a functional T-cell compartment is necessary for wound-promoted tumor growth.
T-lymphocytes can influence tumor progression directly by secreting cytokines such as interleukins, TNF-α, VEGF, or TGF-β, or indirectly by orchestrating other immune cells and modulating angiogenesis (13;16). We next studied whether soluble factors in wound fluid are sufficient to stimulate tumor growth. We therefore injected wound fluid into the proximity of the tumor cell inocculum starting 9 days after tumor cell inoculation. Injections of wound fluid significantly increased tumor growth as compared to injection of DPBS (Fig. 2D), demonstrating that the effect of wounding on tumor growth is partially due to effectors present in the acellular wound fluid that are neither cell-bound nor matrix-bound.
We addressed next whether tumor cells might be direct targets for the growth-promoting effect of wound fluid. We cultured several tumor cells lines (4T1, 4T07, B16F1, TSA, Met1) in vitro with wound fluid or mouse serum. We observed an increase in cell number when cells were stimulated with wound fluid from 9 day old wounds as compared to serum treated cultures (Fig 3A), while at the same time apoptosis was increased in wound fluid treated cultures (Fig. S1). This indicates that wound fluid increases proliferation of tumor cells. For 4T1 cells, we observed a 1.8 fold increase in cell number in cultures treated with wound fluid as compared to mouse serum (p<0.01, ANOVA / Dunnet's Multiple Comparison Test, Fig. 3B). Wound fluid from 6 day to 9 day old wounds was more effective in stimulating proliferation of 4T1 than early (day 3) or late stage (day 10 to 14) wound fluid (Fig. 3); this coincides with the wound healing phase of lymphocyte infiltration(1). Furthermore, while wound fluid from immunocompetent BALB/c mice significantly increased tumor cell proliferation in-vitro, wound fluid from nu/nu BALB/c animals was ineffective (Fig.3B), indicating that wound-activated T-lymphocytes secrete or mediate secretion of effectors into the microenvironment that promote tumor growth. In-vivo, we observed an increase of cell proliferation in tumors of wounded animals (Fig. 3C), confirming that increased cell proliferation is a relevant mechanism of wound promoted tumor growth. Known factors present in wound fluid and capable of promoting tumor cell proliferation include FGF, EGF, PDGF and interleukins(7;17-19). FGF, EGF, and PDGF are not typically secreted by T-lymphocytes and are therefore unlikely to mediate the effect of wounds on tumorigenesis. TGF-β has been shown to promote tumor growth when coinjected with melanoma cells (B16F10)(7) and subcutaneous injection of TGF-β can substitute for wounding in tumor induction in Rous sarcoma virus infected chickens(10). Yet we see no growth promoting or survival-stimulating effect of TGF-β on 4T1 cells in vitro(20), making it unlikely that TGF-β contributes to wound-promoted tumor growth in the 4T1 model by increasing tumor cell proliferation in vivo. It is our goal to eventually identify how T-cells mediate the effect of wounding on tumor growth.
Figure 3.
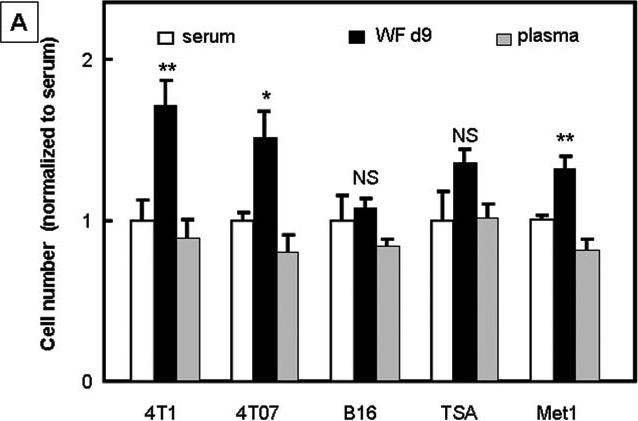

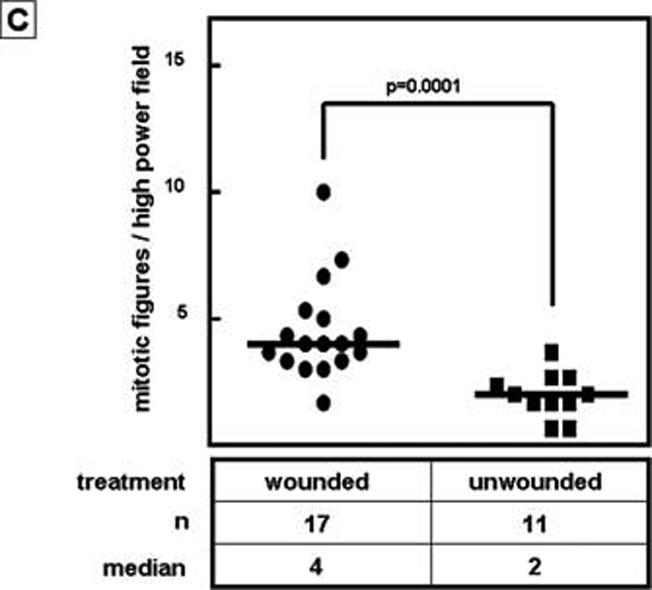
Influence of wound fluid and wounding on tumor cell proliferation. A. Wound fluid increases tumor cell proliferation in vitro. Cells were stimulated with 3% wound fluid or serum or plasma, or were serum starved (0%) for 2 days. Cell numbers were counted and normalized to the average cell number of serum treated cells. ANOVA / Dunnet's Multiple Comparison Test. Data are presented as mean±SEM. B. Wound fluid generated from BALB/c mice but not wound fluid generated from nude mice (BALB/c nu/nu) promotes proliferation of 4T1 cells in vitro. The effect of wound fluid not only depended on the mouse strain, but also on the age of the wound fluid; wound fluid harvested 6 days to 9 days after wounding was most effective in stimulating tumor cell proliferation. ANOVA / Dunnet's Multiple Comparison Test. Data are presented as mean±SEM. C. Wounding increases tumor cell proliferation in-vivo. Mitotic tumor cells were counted in H&E stained sections of tumors obtained from wounded and unwounded animals 23 days after tumor cell inoculation. Student's t-test.
In summary, we have developed a model which allows investigations into the mechanisms of the effect of wounding on pre-existing tumors. The negative impact of wounding or tumor surgery appears to correlate with the inflammatory phase that involves infiltration of the injured tissue by T-lymphocytes and cytokine secretion. Tumors are frequently biopsied or debulked prior to further treatment, procedures that can potentially leave behind residual tumor tissue. Given the negative impact of the wound microenvironment on tumor progression, it will be important to investigate the exact mechanisms by which T-lymphocytes contribute to accelerated tumor growth in the proximity of wounds. Our demonstration that the tumor promoting effect of local wounds on tumor growth can be mimicked at least partially by wound fluid provides an experimental platform for identifying critical T-cell mediated factors driving this process. Understanding and controlling these mechanisms will contribute to avoiding a negative impact of surgery on residual tumor growth.
Reference List
- 1.Park JE, Barbul A. Understanding the role of immune regulation in wound healing. Am J Surg. 2004;187:11S–6S. doi: 10.1016/S0002-9610(03)00296-4. [DOI] [PubMed] [Google Scholar]
- 2.Dvorak HF. Tumors: wounds that do not heal. Similarities between tumor stroma generation and wound healing. N Engl J Med. 1986;315:1650–9. doi: 10.1056/NEJM198612253152606. [DOI] [PubMed] [Google Scholar]
- 3.Haddow A. Molecular repair, wound healing, and carcinogenesis: tumor production a possible overhealing? Adv Cancer Res. 16:181–234. doi: 10.1016/s0065-230x(08)60341-3. [DOI] [PubMed] [Google Scholar]
- 4.Trent JT, Kirsner RS. Wounds and malignancy. Adv Skin Wound Care. 2003 Jan;16:31–4. doi: 10.1097/00129334-200301000-00014. [DOI] [PubMed] [Google Scholar]
- 5.Mueller MM, Fusenig NE. Tumor-stroma interactions directing phenotype and progression of epithelial skin tumor cells. Differentiation. 2002;70:486–97. doi: 10.1046/j.1432-0436.2002.700903.x. [DOI] [PubMed] [Google Scholar]
- 6.Baker DG, Masterson TM, Pace R, Constable WC, Wanebo H. The influence of the surgical wound on local tumor recurrence. Surgery. 1989;106:525–32. [PubMed] [Google Scholar]
- 7.Hofer SO, Shrayer D, Reichner JS, Hoekstra HJ, Wanebo HJ. Wound-induced tumor progression: a probable role in recurrence after tumor resection. Arch Surg. 1998;133:383–9. doi: 10.1001/archsurg.133.4.383. [DOI] [PubMed] [Google Scholar]
- 8.Dolberg DS, Hollingsworth R, Hertle M, Bissell MJ. Wounding and its role in RSV-mediated tumor formation. Science. 1985;230:676–8. doi: 10.1126/science.2996144. [DOI] [PubMed] [Google Scholar]
- 9.Schuh AC, Keating SJ, Monteclaro FS, Vogt PK, Breitman ML. Obligatory wounding requirement for tumorigenesis in v-jun transgenic mice. Nature. 1990;346:756–60. doi: 10.1038/346756a0. [DOI] [PubMed] [Google Scholar]
- 10.Sieweke MH, Thompson NL, Sporn MB, Bissell MJ. Mediation of wound-related Rous sarcoma virus tumorigenesis by TGF-beta. Science. 1990;248:1656–60. doi: 10.1126/science.2163544. [DOI] [PubMed] [Google Scholar]
- 11.Nam JS, Suchar AM, Kang MJ, et al. Bone Sialoprotein Mediates the Tumor Cell-Targeted Prometastatic Activity of Transforming Growth Factor {beta} in a Mouse Model of Breast Cancer. Cancer Res. 2006;66:6327–35. doi: 10.1158/0008-5472.CAN-06-0068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tang B, Vu M, Booker T, et al. TGF-beta switches from tumor suppressor to prometastatic factor in a model of breast cancer progression. J Clin Invest. 2003;112:1116–24. doi: 10.1172/JCI18899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hofer SO, Molema G, Hermens RA, Wanebo HJ, Reichner JS, Hoekstra HJ. The effect of surgical wounding on tumour development. Eur J Surg Oncol. 1999;25:231–43. doi: 10.1053/ejso.1998.0634. [DOI] [PubMed] [Google Scholar]
- 14.Goldfarb Y, Ben-Eliyahu S. Surgery as a risk factor for breast cancer recurrence and metastasis: mediating mechanisms and clinical prophylactic approaches. Breast Dis. 2006;26:99–114. doi: 10.3233/bd-2007-26109. [DOI] [PubMed] [Google Scholar]
- 15.Burmester H, Wolber EM, Freitag P, Fandrey J, Jelkmann W. Thrombopoietin production in wild-type and interleukin-6 knockout mice with acute inflammation. J Interferon Cytokine Res. 2005;25:407–13. doi: 10.1089/jir.2005.25.407. [DOI] [PubMed] [Google Scholar]
- 16.de Visser KE, Eichten A, Coussens LM. Paradoxical roles of the immune system during cancer development. Nat Rev Cancer. 2006;6:24–37. doi: 10.1038/nrc1782. [DOI] [PubMed] [Google Scholar]
- 17.Aiba-Kojima E, Tsuno NH, Inoue K, et al. Characterization of wound drainage fluids as a source of soluble factors associated with wound healing: comparison with platelet-rich plasma and potential use in cell culture. Wound Repair Regen. 2007;15:511–20. doi: 10.1111/j.1524-475X.2007.00259.x. [DOI] [PubMed] [Google Scholar]
- 18.Lanigan F, O'Connor D, Martin F, Gallagher WM. Molecular links between mammary gland development and breast cancer. Cell Mol Life Sci. 2007;64:3159–84. doi: 10.1007/s00018-007-7386-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nicolini A, Carpi A, Rossi G. Cytokines in breast cancer. Cytokine Growth Factor Rev. 2006;17:325–37. doi: 10.1016/j.cytogfr.2006.07.002. [DOI] [PubMed] [Google Scholar]
- 20.Nam JS, Terabe M, Mamura M, et al. An anti-transforming growth factor beta antibody suppresses metastasis via cooperative effects on multiple cell compartments. Cancer Res. 2008;68:3835–43. doi: 10.1158/0008-5472.CAN-08-0215. [DOI] [PMC free article] [PubMed] [Google Scholar]