

Abstract
The primary cause of cancer mortality is not attributed to primary tumor formation, but rather to the growth of metastases at distant organ sites. Tumor cell adhesion to blood vessel endothelium (EC) and subsequent transendothelial migration within the circulation are critical components of the metastasis cascade. Previous studies have shown polymorphonuclear neutrophils (PMNs) may facilitate melanoma cell adhesion to the EC and subsequent extravasation under flow conditions. The melanoma cell–PMN interactions are found to be mediated by the binding between intercellular adhesion molecule-1 (ICAM-1) on melanoma cells and β2 integrin on PMNs and by endogenously secreted interleukin 8 (IL-8) within the tumor-leukocyte microenvironment. In this study, the effects of fluid convection on the IL-8-mediated activation of PMNs and the binding kinetics between PMNs and melanoma cells were investigated. Results indicate that the shear rate dependence of PMN–melanoma cell adhesion and melanoma cell extravasation is due, at least partly, to the convection of tumor-secreted proinflammatory cytokine IL-8.
Keywords: Melanoma, Metastasis, Interleukin 8, Signaling, Neutrophil, Endothelium, Shear stress, Shear rate
INTRODUCTION
Malignant melanoma is highly aggressive and forms metastases with high probability. Most melanoma deaths are due to metastases that are resistant to conventional therapies.23,27 Tumor metastasis consists of a complex cascade, in which adhesion to the vascular endothelium (EC) is a critical step. Human neutrophils (polymorphonuclear neutrophils—PMNs), which comprise 50–70% of circulating leukocytes, have been shown to promote metastasis under certain circumstances.11,12,22,25,26 Binding between intercellular adhesion molecule-1 (ICAM-1) on malignant melanoma cells and β2 integrins (e.g., CD11a/CD18 or LFA-1; and CD11b/CD18 or Mac-1) on PMNs enhances melanoma adhesion to and migration through the EC.12,26 β2 integrins on PMNs are up-regulated when stimulated by chemoattractant, which appears to be essential to PMN adhesion to the EC and subsequent migration to the surrounding tissue in response to inflammation. Recent studies have shown that PMN tethering on the EC is necessary for melanoma cell adhesion to the EC in a shear flow.12,26 It has been widely reported that PMN tethering to the EC is affected by both the shear rate and shear stress. In contrast, PMN-facilitated melanoma cell adhesion to the EC and subsequent transendothelial migration are solely dependent on the shear rate.12,13 However, the mechanism for shear rate-dependence of melanoma cell adhesion to the EC via binding to PMNs has not been understood. The objective of this study is to gain some insights in this void.
Melanomas have been found to constitutively secrete a number of chemokines, including interleukin 8 (IL-8).19,20 Recent reports suggest that the expression of chemokines and chemokine receptors by melanoma may contribute to its ability to escape tumor surveillance and may partially explain preferential patterns of melanoma metastasis.16 IL-8 has a wide range of pro-inflammatory effects on the migration of PMNs from the circulation to a site of injury through CXC chemokine receptors 1 and 2 (CXCR1 and CXCR2).6,17 IL-8 has been shown to trigger functional up-regulation of the β2 integrins binding activity on PMNs.21 This up-regulation may enhance the binding between ICAM-1 on melanoma cells and the up-regulated β2 integrins on PMNs, thus enhancing melanoma cell adhesion to the EC mediated by the PMN. In blood vessels, the chemokines released by melanoma cells may stimulate PMNs within the tumor microenvironment.19 The concentration of chemokine that accumulates at the surface of an effected PMN is governed by the transport of the soluble chemokine by the shear flow. There is little understanding of the diffusion of tumor-secreted chemokines under hydrodynamic shear conditions and their subsequent effects on the activation of PMNs.
Cell adhesion is mediated by the formation of the reversible binding of receptors and ligands. Chemical kinetics is the study of the rates of chemical reactions and has been used to study this binding.2 The association rate is defined as the rate at which a receptor and ligand will combine to form a bond. The dissociation rate is the rate at which a bond will dissociate into its constituent receptor and ligand. These rates are dependent on physical factors such as the number of ligands available to a receptor, the distance between the receptor and ligand, the temperature, and the force applied to the bond. The balance of the association and dissociation rates determines if two cells adhere or not. It is not known how the properties of the shear flow affect these association and dissociation rates of binding between β2 integrins-expressing PMNs and ICAM-1-expressing melanoma cells.
In this study, the effect of IL-8 secreted by melanoma cells on PMN activation and subsequent melanoma–PMN adhesion to the endothelium under various flow conditions is investigated. How IL-8 changes the PMN surface density of LFA-1 and Mac-1 is determined experimentally. To determine the effects of melanoma cell-secreted IL-8 on PMNs in a shear flow, a computational model is used to simulate the transport of IL-8 from a free stream melanoma cell to accumulate near a rolling PMN. The increase in PMN–melanoma cell binding potential due to the IL-8 stimulation is quantified by computing the association rate of LFA-1 and Mac-1 molecules after being stimulated by melanoma cells under various flow conditions. Since bond dissociation is determined by the force applied to the bond, the bond force applied to a melanoma cell by one bond between it and a PMN under various flow conditions is simulated using a fluid dynamics model.
MATERIALS AND METHODS
Reagents
Mouse anti-human CD11a, CD11b, and ICAM-1 monoclonal antibodies (mAbs) were purchased from Invitrogen (Carlsbad, CA). Human recombinant IL-8 was purchased from R&D systems (Minneapolis, MN). Other reagents were purchased from R&D systems unless otherwise mentioned.
Cell Preparations
WM9 melanoma cells (provided by Dr. M. Herlyn, Wistar Institute, Philadelphia, PA) were grown in RPMI-1640 (Invitrogen) supplemented with 10% FBS (Invitrogen) and 100 units mL−1 penicillin–streptomycin (Biosource, Inc.) at 37 °C and 5% CO2. We used a transfected fibroblast L-cell line5 that constitutively expresses human E-selectin and ICAM-1 as an adhesive substrate. This cell line (provided by Dr. S. I. Simon, University of California Davis), which expresses levels of E-selectin and ICAM-1 that are comparable to those found on human umbilical vein endothelial cells, provides a model to explore melanoma cell adhesion. For experiments which required melanoma cells in suspension, cells were detached when confluent using trypsin/versine (Invitrogen) and washed twice with fresh medium. Then the cells were re-suspended in fresh medium and allowed to recover for 1 h while being rocked at 8 rpm at 37 °C.
Following the Pennsylvania State University Institutional Review Board (IRB) approved protocols (#19311), fresh human blood was collected from healthy donors by venipuncture. PMNs were isolated using a Histopaque® (Sigma) density gradient by manufacturer’s instruction and kept at 4 °C in Dulbecco’s PBS (D-PBS) containing 0.1% human serum albumin (HSA) for up to 4 h before use.
Expression of β2 Integrins on PMNs Upon Stimulation
PMNs were incubated with various concentrations of IL-8 at 37 °C for different time periods and then the β2 integrin expression was measured. Briefly, PMNs were incubated with saturating concentrations of primary mAbs directed against either LFA-1 or Mac-1 (1 μg mL−1) in PBS containing 1% BSA for 20 min at 4 °C and then washed twice. After an additional 20 min incubation with TRITC-conjugated goat anti-mouse Fab2 fragment (1 μg/106 cells; Jackson Immuno-Research, West Grove, PA) at 4 °C, the cells were washed twice and fixed with 2% formaldehyde and analyzed by a GUAVA personal flow cytometry (GUAVA technologies Inc., Burlingame, CA.).
PMN Rolling Velocity
PMN rolling velocities on confluent EI monolayer were measured under various shear conditions in a parallel-plate flow chamber (Glycotech, Rockville, MD) mounted on the stage of a phase-contrast optical microscope (Diaphot 330, Nikon, Japan). Briefly, a syringe pump (Harvard Apparatus, South Natick, MA) was used to generate a steady flow field in the flow chamber. A petri dish (35 mm) with a confluent EI cell monolayer (acting as a ligand-binding substrate) was attached to the flow chamber. All experiments were performed at 37 °C. The field of view was 800 μm long (direction of the flow) by 600 μm. The focal plane was set on the EI monolayer. PMNs (1 × 106 cells mL−1 in DPBS containing 0.1% HSA) were perfusion into parallel plate flow chamber at various flow rates. To increase the viscosity in some experiments, 1 or 2% (wt./vol.) dextran (2 × 106 Mr; Sigma) was added to the medium. The viscosity of the medium was measured as 1.0 cP (no dextran), 2.0 cP (1% dextran), or 3.2 cP (2% dextran) at 37 °C in a cone-plate viscometer (RotoVisco 1, Haake, Newington, NH). The interactions of PMNs with EI monolayer in the parallel plate flow chamber were recorded for two minutes using a CCD camera (pco 1600, Cooke Corporation, Romulus, MI) and the rolling velocity for each PMN was determined by tracking individual PMN with Image-ProPlus 5.0 (Media Cybernetics, Inc., Silver Spring, MD). The mean rolling velocity of a PMN under various shear stresses was calculated from a total of 40 rolling PMNs.
Diffusion and Convection of Solutes in a Shear Flow
A two-dimensional Couette flow model was used to simulate the transport of IL-8 released by tumor cells in the circulation. The inputs used in the model are tabulated in Table 1.
TABLE 1.
Description and source of variables as inputs for Comsol simulation.
| Variable | Description | Value | Source |
|---|---|---|---|
| Q | Volumetric flow rate | 0.025, 0.04, and 0.08 mL min−1 | Experimental setting |
| DAB | IL-8 diffusion coefficient | 2.59 × 10−10 m2 s−1 | (15,29) |
| C0 | IL-8 secreted by a melanoma cell | Figure 4 | Detected using ELISA from medium collected after flow assay |
| a | Distance from the center of channel to the wall | 63.5 μm | Parallel plate chamber geometry |
| ρ | Density of fluid | 1.0 × 103 kg m−3 | Known constant |
| u | Melanoma cell velocity | 9.046 × 10−5, 1.447 × 10−4, and 2.894 × 10−4 m s−1 | Calculated from Eqs. (3) and (4) |
| μ | Viscosity | 1.0, 2.0, and 3.2 cP | Experimental setting |
IL-8 distribution under flow conditions was modeled computationally using the commercial software package Comsol Multiphysics 3.2 (Comsol, Stockholm, Sweden), which employs a finite element method to solve the governing partial differential equations. To simulate the IL-8 transport, the convection and diffusion application mode was selected and the Navier–Stokes application mode was used to calculate the velocity pattern. The convection–diffusion equation solved is shown in Eq. (1):
| (1) |
where ci is the IL-8 concentration, u is the velocity, and Di is the solute diffusion coefficient. The diffusion coefficient for IL-8 was obtained from literature.15 It has been shown that for small proteins, the diffusion coefficient is proportional to molecular weight.28,29 A constant flux was assigned at the cell surface and complete dilution was assumed very far from the cell; mathematically stated as:
where C0 is the IL-8 concentration and R is the tumor cell radius. It was assumed that the secreted IL-8 would not be re-consumed by the secreting tumor cell and C0 be uniform over the surface of the cell.
The velocity profile was calculated using the Navier–Stokes application mode, which solves the incompressible momentum and mass conservation:
| (2) |
where u is the velocity, ρ is the density of fluid, μ is the viscosity and p is the pressure. The fluid was assumed to be Newtonian, incompressible, steady, and laminar. A no-slip boundary condition was implemented at the channel walls. Uniform velocity and constant pressure were prescribed at the inlet and outlet, respectively. In the fully developed region, the classical parabolic velocity profile (Poiseuille flow) was obtained. The velocity at a given location in the chamber can be calculated from the equation below:
| (3) |
where the ΔP/L is the driving force, r is the distance from the center of the channel to the location, μ is the viscosity, and a is the distance from the center of the channel to the channel wall. The driving force, ΔP/L, can be derived from the equation:
| (4) |
where Q is the volumetric flow rate.
The relation between diffusion coefficient and solvent viscosity can be calculated using the Stokes–Einstein equation as follows.
| (5) |
where DAB is the diffusion coefficient of solute A in solvent B, RA is the hydrodynamic radius of solute A, κ is Boltzmann’s constant, T is the Kelvin temperature, and μB is the solvent viscosity. Note that the Stokes–Einstein equation usually gives poor accuracy when solute A is a large molecule. Recently, a modification of the Stokes–Einstein equation for small molecules was proposed by Kooijman.10 However, both equations have shown that the diffusion coefficient is inversely proportional to the solution viscosity.
Receptor–Ligand Binding Association Rate
A molecular model of receptor–ligand binding has been developed and used in simulations of cell–cell adhesion.2,3 In this model, the association rate, kon, governs the likelihood of a receptor to form a bond with a ligand on another cell, whose equation is shown in Eq. (6)9:
| (6) |
where AL is the surface area on a ligand-bearing cell that is available to a receptor, nL is the number of ligands on a cell, and nB is the number of bonds already formed. These values and the separation distance between two cells, ε, are determined by the geometry and properties of a chosen cell. The association rate for the receptor–ligand binding under zero-force conditions, , was determined for LFA-1 and Mac-1 binding with ICAM-1 in a companion effort.7 The bond spring constant, σts, and equilibrium length, λ, as well as the Boltzmann’s constant, kb, were assigned values found in the literature.3
Model of Melanoma–PMN Adhesion to the Endothelium in a Shear Flow
A three-dimensional model of a PMN and a tumor cell in flow was developed in a companion effort to simulate the adhesion of a melanoma cell to a tethered PMN under flow conditions.8 The model is used here to compare the force exerted on a melanoma cell, bound to a stationary PMN via a single bond, under varying flow conditions. Since bond formation is a random occurrence, which is based on the proximity of cells, receptor and ligand surface densities and locations, and molecular properties, bonds may form in any position between a melanoma cell and a PMN. Here, a direct comparison of bond force time history in various flow conditions is desired; thus the same initial location is assumed for the melanoma cell and the single bond in all simulations.
One stationary, adherent PMN is modeled on the bottom plate of the parallel plate flow chamber geometry and a free stream tumor cell is modeled above and slightly downstream of the PMN in the flow. A single bond, treated as a linear bond spring, is seeded between the two cells. The forces on the tumor cell due to the fluid and the bond are used to calculate the cell motion.
Two commercial codes were used to simulate the two cell system. Harpoon (Sharc, Manchester, UK) automatically generates the computational grids, and AcuSolve (ACUSIM Software, Mountain View, CA) calculates the flow profile and force distributions. The cell motion calculation was completed by an author-developed Python script, which also controls the overall simulation. Figure 1 shows the order of operations controlled by the script.
FIGURE 1.
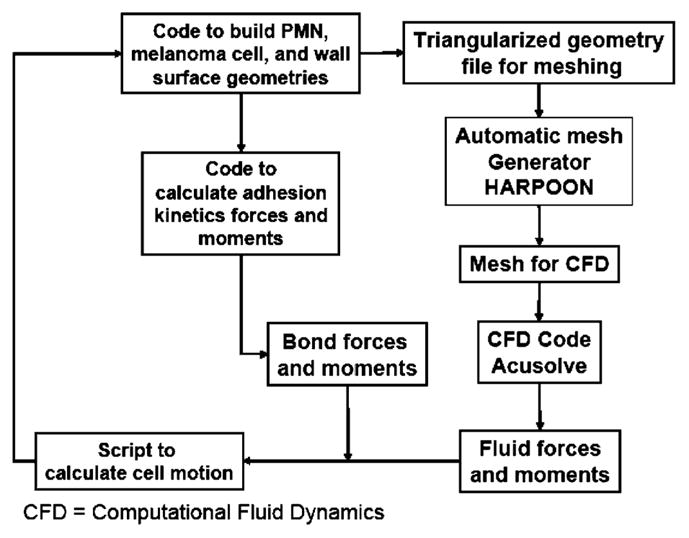
Order of operations for CFD simulations.
Harpoon is a grid generator that creates hexahedra-dominant meshes in a very short time. In the simulations used in this study, a new grid was generated at each time step with the new tumor cell location.
AcuSolve is an incompressible flow solver based on the Galerkin/Least Squares finite element method (GLS). The steady, incompressible Navier–Stokes and continuity equations are solved, as shown in Eq. (2). Variables are also as defined for Eq. (2).
The solution of Newton’s Second Law, shown in Eq. (7), governs the spherical tumor cell motion.
| (7) |
Here x→ and θ→ are the tumor cell translations and rotations, F→ and τ→ are the force and torque vectors on the tumor cell and m and I are the cell’s mass and mass moment of inertia. These equations are solved assuming the tumor cell is a rigid body.
To represent the interactions between the microvilli on the cells’ surfaces, a repulsion force with the form of a non-linear spring force, as shown in Eq. (8), is applied to the tumor cell.
| (8) |
Here k and b are constants and d is the distance between the cells. This force is applied along the normal to the PMN surface at the point of minimum separation distance between the cells. The line of the normal does not generally go through the center of the tumor cell, thus a torque is also applied.
IL-8 Secretion from Melanoma Cells in a Shear Flow
Flow assays were carried out using a continuous circulating loop. Briefly, WM9 cells (2.5 × 106 cells mL−1) suspended in 1640 RPMI with 0.1% BSA were perfused through the flow loop under various shear flow conditions for 4 h in a 37 °C, 5% CO2 incubator. Cell viability at the end of each experiment was determined using Diff Quick stain (Dade Behring Inc., Newark, DE). Medium was collected in the end of this flow assay for ELISA test to detect IL-8 secretion.
ELISA
Cell-free supernatants were collected by a centrifugation at 430g for 5 min and IL-8 was quantified by a sandwich ELISA following standard protocols. Primary and secondary antibody pairs were obtained from R&D Systems (catalog number MAB208 and BAF208). Standard human recombinant IL-8 was also obtained from R&D Systems (catalog number 208-IL) and a standard curve was included in each ELISA plate. Plates were read on a Packard Spectracount at 405 nm and the data analyzed using I-Smart Software. Intra assay variation was typically 10–15%.
Statistical Analysis
All experimental results are reported as the mean ± standard error of the mean (SEM) unless otherwise stated. One-way ANOVA analysis was used for multiple comparisons and t-tests were used for comparisons between 2 groups. p < 0.05 was considered significant.
RESULTS
Expression of LFA-1 and Mac-1 on PMNs upon IL-8 Stimulation
The number of β2 integrins expressed by PMNs at rest had been determined previously to be 45 LFA-1 μm−2 and 5 Mac-1 μm−2.24 To determine the expression of LFA-1 and Mac-1 on PMNs after stimulation, PMNs were treated with various concentrations (0.01, 0.1, 1, 5, and 10 nM) of IL-8 for various time periods (5, 10, 30, 60, 120, 300, and 600 s). By converting the fluorescence intensity changes to a ratio of activated to non-activated intensity and using the known baseline expressions, the site density of LFA-1 and Mac-1 on the activated PMNs was estimated. In response to IL-8 stimulation, both the affinity and avidity of LFA-1 and Mac-1 may change. Here, these changes are assumed to be reflected by a change in the number of active LFA-1 and Mac-1 molecules. The expression of Mac-1 and LFA-1 were upregulated upon IL-8 stimulation in a time- and dose-dependent manner (Fig. 2). After stimulation with a low concentration of 0.01 nM of IL-8, Mac-1, and LFA-1 were not upregulated. Mac-1 and LFA-1 expressions started to increase upon stimulation when IL-8 concentration reached 0.1 nM, indicating a possible threshold concentration for IL-8 to increase the expression of LFA-1 and Mac-1. In addition, there were no significant differences of LFA-1 or Mac-1 expression after increasing the IL-8 concentration from 1 to 5 or 10 nM, which implies that there might be a saturating concentration for IL-8 to up-regulate LFA-1 and Mac-1 expression.
FIGURE 2.

Mac-1 and LFA-1 expression profiles on PMNs after IL-8 stimulation. PMNs were treated with various concentration of IL-8 for various durations. Following treatment, the LFA-1 and Mac-1 expressions on PMNs were detected using flow cytometry. Site density data were calculated by multiplying the ratio of activated to non-activated fluorescence intensity and the known expression numbers of Mac-1 (5 mole μm−2) and LFA-1 (45 mole μm−2) on non-activated PMNs (24). (a) LFA-1 expression on PMNs after IL-8 stimulation; (b) Mac-1 expression on PMNs after IL-8 stimulation. Values were mean ± SEM for N ≥ 3. ○, No stimulation; ●, 0.01 nM; ▼, 0.1 nM; △, 1 nM; ■, 5 nM; □, 10 nM. Inset: first two minutes of stimulation.
The response of LFA-1 upon IL-8 stimulation is rapid. Stimulation with 1 nM IL-8 elicited LFA-1 expression that increased by ~4-fold over the first minute (Fig. 2a inset), culminating in ~5-fold rise within 10 min (Fig. 2a). In contrast, Mac-1 responded to IL-8 in the later time duration (Fig. 2b inset). Stimulation with IL-8 (1 nM) elicited Mac-1 expression that increased by 1-fold only after 2 min, culminating in 9-fold rise within 10 min (Fig. 2b).
Based on these experimental data, curve fitting was used to determine equations for LFA-1/Mac-1 expression (1) with a specific concentration for various time periods of stimulation, and (2) with various concentrations at a specific stimulation time point. Figure 3 shows examples of LFA-1 expression after stimulation by IL-8 with various concentrations for 10 s (Fig. 3a) or after 1 nM IL-8 stimulation for various time periods (Fig. 3b).
FIGURE 3.

LFA-1 expression upon activation of various concentrations of IL-8 for 10 s or of 1 nM IL-8 for different time periods. Based on the data shown in Fig. 2, the LFA-1 or Mac-1 site density on a PMN was derived upon IL-8 stimulation at various concentrations for same period or at the same concentration for various time periods. Using curve fitting program in SigmaPlot 8.0 (Systat Software Inc., San Jose, CA), the expression equation of the site density was plotted. (a) LFA-1 expression upon stimulation of various IL-8 concentrations for 10 s; (b) LFA-1 expression upon 1 nM IL-8 stimulation for various time periods. LFA-1 expression under other stimulation conditions as well as Mac-1 expression were derived the same way.
IL-8 Secretion from Melanoma Cells in a Shear Flow
To determine the secretion of IL-8 from melanoma cells under flow conditions, WM9 cells were perfused into a continuous circulation flow loop driven by a peristaltic pump, and the amount of secreted IL-8 in the suspension was detected by ELISA. Results showed that the secretion of IL-8 from WM9 cells was not affected by the flow conditions (Fig. 4). IL-8 secreted by a single WM9 cell (calculated by dividing the concentration by the number of cells) was used in the Comsol Multiphysics simulation.
FIGURE 4.

IL-8 secretion from a single WM9 under various flow conditions. WM9 cells (2.5 × 106 cells mL−1) suspended in 1640 RPMI with 0.1% BSA were perfused through a flow loop under various shear flow conditions for 4 h. Cell viability at the end of each experiment was determined using Diff Quick stain. Medium was collected at the end of this flow assay for ELISA test to detect IL-8 secretion. IL-8 concentration from a single WM9 cell was calculated based on the experimental data. Values were mean ± SEM for N ≥ 3.
Rolling Velocity of PMN Under Flow Conditions
Previous study has shown that PMN-facilitated melanoma adhesion is a multiple step process which begins with PMNs tethering on the EC, followed by melanoma cells adhering to tethered PMNs, thus bringing tumor cells into close contact with the EC.12 Since the PMN tethering step is an important step in this process, PMN rolling velocity was measured under various shear conditions. Results indicate that PMN rolling velocity was dependent on the shear rate (Fig. 5). Under a fixed shear rate, an increase in shear stress, via increasing the media viscosity, did not change the PMN rolling velocity significantly. However, under the same shear stress, an increase in shear rate (via decreasing the media viscosity) resulted in increased PMN rolling velocity.
FIGURE 5.

PMN rolling velocity. PMNs (1 × 106 cells mL−1 in DPBS containing 0.1% HSA) were perfused into parallel plate flow chamber at various flow rates. To increase the viscosity in some experiments, 1 or 2% (wt./vol.) dextran (2 × 106 Mr) was added to the media. The viscosity of the media was measured as 1.0 cP (no dextran), 2.0 cP (1% dextran), or 3.2 cP (2% dextran) at 37 °C in a cone-plate viscometer. The interactions of PMNs with EI monolayer in the parallel plate flow chamber were recorded for two minutes using a CCD camera and the rolling velocity for each PMN was determined by tracking individual PMN with Image-ProPlus 5.0. The line in each figure indicates the mean rolling velocity of a PMN under various shear stresses, as calculated from 40 rolling PMNs.
Fluid Convection Affects IL-8-Mediated PMN Activation Within the Tumor Microenvironment
To investigate how the fluid convection affects IL-8 secretion by melanoma cells, which in turn affects the expression of LFA-1 and Mac-1 on PMNs and modifies tumor cell adhesion to the EC, Comsol Multi-physics was used to simulate the convection and diffusion of IL-8 from a moving cell under flow conditions (Fig. 6). To simulate the system, a tumor cell was assumed to be in the free stream in the near-wall region and its velocity was derived based on the shear flow condition, using Eq. (3). A PMN was assumed to roll on the EC at the experimentally derived average rolling velocity for the shear condition. The local IL-8 concentration near a rolling PMN and the time at which the PMN is activated by the transported IL-8 were derived to determine the extent of LFA-1 and Mac-1 activation.
FIGURE 6.
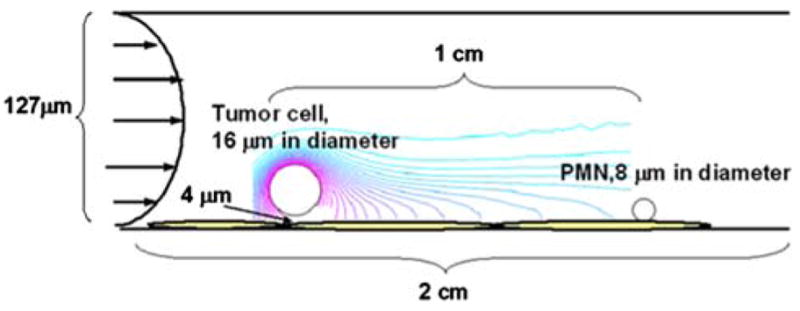
Schematic of IL-8 transport simulation. A parallel plate is 2 cm in length and 127 μm in height. IL-8 was assumed to be constitutively secreted from a moving tumor cell and evenly distributed on cell surface. The concentration of IL-8 secreted by one tumor cell was determined by ELISA as described in “Materials and Methods”. The tumor cell was assumed to travel in a shear flow near the EC substrate; while a PMN was assumed to be rolling on the EC in the downstream. Simulation started when a tumor cell and a PMN were 1 cm apart. The concentration of IL-8 secreted by the moving tumor cell near the rolling PMN was derived by using Comsol Multiphysics simulation under various shear flow conditions.
Results indicate that altering the shear rate affects the IL-8 concentration near a rolling PMN at different time points (Fig. 7). When the shear rate was constant at 62.5 s−1, the IL-8 concentration near a rolling PMN and the stimulation time remained constant under various shear stresses (Fig. 7a). In contrast, when the shear stress was constant at 2 dyn cm−2 and shear rate increased from 62.5 to 200 s−1, it took less time to activate the PMNs under a higher shear rate (Fig. 7b). These data suggest that the fluid convection affects the local concentration of IL-8 near a downstream PMN as well as the time needed to activate the PMN, which is important in upregulating the expression of LFA-1 and Mac-1.
FIGURE 7.

The concentration of IL-8 near PMN and the time it activates PMN under different shear flow conditions. Comsol Multiphysics was used to simulate the diffusion of IL-8 as described in “Materials and Methods”, and then the concentration of IL-8 near the PMN was derived. The activation time of IL-8 on the PMN was determined by the time necessary for the melanoma cell to reach the PMN. (a) IL-8 concentration near a rolling PMN and the time it activated the PMN was very similar under same shear rate but different shear stresses; (b) Higher shear rate led to faster melanoma cell velocities, resulting in shorter stimulation times on PMN under constant shear stress but different shear rates.
The changes of LFA-1 and Mac-1 expression on a rolling PMN upon stimulation from IL-8 secreted by a tumor cell in a shear flow were then calculated (Fig. 8). Results indicate that at a constant shear rate, 62.5 s−1, varying the shear stress does not alter the expressions of LFA-1 and Mac-1 on PMNs. However, at a constant shear stress, 2 dyn cm−2, LFA-1 and Mac-1 expressions on PMNs were reduced when shear rate is increased.
FIGURE 8.

LFA-1 and Mac-1 expressions upon activation of IL-8 from tumor cell under different shear flow conditions. LFA-1 and Mac-1 site densities on the PMN under various shear conditions were derived using the site density expression data as exemplified in Fig. 3 and the simulation data shown in Fig. 7. (a) LFA-1 expression; (b) Mac-1 expression. At the same shear rate, LFA-1 and Mac-1 expressions on PMN were not significantly different although the shear stresses were different. In contrast, when the shear stress was the same, the increase of shear rates reduced LFA-1 and Mac-1 expressions.
Melanoma–PMN Binding Association Rate is Affected by the Shear Rate
To quantify the increase in binding potential when a PMN is stimulated by IL-8 secreted from a melanoma cell, the association rate governing the binding of the cells was calculated. The likelihood of binding between the two cells is determined by the number and availability of adhesion molecules on both cells, as well as the intrinsic binding properties of the molecules. Equation (6) was used to calculate the association rate for an LFA-1 and a Mac-1 molecule binding with an ICAM-1 molecule when a melanoma cell is 1 μm from a rolling PMN under various shear conditions. The surface densities of LFA-1 and Mac-1 molecules on a PMN stimulated by melanoma cell-secreted IL-8 (Fig. 8) were used for nL. A constant separation distance and contact area were assumed for the six shear flow conditions.
The association rates indicate that the likelihood of an LFA-1/ICAM-1 bond forming between a melanoma cell and PMN is almost doubled when the shear rate is decreased from 200 to 62.5 s−1 and the shear stress is considered constant (Fig. 9). For Mac-1/ICAM-1 binding, the likelihood increases by more than twice when the shear rate is decreased. When the shear rate remains constant, however, and the shear stress is increased, the binding potential for both molecules remains constant.
FIGURE 9.

Association-rates for LFA-1 and Mac-1 binding to ICAM-1 when the molecules are up-regulated due to IL-8 stimulation from an upstream melanoma cell. (a) LFA-1/ICAM-1 binding association-rate; (b) Mac-1/ICAM-1 binding association-rate. Both the LFA-1/ICAM-1 and Mac-1/ICAM-1 association-rates are dependent on the shear rate and not the shear stress of the flow.
Bond Force Between Melanoma Cell and PMN Under Different Shear Conditions
The dissociation of β2 integrin bonds with ICAM-1 is governed by the force applied to them.3 In order to compare the affect the shear rate and shear stress have on the bond dissociation, a single bond was examined under various flow conditions. A numerical model was used to simulate a melanoma cell and a stationary PMN to determine the peak force applied to the melanoma cell due to a single bond between the cells under various shear conditions. In all simulations, the melanoma cell was initially located above and slightly downstream of the stationary PMN (Fig. 10) and then was allowed to move freely at its steady state velocity for each flow condition. The melanoma cell was acted upon by the fluid, bond, and repulsion forces until a maximum bond force was reached. Although adhesion between a melanoma cell and a PMN may be mediated by more than a single bond, the trend in bond dissociation over various flow conditions is expected to be similar for all bonds.
FIGURE 10.
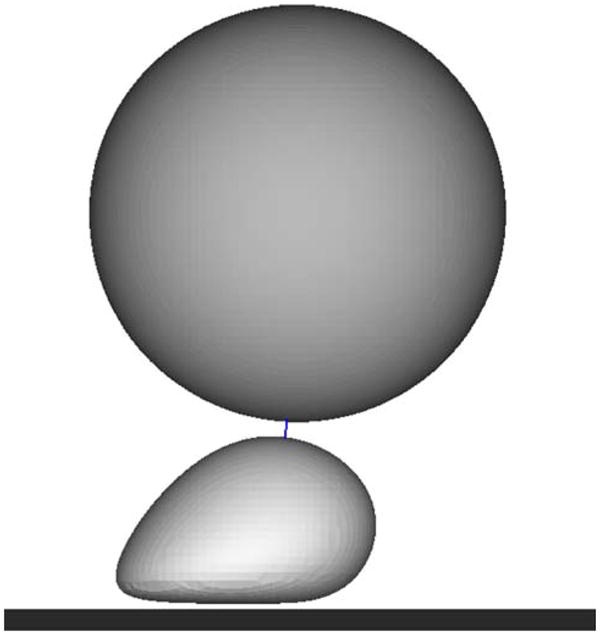
Melanoma cell bound to a stationary PMN via a single bond. Initial configuration of the cells, with a line between them to represent the location of the bond.
Results indicate that increasing the shear rate from 62.5 to 200 s−1, while maintaining a constant shear stress, increased the maximum bond force by approximately 11%, but increased the rate at which it was reached by almost four times (Fig. 11). Increasing the shear stress from 0.625 to 2 dyn cm−2, while keeping a constant shear rate, increased the maximum bond force by almost twice, and decreased the rate which it was reached by almost five times. Together, these results suggest that the dissociation of bonds between the melanoma cell and PMN is governed by both the shear stress and the shear rate.
FIGURE 11.

Force on a melanoma cell due to a single bond with a stationary PMN under various shear conditions. Melanoma cell motion was simulated numerically due to the action of a single bond with a PMN. (a) Under the same shear rate, at higher shear stress (due to higher viscosity), the bond force peaks slower, but at a greater force than at lower shear stress; (b) Under the same shear stress, at a higher shear rate, the bond force reaches a maximum faster and at a greater force than at lower shear rates. These results indicate the maximum force applied to the melanoma cell due to the bond and the time it takes to reach it is dependent on both shear rate and shear stress.
DISCUSSION
Recent studies have indicated that PMN-facilitated melanoma adhesion to the EC is shear rate dependent and is influenced by endogenous IL-8 chemokine, either liberated from melanoma cells or from PMNs after stimulated by melanoma cells.12,13,19 However, little is known regarding the shear flow-regulated mechanisms involved in this interaction. In this study, we have investigated the effects of: (1) the chemokine IL-8 on the PMN surface density of β2 integrins, (2) the transport of IL-8 from a melanoma cell to a PMN under various flow conditions, (3) increased β2 integrin surface density on the likelihood of binding between a melanoma cell and a PMN, and (4) various flow conditions on the force applied to a melanoma cell by a single bond with a stationary PMN.
Chemokines, which comprise the largest family of cytokines, are small secreted proteins that regulate leukocyte transport by mediating the adhesion of leukocytes to endothelial cells, which is the initial step in transendothelial migration and tissue invasion.30 Specifically, the up-regulation of LFA-1 and Mac-1 upon stimulation by IL-8 has been reported.1,4 In agreement with those studies, we found that the stimulation of PMNs using recombinant IL-8 increased the expression of LFA-1 and Mac-1 on PMNs. Moreover, the up-regulation was both time and dose dependent. There existed both a threshold and a saturating concentration of IL-8 in stimulating LFA-1 and Mac-1 on PMNs. Reports from other groups have supported this finding. For example, Lum et al.14 found a threshold dose of 75 pM IL-8 in activating PMNs to adhere to the ICAM-1 coated beads. Seo et al.21 observed that an increase in IL-8 concentration from 1 to 5 nM did not induce significant changes in PMNs adhesion to ICAM-1 coated beads. LFA-1 and Mac-1 expressions also respond to stimulation by IL-8 in different ways. The response of LFA-1 upon IL-8 stimulation is rapid, while Mac-1 responds to the stimulation in a later time period. This behavior is consistent with studies that have shown that LFA-1 is important for rapid binding between cells, while Mac-1 is important for the maintenance of adhesion.18
One important mode of intercellular communication in cancer hematogenous metastasis occurs through the soluble chemokines released by involved cells, which may be affected by hydrodynamic shear flow. Melanoma cells constitutively secrete the inflammatory chemokine IL-8.1,20 When in a shear flow, this IL-8 is transported and stimulates other cells in the flow. A numerical model was employed to investigate the effects of the transported IL-8 on PMNs in a shear flow. Results show that shear rate, rather than shear stress, affects the IL-8 concentration near a rolling PMN at different time points. Thus, the surface density of LFA-1 and Mac-1 on the stimulated PMN is also shear rate dependent. The binding potential of IL-8 stimulated PMNs (mediated by both LFA-1/ICAM-1 and Mac-1/ICAM-1 binding) followed the same shear rate dependent trend, which implies the transport of IL-8 to a rolling PMN contributes to the shear rate dependence of melanoma cell adhesion to the EC in the presence of PMNs in a hydrodynamic shear flow.
The dissociation of melanoma cells from PMNs is determined by the force applied to the intercellular bonds. In the numerical simulations presented, the force applied to a single bond between the cells was modeled in order to directly compare the results under various flow conditions. Though cell adhesion is expected to be mediated by more than one bond, the trend of bond dissociation is expected to be similar to the trend seen in the single bond situation. Thus, we can make a general conclusion on the trend in bond dissociation between the cells based on the numerical simulations.
Changing either the shear rate or the shear stress altered the trend of the force applied to the melanoma cell by a single bond. This suggests that the dissociation of the bonds between a melanoma cell and a PMN is dependent on both the shear rate and shear stress. Since the melanoma cell–PMN aggregation through β2 integrins and ICAM-1 is affected by shear rate, but the dissociation of bonds between the cells is dependent on both the shear rate and shear stress, it is probable that the formation of bonds between the two cells plays a more important role in determining the aggregation potential than the bond dissociation.
Acknowledgments
The authors thank Dr. Scott Simon (UC Davis, CA) for providing EI cells and Dr. Meenhard Herlyn (Wistar Institute, Philadelphia, PA) for providing WM9 cells. This work was supported by the National Institutes of Health grant CA-97306, CA-125707 and National Science Foundation grant CBET-0729091. The authors acknowledge and appreciate complimentary software licenses provided to the authors by CEI, Inc. (Harpoon and Ensight) and ACUSIM Software, Inc. (AcuSolve). The authors appreciate the support of the Pennsylvania State University Applied Research Laboratory and Penn State General Clinical Research Center (GCRC), the sta3 that provided nursing care. The GCRC is supported by NIH Grant M01-RR-10732.
References
- 1.Baggiolini M, Dewald B, Moser B. Interleukin-8 and related chemotactic cytokines—CXC and CC chemokines. Adv Immunol. 1994;55:97–179. [PubMed] [Google Scholar]
- 2.Bell GI. Models for the specific adhesion of cells to cells. Science. 1978;200:618–627. doi: 10.1126/science.347575. [DOI] [PubMed] [Google Scholar]
- 3.Dembo M, Torney DC, Saxaman K, Hammer D. The reaction-limited kinetics of membrane-to surface adhesion and detachment. Proc R Soc Lond. 1988;234:55–83. doi: 10.1098/rspb.1988.0038. [DOI] [PubMed] [Google Scholar]
- 4.Diamond MS, Springer TA. The dynamic regulation of integrin adhesiveness. Curr Biol. 1994;4:506–507. doi: 10.1016/s0960-9822(00)00111-1. [DOI] [PubMed] [Google Scholar]
- 5.Gopalan P, Smith C, Lu H, Berg E, Mcintire L, Simon SI. PMN CD18-dependent arrest on ICAM-1 in shear flow can be activated through L-selectin. J Immunol. 1997;158:367–375. [PubMed] [Google Scholar]
- 6.Holmes WE, Lee J, Kuang WJ, Rice GC, Wood WI. Structure and functional expression of human interleukin-8 receptor. Science. 1991;253:1278–1280. doi: 10.1126/science.1840701. [DOI] [PubMed] [Google Scholar]
- 7.Hoskins MH, Dong C. Kinetics analysis of binding between melanoma cells and neutrophils. Mole Cell Biomech. 2006;3:79–87. [PMC free article] [PubMed] [Google Scholar]
- 8.Hoskins MH, Kunz RF, Bistline JE, Dong C. Coupled flow-structure-biochemistry simulations of dynamic systems of blood cells using an adaptive surface tracking method. 2008 doi: 10.1016/j.jfluidstructs.2009.02.002. (submitted for review) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Jadhav S, Eggleton CD, Konstantopoulos K. A 3-D computational model predicts that cell deformation affects selectin-mediated leukocyte rolling. Biophys J. 2005;88:96–104. doi: 10.1529/biophysj.104.051029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kooijman HA. A modification of the Stokes–Einstein equation for diffusivities in dilute binary mixtures. Ind Eng Chem Res. 2002;41:3326–3328. [Google Scholar]
- 11.Liang S, Sharma A, Peng H, Robertson GP, Dong C. Targeting mutant (V600E) B-Raf in melanoma interrupts immunoediting of leukocyte functions and melanoma extravasation. Cancer Res. 2007;67:5814–5820. doi: 10.1158/0008-5472.CAN-06-4233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Liang S, Slattery M, Dong C. Shear stress and shear rate differentially affect the multi-step process of leukocyte-facilitated melanoma adhesion. Exp Cell Res. 2005;310:282–292. doi: 10.1016/j.yexcr.2005.07.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Liang S, Slattery M, Wagner D, Simon SI, Dong C. Hydrodynamic shear rate regulates melanoma–leukocyte aggregations, melanoma adhesion to the endothelium and subsequent extravasation. Ann Biomed Eng. 2008;36:661–671. doi: 10.1007/s10439-008-9445-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lum AFH, Green CE, Lee GR, Staunton DE, Simon SI. Dynamic regulation of LFA-1 activation and neutrophil arrest on intercellular adhesion molecule 1 (ICAM-1) in shear flow. J Biol Chem. 2002;277:20660–20670. doi: 10.1074/jbc.M202223200. [DOI] [PubMed] [Google Scholar]
- 15.Moghe PV, Nelson RD, Tranquillo RT. Cytokine-stimulated chemotaxis of human neutrophils in a 3-D conjoined fibrin gel assay. J Immunol Meth. 1995;180:193–211. doi: 10.1016/0022-1759(94)00314-m. [DOI] [PubMed] [Google Scholar]
- 16.Muller A, Homey B, Soto H, Ge N, Catron D, Buchanan EM, McClanahan T, Murphy E, Yuan W, Wagner NS, Barrera JL, Mohar A, Verástegui E, Zlotnik A. Involvement of chemokine receptors in breast cancer metastasis. Nature. 2001;410:50–56. doi: 10.1038/35065016. [DOI] [PubMed] [Google Scholar]
- 17.Murphy PM, Tiffany HL. Cloning of complementary DNA encoding a functional human interleukin-8 receptor. Science. 1991;253:1280–1283. doi: 10.1126/science.1891716. [DOI] [PubMed] [Google Scholar]
- 18.Neelamegham S, Taylor AD, Burns AR, Smith CW, Simon SI. Hydrodynamic shear shows distinct roles for LFA-1 and MAC-1 in neutrophil adhesion to intercellular adhesion molecule-1. Blood. 1998;92:1626–1638. [PubMed] [Google Scholar]
- 19.Peng H, Liang S, Henderson AJ, Dong C. Regulation of interleukin-8 expression in melanoma-stimulated neutrophil inflammatory response. Exp Cell Res. 2007;313:551–559. doi: 10.1016/j.yexcr.2006.10.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Schadendorf D, Moller A, Algermissen B, Worm M, Sticherling M, Czarnetzki BM. IL-8 produced by human malignant melanoma cells in vitro is an essential autocrine growth factor. Arch Dermatol Res. 1993;151:2667–2675. [PubMed] [Google Scholar]
- 21.Seo SM, Mcintire LV, Smith CW. Effects of IL-8, Groa, and LTB4 on the adhesive kinetics of LFA-1 and Mac-1 on human neutrophils. Am J Physiol Cell Physiol. 2001;281:C1568–C1578. doi: 10.1152/ajpcell.2001.281.5.C1568. [DOI] [PubMed] [Google Scholar]
- 22.Sharma A, Tran MA, Liang S, Sharma AK, Amin S, Smith CD, Dong C, Robertson GP. Targeting Mek in the mutant (V600E) B-Raf signaling cascade effectively inhibits melanoma lung metastases. Cancer Res. 2006;66:8200–8209. doi: 10.1158/0008-5472.CAN-06-0809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shevde LA, Welch DR. Metastasis suppressor pathways—an evolving paradigm. Cancer Lett. 2003;198:1–20. doi: 10.1016/s0304-3835(03)00304-5. [DOI] [PubMed] [Google Scholar]
- 24.Simon SI, Green CE. Molecular mechanics and dynamics of leukocyte recruitment during inflammation. Annu Rev Biomed Eng. 2005;7:151–185. doi: 10.1146/annurev.bioeng.7.060804.100423. [DOI] [PubMed] [Google Scholar]
- 25.Slattery M, Dong C. Neutrophils influence melanoma adhesion and migration under flow conditions. Intl J Cancer. 2003;106:713–722. doi: 10.1002/ijc.11297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Slattery M, Liang S, Dong C. Distinct role of hydrodynamic shear in leukocyte-facilitated tumor cell extravasation. Am J Physiol Cell Physiol. 2005;288:C831–C839. doi: 10.1152/ajpcell.00439.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Soengas MS, Lowe SW. Apoptosis and melanoma chemoresistance. Oncogene. 2003;22:3138–3151. doi: 10.1038/sj.onc.1206454. [DOI] [PubMed] [Google Scholar]
- 28.You J, Mastro AM, Dong C. Application of the dual micropipette technique to the measurement of tumor cell locomotion. Exp Cell Res. 1999;248:160–171. doi: 10.1006/excr.1999.4388. [DOI] [PubMed] [Google Scholar]
- 29.Young ME, Carroad PA, Bell RL. Estimation of diffusion coefficients of proteins. Biotechnol Bioeng. 1980;22:947–955. [Google Scholar]
- 30.Zlotnik A, Yoshie O. Chemokines: a new classification system and their immunity. Immunity. 2000;12:121–127. doi: 10.1016/s1074-7613(00)80165-x. [DOI] [PubMed] [Google Scholar]