

Abstract
Endocannabinoids are part of a bioactive lipid signaling system, not only in the central nervous system but also in various peripheral organs. Accumulating evidence implicates dysregulation of the endocannabinoid system (ECS) in the pathogenesis of various cardiovascular diseases, including hypertension, atherosclerosis, myocardial infarction, hemorrhagic or septic shock, heart failure and cardiovascular complications of liver cirrhosis. Even though the benefit of chronic cannabinoid 1 (CB1) receptor blockade with the currently available compounds may not outweigh the risks in chronic conditions such as obesity, modulation of the ECS may hold great therapeutic promise in various cardiovascular conditions/disorders.
In this review we will discuss recent advances in understanding the role of CB1 receptors and endocannabinoids in the regulation of cardiac function in cirrhotic cardiomyopathy and in doxorubicin-induced heart failure.
Keywords: cannabinoids, cardiovascular disease, cardiac function, heart failure, cardiomyopathy, cirrhosis, doxorubicin
Introduction
Endogenous cannabinoids may exert a wide range of biological effects in a variety of physiological and pathological conditions (1, 2). Initially identified in the brain, these bioactive lipids are part of a novel signaling system along with their G-protein coupled receptors and the enzymes involved in their biosynthesis and degradation. The growing availability of pharmacological tools and knockout mouse strains has been crucial in uncovering the biological functions that are modulated by the endocannabinoid system (ECS) under normal and pathological conditions (1, 3). Arachidonoyl ethanolamine or anandamide (AEA) and 2-arachidonoylglycerol (2-AG), are the two most studied endocannabinoids with well characterized cardiovascular effects. In contrast, the cardiovascular effects of other putative endocannabinoid ligands, such as the 2-arachidonyl-glyceryl ether (noladin ether), O-arachidonoylethanolamide (virhodamine) and N-arachidonyl-dopamine are lesser known (1). AEA and 2-AG are generated from membrane glycerophospholipids by distinct biosynthetic pathways in response to a rise in intracellular calcium and/or stimulation of metabotropic receptors (4). AEA is metabolized predominantly by fatty acid amide hydrolase (FAAH) and 2-AG is preferentially degraded by monoglyceride lipase (MGL). Endocannabinoids share the pharmacology of THC, the psychoactive ingredient of marijuana (cannabis), and produce similar biological effects. Two types of cannabinoid receptors, CB1 and CB2, have been identified in mammalian tissues. However, based on pharmacological evidence, several other receptors as well as non-receptor mediated pathways have also been implicated in the effects of endocannabinoids (1, 5, 6). CB1 is the most abundant G-protein coupled receptor in the brain but it has been identified at lower, yet functionally relevant levels in various peripheral tissues (e.g. adipose tissue (7, 8), liver (9, 10), myocardium (11-14), and the vascular endothelium and smooth muscle cells (15-17). CB2 receptors are expressed primarily in cells of immune and hematopoietic origin, but their presence has recently been also confirmed in the brain (18), liver (9), myocardium (13, 19), vascular endothelial and smooth muscle cells (16). Both receptors are coupled to Gi/o, inhibiting adenylate cyclase, but they can also signal through other pathways (e.g. cyclooxygenase-2, mitogen-activated protein kinases and protein kinases A and C, just to mention a few (1, 20). Therapeutic modulation of the ECS may be beneficial in various cardiovascular disorders, including hypertension, myocardial infarction, circulatory shock, cirrhotic cardiomyopathy and heart failure, which have been covered in recent overviews (21, 22). In this review we will focus on recent advances in understanding the role of the CB1 receptor and endocannabinoids in the regulation of cardiac function with special emphasis on the potential benefits of CB1 antagonism in cirrhotic cardiomyopathy and in doxorubicin-induced heart failure.
Endocannabinoids and the heart
The cardiovascular effects of cannabinoid compounds have been known for a long time. Studies in humans indicated that chronic use of marijuana causes long lasting decrease in blood pressure and heart rate, whereas the acute use increases heart rate without affecting blood pressure (14, 23). In anesthetized animals, Δ9-tetrahydrocannabidiol (THC), its synthetic analog HU-210, and anandamide decrease blood pressure and cardiac contractility and cause bradycardia through the activation of CB1 receptors (14). Cardiovascular functions are regulated by complex central and local mechanisms; the ECS recently emerged as a potent cardiovascular modulator mechanism that may become particularly relevant under disease conditions. Cannabinoid receptors, located centrally and on presynaptic autonomic nerve terminals may inhibit neurotransmitter release, thereby modulating the sympathetic outflow, which maintains constant control of cardiovascular function (24, 25). At the peripheral level, functional CB1 receptors in the vessel wall appear to be involved in certain tissue- and species-specific vasodilatory actions of cannabinoids, however these effects may also be mediated by other receptors (such as TRPV1, GPR55, and yet unidentified ones) (5, 14, 26, 27). Myocardial CB1 receptors mediate negative inotropy in papillary muscles in vitro, in isolated hearts ex vivo, as well as in anesthetized rodents in vivo (14, 28) (Figure 1.). The latter evidence comes from recent studies in which the hypotensive effect of AEA has been found to be predominantly mediated by the decrease in cardiac contractility through CB1 receptors (11, 29, 30). These effects are less pronounced in conscious cannulated animals (31). In addition to the above-discussed actions, endocannabinoids are rapidly cleaved in vivo by hydrolysis, and cycloxygenase-2, lipoxygenases and cytochrom P450-mediated oxidative metabolism. The rapidly generated bioactive derivatives, such as prostanoids, exert additional cardiovascular and other biological effects that may be confounding (32-34).
Figure 1.
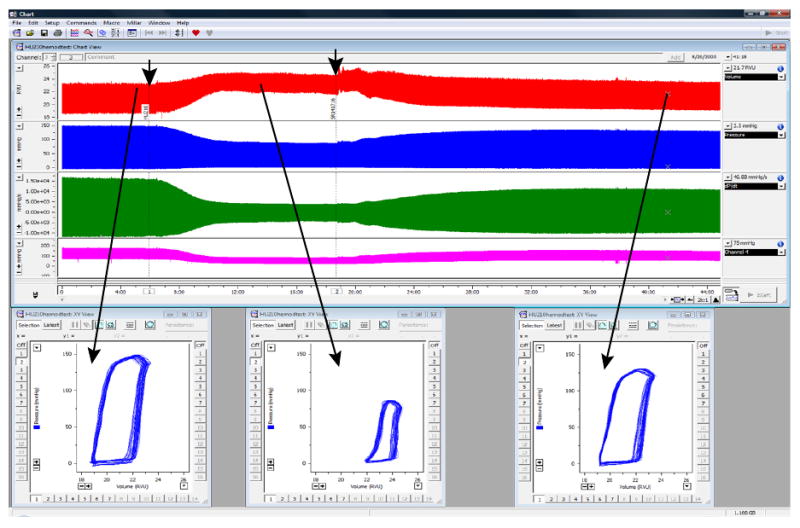
Characteristic changes in rat LV volume (red trace) and pressure (blue trace), +dP/dt and -dP/dt derived from LV pressure signal (green trace), and arterial pressure (purple trace) before (at baseline) or after an administration of a drug with known cardiodepressive properties (a cannabinoid type 1 (CB1) receptor agonist HU210), followed by the recovery after the administration of the CB1 antagonist SR141716 (drug administrations are indicated by arrows). Lower panels show characteristic PV loops at baseline and following the drug administrations. Note that even without any calibrations, the volume traces and PV relationships are very informative. (Used with permission (28))
The available evidence suggests that endocannabinoids play only limited, if any role in the regulation of the cardiovascular system under normal conditions in healthy individuals. Administration of CB1 antagonists does not affect blood pressure, heart rate or cardiac function in normotensive rodents (11, 13, 35) or in humans. In obese but normotensive patients one-year treatment with CB1 antagonist rimonabant slightly but significantly decreased both systolic and diastolic blood pressure, presumably as a result of the concomitant body weight loss (36).
On the other hand, activation of the ECS, which may result from an imbalance between endocannabinoid biosynthesis and degradation and/or from changes at the level of receptors and signaling pathways, has been associated with various pathological cardiovascular conditions associated with decreased myocardial function.
The endocannabinoid system in advanced hepatic cirrhosis
Hepatic cirrhosis, the end stage of chronic liver disease, is associated with impaired hepatic function, portal hypertension and significant resultant morbidity and mortality. The underlying pathomechanism is the progressive replacement of the liver tissue by fibrous scar tissue and by regenerative nodules leading to the disruption of the entire liver architecture. The transformation of the liver parenchyma results in the reorganization of the intrahepatic vasculature. The pathological features of cirrhosis may be clinically silent for a long time, followed by a period of nonspecific signs. In advanced cirrhosis, the symptoms are related to the progressive development of liver failure and the consequences of portal hypertension. The prognosis for the patients with cirrhosis depends on several factors, such as the degree of hepatic injury, presence of complications of advanced liver disease. When complications cannot further be controlled or the hepatic failure is complete, liver transplantation remains the only therapeutic option. Cardiovascular complications of cirrhosis include hemodynamic changes, abnormalities in the central, splanchnic and peripheral circulation and cardiac dysfunction. Patients with advanced cirrhosis may present characteristic cardiovascular symptoms such as tachycardia, systemic hypotension, increased baseline cardiac output and decreased total vascular resistance, the signs of hyperdynamic circulation (37). In addition to the above-mentioned hemodynamic constellation, cardiac contractile function in patients with advanced cirrhosis is often impaired; a condition also termed cirrhotic cardiomyopathy (38). The signs of cirrhotic cardiomyopathy are often subclinical; they are generally masked by the reduced left ventricular afterload. However, the attenuated ventricular contractile response can be revealed under conditions of physiological or pharmacological stress. These include response to inotropic drugs, physical exercise, or therapeutic interventions such as transjugular intrahepatic portosystemic shunt or liver transplantation. Electrophysiological abnormalities of the heart, such as prolongation of repolarization (increased QT interval), electromechanical uncoupling, and cardiac chamber hypertrophy or enlargement (38, 39) are often present in cirrhotic patients. Cirrhotic cardiomyopathy may contribute to poor outcomes following liver transplantation and may be implicated in the pathogenesis of the hepatorenal syndrome. The findings that the hypotension, induced by endotoxin (40) or associated with hemorrhagic shock can be reversed by CB1 antagonist and endocannabinoid levels are elevated in these conditions (41, 42) led to the hypothesis that the ECS might also be involved in the pathogenesis of cirrhosis. Early studies mainly focused on the peripheral vascular actions of endocannabinoids in cirrhosis. These studies (43, 44) have suggested that an overactivated ECS contributes to the decreased blood pressure in advanced cirrhosis. A single bolus dose of a selective CB1 receptor antagonist reversed the systemic hypotension, decreased portal venous pressure, mesenteric flow (43) and cardiac output; and increased total peripheral resistance and blood pressure in the cirrhotic animal (44). A more recent study investigated the effect of chronic CB1 antagonist treatment in the carbontetrachloride model of rat cirrhosis and found that it dose-dependently improved hemodynamic alterations and resulted in decreased ascites formation in cirrhotic animals (45).
Given that bacterial lipopolysaccharide (LPS) represents a major trigger for elevated endocannabinoid levels, as it has been shown in peripheral lymphocytes (46), monocytes and platelets (47), hepatocytes (48), and possibly in other cell types, and endotoxemia is common in advanced cirrhosis due to the leaky gut syndrome, it can be hypothesized that the elevated circulating levels of LPS stimulate the production or decrease the degradation of anandamide and possibly other endocannabinoids or endocannabinoid-like substances. Indeed, earlier preclinical studies demonstrated that isolated monocytes and platelets from cirrhotic animals, as well as from patients with the condition, had significantly elevated levels of anandamide (43, 44). Furthermore, the injection of these cells into normal recipient animals elicited CB1 receptor sensitive hypotension. Recent studies have established that endocannabinoid levels are elevated in patients with advanced cirrhosis compared to healthy volunteers (49, 50). However, the clinical significance of these findings remains to be explored.
These studies, however, did not address the question whether the activated ECS has a differential effect on target organs and vascular beds. One of the first documented peripheral effects of anandamide was its vasodilator action mediated by CB1, TRPV1, nonCB1/CB2 receptors, via endothelium and/or nitric oxide-dependent and/or -independent mechanisms (14, 29, 35, 51). As already mentioned above, the cellular distribution of CB1 receptor is altered in cirrhosis. A marked increase in CB1 receptor expression was found in human cirrhotic liver endothelial cells (43) and liver fibrogenic stellate cells (52), as well as in mesenteric arteries from cirrhotic rats (53) and in cirrhotic rat liver samples (54). Experiments using isolated vessels provided functional evidence that cirrhosis is associated with altered vascular function. Isolated mesenteric arteries from cirrhotic animals were more sensitive to the vasorelaxant effects of anandamide than the vessels from control rats. This vasodilator effect was independent from endothelium and nitric oxide and involved CB1 and TRPV1 receptors, the latter presumably located in the perivascular sensory nerve terminals of the mesenteric artery (53).
The initial event in the development of portal hypertension is the increase in intrahepatic resistance caused by the intrahepatic vascular rearrangement in cirrhosis. As opposed to its effect in the mesenteric vasculature, anandamide has been found to contribute to the increased hepatic vasoconstriction in cirrhotic animals (55): CB1 antagonist treatment was able to reverse the increased intrahepatic resistance and portal venous pressure (54). Since cycloxygenase-2 and 5-lipoxygenase expressions were found to be increased in the cirrhotic liver, this hepatic “hyperreactivity” to anandamide may be mediated by the increased vasoactive eicosanoid production. Indeed, there is in vitro and in vivo evidence that cycloxygenase-2 might contribute to the metabolism and cardiovascular effects of anandamide (34).
Apart from the above-mentioned effects, endocannabinoids may modulate apoptosis in various cell types (56). Indeed, studies demonstrated that both anandamide and 2-AG can induce dose-dependent apoptosis in fibrogenic hepatic stellate cells both in vitro and in vivo (57, 58). If so, this effect of the endocannabinoids may have pathogenetic role in cirrhotic fibrogenesis and liver injury and the resultant impairment of intrahepatic microcirculation, leading to the development of portal hypertension. Indeed, CB1 antagonism attenuates liver fibrosis in various animal models (52). Taken together, the porto-hepatic ECS appears to play an important role in the pathogenesis of hemodynamic alterations associated with liver cirrhosis.
Endocannabinoids and the cirrhotic contractile dysfunction
The emergence of in vitro and in vivo studies that demonstrated the negative inotropic effect of endogenous and exogenous cannabinoids in various experimental models suggested the possible pathogenic role of endocannabinoids in contractile dysfunction associated with advanced hepatic cirrhosis. The first indication that CB1 receptors are involved in some aspects of abnormal myocardial contractility in liver cirrhosis came from an in vitro study (59) using isolated left ventricular papillary muscle from bile duct-ligated cirrhotic rats, in which CB1 blockade reversed the decreased β-adrenergic responsiveness (59). A recent study has characterized in vivo left ventricular pressure-volume relationships in rats with carbon tetrachloride-induced advanced micronodular cirrhosis and provided additional evidence on the involvement of the ECS in the pathogenesis of hemodynamic alterations associated with hepatic cirrhosis (60). The cardiac contractile dysfunction was evident by the marked changes in both load-dependent and load-independent indices of myocardial contractility (Figure 2.). Interestingly, cardiac index was increased in cirrhotic, compared with normal rats, and was unaffected by CB1 blockade with CB1 antagonist AM251, in agreement with previous reports in cirrhotic rats (61). The maximum first derivative of ventricular pressure with respect to time (+dP/dt), an indicator of left ventricular contractility, was markedly reduced in cirrhosis. Since the reduced cardiac contractile function in cirrhotic rats was likely offset by increased venous return and decreased peripheral resistance, load independent parameters of contractile function were also evaluated, such as the slope of the end systolic pressure-volume ratio (ESPVR), Emax and preload recruitable stroke work (PRSW), in order to uncover contractile impairment in advanced cirrhosis. ESPVR and Emax were markedly decreased in cirrhotic rats and increased toward control levels following CB1 blockade (60). The marked decrease in these functional indices in cirrhotic rats and their return to near control levels following AM251 treatment clearly demonstrated that endocannabinoids are directly involved in systolic dysfunction through directly affecting myocardial contractility of the cirrhotic heart. Parameters of diastolic function, such as the end-diastolic pressure and τ were increased and the maximum first derivative of ventricular pressure with respect to time was decreased in cirrhosis, which could reflect impaired diastolic relaxation, an active process reflecting myocardial calcium sequestration. However, end-diastolic pressure may also be influenced by changes in preload, and its increase may have resulted from increased venous return related to the low peripheral resistance in cirrhosis. The finding that the load-independent measure of stiffness, EDPVR, was unaffected in hepatic cirrhosis suggests the absence of significant myocardial fibrosis, and that the observed diastolic dysfunction is rather related to impaired inotropy. In addition to such pharmacological evidence, myocardial and liver AEA and/or 2-AG levels were elevated in cirrhotic animals (60), implicating tissue specific overactivation of the ECS in the pathogenesis of liver cirrhosis. Taken together, these findings suggest that the increased activity of the endocannabinoid/CB1-receptor system is largely responsible for the impaired myocardial contractile function associated with hepatic cirrhosis in experimental disease models. Importantly, this cardiac dysfunction could be corrected by CB1 blockade, highlighting the therapeutic potential of CB1 antagonists in this disease condition.
Figure 2.

Effects of AM251 on load-independent contractile parameters. A: Representative pressure-volume (PV) loops obtained with a P-V conductance catheter system at different preloads. Note that the slope of the end-systolic P-V relation (ESPVR, dotted lines) is less steep in cirrhotic compared to control animals, indicating decreased contractile function, and is acutely increased following treatment of the cirrhotic rats with 3 mg/kg AM251. B: Effect of AM251 on load-independent contractile parameters measured as Emax, ESPVR, PRSW for systolic and EDPVR for diastolic function. Control and cirrhotic animals were treated with vehicle (white or gray columns, respectively) or 3 mg/kg AM251 (black columns). P<0.05 control vs cirrhotic (*) or vehicle vs. AM251 value (#). N=6-8 rats per group, vertical bars indicate SEM. (Used with permission (59))
Endocannabinoids in myocardial injury and dysfunction
It has already been stated that the ECS is activated in conditions associated with decreased blood pressure and cardiac function, such as hemorrhagic, septic and cardiogenic shock (reviewed in (1)) and in advanced liver cirrhosis, as discussed above. In addition to the well established role of myocardial CB1 receptors in negative inotropy, and the role of elevated levels of endocannabinoids in CB1 receptor-mediated hypotension and decreased myocardial contractility, endocannabinoids may also be involved in the pathogenesis of acute myocardial impairment and the subsequent ventricular dysfunction. In earlier studies, endocannabinoids appeared to be cardioprotective, limiting myocardial damage and dysfunction through preconditioning in models of ischemia-reperfusion (62-65). In most of these studies, endocannabinoid levels were not measured and the conclusion was based mainly on the use of non-selective pharmacological tools and ex-vivo perfused model systems, which cannot address the potential important interactions between the ECS and the local and systemic inflammatory elements (64). In some of these studies the observed protective effects were CB receptor dependent, but not in others (64).
Earlier in vitro studies provided conflicting results regarding the role of endocannabinoids and CB1 in the pathogenesis of myocardial injury and myocardial remodeling (66, 67). Indeed, chronic CB1 antagonist treatment was found to promote cardiac remodeling while cardiac function, but not the survival, was improved (67). In the same model HU-210, a non-selective cannabinoid receptor agonist (stronger agonist on CB1), was shown to prevent endothelial dysfunction, however it increased the left ventricular end-diastolic pressure, which are conflicting findings. However, the limitations of the above mentioned studies are the insufficient dose of CB1 antagonists used, and the high dose of HU-210, which is known to be associated with psychotropic effects and centrally mediated hypothermia. In a separate study, HU-210 decreased the incidence of ventricular arrhythmias (68). Another nonselective CB receptor agonist, WIN-55212,2 significantly reduced infarct size in an in vivo mouse model of ischemia, where the protective effect was mediated presumably by CB2 rather than CB1 receptors (69). A recent in vivo study provided further evidence for the protective effect of CB2 receptor activation against myocardial ischemia/reperfusion injury (19). This is consistent with the results previously observed in a hepatic model of I/R injury (48).
Based on these studies, it can be concluded that the ECS appears to be activated during myocardial damage, however its precise role is still elusive (for more detailed review on this subject see (64, 70).
The endocannabinoid system in doxorubicin induced heart failure
Doxorubicin is one of the most effective anticancer drugs available. This anthracycline antibiotic is frequently used to treat breast cancer, childhood solid tumors, soft tissue sarcomas and lymphomas (71). Unfortunately, the use of doxorubicin is associated with cardiotoxicity, limiting its clinical use. Doxorubicin may induce cardiomyopathy that leads to the development of heart failure (72). The pathomechanism of doxorubicin-induced cardiotoxicity is multifaceted. Most studies are in agreement that it involves oxidative and nitrosative stress, increased generation of reactive oxygen and nitrogen species and consequent tissue injury. It also involves increased nitric oxide production via the activation of the inducible nitric oxide synthase, and increased peroxynitrite formation through the diffusion-limited reaction of superoxide and NO (73, 74). Peroxynitrite is capable of modifying and inactivating key proteins involved in various cellular functions (75). Other possible mechanisms include the activation of various mitogen activated protein kinases and inflammatory pathways, dysregulation of iron and intracellular calcium homeostasis, alterations in mitochondrial respiratory chain function and β-adrenergic receptor signaling, eventually leading to cell death by necrosis or apoptosis (72, 73, 76-78).
A recent study evaluated the role of the ECS in myocardial injury and subsequent ventricular dysfunction, induced by doxorubicin (13), using a well-established mouse model of doxorubicin-induced cardiomyopathy/heart failure (13, 73, 79). Mice develop severe cardiomyopathy 5 days after a single intraperitoneal injection of doxorubicin, with characteristic severe depression of left ventricular systolic pressure, +dP/dt, cardiac output, stroke work, ejection fraction (Figure 3.). The load-independent parameters of left ventricular contractile function (ESPVR, PRSW, +dP/dt-end diastolic volume relation) also indicated severe myocardial contractile impairment. At the cellular level, doxorubicin induced apoptosis as well as cardiomyocyte necrosis. Injections of the CB1 antagonist rimonabant or AM281 significantly attenuated the left ventricular dysfunction caused by doxorubicin (13). Treatment with CB1 antagonists also prevented doxorubicin-induced cell death in vivo and in cultured cardiomyocytes. Indeed, others (80, 81) have confirmed the anti-apoptotic properties of the selective CB1 antagonist rimonabant. In contrast, the CB1 agonist HU-210 by itself induced cell death in cardiomyocytes (13).
Figure 3.

Effect of CB1 antagonists on DOX-induced depression of load-independent indexes of cardiac contractility. A) Representative P-V loops obtained with a P-V conductance catheter system at different preloads following vena cava occlusion, showing differences in the end-systolic P-V relation (ESPVR) and between mice pretreated with vehicle, rimonabant or AM281 and treated with vehicle or DOX. The less steep ESPVR in DOX-treated mice indicates decreased contractile function, which was improved by CB1 antagonists. B) Effects CB1 antagonists on DOX-induced depression of load-independent indexes of cardiac contractility. Results are mean±SEM of 9 to 18 experiments in each group. *P<0.05 vs vehicle; #P<0.05 vs DOX. PRSW: preloadrecruitable stroke work; dP/dt–end diastolic volume (EDV) relation (dP/d –EDV); ESPVR or Emax: end-systolic pressure–volume relation. (Used with permission (13))
The CB1-selective nature of this effect was further supported by the absence of effects of CB2 agonists/antagonists on doxorubicin-induced cell death in vitro. In addition, doxorubicin treatment was associated with elevated levels of AEA in the myocardium, indicating an overactivation of the ECS in vivo, likewise in cultured cardiomyocytes in vitro. The myocardial CB1 and CB2 receptor expression remained unchanged in vivo, as well as in cells exposed to doxorubicin in vitro. These findings indicate that the activated ECS contributes to the myocardial injury and functional impairment following doxorubicin administration. Further investigation of the signaling pathways associated with doxorubicin-endocannabinoid interaction will be required. These findings strongly suggest that CB1 blockade may represent a novel cardioprotective strategy against doxorubicin, or perhaps other cardiac injuries involving cell death (both apoptotic and necrotic).
Conclusion
The endocannabinoid research field is rapidly moving forward through recognizing clinically relevant conditions associated with overactivation of the ECS. Such conditions may include impaired cardiac function in various forms of cardiomyopathies and heart failure. Based on published studies, there is growing evidence that CB1 receptor antagonism may be beneficial and, therefore can be therapeutically exploited in these conditions. One possible therapeutic strategy may involve the acute administration of a CB1 antagonist to abrogate the undesirable hemodynamic and other effects of pathologically overproduced endocannabinoids. Another intriguing possibility lies in exploiting the tissue protective effects of CB1 antagonism in various forms of acute tissue injury. Despite the setback with the first class of CB1 antagonists, a better understanding of the role of the ECS in various cardiovascular pathologies, and the lessons learned from the clinical experience with chronic CB1 blockade should ultimately lead to the development of better therapeutic approaches (e.g. peripherally acting or neutral CB1 receptor antagonists, devoid of centrally mediated side effects, e.g. anxiety and depression (82)), to limit myocardial injury and improve cardiac function in a multitude of disparate cardiovascular diseases.
Acknowledgments
This work was supported by the Division of Intramural Clinical and Biological Research, NIAAA, NIH. Authors are indebted to George Kunos for reading the manuscript and for valuable suggestions.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Pacher P, Batkai S, Kunos G. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol Rev. 2006;58:389–462. doi: 10.1124/pr.58.3.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mechoulam R, Fride E, Di Marzo V. Endocannabinoids. Eur J Pharmacol. 1998;359:1–18. doi: 10.1016/s0014-2999(98)00649-9. [DOI] [PubMed] [Google Scholar]
- 3.Di Marzo V. Targeting the endocannabinoid system: to enhance or reduce? Nat Rev Drug Discov. 2008;7:438–455. doi: 10.1038/nrd2553. [DOI] [PubMed] [Google Scholar]
- 4.Liu J, Wang L, Harvey-White J, Huang BX, Kim HY, Luquet S, Palmiter RD, Krystal G, Rai R, Mahadevan A, Razdan RK, Kunos G. Multiple pathways involved in the biosynthesis of anandamide. Neuropharmacology. 2008;54:1–7. doi: 10.1016/j.neuropharm.2007.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Begg M, Pacher P, Batkai S, Osei-Hyiaman D, Offertaler L, Mo FM, Liu J, Kunos G. Evidence for novel cannabinoid receptors. Pharmacol Ther. 2005;106:133–145. doi: 10.1016/j.pharmthera.2004.11.005. [DOI] [PubMed] [Google Scholar]
- 6.Lauckner JE, Jensen JB, Chen HY, Lu HC, Hille B, Mackie K. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc Natl Acad Sci U S A. 2008;105:2699–2704. doi: 10.1073/pnas.0711278105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cota D, Marsicano G, Tschop M, Grubler Y, Flachskamm C, Schubert M, Auer D, Yassouridis A, Thone-Reineke C, Ortmann S, Tomassoni F, Cervino C, Nisoli E, Linthorst AC, Pasquali R, Lutz B, Stalla GK, Pagotto U. The endogenous cannabinoid system affects energy balance via central orexigenic drive and peripheral lipogenesis. J Clin Invest. 2003;112:423–431. doi: 10.1172/JCI17725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Engeli S, Bohnke J, Feldpausch M, Gorzelniak K, Janke J, Batkai S, Pacher P, Harvey-White J, Luft FC, Sharma AM, Jordan J. Activation of the peripheral endocannabinoid system in human obesity. Diabetes. 2005;54:2838–2843. doi: 10.2337/diabetes.54.10.2838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mallatand A, Lotersztajn S. Endocannabinoids and liver disease. I. Endocannabinoids and their receptors in the liver. Am J Physiol Gastrointest Liver Physiol. 2008;294:G9–G12. doi: 10.1152/ajpgi.00467.2007. [DOI] [PubMed] [Google Scholar]
- 10.Osei-Hyiaman D, DePetrillo M, Pacher P, Liu J, Radaeva S, Batkai S, Harvey-White J, Mackie K, Offertaler L, Wang L, Kunos G. Endocannabinoid activation at hepatic CB1 receptors stimulates fatty acid synthesis and contributes to diet-induced obesity. J Clin Invest. 2005;115:1298–1305. doi: 10.1172/JCI23057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Batkai S, Pacher P, Osei-Hyiaman D, Radaeva S, Liu J, Harvey-White J, Offertaler L, Mackie K, Rudd MA, Bukoski RD, Kunos G. Endocannabinoids acting at cannabinoid-1 receptors regulate cardiovascular function in hypertension. Circulation. 2004;110:1996–2002. doi: 10.1161/01.CIR.0000143230.23252.D2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bonz A, Laser M, Kullmer S, Kniesch S, Babin-Ebell J, Popp V, Ertl G, Wagner JA. Cannabinoids acting on CB1 receptors decrease contractile performance in human atrial muscle. J Cardiovasc Pharmacol. 2003;41:657–664. doi: 10.1097/00005344-200304000-00020. [DOI] [PubMed] [Google Scholar]
- 13.Mukhopadhyay P, Batkai S, Rajesh M, Czifra N, Harvey-White J, Hasko G, Zsengeller Z, Gerard NP, Liaudet L, Kunos G, Pacher P. Pharmacological inhibition of CB1 cannabinoid receptor protects against doxorubicin-induced cardiotoxicity. J Am Coll Cardiol. 2007;50:528–536. doi: 10.1016/j.jacc.2007.03.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pacher P, Batkai S, Kunos G. Cardiovascular pharmacology of cannabinoids. Handb Exp Pharmacol. 2005:599–625. doi: 10.1007/3-540-26573-2_20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Liu J, Gao B, Mirshahi F, Sanyal AJ, Khanolkar AD, Makriyannis A, Kunos G. Functional CB1 cannabinoid receptors in human vascular endothelial cells. Biochem J. 2000;346(Pt 3):835–840. [PMC free article] [PubMed] [Google Scholar]
- 16.Rajesh M, Mukhopadhyay P, Batkai S, Hasko G, Liaudet L, Huffman JW, Csiszar A, Ungvari Z, Mackie K, Chatterjee S, Pacher P. CB2-receptor stimulation attenuates TNF-alpha-induced human endothelial cell activation, transendothelial migration of monocytes, and monocyte-endothelial adhesion. Am J Physiol Heart Circ Physiol. 2007;293:H2210–2218. doi: 10.1152/ajpheart.00688.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rajesh M, Mukhopadhyay P, Hasko G, Pacher P. Cannabinoid CB1 receptor inhibition decreases vascular smooth muscle migration and proliferation. Biochem Biophys Res Commun. 2008;377:1248–1252. doi: 10.1016/j.bbrc.2008.10.159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Van Sickle MD, Duncan M, Kingsley PJ, Mouihate A, Urbani P, Mackie K, Stella N, Makriyannis A, Piomelli D, Davison JS, Marnett LJ, Di Marzo V, Pittman QJ, Patel KD, Sharkey KA. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science. 2005;310:329–332. doi: 10.1126/science.1115740. [DOI] [PubMed] [Google Scholar]
- 19.Montecucco F, Lenglet S, Braunersreuther V, Burger F, Pelli G, Bertolotto M, Mach F, Steffens S. CB(2) cannabinoid receptor activation is cardioprotective in a mouse model of ischemia/reperfusion. J Mol Cell Cardiol. 2009 doi: 10.1016/j.yjmcc.2008.12.014. [DOI] [PubMed] [Google Scholar]
- 20.Howlett AC. Cannabinoid receptor signaling. Handb Exp Pharmacol. 2005:53–79. doi: 10.1007/3-540-26573-2_2. [DOI] [PubMed] [Google Scholar]
- 21.Mukhopadhyay P, Mohanraj R, Batkai S, Pacher P. CB1 cannabinoid receptor inhibition: promising approach for heart failure? Congest Heart Fail. 2008;14:330–334. doi: 10.1111/j.1751-7133.2008.00016.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Pacher P, Mukhopadhyay P, Mohanraj R, Godlewski G, Batkai S, Kunos G. Modulation of the endocannabinoid system in cardiovascular disease: therapeutic potential and limitations. Hypertension. 2008;52:601–607. doi: 10.1161/HYPERTENSIONAHA.105.063651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gorelick DA, Heishman SJ, Preston KL, Nelson RA, Moolchan ET, Huestis MA. The cannabinoid CB1 receptor antagonist rimonabant attenuates the hypotensive effect of smoked marijuana in male smokers. Am Heart J. 2006;151:754 e751–754 e755. doi: 10.1016/j.ahj.2005.11.006. [DOI] [PubMed] [Google Scholar]
- 24.Niederhofferand N, Szabo B. Effect of the cannabinoid receptor agonist WIN55212-2 on sympathetic cardiovascular regulation. Br J Pharmacol. 1999;126:457–466. doi: 10.1038/sj.bjp.0702337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Varga K, Lake KD, Huangfu D, Guyenet PG, Kunos G. Mechanism of the hypotensive action of anandamide in anesthetized rats. Hypertension. 1996;28:682–686. doi: 10.1161/01.hyp.28.4.682. [DOI] [PubMed] [Google Scholar]
- 26.Johns DG, Behm DJ, Walker DJ, Ao Z, Shapland EM, Daniels DA, Riddick M, Dowell S, Staton PC, Green P, Shabon U, Bao W, Aiyar N, Yue TL, Brown AJ, Morrison AD, Douglas SA. The novel endocannabinoid receptor GPR55 is activated by atypical cannabinoids but does not mediate their vasodilator effects. Br J Pharmacol. 2007;152:825–831. doi: 10.1038/sj.bjp.0707419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zygmunt PM, Petersson J, Andersson DA, Chuang H, Sorgard M, Di Marzo V, Julius D, Hogestatt ED. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature. 1999;400:452–457. doi: 10.1038/22761. [DOI] [PubMed] [Google Scholar]
- 28.Pacher P, Nagayama T, Mukhopadhyay P, Batkai S, Kass DA. Measurement of cardiac function using pressure-volume conductance catheter technique in mice and rats. Nat Protoc. 2008;3:1422–1434. doi: 10.1038/nprot.2008.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pacher P, Batkai S, Kunos G. Haemodynamic profile and responsiveness to anandamide of TRPV1 receptor knock-out mice. J Physiol. 2004;558:647–657. doi: 10.1113/jphysiol.2004.064824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pacher P, Batkai S, Osei-Hyiaman D, Offertaler L, Liu J, Harvey-White J, Brassai A, Jarai Z, Cravatt BF, Kunos G. Hemodynamic profile, responsiveness to anandamide, and baroreflex sensitivity of mice lacking fatty acid amide hydrolase. Am J Physiol Heart Circ Physiol. 2005;289:H533–541. doi: 10.1152/ajpheart.00107.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hoand WS, Gardiner SM. Acute hypertension reveals depressor and vasodilator effects of cannabinoids in conscious rats. Br J Pharmacol. 2009;156:94–104. doi: 10.1111/j.1476-5381.2008.00034.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mendizabaland VE, Adler-Graschinsky E. Cannabinoids as therapeutic agents in cardiovascular disease: a tale of passions and illusions. Br J Pharmacol. 2007;151:427–440. doi: 10.1038/sj.bjp.0707261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Randall MD, Harris D, Kendall DA, Ralevic V. Cardiovascular effects of cannabinoids. Pharmacol Ther. 2002;95:191–202. doi: 10.1016/s0163-7258(02)00258-9. [DOI] [PubMed] [Google Scholar]
- 34.Fowler CJ. The contribution of cyclooxygenase-2 to endocannabinoid metabolism and action. Br J Pharmacol. 2007;152:594–601. doi: 10.1038/sj.bjp.0707379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Batkai S, Pacher P, Jarai Z, Wagner JA, Kunos G. Cannabinoid antagonist SR-141716 inhibits endotoxic hypotension by a cardiac mechanism not involving CB1 or CB2 receptors. Am J Physiol Heart Circ Physiol. 2004;287:H595–600. doi: 10.1152/ajpheart.00184.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ruilope LM, Despres JP, Scheen A, Pi-Sunyer X, Mancia G, Zanchetti A, Van Gaal L. Effect of rimonabant on blood pressure in overweight/obese patients with/without co-morbidities: analysis of pooled RIO study results. J Hypertens. 2008;26:357–367. doi: 10.1097/HJH.0b013e3282f2d625. [DOI] [PubMed] [Google Scholar]
- 37.Kowalskiand HJ, Abelmann WH. The cardiac output at rest in Laennec's cirrhosis. J Clin Invest. 1953;32:1025–1033. doi: 10.1172/JCI102813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Maand Z, Lee SS. Cirrhotic cardiomyopathy: getting to the heart of the matter. Hepatology. 1996;24:451–459. doi: 10.1002/hep.510240226. [DOI] [PubMed] [Google Scholar]
- 39.Gaskari SA, Honar H, Lee SS. Therapy insight: Cirrhotic cardiomyopathy. Nat Clin Pract Gastroenterol Hepatol. 2006;3:329–337. doi: 10.1038/ncpgasthep0498. [DOI] [PubMed] [Google Scholar]
- 40.Varga K, Wagner JA, Bridgen DT, Kunos G. Platelet- and macrophage-derived endogenous cannabinoids are involved in endotoxin-induced hypotension. FASEB J. 1998;12:1035–1044. doi: 10.1096/fasebj.12.11.1035. [DOI] [PubMed] [Google Scholar]
- 41.Wagner JA, Varga K, Ellis EF, Rzigalinski BA, Martin BR, Kunos G. Activation of peripheral CB1 cannabinoid receptors in haemorrhagic shock. Nature. 1997;390:518–521. doi: 10.1038/37371. [DOI] [PubMed] [Google Scholar]
- 42.Wagner JA, Varga K, Kunos G. Cardiovascular actions of cannabinoids and their generation during shock. J Mol Med. 1998;76:824–836. doi: 10.1007/s001090050287. [DOI] [PubMed] [Google Scholar]
- 43.Batkai S, Jarai Z, Wagner JA, Goparaju SK, Varga K, Liu J, Wang L, Mirshahi F, Khanolkar AD, Makriyannis A, Urbaschek R, Garcia N, Jr, Sanyal AJ, Kunos G. Endocannabinoids acting at vascular CB1 receptors mediate the vasodilated state in advanced liver cirrhosis. Nat Med. 2001;7:827–832. doi: 10.1038/89953. [DOI] [PubMed] [Google Scholar]
- 44.Ros J, Claria J, To-Figueras J, Planaguma A, Cejudo-Martin P, Fernandez-Varo G, Martin-Ruiz R, Arroyo V, Rivera F, Rodes J, Jimenez W. Endogenous cannabinoids: a new system involved in the homeostasis of arterial pressure in experimental cirrhosis in the rat. Gastroenterology. 2002;122:85–93. doi: 10.1053/gast.2002.30305. [DOI] [PubMed] [Google Scholar]
- 45.Domenicali M, Caraceni P, Giannone F, Pertosa AM, Principe A, Zambruni A, Trevisani F, Croci T, Bernardi M. Cannabinoid Type 1 Receptor Antagonism Delays Ascites Formation in Rats with Cirrhosis. Gastroenterology. 2009 doi: 10.1053/j.gastro.2009.01.004. [DOI] [PubMed] [Google Scholar]
- 46.Maccarrone M, De Petrocellis L, Bari M, Fezza F, Salvati S, Di Marzo V, Finazzi-Agro A. Lipopolysaccharide downregulates fatty acid amide hydrolase expression and increases anandamide levels in human peripheral lymphocytes. Arch Biochem Biophys. 2001;393:321–328. doi: 10.1006/abbi.2001.2500. [DOI] [PubMed] [Google Scholar]
- 47.Liu J, Batkai S, Pacher P, Harvey-White J, Wagner JA, Cravatt BF, Gao B, Kunos G. Lipopolysaccharide induces anandamide synthesis in macrophages via CD14/MAPK/phosphoinositide 3-kinase/NF-kappaB independently of platelet-activating factor. J Biol Chem. 2003;278:45034–45039. doi: 10.1074/jbc.M306062200. [DOI] [PubMed] [Google Scholar]
- 48.Batkai S, Osei-Hyiaman D, Pan H, El-Assal O, Rajesh M, Mukhopadhyay P, Hong F, Harvey-White J, Jafri A, Hasko G, Huffman JW, Gao B, Kunos G, Pacher P. Cannabinoid-2 receptor mediates protection against hepatic ischemia/reperfusion injury. FASEB J. 2007;21:1788–1800. doi: 10.1096/fj.06-7451com. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Caraceni P, Domenicali M, Bernardi M. The endocannabinoid system and liver diseases. J Neuroendocrinol. 2008;20(Suppl 1):47–52. doi: 10.1111/j.1365-2826.2008.01679.x. [DOI] [PubMed] [Google Scholar]
- 50.Fernandez-Rodriguez CM, Romero J, Petros TJ, Bradshaw H, Gasalla JM, Gutierrez ML, Lledo JL, Santander C, Fernandez TP, Tomas E, Cacho G, Walker JM. Circulating endogenous cannabinoid anandamide and portal, systemic and renal hemodynamics in cirrhosis. Liver Int. 2004;24:477–483. doi: 10.1111/j.1478-3231.2004.0945.x. [DOI] [PubMed] [Google Scholar]
- 51.Kunos G, Batkai S, Offertaler L, Mo F, Liu J, Karcher J, Harvey-White J. The quest for a vascular endothelial cannabinoid receptor. Chem Phys Lipids. 2002;121:45–56. doi: 10.1016/s0009-3084(02)00145-7. [DOI] [PubMed] [Google Scholar]
- 52.Teixeira-Clerc F, Julien B, Grenard P, Tran Van Nhieu J, Deveaux V, Li L, Serriere-Lanneau V, Ledent C, Mallat A, Lotersztajn S. CB1 cannabinoid receptor antagonism: a new strategy for the treatment of liver fibrosis. Nat Med. 2006;12:671–676. doi: 10.1038/nm1421. [DOI] [PubMed] [Google Scholar]
- 53.Domenicali M, Ros J, Fernandez-Varo G, Cejudo-Martin P, Crespo M, Morales-Ruiz M, Briones AM, Campistol JM, Arroyo V, Vila E, Rodes J, Jimenez W. Increased anandamide induced relaxation in mesenteric arteries of cirrhotic rats: role of cannabinoid and vanilloid receptors. Gut. 2005;54:522–527. doi: 10.1136/gut.2004.051599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Yang YY, Lin HC, Huang YT, Lee TY, Hou MC, Wang YW, Lee FY, Lee SD. Effect of chronic CB1 cannabinoid receptor antagonism on livers of rats with biliary cirrhosis. Clin Sci (Lond) 2007;112:533–542. doi: 10.1042/CS20060260. [DOI] [PubMed] [Google Scholar]
- 55.Yang YY, Lin HC, Huang YT, Lee TY, Hou MC, Wang YW, Lee FY, Lee SD. Roles of anandamide in the hepatic microcirculation in cirrhotic rats. Am J Physiol Gastrointest Liver Physiol. 2006;290:G328–334. doi: 10.1152/ajpgi.00367.2005. [DOI] [PubMed] [Google Scholar]
- 56.Maccarroneand M, Finazzi-Agro A. The endocannabinoid system, anandamide and the regulation of mammalian cell apoptosis. Cell Death Differ. 2003;10:946–955. doi: 10.1038/sj.cdd.4401284. [DOI] [PubMed] [Google Scholar]
- 57.Siegmund SV, Qian T, de Minicis S, Harvey-White J, Kunos G, Vinod KY, Hungund B, Schwabe RF. The endocannabinoid 2-arachidonoyl glycerol induces death of hepatic stellate cells via mitochondrial reactive oxygen species. FASEB J. 2007;21:2798–2806. doi: 10.1096/fj.06-7717com. [DOI] [PubMed] [Google Scholar]
- 58.Siegmund SV, Uchinami H, Osawa Y, Brenner DA, Schwabe RF. Anandamide induces necrosis in primary hepatic stellate cells. Hepatology. 2005;41:1085–1095. doi: 10.1002/hep.20667. [DOI] [PubMed] [Google Scholar]
- 59.Gaskari SA, Liu H, Moezi L, Li Y, Baik SK, Lee SS. Role of endocannabinoids in the pathogenesis of cirrhotic cardiomyopathy in bile duct-ligated rats. Br J Pharmacol. 2005;146:315–323. doi: 10.1038/sj.bjp.0706331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Batkai S, Mukhopadhyay P, Harvey-White J, Kechrid R, Pacher P, Kunos G. Endocannabinoids acting at CB1 receptors mediate the cardiac contractile dysfunction in vivo in cirrhotic rats. Am J Physiol Heart Circ Physiol. 2007;293:H1689–1695. doi: 10.1152/ajpheart.00538.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Moezi L, Gaskari SA, Liu H, Baik SK, Dehpour AR, Lee SS. Anandamide mediates hyperdynamic circulation in cirrhotic rats via CB(1) and VR(1) receptors. Br J Pharmacol. 2006;149:898–908. doi: 10.1038/sj.bjp.0706928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Bouchard JF, Lepicier P, Lamontagne D. Contribution of endocannabinoids in the endothelial protection afforded by ischemic preconditioning in the isolated rat heart. Life Sci. 2003;72:1859–1870. doi: 10.1016/s0024-3205(02)02474-8. [DOI] [PubMed] [Google Scholar]
- 63.Lepicier P, Bouchard JF, Lagneux C, Lamontagne D. Endocannabinoids protect the rat isolated heart against ischaemia. Br J Pharmacol. 2003;139:805–815. doi: 10.1038/sj.bjp.0705313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Pacherand P, Hasko G. Endocannabinoids and cannabinoid receptors in ischaemia-reperfusion injury and preconditioning. Br J Pharmacol. 2008;153:252–262. doi: 10.1038/sj.bjp.0707582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Underdown NJ, Hiley CR, Ford WR. Anandamide reduces infarct size in rat isolated hearts subjected to ischaemia-reperfusion by a novel cannabinoid mechanism. Br J Pharmacol. 2005;146:809–816. doi: 10.1038/sj.bjp.0706391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wagner JA, Hu K, Bauersachs J, Karcher J, Wiesler M, Goparaju SK, Kunos G, Ertl G. Endogenous cannabinoids mediate hypotension after experimental myocardial infarction. J Am Coll Cardiol. 2001;38:2048–2054. doi: 10.1016/s0735-1097(01)01671-0. [DOI] [PubMed] [Google Scholar]
- 67.Wagner JA, Hu K, Karcher J, Bauersachs J, Schafer A, Laser M, Han H, Ertl G. CB(1) cannabinoid receptor antagonism promotes remodeling and cannabinoid treatment prevents endothelial dysfunction and hypotension in rats with myocardial infarction. Br J Pharmacol. 2003;138:1251–1258. doi: 10.1038/sj.bjp.0705156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Krylatov AV, Ugdyzhekova DS, Bernatskaya NA, Maslov LN, Mekhoulam R, Pertwee RG, Stephano GB. Activation of type II cannabinoid receptors improves myocardial tolerance to arrhythmogenic effects of coronary occlusion and reperfusion. Bull Exp Biol Med. 2001;131:523–525. doi: 10.1023/a:1012381914518. [DOI] [PubMed] [Google Scholar]
- 69.Di Filippo C, Rossi F, Rossi S, D'Amico M. Cannabinoid CB2 receptor activation reduces mouse myocardial ischemia-reperfusion injury: involvement of cytokine/chemokines and PMN. J Leukoc Biol. 2004;75:453–459. doi: 10.1189/jlb.0703303. [DOI] [PubMed] [Google Scholar]
- 70.Pacherand P, Gao B. Endocannabinoids and liver disease. III. Endocannabinoid effects on immune cells: implications for inflammatory liver diseases. Am J Physiol Gastrointest Liver Physiol. 2008;294:G850–854. doi: 10.1152/ajpgi.00523.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Young RC, Ozols RF, Myers CE. The anthracycline antineoplastic drugs. N Engl J Med. 1981;305:139–153. doi: 10.1056/NEJM198107163050305. [DOI] [PubMed] [Google Scholar]
- 72.Takemuraand G, Fujiwara H. Doxorubicin-induced cardiomyopathy from the cardiotoxic mechanisms to management. Prog Cardiovasc Dis. 2007;49:330–352. doi: 10.1016/j.pcad.2006.10.002. [DOI] [PubMed] [Google Scholar]
- 73.Pacher P, Liaudet L, Bai P, Mabley JG, Kaminski PM, Virag L, Deb A, Szabo E, Ungvari Z, Wolin MS, Groves JT, Szabo C. Potent metalloporphyrin peroxynitrite decomposition catalyst protects against the development of doxorubicin-induced cardiac dysfunction. Circulation. 2003;107:896–904. doi: 10.1161/01.cir.0000048192.52098.dd. [DOI] [PubMed] [Google Scholar]
- 74.Mukhopadhyay P, Rajesh M, Batkai S, Yoshihiro K, Hasko G, Liaudet L, Szabo C, Pacher P. Role of superoxide, nitric oxide and peroxynitrite in doxorubicin-induced cell death in vivo and in vitro. Am J Physiol Heart Circ Physiol. 2009 doi: 10.1152/ajpheart.00795.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Pacher P, Beckman JS, Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiol Rev. 2007;87:315–424. doi: 10.1152/physrev.00029.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Bai P, Mabley JG, Liaudet L, Virag L, Szabo C, Pacher P. Matrix metalloproteinase activation is an early event in doxorubicin-induced cardiotoxicity. Oncol Rep. 2004;11:505–508. [PubMed] [Google Scholar]
- 77.Myers CE, McGuire WP, Liss RH, Ifrim I, Grotzinger K, Young RC. Adriamycin: the role of lipid peroxidation in cardiac toxicity and tumor response. Science. 1977;197:165–167. doi: 10.1126/science.877547. [DOI] [PubMed] [Google Scholar]
- 78.Tokarska-Schlattner M, Zaugg M, Zuppinger C, Wallimann T, Schlattner U. New insights into doxorubicin-induced cardiotoxicity: the critical role of cellular energetics. J Mol Cell Cardiol. 2006;41:389–405. doi: 10.1016/j.yjmcc.2006.06.009. [DOI] [PubMed] [Google Scholar]
- 79.Pacher P, Liaudet L, Bai P, Virag L, Mabley JG, Hasko G, Szabo C. Activation of poly(ADP-ribose) polymerase contributes to development of doxorubicin-induced heart failure. J Pharmacol Exp Ther. 2002;300:862–867. doi: 10.1124/jpet.300.3.862. [DOI] [PubMed] [Google Scholar]
- 80.Bifulco M, Grimaldi C, Gazzerro P, Pisanti S, Santoro A. Rimonabant: just an antiobesity drug? Current evidence on its pleiotropic effects. Mol Pharmacol. 2007;71:1445–1456. doi: 10.1124/mol.106.033118. [DOI] [PubMed] [Google Scholar]
- 81.Malfitano AM, Laezza C, Pisanti S, Gazzerro P, Bifulco M. Rimonabant (SR141716) exerts anti-proliferative and immunomodulatory effects in human peripheral blood mononuclear cells. Br J Pharmacol. 2008;153:1003–1010. doi: 10.1038/sj.bjp.0707651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kunos G, Osei-Hyiaman D, Batkai S, Sharkey KA, Makriyannis A. Should peripheral CB(1) cannabinoid receptors be selectively targeted for therapeutic gain? Trends Pharmacol Sci. 2009;30:1–7. doi: 10.1016/j.tips.2008.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]