

Abstract
AIM: To investigate the effect of activation of canonical Wnt signaling pathway on the proliferation and differentiation of hepatic oval cells in vitro.
METHODS: WB-F344 cells were treated with recombinant Wnt3a (20, 40, 80, 160, 200 ng/mL) in serum-free medium for 24 h. Cell proliferation was measured by Brdu incorporation analysis; untreated WB-F344 cells were taken as controls. After treatment with Wnt3a (160 ng/mL) for 24 h, subcellular localization and protein expression of β-catenin in WB-F344 cells treated and untreated with Wnt3a were examined by immunofluorescence staining and Western blot analysis. CyclinD1 mRNA expression was determined by semi-quantitative reverse-transcript polymerase chain reaction (RT-PCR). The mRNA levels of some phenotypic markers (AFP, CK-19, ALB) and two hepatic nuclear factors (HNF-4, HNF-6) were measured by RT-PCR. Expressions of CK-19 and AFP protein were detected by Western blot analysis.
RESULTS: Wnt3a promoted proliferation of WB-F344 cells. Stimulation of WB-F344 cells with recombinant Wnt3a resulted in accumulation of the transcriptional activator β-catenin, together with its translocation into the nuclei, and up-regulated typical Wnt target gene CyclinD1. After 3 d of Wnt3a treatment in the absence of serum, WB-F344 cells retained their bipotential to express several specific phenotypic markers of hepatocytes and cholangiocytes, such as AFP and CK-19, following activation of the canonical Wnt signaling pathway.
CONCLUSION: The canonical Wnt signaling pathway promotes proliferation and self-renewal of rat hepatic oval cells.
Keywords: Canonical Wnt signaling pathway, Oval cells, Cell proliferation, Self-renewal of cells
INTRODUCTION
The liver is an organ with a remarkable regenerative capacity, provided by preexisting hepatocytes and cholangiocytes under normal conditions[1]. However, viral infection, alcohol, some drugs and self-immunity may cause liver inflammation and fibrosis. When the regenerative ability of hepatocytes to divide and replace the damaged tissue is compromised, oval cells are activated, and then may give rise to both hepatocytes and biliary epithelial cells[2,3]. Oval cell activation and proliferation serve as a source of cell replenishment and tissue repair as they differentiate into functional mature hepatocytes, thus aiding in the process of liver regeneration[4,5]. Therefore, oval cells are also designated as facultative hepatic stem cells because they proliferate only in response to chronic regenerative stimuli and have bipotential differentiation capabilities for both hepatocytes and cholangiocytes[6,7]. Although oval cells are usually found in normal human fetal livers, they are also observed in numerous hepatic pathologies[8,9]. Oval cell activation in malignancies such as hepatocellular carcinoma (HCC) and cholangiocarcinoma is of special relevance in light of the theories supporting the existence of cancer stem cells, which may serve as a tumor source, or as a mechanism of cancer recurrence or metastasis[10,11]. However, little is known about the signaling mechanisms involved in oval cell proliferation and differentiation.
The canonical Wnt signaling pathway is highly conserved throughout animal development, during which it exerts pleiotropic effects on cell proliferation, differentiation, and polarity or migration[12]. The Wnt signaling pathway, identified recently, critically regulates various postnatal stem cell compartments, including the hematopoietic, skin, and enteric systems. In this respect, it has been demonstrated that hematopoietic stem cells maintain an undifferentiated self-renewing state through constitutive activation of the canonical Wnt signaling pathway with Wnt3a, a prominent member of the Wnt family[13,14]. In the liver, there is accumulating evidence that Wnt/β-catenin signaling plays a central role in various aspects of hepatic biology, including liver development, regeneration, growth, and oncogenesis. During liver development, β-catenin critically regulates hepatic progenitor cell proliferation, and over-expression or inhibition of β-catenin either increases or decreases the overall liver size, respectively[15-19]. Studies of pathological specimens and rodent models of liver diseases have demonstrated aberrations in the Wnt/β-catenin signaling pathway in conditions ranging from hepatitis to HCC[20]. However, no studies have definitively addressed the role of canonical Wnt signaling in proliferation and differentiation of hepatic oval cells. Therefore, we hypothesize that Wnt signaling can regulate the proliferation and differentiation of oval cells.
WB-F344, a rat hepatic stem-like epithelial cell line, isolated from the liver of an adult male Fischer-344 rat, can express a phenotypic repertory of both hepatocytes and bile duct epithelial cells compared with those of normal hepatocytes, biliary epithelial (ductular) and “oval” cells isolated from liver treated with chemical carcinogens. The phenotypic properties of cultured liver epithelial cell line most resemble those of the “oval” cells. Thus, it is considered to be an in vitro model of bipotent oval cells as it shares their phenotype[21,22]. After transplantation into livers of adult syngeneic German-strain Fischer-344 rats that are deficient in bile canalicular enzyme dipeptidyl peptidase IV (DPP-IV), WB-F344 cells integrate into hepatic plates and differentiate into mature hepatocytes[23]. Moreover, when treated with sodium butyrate and cultured on Matrigel, WB-F344 cells can differentiate along the biliary phenotype in vitro[24]. In the present study, we provided direct evidence for the activation of canonical Wnt signal transduction in WB-F344 cells in response to Wnt ligands, and showed that activation of canonical Wnt signaling regulates the proliferation and bipotential of WB-F344 cells. A better understanding of its role in hepatic stem cell proliferation and differentiation can lead to the successful manipulation of liver biology for therapeutic purposes.
MATERIALS AND METHODS
Cell line culture
The rat hepatic oval cell line (WB-F344) was obtained from Academy of Military Medical Sciences. The cells were cultured in Dulbecco’s modified Eagle’s medium/F12 (Invitrogen, Carlsbad, CA), supplemented with 10% fetal bovine serum (Gibco BRL, USA), 10 mL of 200 mmol/L L-glutamine and 0.5 mL of penicillin-streptomycin mixture. All cultures were maintained at 37°C in 50 mL/L CO2.
Proliferation test (Brdu incorporation assays)
Cell proliferation was detected by Brdu incorporation assays. A total of 6000 WB-F344 cells were seeded into 96-well culture plates. Cells were brought to 90% confluence and then fasted overnight in serum-free media before addition of Brdu to a final concentration of 10 μmol/L. Two hours later, the cells were stimulated with escalating doses of Wnt3a (R&D, Systems, Inc) (20, 40, 80, 160, and 200 ng/mL) for 24 h, washed free of BrdU and harvested. The cells were fixed with 4% paraformaldehyde for 30 min, blocked with 1% BSA in phosphate-buffered saline containing 0.2% Triton X-100 for 30 min, incubated with a HRP-Brdu antibody (1:200) for 2 h at 37°C, and washed. A TMB substrate solution was added to the wells and color developed in proportion to the amount of Brdu bound. The stop solution changed the color from blue to yellow, and intensity of the color was measured at 450 nm with a spectrophotometer.
Immunofluorescence staining
For subcellular localization of β-catenin by immunocytochemistry, WB-F344 cells were plated onto chamber slides resulting in 90% confluence, and then fasted overnight in serum-free media before the addition of 160 ng/mL Wnt3a (R&D, Systems, Inc). The cells were fixed with 4% paraformaldehyde for 30 min at 37°C. Permeabilization of the cells was achieved after incubation for 30 min at 37°C with PBS containing 0.2% Triton X-100. To minimize nonspecific binding of the antibody, blocking was carried out with a buffer containing 1% bovine serum albumin for 1 h. β-catenin antibody (R&D, Systems, Inc) was applied at a 1:25 dilution for 90 min at 37°C. As a negative control, PBS was used instead of the primary antibody to exclude the unspecific binding of the secondary antibody. No fluorescent labeling was observed in the negative control. After repeated washing with PBS, the cells were incubated with a goat-anti-mouse antibody labeled with fluorescein isothiocyanate (1:10) for an additional 30 min. Finally, cell nuclei were counterstained with Hoechst 33258. Images were obtained using a confocal laser scanning microscope.
RNA isolation and reverse-transcription polymerase chain reaction (RT-PCR)
WB-F344 cells were brought to 90% confluence and then fasted overnight in serum-free media. After stimulation with or without Wnt3a, cells were cultivated for 1 and 3 d, respectively. Total RNA was extracted from WB-F344 cells treated with Wnt3a with Trizol according to the manufacture’s instructions. RNA (1 μg) was reverse transcribed. For semi-quantitative PCR, the number of cycles corresponded to the mid-logarithmic phase. Primers were designed using GenBank sequences (Table 1). PCR amplification was performed using PCR Master Mix (Taqman) according to the manufacturer’s instructions. PCR products were analyzed by electrophoresis on a 2% agarose gel.
Table 1.
Primer sequences used for RT-PCR
| Gene | Primer sequences | Annealing temp (°C) | Cycles | Ampliconsize (bp) |
| CyclinD1 | 5’-ATTGAAGCCCTTCTGGAGTCAAGCC-3’ | 56 | 26 | 420 |
| 5’-TCTATTTTTGTAGCACCCCCCCGTC-3’ | ||||
| AFP | 5’-GCTGAACCCAGAGTACTG AC-3’ | 60 | 34 | 472 |
| 5’-GACACGTCGTAGATGAACGTG-3’ | ||||
| ALB | 5’-AAGGCACCCCGATTACTCCG-3’ | 56 | 36 | 608 |
| 5’-TGCGAAGTCACCCATCACCG-3’ | ||||
| CK-19 | 5’-ATGACTTCCTATAGCTATCG-3’ | 64 | 34 | 340 |
| 5’-CACCTCCAGCTCGCCATTAG-3’ | ||||
| HNF-6 | 5’-GACAAATGGCAGGACGAGGG-3’ | 62 | 36 | 781 |
| 5’-AGCGTACTGGTTTAGGTGCC-3’ | ||||
| HNF-4α | 5’-CTTCCTTCTTCATGCCAG-3’ | 62 | 34 | 269 |
| 5’-ACACGTCCCCATCTGAAG-3’ | ||||
| GAPDH | 5’-ACCACAGTCCATGCCATCAC-3’ | 56 | 24 | 469 |
| 5’-TCCACCACCCTGTTGCTGTA-3’ |
Western blot analysis
After stimulation with Wnt3a, cells were cultivated for 1 and 3 d, respectively. Trypsinized protein was isolated using a lysis buffer (50 mmol/L Na2HPO4, 50 mmol/L NaH2PO4, 0.2 mol/L NaCl, 5 mmol/L EDTA, 1% Triton X-100, pH 6.0). After incubation on ice for 30 min, samples were centrifuged at 13 000 r/min for 20 min at 4°C. Then, a 2 × dithiothreitol (DTT) loading buffer containing 0.4 mol/L Tris (pH 6.8), 4% SDS, 20% glycerol, and 10% DTT was added to the sample supernatants, and incubated for 5 min at 95°C. Following electrophoretic separation by 10% SDS-polyacrylamide gel electrophoresis, proteins were electroblotted onto nitrocellulose membranes. The membranes were blocked in a NET buffer (150 mmol/L NaCl; 5 mmol/L EDTA, pH 8.0; 50 mmol/L Tris/HCl, pH 7.5; 0.05% Triton X-100) containing 2.5% gelatin (Merck) for 1 h at room temperature. Polyclonal antibodies against β-catenin (R&D, Systems, Inc), cytokeratin-19 (CK-19) (Santa Cruz Biotechnology Inc), AFP (Santa Cruz Biotechnology Inc) were used at a dilution of 1:25 (β-catenin), 1:200 (AFP, CK-19) and 1:400 (β-actin), respectively, and incubated for 1 h at room temperature. Thereafter, the membranes were washed in a NET buffer, and further incubated with a peroxidase-conjugated antibody at a dilution of 1:20 000. Antibody binding was visualized by DAB. The bands were semi-quantitatively evaluated by densitometric analysis. Protein expression levels of β-catenin were thereby normalized to those of the housekeeping gene β-actin.
Differentiation assay
WB-F344 cells grown in a Wnt3a-containing medium in the absence of serum were cultured with or without Wnt3a for 3 d. The medium was changed every day. After 3 d of coculture, total Mrna and protein were extracted and analyzed for expression of CK-19, ALB, AFP by RT-PCR and Western blot analysis.
Statistical analysis
All results were expressed as mean ± SD. Measurement data were analyzed using one-way analysis of variances (ANOVA, SPSS 11.5). P < 0.05 was considered statistically significant.
RESULTS
Activation of the Wnt pathway, accumulation of β-catenin, its translocation into nuclei, and transcriptional activation of Wnt target genes
To investigate whether treatment of WB-F344 cells with recombinant Wnt3a (160 ng/mL) influences β-catenin in WB-F344 cells at the protein level, we monitored the expression of β-catenin by semi-quantitative Western blot analysis. Accumulation of β-catenin protein (1.2-fold) was observed 1 d after addition of 160 ng/mL Wnt3a (Figure 1A). To confirm the data obtained by Western blot analysis and prove whether Wnt3a stimulation has any influence on the subcellular localization of β-catenin, immunocytochemistry was performed. Clear nuclear staining for β-catenin was observed after treatment with 160 ng/mL Wnt3a for 1 d, suggesting that Wnt3a stimulation is responsible for the accumulation of β-catenin and its translocation from cytoplasm into nuclei (Figure 1B). To evaluate whether Wnt3a-mediated accumulation of β-catenin would result in the activation of typical Wnt target genes, the mRNA expression levels of CyclinD1 was semi-quantified by RT-PCR 1 d after Wnt3a stimulation (160 ng/mL). A significant induction in CyclinD1 was observed (Figure 2). These findings indicate that activation of the Wnt pathway in WB-F344 cells can result in accumulation of β-catenin, its translocation into nuclei, and enhanced expression of Wnt target genes.
Figure 1.
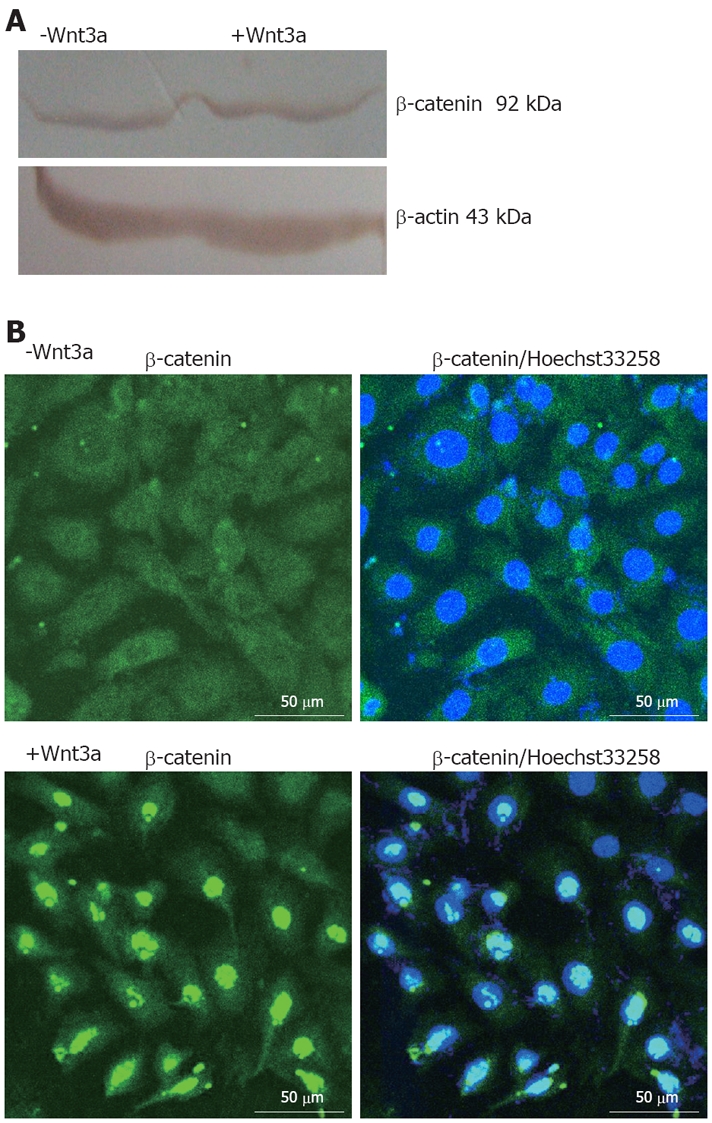
Effects of Wnt3a on β-catenin expression, its subcellular localization, and induction of typical Wnt target genes. A: Stimulation of WB-F344 cells with 160 ng/mL Wnt3a for 1 d revealing a slight increase in β-catenin protein level as shown by Western blot analysis and densitometric analysis; B: Immunocytochemistry analysis of β-catenin exhibiting perinuclear staining for β-catenin in unstimulated WB-F344 cells (upper panels), whereas addition of 160 ng/mL Wnt3a for 1 d (lower panels) showing clear nuclear staining for β-catenin. Immunofluorescence was performed using a polyclonal antibody against β-catenin (left panels). In addition, nuclei of WB-F344 cells were stained with Hoechst33258 (right panels). Scale bars: 50 μm.
Figure 2.
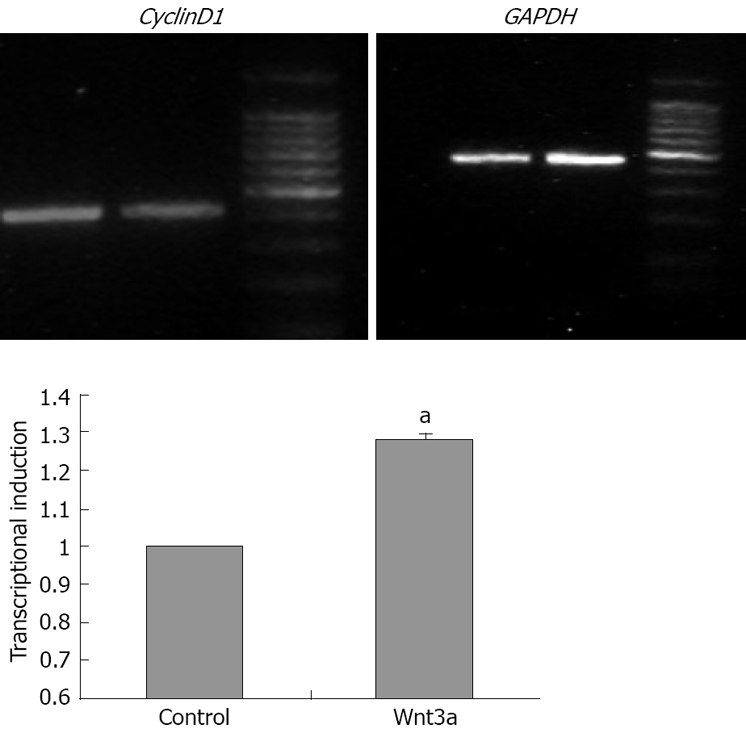
The mRNA expression levels of the known Wnt target genes CyclinD1 was semi-quantified at day 1 during stimulation with Wnt3a (160 ng/mL) and normalized to the expression levels in untreated WB-F344 cells (set as 100%). The mRNA expression level of CyclinD1, one of the known Wnt target genes in WB-F344 cells after stimulation by Wnt3a (160 ng/mL) for 1 d was semi-quantified by RT-PCR and the results was normalized to the expression levels in untreated WB-F344 cells (set as 1). CyclinD1 was upregulated under stimulating conditions on day 1. For quantification, CyclinD1 mRNA was scanned by densitometric analysis and normalized to GAPDH. Data are presented as mean ± SD of triplicate experiments. aP < 0.05 in comparison with un-treated cells.
Effect of Wnt3a on WB-F344 cell proliferation
To determine whether WB-F344 cells respond to Wnt ligand stimulation with activation of the canonical Wnt pathway, WB-F344cells in culture were stimulated by purified Wnt3a protein. Brdu incorporation assays were performed to determine whether Wnt stimulates the proliferation of WB-F344 cells. After serum starvation, Wnt3a stimulation of WB-F344 cells resulted in significantly more uptake of Brdu compared with controls in the absence of serum. The proliferation of WB-F344 cells increased after the addition of Wnt3a, and reached its peak at 160 ng/mL Wnt3a (Figure 3). To evaluate whether the expression of Wnt target gene CyclinD1 is directly correlated with the proliferation of WB-F344 cells, the mRNA expression level of CyclinD1 was determined by semi-quantitative RT-PCR 1 d after Wnt3a stimulation (160 ng/mL). A significant up-regulation of CyclinD1 expression was observed, suggesting that CyclinD1 has a major impact on the proliferation of WB-F344cells. These findings indicate that proliferation of WB-F344 cells is stimulated by the canonical Wnt signaling pathway, which may be the mechanism underlying the up-regulated CyclinD1 expression.
Figure 3.

Proliferation of WB-F344 cells upon treatment with Wnt3a measured by Brdu incorporation assay. The proliferation of WB-F344 cells was significantly enhanced by stimulation with recombinant Wnt3a for 1 d Data are presented as mean ± SD. aP < 0.05 vs untreated WB-F344 cells.
Activation of the canonical Wnt signaling pathway promoted self-renewal of WB-F344 cells
Although Wnt3a can activate the canonical Wnt signaling pathway, it remains unclear whether activation of the canonical Wnt signaling pathway results in differentiation of WB-F344 cells. If Wnt3a can promote cell differentiation, WB-F344 cells treated with Wnt3a should express specific markers after activation of the canonical Wnt signaling pathway. To test this hypothesis, WB-F344 cells were cultured in the presence or absence of Wnt3a-containing medium for 3 d under serum-free conditions. RT-PCR and Western blot were performed for the expression of markers in WB-F344 cells treated or untreated with Wnt3a. Two cell–specific markers, AFP and CK-19 and two hepatocyte nuclear factors, HNF-4α, HNF-6 were expressed in both Wnt3a-treated and untreated WB-F344 cells. In respect of these markers, Wnt3a-treated WB-F344 cells did not show any marked deviation compared with untreated WB-F344 cells (Figure 4). The transcription factors, known to play a key role in differentiation of hepatocytes and cholangiocytes, were expressed in both Wnt3a-treated and untreated WB-F344 cells. These findings suggest that self-renewal of WB-F344 cells is stimulated by the canonical Wnt signal transduction.
Figure 4.

RT-PCR and Western blot analysis of differentiated WB-F344 cells treated or untreated with Wnt3a. A: Wnt3a-treated WB-F344 cells expressing two phenotypic markers (CK-19 and AFP) and two hepatic nuclear factors (HNF4α and HNF-6) at mRNA level with untreated WB-F344 cells as controls; B: Wnt3a-treated cells expressing two phenotypic markers (CK-19 and AFP) at protein level.
DISCUSSION
Oval cell activation occurs in the majority of chronic liver diseases and increases with the severity of the disease. In moderate and severe degrees of inflammation, intermediate hepatocytes occur, having a phenotype intermediate between progenitor cells/ductular cells and mature hepatocytes. The number of these intermediate hepatocytes gradually increases with higher degrees of inflammation and necrosis in necrotizing hepatitis or with more advanced stages of (non) alcoholic steatohepatitis[25-27]. However, little is known about the signaling pathways involved in controlling hepatic oval cell proliferation and differentiation. The present study demonstrated that Wnt/β-catenin signaling plays an important role in the proliferation and renewal of oval cells in vitro.
Although multiple Wnts are now known to play a role in the proliferation and renewal of stem cells via the canonical or non-canonical pathway, Wnt-1 and -3 have a similar mode of action leading to β-catenin accumulation[28,29]. Taking advantage of this redundancy and availability of biologically active Wnt3a, we tested the impact of Wnt enrichment on WB-F344 cells, at beginning of cultures. Thus, this study directly addressed the effect of Wnt3a on the proliferation and differentiation of WB-F344 cells.
We performed this study in serum-free conditions to minimize other confounding factors. Our initial results demonstrate that Wnt proteins, showing high transforming activity in WB-F344 cells, could activate the canonical Wnt signaling pathway. To activate the canonical Wnt signaling pathway, WB-F344 cells were stimulated with recombinant Wnt3a (160 ng/mL) for 24 h. β-catenin, upon treatment with Wnt3a, translocated into the nuclei of WB-F344 cells where it promotes TCF/LEF-dependent transcription, with a slightly increased accumulation of β-catenin at protein level, suggesting that degradation of β-catenin diminished by the destructive complex is the underlying mechanism[30]. Nuclear accumulation of this protein indicates that activity of the Wnt signal pathway is increased[31]. Furthermore, Wnt3a-mediated nuclear translocation of β-catenin results in the up-regulation of CyclinD1, suggesting that the canonical Wnt target gene, originally identified in colon carcinoma cells[32], can be induced in WB-F344 cells. Accumulated β-catenin is translocated into the nuclei of WB-F344 cells where it binds to the transcription factors, T cell factor (Tcf)/lymphoid enhancer factor (Lef), thereby stimulates expression of target genes[33]. Therefore, activation of the Wnt signaling pathway in WB-F344 cells results in accumulation of β-catenin, its translocation into nuclei, and enhanced expression of Wnt target genes.
We also investigated the effect of recombinant Wnt3a on proliferation of WB-F344 cells in serum-free environment, showing that recombinant Wnt3a could stimulate the proliferation of WB-F344 cells. This observation is consistent with the reported findings that β-catenin plays a central role in regulating the proliferation and regeneration of hepatocytes[34-36]. To investigate whether the expression of CyclinD1 is directly correlated with the proliferation of WB-F344 cells, mRNA expression in CyclinD1, which plays a fundamental role in cell cycle at the G1-S phase transition, was semi-quantified by RT-PCR 1 d after Wnt3a stimulation (160 ng/mL). A significant mRNA expression was induced in CyclinD1. This effect was even more pronounced under Wnt stimulatory conditions. Moreover, it was reported that proliferation of WB-F344 cells is severely impaired in the absence of β-catenin, secondary to the decreased expression of downstream targets such as CyclinD1, which are critical in proliferation[37]. It has also been shown that WB-F344 cells proliferate in postnatal liver development, ex vivo embryonic liver development, and in facultative liver stem and oval cells[38-41]. These findings further support the idea that the Wnt signaling pathway is largely involved in controlling the proliferation of WB-F344 cells.
Oval cells are known to be tumorigenic[42]. The Wnt/β-catenin signaling pathway plays an important role in the pathogenesis of hepatic adenoma and its progression to HCC[43] and also in self-renewal of stem cells in several tissue types[44]. β-catenin is a mediator of cancer stem cells[43] and plays an important role in early liver development at the stage of ongoing hepatic progenitor proliferation[39,45]. In the present study, the hepatobiliary phenotype was confirmed by RT-PCR and Western blot analysis. Wnt3a-treated and untreated WB-F344 cells cells were positive for AFP and CK-19 but negative for ALB, one of the final maturation phase markers, indicating that they were immature cells. The bipotential phenotype of WB-F344 cells was observed in oval cells, presumed precursors of hepatocytes and biliary cells. HNF4α and HNF6 are known to play a key role in differentiation of hepatocytes and cholangiocytes[46-50]. Our results also demonstrate that WB-F344 cells treated or untreated with Wnt3a expressed HNF4α and HNF6. Studies in knockout mice showed that hepatocyte nuclear factor HNF4α regulates transcription of genes essential for hepatocytic cell lineage[51,52], whereas HNF6 is involved in the development of gallbladder and bile ducts[53,54]. HNF4α and HNF6 are expressed in fetal hepatoblasts and show different expression patterns in adult liver. HNF4α is exclusively expressed in fetal and adult hepatocytes. No HNF4α expression has been observed in fetal ductal plate or in bile duct epithelium or in normal adult liver. HNF6 is also expressed in fetal and adult hepatocytes and in fetal BEC. HNF6 is completely lost from the BEC with mature biliary phenotype[55]. It was reported more recently that the Wnt/β-catenin signaling pathway plays a critical role in oval cell activation[40]. HNF4α and HNF6 are expressed in oval cells of the liver activated by 2-AAF/PH[56]. Our results show that activation of the Wnt/β-catenin signaling pathway could promote self-renewal of WB-F344 cells, indicating that some molecules involved in the canonical Wnt pathway may be therapeutic targets. β-catenin targeting might be of essence in preneoplastic and early or late HCC as a chemopreventive or che-motherapeutic measure.
In conclusion, the canonical Wnt signaling pathway plays a key role in regulating the proliferation and self-renewal of hepatic oval cells. The detailed mechanism of Wnt3a underlying the differentiation of WB-F344 cells and whether β-catenin can directly control the expression of HNF4α and HNF6 need further study.
COMMENTS
Background
Oval cell activation occurs in the majority of chronic liver diseases and increases with the severity of the disease. However, little is known about the signaling pathways involved in controlling hepatic oval cell proliferation and differentiation. The canonical Wnt signaling pathway is highly conserved throughout animal development during which it exerts pleiotropic effects on cell proliferation, differentiation, and polarity or migration. In the present study, the authors aimed to investigate the effect of activation of canonical Wnt signaling pathway on the proliferation and differentiation of heaptic oval cells in vitro.
Research frontiers
There is accumulating evidence that Wnt/β-catenin signaling in the liver plays a central role in various aspects of hepatic biology, including liver development, regeneration, growth, and oncogenesis. Studies of pathological specimens and rodent models of liver diseases have demonstrated aberrations in the Wnt/β-catenin signaling pathway in conditions ranging from hepatitis to hepatocellular carcinoma (HCC). However, no studies have definitively addressed the role of canonical Wnt signaling in proliferation and differentiation of hepatic oval cells.
Innovations and breakthroughs
This is the first study addressing the role of canonical Wnt signaling in proliferation and differentiation of hepatic oval cells. The canonical Wnt signaling pathway plays a key role in regulating the proliferation and self-renewal of hepatic oval cells.
Applications
Oval cells are known to be tumorigenic. The Wnt/β-catenin signaling pathway plays an important role in the pathogenesis of hepatic adenoma and its progression to HCC, and also in self-renewal of stem cells in several tissue types. β-catenin is a mediator of cancer stem cells. Activation of the Wnt/β-catenin signaling pathway could promote self-renewal of oval cells, indicating that some molecules involved in the canonical Wnt pathway may be therapeutic targets. β-catenin targeting might be of essence in preneoplastic and early or late HCC as a chemopreventive or chemotherapeutic measure.
Terminology
The Wnt signaling pathway, identified recently, critically regulates various postnatal stem cell compartments, including the hematopoietic, skin, and enteric systems. In this respect, it has been demonstrated that hematopoietic stem cells maintain an undifferentiated self-renewing state through constitutive activation of the canonical Wnt signaling pathway with Wnt3a, a prominent member of the Wnt family.
Peer review
In this study, the authors demonstrated the role of canonical Wnt signaling in proliferation and differentiation of hepatic oval cells. This work adds significant information that activation of the Wnt/β-catenin signaling pathway could promote proliferation and self-renewal of oval cells, indicating that some molecules involved in the canonical Wnt pathway may be therapeutic targets.
Footnotes
Peer reviewers: Ezio Laconi, Professor, University of Cagliari, Via Porcell 4, Cagliari 09125, Italy; Chee-Kin Hui, MD, University of Hong Kong, Queen Mary Hospital, Department of Medicine, 102 Pokfulam Road, Hong Kong SAR, China
S- Editor Li DL L- Editor Li M E- Editor Lin YP
References
- 1.Taub R. Liver regeneration: from myth to mechanism. Nat Rev Mol Cell Biol. 2004;5:836–847. doi: 10.1038/nrm1489. [DOI] [PubMed] [Google Scholar]
- 2.Lowes KN, Croager EJ, Olynyk JK, Abraham LJ, Yeoh GC. Oval cell-mediated liver regeneration: Role of cytokines and growth factors. J Gastroenterol Hepatol. 2003;18:4–12. doi: 10.1046/j.1440-1746.2003.02906.x. [DOI] [PubMed] [Google Scholar]
- 3.Walkup MH, Gerber DA. Hepatic stem cells: in search of. Stem Cells. 2006;24:1833–1840. doi: 10.1634/stemcells.2006-0063. [DOI] [PubMed] [Google Scholar]
- 4.Fausto N, Campbell JS. The role of hepatocytes and oval cells in liver regeneration and repopulation. Mech Dev. 2003;120:117–130. doi: 10.1016/s0925-4773(02)00338-6. [DOI] [PubMed] [Google Scholar]
- 5.Petersen BE. Hepatic "stem" cells: coming full circle. Blood Cells Mol Dis. 2001;27:590–600. doi: 10.1006/bcmd.2001.0422. [DOI] [PubMed] [Google Scholar]
- 6.Fausto N. Liver regeneration and repair: hepatocytes, progenitor cells, and stem cells. Hepatology. 2004;39:1477–1487. doi: 10.1002/hep.20214. [DOI] [PubMed] [Google Scholar]
- 7.Newsome PN, Hussain MA, Theise ND. Hepatic oval cells: helping redefine a paradigm in stem cell biology. Curr Top Dev Biol. 2004;61:1–28. doi: 10.1016/S0070-2153(04)61001-5. [DOI] [PubMed] [Google Scholar]
- 8.Masson NM, Currie IS, Terrace JD, Garden OJ, Parks RW, Ross JA. Hepatic progenitor cells in human fetal liver express the oval cell marker Thy-1. Am J Physiol Gastrointest Liver Physiol. 2006;291:G45–G54. doi: 10.1152/ajpgi.00465.2005. [DOI] [PubMed] [Google Scholar]
- 9.Roskams T, De Vos R, Van Eyken P, Myazaki H, Van Damme B, Desmet V. Hepatic OV-6 expression in human liver disease and rat experiments: evidence for hepatic progenitor cells in man. J Hepatol. 1998;29:455–463. doi: 10.1016/s0168-8278(98)80065-2. [DOI] [PubMed] [Google Scholar]
- 10.Knight B, Matthews VB, Olynyk JK, Yeoh GC. Jekyll and Hyde: evolving perspectives on the function and potential of the adult liver progenitor (oval) cell. Bioessays. 2005;27:1192–1202. doi: 10.1002/bies.20311. [DOI] [PubMed] [Google Scholar]
- 11.Lee JS, Heo J, Libbrecht L, Chu IS, Kaposi-Novak P, Calvisi DF, Mikaelyan A, Roberts LR, Demetris AJ, Sun Z, et al. A novel prognostic subtype of human hepatocellular carcinoma derived from hepatic progenitor cells. Nat Med. 2006;12:410–416. doi: 10.1038/nm1377. [DOI] [PubMed] [Google Scholar]
- 12.Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol. 2004;20:781–810. doi: 10.1146/annurev.cellbio.20.010403.113126. [DOI] [PubMed] [Google Scholar]
- 13.Lowry WE, Blanpain C, Nowak JA, Guasch G, Lewis L, Fuchs E. Defining the impact of beta-catenin/Tcf transactivation on epithelial stem cells. Genes Dev. 2005;19:1596–1611. doi: 10.1101/gad.1324905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wodarz A, Nusse R. Mechanisms of Wnt signaling in development. Annu Rev Cell Dev Biol. 1998;14:59–88. doi: 10.1146/annurev.cellbio.14.1.59. [DOI] [PubMed] [Google Scholar]
- 15.Devereux TR, Stern MC, Flake GP, Yu MC, Zhang ZQ, London SJ, Taylor JA. CTNNB1 mutations and beta-catenin protein accumulation in human hepatocellular carcinomas associated with high exposure to aflatoxin B1. Mol Carcinog. 2001;31:68–73. doi: 10.1002/mc.1041. [DOI] [PubMed] [Google Scholar]
- 16.Micsenyi A, Tan X, Sneddon T, Luo JH, Michalopoulos GK, Monga SP. Beta-catenin is temporally regulated during normal liver development. Gastroenterology. 2004;126:1134–1146. doi: 10.1053/j.gastro.2003.12.047. [DOI] [PubMed] [Google Scholar]
- 17.Monga SP, Monga HK, Tan X, Mule K, Pediaditakis P, Michalopoulos GK. Beta-catenin antisense studies in embryonic liver cultures: role in proliferation, apoptosis, and lineage specification. Gastroenterology. 2003;124:202–216. doi: 10.1053/gast.2003.50000. [DOI] [PubMed] [Google Scholar]
- 18.Monga SP, Pediaditakis P, Mule K, Stolz DB, Michalopoulos GK. Changes in WNT/beta-catenin pathway during regulated growth in rat liver regeneration. Hepatology. 2001;33:1098–1109. doi: 10.1053/jhep.2001.23786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Taniguchi K, Roberts LR, Aderca IN, Dong X, Qian C, Murphy LM, Nagorney DM, Burgart LJ, Roche PC, Smith DI, et al. Mutational spectrum of beta-catenin, AXIN1, and AXIN2 in hepatocellular carcinomas and hepatoblastomas. Oncogene. 2002;21:4863–4871. doi: 10.1038/sj.onc.1205591. [DOI] [PubMed] [Google Scholar]
- 20.Thompson MD, Monga SP. WNT/beta-catenin signaling in liver health and disease. Hepatology. 2007;45:1298–1305. doi: 10.1002/hep.21651. [DOI] [PubMed] [Google Scholar]
- 21.Thorgeirsson SS, Grisham JW. Overview of recent experimental studies on liver stem cells. Semin Liver Dis. 2003;23:303–312. doi: 10.1055/s-2004-815559. [DOI] [PubMed] [Google Scholar]
- 22.Tsao MS, Smith JD, Nelson KG, Grisham JW. A diploid epithelial cell line from normal adult rat liver with phenotypic properties of 'oval' cells. Exp Cell Res. 1984;154:38–52. doi: 10.1016/0014-4827(84)90666-9. [DOI] [PubMed] [Google Scholar]
- 23.Coleman WB, McCullough KD, Esch GL, Faris RA, Hixson DC, Smith GJ, Grisham JW. Evaluation of the differentiation potential of WB-F344 rat liver epithelial stem-like cells in vivo. Differentiation to hepatocytes after transplantation into dipeptidylpeptidase-IV-deficient rat liver. Am J Pathol. 1997;151:353–359. [PMC free article] [PubMed] [Google Scholar]
- 24.Couchie D, Holic N, Chobert MN, Corlu A, Laperche Y. In vitro differentiation of WB-F344 rat liver epithelial cells into the biliary lineage. Differentiation. 2002;69:209–215. doi: 10.1046/j.1432-0436.2002.690414.x. [DOI] [PubMed] [Google Scholar]
- 25.Roskams T, Yang SQ, Koteish A, Durnez A, DeVos R, Huang X, Achten R, Verslype C, Diehl AM. Oxidative stress and oval cell accumulation in mice and humans with alcoholic and nonalcoholic fatty liver disease. Am J Pathol. 2003;163:1301–1311. doi: 10.1016/S0002-9440(10)63489-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lowes KN, Brennan BA, Yeoh GC, Olynyk JK. Oval cell numbers in human chronic liver diseases are directly related to disease severity. Am J Pathol. 1999;154:537–541. doi: 10.1016/S0002-9440(10)65299-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Libbrecht L, Desmet V, Van Damme B, Roskams T. Deep intralobular extension of human hepatic 'progenitor cells' correlates with parenchymal inflammation in chronic viral hepatitis: can 'progenitor cells' migrate? J Pathol. 2000;192:373–378. doi: 10.1002/1096-9896(2000)9999:9999<::AID-PATH700>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- 28.Kispert A, Vainio S, McMahon AP. Wnt-4 is a mesenchymal signal for epithelial transformation of metanephric mesenchyme in the developing kidney. Development. 1998;125:4225–4234. doi: 10.1242/dev.125.21.4225. [DOI] [PubMed] [Google Scholar]
- 29.Ikeya M, Lee SM, Johnson JE, McMahon AP, Takada S. Wnt signalling required for expansion of neural crest and CNS progenitors. Nature. 1997;389:966–970. doi: 10.1038/40146. [DOI] [PubMed] [Google Scholar]
- 30.Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherin pathways. Science. 2004;303:1483–1487. doi: 10.1126/science.1094291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kikuchi A, Yamamoto H, Kishida S. Multiplicity of the interactions of Wnt proteins and their receptors. Cell Signal. 2007;19:659–671. doi: 10.1016/j.cellsig.2006.11.001. [DOI] [PubMed] [Google Scholar]
- 32.Tetsu O, McCormick F. Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature. 1999;398:422–426. doi: 10.1038/18884. [DOI] [PubMed] [Google Scholar]
- 33.Cadigan KM, Nusse R. Wnt signaling: a common theme in animal development. Genes Dev. 1997;11:3286–3305. doi: 10.1101/gad.11.24.3286. [DOI] [PubMed] [Google Scholar]
- 34.Suksaweang S, Lin CM, Jiang TX, Hughes MW, Widelitz RB, Chuong CM. Morphogenesis of chicken liver: identification of localized growth zones and the role of beta-catenin/Wnt in size regulation. Dev Biol. 2004;266:109–122. doi: 10.1016/j.ydbio.2003.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Monga SP, Pediaditakis P, Mule K, Stolz DB, Michalopoulos GK. Changes in WNT/beta-catenin pathway during regulated growth in rat liver regeneration. Hepatology. 2001;33:1098–1109. doi: 10.1053/jhep.2001.23786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tan X, Behari J, Cieply B, Michalopoulos GK, Monga SP. Conditional deletion of beta-catenin reveals its role in liver growth and regeneration. Gastroenterology. 2006;131:1561–1572. doi: 10.1053/j.gastro.2006.08.042. [DOI] [PubMed] [Google Scholar]
- 37.Shtutman M, Zhurinsky J, Simcha I, Albanese C, D'Amico M, Pestell R, Ben-Ze'ev A. The cyclin D1 gene is a target of the beta-catenin/LEF-1 pathway. Proc Natl Acad Sci USA. 1999;96:5522–5527. doi: 10.1073/pnas.96.10.5522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sekine S, Gutierrez PJ, Lan BY, Feng S, Hebrok M. Liver-specific loss of beta-catenin results in delayed hepatocyte proliferation after partial hepatectomy. Hepatology. 2007;45:361–368. doi: 10.1002/hep.21523. [DOI] [PubMed] [Google Scholar]
- 39.Tan X, Apte U, Micsenyi A, Kotsagrelos E, Luo JH, Ranganathan S, Monga DK, Bell A, Michalopoulos GK, Monga SP. Epidermal growth factor receptor: a novel target of the Wnt/beta-catenin pathway in liver. Gastroenterology. 2005;129:285–302. doi: 10.1053/j.gastro.2005.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Apte U, Thompson MD, Cui S, Liu B, Cieply B, Monga SP. Wnt/beta-catenin signaling mediates oval cell response in rodents. Hepatology. 2008;47:288–295. doi: 10.1002/hep.21973. [DOI] [PubMed] [Google Scholar]
- 41.Hu M, Kurobe M, Jeong YJ, Fuerer C, Ghole S, Nusse R, Sylvester KG. Wnt/beta-catenin signaling in murine hepatic transit amplifying progenitor cells. Gastroenterology. 2007;133:1579–1591. doi: 10.1053/j.gastro.2007.08.036. [DOI] [PubMed] [Google Scholar]
- 42.Roskams T. Liver stem cells and their implication in hepatocellular and cholangiocarcinoma. Oncogene. 2006;25:3818–3822. doi: 10.1038/sj.onc.1209558. [DOI] [PubMed] [Google Scholar]
- 43.Zucman-Rossi J, Jeannot E, Nhieu JT, Scoazec JY, Guettier C, Rebouissou S, Bacq Y, Leteurtre E, Paradis V, Michalak S, et al. Genotype-phenotype correlation in hepatocellular adenoma: new classification and relationship with HCC. Hepatology. 2006;43:515–524. doi: 10.1002/hep.21068. [DOI] [PubMed] [Google Scholar]
- 44.Moon RT, Kohn AD, De Ferrari GV, Kaykas A. WNT and beta-catenin signalling: diseases and therapies. Nat Rev Genet. 2004;5:691–701. doi: 10.1038/nrg1427. [DOI] [PubMed] [Google Scholar]
- 45.Hussain SZ, Sneddon T, Tan X, Micsenyi A, Michalopoulos GK, Monga SP. Wnt impacts growth and differentiation in ex vivo liver development. Exp Cell Res. 2004;292:157–169. doi: 10.1016/j.yexcr.2003.08.020. [DOI] [PubMed] [Google Scholar]
- 46.Duncan SA. Transcriptional regulation of liver development. Dev Dyn. 2000;219:131–142. doi: 10.1002/1097-0177(2000)9999:9999<::aid-dvdy1051>3.3.co;2-e. [DOI] [PubMed] [Google Scholar]
- 47.Lee CS, Friedman JR, Fulmer JT, Kaestner KH. The initiation of liver development is dependent on Foxa transcription factors. Nature. 2005;435:944–947. doi: 10.1038/nature03649. [DOI] [PubMed] [Google Scholar]
- 48.Zaret KS. Liver specification and early morphogenesis. Mech Dev. 2000;92:83–88. doi: 10.1016/s0925-4773(99)00326-3. [DOI] [PubMed] [Google Scholar]
- 49.Zhou H, Rogler LE, Teperman L, Morgan G, Rogler CE. Identification of hepatocytic and bile ductular cell lineages and candidate stem cells in bipolar ductular reactions in cirrhotic human liver. Hepatology. 2007;45:716–724. doi: 10.1002/hep.21557. [DOI] [PubMed] [Google Scholar]
- 50.Lemaigre FP. Development of the biliary tract. Mech Dev. 2003;120:81–87. doi: 10.1016/s0925-4773(02)00334-9. [DOI] [PubMed] [Google Scholar]
- 51.Li J, Ning G, Duncan SA. Mammalian hepatocyte differentiation requires the transcription factor HNF-4alpha. Genes Dev. 2000;14:464–474. [PMC free article] [PubMed] [Google Scholar]
- 52.Hayhurst GP, Lee YH, Lambert G, Ward JM, Gonzalez FJ. Hepatocyte nuclear factor 4alpha (nuclear receptor 2A1) is essential for maintenance of hepatic gene expression and lipid homeostasis. Mol Cell Biol. 2001;21:1393–1403. doi: 10.1128/MCB.21.4.1393-1403.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Clotman F, Lannoy VJ, Reber M, Cereghini S, Cassiman D, Jacquemin P, Roskams T, Rousseau GG, Lemaigre FP. The onecut transcription factor HNF6 is required for normal development of the biliary tract. Development. 2002;129:1819–1828. doi: 10.1242/dev.129.8.1819. [DOI] [PubMed] [Google Scholar]
- 54.Yamasaki H, Sada A, Iwata T, Niwa T, Tomizawa M, Xanthopoulos KG, Koike T, Shiojiri N. Suppression of C/EBPalpha expression in periportal hepatoblasts may stimulate biliary cell differentiation through increased Hnf6 and Hnf1b expression. Development. 2006;133:4233–4243. doi: 10.1242/dev.02591. [DOI] [PubMed] [Google Scholar]
- 55.Limaye PB, Alarcon G, Walls AL, Nalesnik MA, Michalopoulos GK, Demetris AJ, Ochoa ER. Expression of specific hepatocyte and cholangiocyte transcription factors in human liver disease and embryonic development. Lab Invest. 2008;88:865–872. doi: 10.1038/labinvest.2008.56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Shafritz DA, Oertel M, Menthena A, Nierhoff D, Dabeva MD. Liver stem cells and prospects for liver reconstitution by transplanted cells. Hepatology. 2006;43:S89–S98. doi: 10.1002/hep.21047. [DOI] [PubMed] [Google Scholar]