

Abstract
Weill-Marchesani syndrome (WMS) is a well-characterized disorder in which patients develop eye and skeletal abnormalities. Autosomal-recessive and autosomal-dominant forms of WMS are caused by mutations in ADAMTS10 and FBN1 genes, respectively. Here we report on 13 patients from seven unrelated families from the Arabian Peninsula. These patients have a constellation of features that fall within the WMS spectrum and follow an autosomal-recessive mode of inheritance. Individuals who came from two families and met the diagnostic criteria for WMS were each found to have a different homozygous missense mutation in ADAMTS10. Linkage analysis and direct sequencing of candidate genes in another two families and a sporadic case with phenotypes best described as WMS-like led to the identification of three homozygous mutations in the closely related ADAMTS17 gene. Our clinical and genetic findings suggest that ADAMTS17 plays a role in crystalline lens zonules and connective tissue formation and that mutations in ADAMTS17 are sufficient to produce some of the main features typically described in WMS.
Introduction
Weill-Marchesani syndrome (WMS; MIM 277600) is a rare connective-tissue disorder, mainly characterized in the eye by subluxation of microspherophakic lenses and, consequently, severe myopia and possible glaucoma.1,2 Other symptoms include short stature, brachydactyly, joint stiffness, and heart defects.1,3–6 Both autosomal-recessive (AR) and autosomal-dominant (AD) forms of the disease have been described.7–10 Three mutations in ADAMTS10 (MIM ∗608990, ADAM metallopeptidase with thrombospondin type 1 motif, 10) were reported in two Middle Eastern families with an AR form of the disease and in a single sporadic case of WMS. These include a nonsense mutation (p.R237X) and two splice mutations (810+1 G > A and 1190+1 G > A). All three mutations were predicted to affect the catalytic domain of ADAMTS10, which plays a role in the remodeling of connective tissue.8 Compound heterozygous p.A25T and p.E318X mutations in ADAMTS10 were also identified in a sporadic case of WMS.8,11 In a large family with AD WMS, an in-frame (24 bp) deletion in FBN1 (MIM ∗134797, fibrillin1), which is implicated in Marfan syndrome12,13, was identified, confirming the genetic heterogeneity of this condition.7,12
The purpose of the present study was to describe the clinical and molecular characterization of 13 affected individuals (twelve Saudi and one Yemeni) representing seven unrelated families who presented with varying manifestations of AR WMS. Two homozygous missense mutations were identified in ADAMTS10 in four WMS patients from two different families. Additionally, we identified three different homozygous truncating mutations in ADAMTS17 (MIM ∗607511, ADAM metallopeptidase with thrombospondin type 1 motif, 17), a gene that we mapped by using linkage analysis in four affected members of a consanguineous family. In contrast to the phenotype related to ADAMTS10, the ADAMTS17-related phenotype is best described as WMS-like because none of the patients harboring ADAMTS17 mutations met the full clinical definition of WMS. To the best of our knowledge, this is the first report of ADAMTS17 mutations in humans, and our characterization of this ADAMTS protein supports its role in the development of a connective-tissue disorder that resembles WMS.
Material and Methods
Subjects and Samples
Blood samples for molecular analysis were obtained from participants, including patients (13 individuals) and unaffected family members (seven participants), who provided approved written informed consent in adherence to institutional and international guidelines (RAC# 2070008).
Clinical Assessment
Medical and family history, particularly with regard to consanguinity and ocular and cardiac involvement, was obtained. Ocular history specifically addressed the need for spectacle correction and the age at which any correction occurred, medical or surgical treatment of glaucoma, and crystalline lens extraction. Physical examination of all affected individuals and available unaffected family members included assessment of height, weight, joint flexibility, and the presence of brachydactyly. Ophthalmologic evaluation included assessment of visual acuity, ocular motility status, slit lamp biomicroscopy, and gonioscopy, intraocular pressure measurement by Goldmann applanation tonometry, and dilated examination of the lens, retina, and optic nerve. Biometric ultrasound measurements of axial length, lens thickness, and depth of the anterior chamber were obtained for affected individuals. Goldmann manual kinetic perimetry (Haag Streit International, Koniz-Bern, Switzerland) or automated Humphrey perimetry (Humphrey Field Analyzer II; Carl Zeiss Meditec, Dublin, CA) was obtained in patients with elevated intraocular pressure. Shallow anterior chambers, microspherophakia, and lens subluxation were documented whenever possible by anterior segment photography (Carl Seiss Meditec) and rotating Scheimpflug camera measurements (Pentacam HR, Oculus, Wetzlar, Germany).
Definitions of Clinical Abnormalities
Lenticular Myopia
High myopia (spherical equivalent greater than or equal to −5.00 Diopters in both eyes) without significant (>24.5 mm) enlargement of globe axial length.
Ectopia Lentis
Presence of crystalline lens subluxation either axially or laterally as determined by slit lamp biomicroscopy.
Glaucoma
Glaucomatous damage, as evidenced by characteristic glaucomatous optic nerve cupping of more than 0.6 vertically and/or glaucomatous changes in the visual field as determined by Humphrey automated perimetry or Goldmann static manual perimetry.
Spherophakia
Rounded shape of crystalline lens as evidenced by slit-lamp biomicroscopy exam and an increased antero-posterior diameter (>4 mm) documented by ultrasound.
Short Stature
Abnormally short stature was defined as that in the third percentile or less as determined by growth charts, recently studied and validated, for average height in the region.14
Brachydactyly
Presence of short, stubby fingers in comparison to normal-appearing hands in unaffected family members.
Stiffness of Hand Joints
Inability to flex hand completely, i.e., to make a full fist.
Congenital Heart Abnormalities
History of clinical heart problems detected since childhood.
Automated Sequencing
Purified PCR products covering genes of interest were sequenced with an ABI Prism Big Dye Terminator v3.1 Cycle Sequencing Kit as instructed by the manufacturer. Results were exported in one of several formats for visualization, and sequences were analyzed with SeqMan 6.1 (Lasergene 6 software package).
Screening for Variants in ADAMTS10 and FBN1
PCR products covering all coding exons, along with primers as previously described8, were used for sequencing the entire ORF of ADAMTS10 (NM_030957.2) in patients. Primers covering the 66 coding exons of FBN1 (NM_000138.3) were designed, and amplified products from affected individuals were sequenced (Table S1).
Linkage Analysis
SNP genotyping was performed with Affymetrix GeneChip Human Mapping 10K Arrays. SNP genotypes were called with Affymetrix GCOS 1.4 software and an overall SNP call rate of 95%–99%. Multipoint LOD score calculations were performed with the Allegro module of the Easy Linkage software package15 under the assumption of an autosomal-recessive mode of inheritance with 100% penetrance and disease allele frequency of 0.01%.
Mutation Detection in ADAMTS17
Genomic DNA (10–50 ng) was amplified by PCR in 25 μl reactions via intronic primers that flank each of the 22 exons of ADAMTS17 (NM_139057.2) (Table S2). Standard PCR conditions were as follows: 94°C for 12 min, 30 cycles of 94°C for 45 s, 56°C for 45 s, and 72°C for 45 s, followed by an extension step at 72°C for 10 min. Mutations and variants in ADAMTS17 were identified after PCR products were sequenced. Sequencing new PCR products in both directions confirmed mutations identified in each patient, and the segregation pattern was checked whenever applicable. Sequencing 300 ethnically matched normal controls further confirmed the mutations.
Characterization of Splicing Pattern for c.1721 +1 G > A in ADAMTS17
To ascertain the effect of the c.1721 +1 G > A splice mutation in lymphoblasts from patient WMS-D1, we amplified exons 11-13 and 11-14 from cDNA by using the common forward primer ADAMTS17-x11Frna, 5′-GACACATCCTGCAAGACCAA-3′ and reverse primers ADAMTS17-x13Rrna, 5′-GAAGCTGGGCAGACCCTTG-3′ and ADAMTS17-x14Rrna, 5′-AGGCTTATCGTCAACCAC-3′ to generate wild-type amplicons of 300 bp and 384 bp, respectively.
Expression Analysis of ADAMTS17
First-strand cDNA libraries from multiple human adult and fetal tissues were obtained commercially (Genemed Synthesis, South San Francisco, CA and Capital Biosciences, Rockville, MD, USA). The following primers specific for the two reported ADAMTS17 transcripts were designed: ADAMTS17-x17Frna, 5′-CCGCAGGACTCTTTGTTCAT-3′ and ADAMTS17x19Rrna, 5′-ACTCAGACGCCTCCCAGAT-3′ (used for generation of a 400 bp product specific for transcript ENST00000268070); and ADAMTS17-x14F, 5′-GCCATGTGAACTCTACTGCTC-3′ and ADAMTS17-x16Rrna, 5′-TCCACCACACGGATCCTC-3′ (used for generation of a 300 bp product from transcript ENST00000378898). A 100 bp β-actin fragment was used as a positive control for gene expression. Representative bands were excised from the gel, purified, and sequenced so that the origin of each isoform could be confirmed.
Immunoblot
ADAMTS17 mouse monoclonal antibody that recognizes amino acids 543–651 was obtained commercially (ab58099, Abcam, Cambridge, MA, USA). To assess the ADAMTS17 antibody, we purified proteins from cultures of the following cell lines: normal breast cell line MCF-10 and breast cancer cell lines MCF-7 and T47D. In brief, cells were washed with PBS, then lysed in RIPA lysis buffer (50 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1% NP-40, 0.5% deoxycholate, and 0.1% SDS) containing complete protease inhibitor cocktail (Roche, Indianapolis, IN, USA). The protein concentration was analyzed by Bradford assay. Subsequently, for immunoblotting, 30 μg of each cell lysate was separated on an 8% SDS-PAGE gel and then transferred onto polyvinylidene difluoride membrane (PVDF). The membrane was incubated overnight with ADAMTS17 monoclonal antibody at a 1:500 dilution. The secondary horseradish peroxidase-conjugated rabbit anti-mouse antibody was visualized with a chemiluminescence detection procedure (Amersham Pharmacia Biotech [GE Healthcare Life Science], Piscataway, NJ, USA) according to the manufacturer's protocol, and by exposure to Kodak biomax MR film (Sigma Aldrich, St Louis, MO, USA).
Construction of Tissue Microarray
Tissue microarrays (TMA) were constructed as described previously.16 Two tissue cylinders with a diameter of 0.6 mm were punched from archival paraffin blocks and transferred into a recipient paraffin block via a semi-automatic precision instrument (Beecher Instruments, Silver Spring, MD, USA).
Immunohistochemistry
To validate the ADAMTS17 antibody, we used ADAMTS17 recombinant protein (Q01) specific for ADAMTS17 (NP_620688.2, amino acids 543–650) (Abnova, Taiwan) on MCF-7 breast cancer cell line and a control tissue microarray comprising various adult human normal tissues (colon, liver, kidney, pancreas, thyroid, lymph node, spleen, heart, lung, smooth muscle, ovary, and testis). Human TMA slides and mouse E14.5 embryo slides were incubated overnight in a 1:6000 dilution of anti-ADAMTS17 (ab58099, Abcam), and the Dako Envision Plus System kit was used as the secondary detection system with DAB as chromogen. Endogenous peroxidase activity was quenched with 3% hydrogen peroxidase in methanol. Endogenous biotin was blocked, and all slides were counterstained with hematoxylin, dehydrated, cleared, and coverslipped with premount. Cell line blocks of MCF-7 and T47D and tumor tissue samples of ovarian and colon cancers served as a positive control. Finally, two types of negative controls were used: the first was the exclusion of the primary antibody, and the second was a peptide competition assay. For the latter, ADAMTS17 antibody was preincubated with the ADAMTS17 recombinant protein (Q01) at a 1:2 ratio (w/w) for 1 hr at room temperature prior to staining. Tissues were scored on a four-tier system: 0, absence of staining; and 1, 2, and 3, weak, moderate, and strong staining, respectively.
Results
Clinical Manifestations
A total of 13 patients from seven unrelated families, including three with single affected individuals, were investigated. There were five cases (two families and one sporadic case) with complete features of AR WMS (lenticular myopia, ectopia lentis, spherophakia, short stature, brachydactyly, and decreased joint flexibility) and eight cases (two families and two sporadic cases) who had the previously mentioned WMS manifestations except for brachydactyly and decreased joint flexibility and who were labeled as having an AR WMS-like condition (Figure 1). A summary of the main ophthalmological and clinical features is presented in Table 1.
Figure 1.
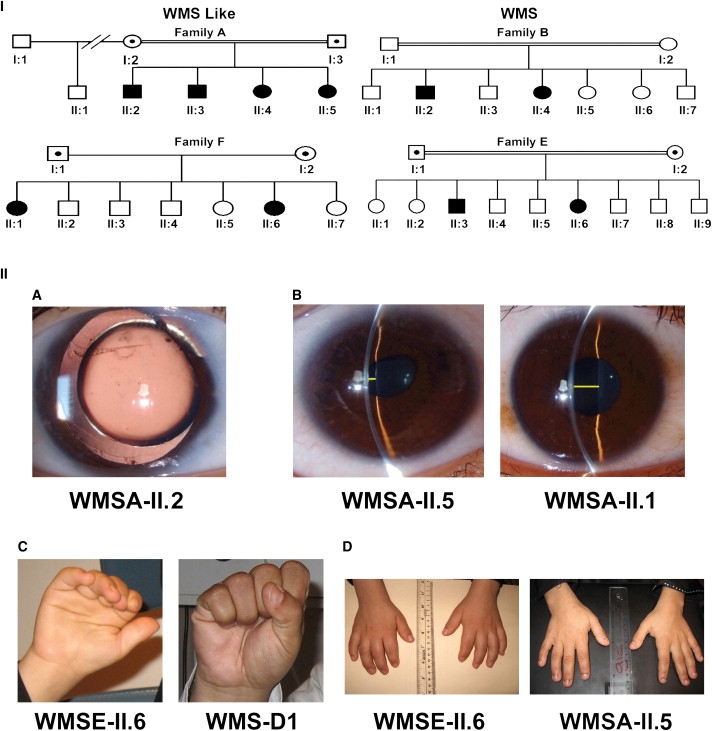
Family Pedigrees and Clinical Signs
(I) The pedigrees of the four families with more than one affected individual. Families A and F show autosomal-recessive inheritance and a WMS-like condition, and families B and E are classical autosomal-recessive WMS families.
(II) (A) Slit-lamp photograph of the right eye of patient WMSA-II.2 displaying clear microspherophakia and showing the equator of an apparently smaller-than-normal crystalline lens within a dilated pupil. There is evidence of prior glaucoma surgery, including a peripheral iridectomy and some peripheral iris synechiae. (B) Comparison of a very shallow anterior chamber highlighted by a yellow line outlining the space between the lens and the inner surface of the cornea on patient WMSA-II.5 and a normal deep anterior chamber of her unaffected half-sibling, WMSA-II.1. (C) Stiffness of joints is illustrated by the inability of patient WMSE-II.6 to make a fist; in contrast, patient WMS-D1 (not displayed on pedigrees, but corresponding to patient number five in Table 1) is making a normal full fist. (D) The hands of patient WMSE-II.6 with short, stubby fingers (brachydactyly) in comparison with the normal hands of patient WMSA-II.5.
Table 1.
Summary of Clinical Features of Affected Individuals with WMS and WMS-like Syndrome
| Number | Family ID | Patient ID | Agea | Gender | Myopia Levelb | Highest IOP in mmHgc | Glaucomatous Nerve Damage | Height in cm | Percentiled | Brachydactyly | Joint Stiffness | Congenital Heart Abnormalitiese | Average Axial Lengthf | Shallow Anterior Chamber | Peripheral Iris Synechiae | Gene/Mutation |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WMS-like | ||||||||||||||||
| 1 | Family A | II.2 | 16 | M | −7.8 | NA | Yes | 160 | 0.25 | No | No | No | 23.0 | Yes | Yes | ADAMTS17/ c.2458_2459insG |
| 2 | Family A | II.3 | 15 | M | −12.0 | 21 | No | 150 | <0.25 | No | No | No | 22.8 | No | No | ADAMTS17/ c.2458_2459insG |
| 3 | Family A | II.4 | 12 | F | −11.6 | 42 | Yes | 127 | <0.03 | No | No | No | 22.5 | Yes | Yes | ADAMTS17/ c.2458_2459insG |
| 4 | Family A | II.5 | 10 | F | −10.5 | 23 | No | 118 | <0.03 | No | No | No | 21.5 | Yes | Yes | ADAMTS17/ c.2458_2459insG |
| 5 | Family D | WMS-D1 | 33 | M | −11.9 | 33 | Yes | 155 | 0.03 | No | No | No | 21.3 | Yes | No | ADAMTS17/ c.1721+1G > A |
| 6 | Family F | II.1 | 30 | F | −8.6 | 41 | Yes | 145 | 0.05 | No | No | No | 21.7 | Yes | Yes | ADAMTS17/ c.760 C > T |
| 7 | Family F | II.6 | 11 | F | −5.5 | 40 | No | 128 | <0.05 | No | No | No | 20.7 | Yes | Yes | ADAMTS17/ c.760 C > T |
| 8 | Family G | WMS-G1 | 36 | F | −9.0 | 60 | No | 134 | <0.03 | No | No | Yes | 21.9 | Yes | Yes | No mutations in ADAMTS10, ADAMTS17 and FBN1 |
| WMS | ||||||||||||||||
| 9 | Family B | II.2 | 17 | M | −14.4 | 35 | No | 142 | <0.03 | Yes | Yes | No | 22.4 | Yes | No | ADAMTS10/ c.1553 G > A |
| 10 | Family B | II.4 | 14 | F | −8.9 | 19 | No | 133 | <0.03 | Yes | Yes | No | 21.4 | Yes | No | ADAMTS10/ c.1553 G > A |
| 11 | Family E | II.3 | 22 | M | −19.1 | 23 | No | 148 | <0.03 | Yes | Yes | No | 23.7 | Yes | No | ADAMTS10/ c.2098 G > T |
| 12 | Family E | II.6 | 13 | F | −11.0 | 27 | No | 132 | <0.03 | Yes | Yes | Yes | 20.1 | Yes | No | ADAMTS10/ c.2098 G > T |
| 13 | Family C | II.1 | 70 | M | 13.8 | 30 | Yes | 142 | <0.03 | Yes | Yes | No | 21.4 | Yes | No | No mutations in ADAMTS10, ADAMTS17 and FBN1 |
Family and Patient ID: family and patient identification as per pedigree (all families displayed autosomal-recessive inheritance).
Age: in years at the time of the study. M, male; F, female.
Average spherical equivalent in diopters for both eyes.
Maximal IOP in mmHg in either eye.
Percentile as per Saudi growth charts.14
Patients WMEII.6 and WMS-G1 had congenital cardiac valvular abnormalities as per clinical history; they required catheterism and valvuloplasty at ages 4 and 6, respectively.
Average eye globe axial length from both eyes, measured by ultrasound.
Regarding height, two patients were at or below the 25th percentile, two were at or below the fifth percentile, and the rest were at or below the third percentile according to growth tables recently established for the Saudi population.14
Mean globe axial length, antero-posterior lens diameter, and anterior chamber depth for both eyes were 21.9 mm (range 20.1–23.0 mm), 5.1 mm (range 4.2–7.6 mm), and 2.1 mm (range 1.1–3.1), respectively.
Glaucomatous damage to the optic nerve and impairment of the visual field were documented in five patients, elevated intraocular pressures were documented in 11 patients, and clinically shallow anterior chambers were documented in all except one patient.
Mutations and Variants Identified in ADAMTS10 and FBN1
Two missense homozygous mutations in the ADAMTS10 gene, c.1553 G > A (p.G518D) and c.2098 G > T (p.G700C) (accession NP_112219.2), were identified in patients from families B and E, respectively, and were absent in 240 normal controls. Moreover, a number of previously reported coding and noncoding variants were identified by sequencing. These include c.401 C > G (p.T134S, rs7255721), c.2283 T > C (p.P761, rs4476282), c.2423 T > G (p.L808R, rs10420313), c.2584 G > T (p.A862S, rs10418929), c.435+24 A > G (rs7260282), and c.1085-47 A > G (rs7247944) in the ADAMTS10 gene. In the FBN1 gene, the following noncoding variants were identified; c.1960+76 G > A (rs17361868), c.4748-77 T > G (rs55694948), c.6037+54 T > A (rs2303502), and 6997+17 C > G (rs363832) (accession NP_000129.2).
Mutations in ADAMTS17 in Patients with Partial WMS
Multipoint linkage analysis of family A, including unaffected parents, all four affected siblings, and the unaffected maternal half sibling, identified a disease locus on chromosome 15q26.3 (Ensembl cytogenetic band) with a maximum logarithm of odds (LOD) score of 3 (Figure 2). The linkage region spanned 2.05 Mb, and IGF1R (NM_000875), SYNM (NM_145728), TTC23 (NM_022905), MEF2A (NM_001130926), LYSMD4 (NM_152449), and ADAMTS17 (NM_139057.2) from within this interval were selected as plausible candidates. A homozygous 1 bp insertion in exon 18, c.2458_2459insG (p.E820GfsX23; accession NP_620688.2) of ADAMTS17 was identified. This out-of-frame insertion fully segregated with the phenotype in the family used for linkage analysis and was consistent with an autosomal-recessive mode of inheritance (Figure 2).
Figure 2.
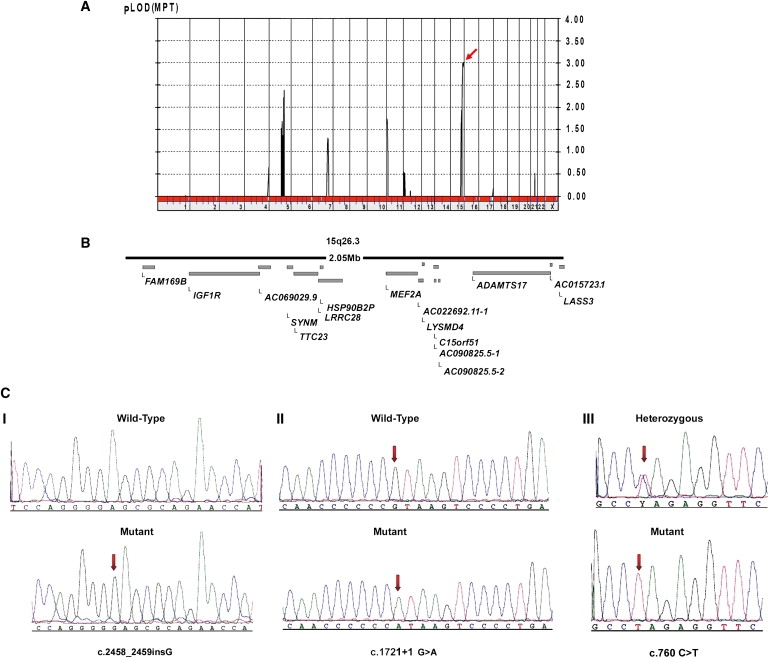
ADAMTS17 Mutations in Patients with Autosomal-Recessive WMS-like Syndrome
(A) Linkage analysis via the Allegro module reveals a LOD score of 3 on chromosome 15, as indicated by an arrow.
(B) Linkage interval of 2.05 Mb as annotated in the Ensembl genome database.
(C) Sequence chromatograms of homozygous mutations encountered in ADAMTS17. (I) c.2458_2459insG (p.E820GfsX23) in exon 18. (II) c.1721+1 G > A splice mutation in intron 12. (III) c.760 C > T (Q254X) in exon 4. Arrows point to mutation sites.
Screening the entire ORF of the ADAMTS17 gene in similar WMS-like individuals identified a homozygous truncating mutation, c.760 C > T (p.Q254X), in the two affected members of family F (both parents were heterozygous), and a third homozygous mutation, c.1721+1 G > A, was identified in a sporadic case (WMS-D1). These mutations were absent in 300 ethnically matched normal controls (Figure 2).
mRNA analysis with two primer sets flanking exon 12 indicated the presence of three aberrant transcripts in patient WMS-D1 lymphoblasts as a result of the c.1721+1 G > A mutation. The first transcript resulted from exon 12 skipping. The second and third transcripts retained exon 12 but also included different segments of intron 12 (Figure 3). All three transcripts harbored a frameshift and are predicted to produce truncated proteins of 579 and 574 amino acids, respectively.
Figure 3.
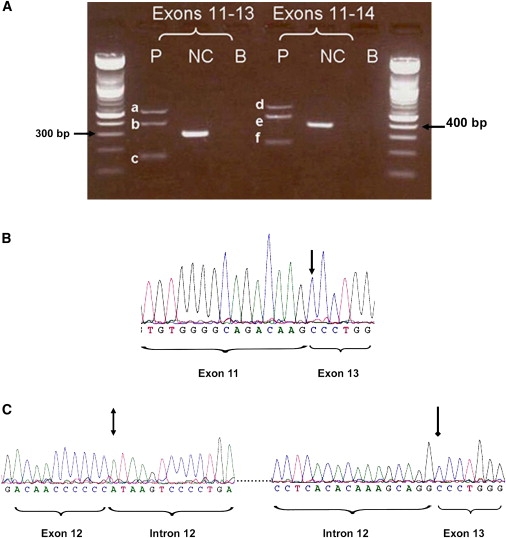
Splicing Patterns of the c.1721 +1 G > A Mutation in ADAMTS17 in Patient WMS-D1
(A) PCR products of cDNA from the patient (P) and a normal control (NC). Two sets of primers were used for the PCR. The patient cDNA gave three PCR products of different sizes with both sets of primers, whereas the NC gave only the wild-type expected product size.
(B) The sequence chromatogram of band “c” from PCR using the forward primer confirms exon 12 skipping. An arrow points to the end of exon 11 and the beginning of exon 13.
(C) Sequence chromatogram of PCR band “a.” A double-headed arrow points to the end of exon 12 and the beginning of intron 12, and a single-headed arrow points to the end of the “spliced-in” part of intron 12 and the beginning of exon 13.
Variants Identified in ADAMTS17
A number of coding and noncoding variants were identified in ADAMTS17. These include 19 intronic variants, 7 synonymous and 3 non-synonymous variants (Table S3). Some of these variants are likely to represent previously undescribed SNPs.
Expression Analysis of ADAMTS17
There are two reported isoforms of ADAMTS17 with different initiation codons: a transcript that spans 22 exons and codes for 1095 amino acids and a shorter transcript of 16 exons that codes for 502 amino acids. These will be designated in the text as isoforms “a” and “b,” respectively. Expression of the two transcripts was investigated in cDNA libraries from different adult and fetal tissues (Figure 4). The lung, brain, whole eye, and retina showed high expression of both transcripts. Isoform “a” showed weaker expression in the heart, kidney, and skeletal muscle, whereas bone marrow lacked expression of this transcript. Expression of isoform “b” in the kidney, bone marrow, and skeletal muscle was weak. Both transcripts demonstrated high expression of ADAMTS17 in the fetal brain, heart, kidney, and whole eye, whereas expression was weak in the liver.
Figure 4.
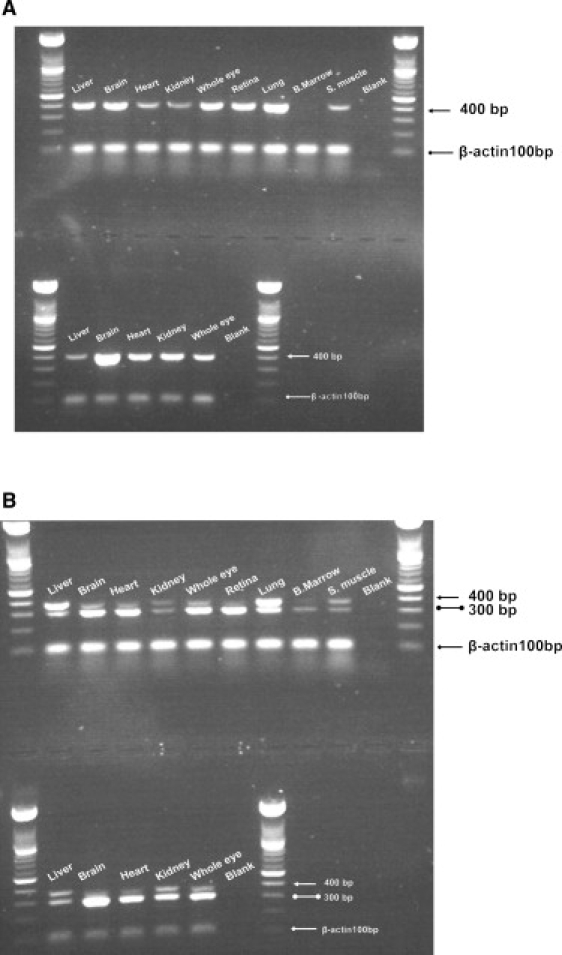
Expression of ADAMTS17 in cDNA Libraries
Expression of ADAMTS17 in cDNA libraries from different human adult (top panel) and fetal (bottom panel) tissues.
(A) An arrow points to a 400 bp ADAMTS17 product specific for isoform “a.”
(B) A double-ball-headed arrow indicates a 300 bp ADAMTS17 product specific for isoform “b.” A Single-headed arrow points to a third transcript detected.
Our analysis using primers specific for transcript “b” uncovered a previously uncharacterized third transcript where there was a 69 bp inclusion from intron 15; this was confirmed by sequencing of the PCR band. This exon might represent an alternatively spliced exon originating from transcript “b.” This transcript showed high expression in adult liver and lungs and weak expression in the remaining adult and fetal tissues studied and could represent one of the ADAMTS17 minor isoforms.
Tissue Distribution of ADAMTS17
ADAMTS17 is a member of the secreted metalloproteinase family of proteins that are believed to bind to the extra cellular matrix (ECM). Immunohistochemistry for which a mouse monoclonal ADAMTS17 antibody was used on different normal and cancer human tissues showed strong staining in the spleen and testis and moderate staining in the liver, kidney, placenta, and lymph node, whereas the expression was weak in the heart, lung, colon, ovary and uterine wall smooth muscle. The pancreas and thyroid gland lacked expression of ADAMTS17 (Table S4). Importantly, eye sections from E14.5 mouse embryos showed some diffuse expression of ADAMTS17 within the area of a future ciliary body (Figure S1).
Discussion
This study reports the identification of ADAMTS17 as another WMS gene, for which three mutations were identified in seven patients who had the typical WMS ocular manifestations in addition to short stature. The three mutations consist of a 1 bp insertion in exon 18, which is predicted to produce a truncated protein of 841 amino acids, a splice-site mutation that completely abolished the consensus donor sequence of intron 12 and produced three truncated transcripts, and a nonsense mutation in exon 4. The c.2458_2459insG mutation and the splice mutation c.1721+1 G > A are predicted to generate a truncated protein that retains the catalytic domain but lacks the central and/or C-terminal TS repeats, which are thought to be essential for ECM binding.17 On the other hand, the nonsense mutation c.760 C > T in exon 4 would create a truncated protein that lacks the catalytic domain in addition to the central and C-terminal TS repeats. Interestingly, RNA analysis on lymphoblasts from patients suggests that all of these mutations created stable mRNA transcripts. c.760 C > T and c.1721+1 G > A mutations affect all known ADAMTS17 isoforms, whereas c.2458_2459insG in exon 18 only affects isoform “a.” The nature of these mutations suggests that they are pathogenic, and we confirmed their absence in 600 ethnically matched normal control chromosomes.
Proteases have important roles in ocular physiology and pathophysiology. These proteases include caspases, calpains, matrix metalloproteases, ADAMs, ADAMTSs, those in the ubiquitin-protesome pathway (UPP), tissue plasminogen activator, and secretase.18 ADAMTS is a family of 19 homologous secreted metalloproteinases that are believed to be anchored to the ECM via their C-terminal thrombospondin (TS) repeats.19 These proteins undergo N-terminal processing and removal of the prodomain during their transit through the membrane of the endoplasmic reticulum; they also undergo C-terminal processing that affects substrate specificity and ECM binding.20 Previous characterization of the ADAMTS proteins showed their involvement in different cellular mechanisms17,19, and some of these metalloproteinases have been implicated in a number of connective-tissue disorders and ectopia lentis.8,21,22 ADAMTS17 is a member of the same protein family as ADAMTS10, which is mutated in AR WMS.8,17 However, only the structure of ADAMTS17 was characterized, and its putative role and substrate remain to be elucidated.
The previously reported ADAMTS10 mutations identified in WMS patients disrupted the catalytic domain of the enzyme, and it was suggested that the catalytic domain of ADAMTS10 could play a role in connective tissue remodeling and that this might explain the pathogenic mechanism of AR WMS.8 In contrast, two of our mutations in ADAMTS17 resulted in transcripts that retained the catalytic domain but lacked the C-terminal TS repeats (Figure 5), suggesting that ADAMTS17 might play more of a structural role in the assembly or integrity of ECM, similar to what was proposed for ADAMTSL4.22
Figure 5.
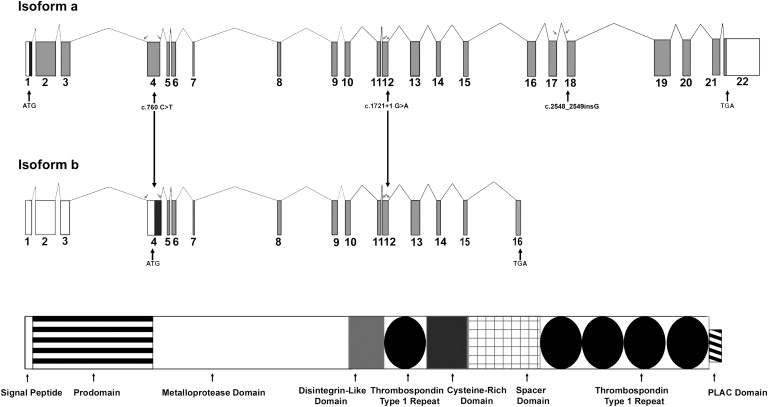
The Two Major Isoforms of the ADAMTS17 Gene
Isoform “a” based on Ensembl transcript (ENST00000268070) and isoform “b” is based on Ensembl transcript (ENST00000378898). The protein domains are illustrated, and the position of each of the three mutations is specified. Arrows point to the position of primers flanking exons 4,12, and 18.
Our patients with truncating mutations in ADAMTS17 had the typical eye anomalies described in WMS (these anomalies include spherophakia, ectopia lentis, high myopia, and glaucoma secondary to shallow anterior-chamber angles) but lacked joint stiffness, brachydactyly, and cardiac valvular abnormalities. Expression analysis of all known normal transcripts of ADAMTS17 on cDNA libraries from multiple fetal and adult tissues confirmed the previously reported high expression in the lungs19 and showed high expression in the whole eye and retina, which is consistent with the eye abnormalities observed in these patients. Moreover, ADAMTS17 expression was observed in the ciliary body of E.14.5 mouse embryos, which might explain the role of this protein in zonule development. Because we did not examine the expression of ADAMTS17 in developing limb buds, we cannot determine whether the lack of hand manifestations is due to a lack of expression or whether it results from redundant expression of other ADAMTS proteins.
We found two homozygous missense ADAMTS10 mutations, c.1553 G > A and c.2098 G > T, in patients with autosomal-recessive inheritance and full WMS manifestations, including joint stiffness and brachydactyly. This confirms previous reports about the involvement of ADAMTS10 in this condition and the genetic heterogeneity of the disorder already pointed out by Faivre et al.23 Because we found ADAMTS17 mutations only in patients with incomplete WMS manifestations, there is a possibility that different members of the ADAMTS family of proteins could be responsible for different subtypes of the syndrome. In addition, some ADAMTS proteins could have overlapping functions and interactions in such a way that a mutation in one of them can be complemented by the function of others or affect their activity in a limited fashion and thus cause a partial presentation of symptoms. However, ADAMTS17 mutations seem to be capable of producing only the ophthalmic manifestations and short stature encountered in WMS.
To the best of our knowledge, this is the first report of homozygous mutations of ADAMTS17 in patients with ectopia lentis and short stature. This highlights the importance of the ADAMTS protein family in connective-tissue biology, particularly on the crystalline lens zonules because the main impact in the eye seems to be through abnormalities of the elastic zonules. This in turn causes lens subluxation, spherophakia, and high myopia and causes the anterior chamber to become shallow, which results in glaucoma. ADAMTS17 and ADAMTS10 have structural similarities, but on the basis of the level of conservation of the protease active site19,24, they belong to two different phylogenetic nodes, and so far we do not have an explanation for the fact that ADAMTS17 seems to cause less systemic abnormalities. ADAMTS10 seems to be more highly expressed than ADAMTS17 in the heart, which could partially explain why patients with ADAMTS17 mutations lacked congenital cardiac abnormalities. Because no echocardiographic studies were done, we cannot completely rule out subclinical cardiac abnormalities in these individuals. Among our cohort, we were not able to find any variants in the ADAMTS17, ADAMTS10, or FBN1 genes that segregated with the disease in two patients. One of them (WMS-G1) had incomplete features of WMS without brachydactyly or joint stiffness. The second patient (WMSC-II.1) seemed to have all the features, but the disease had a late onset, and lens subluxation and glaucoma appeared in middle age instead of childhood or adolescence. Although we cannot exclude deep intronic or regulatory-element mutations in these genes, our results suggest that additional loci might have caused WMS or WMS-like manifestations in these two patients.
Verloes et al. suggested the acronym GEMSS (glaucoma, ectopia lentis, microspherophakia, short stature, and stiffness of joints) for what they considered to be a dominant form of WMS.25 We believe that rather than constraining the definition of the syndrome to a particular type of hereditary transmission, it might be more beneficial at this stage, when more mutations are being discovered, to reach better standardization and a consensus regarding what manifestations constitute WMS, as was done with Marfan syndrome.26 It is worth highlighting that lenticular myopia, ectopia lentis, glaucoma, spherophakia, and short stature were present in patients with mutations in ADAMTS10 and ADAMTS17 as well as in the two patients with no identified mutations in the three genes.
Although not all patients developed glaucoma, most had shallow anterior chambers secondary to anterior lens subluxation and elevated intraocular pressures at some point, which would put them at risk of developing secondary angle-closure glaucoma. The average axial globe length was shorter than usual. This confirms the lenticular component of the high myopia encountered in these patients and also suggests that some of these patients might be more predisposed to secondary angle-closure glaucoma due to the combination of a short globe and anterior crystalline lens subluxation's crowding the anterior chamber angle and blocking the trabecular meshwork.
Six of the eight patients with WMS-like manifestations had peripheral irido-corneal adhesions, some of which were noted before any intraocular surgical procedure was undertaken. It is possible that they could have formed as a result of intermittent anterior lens movement and repeated contact between the iris and the cornea, but we cannot rule this out as another possible clinical ophthalmological feature in some patients with this condition. We did not observe these kinds of irido-corneal adhesions in any of the patients with classical WMS.
For phenotype classification, we prefer to describe the lens abnormality as spherophakia instead of microspherophakia because the smaller equatorial diameter was not clearly seen in all of our patients, whereas the globular shape of the lens was easier to identify by slit-lamp biomicroscopy and to document consistently by ultrasound measurements. Besides, it is possible that the lens is not actually anatomically small but that the globular shape and the decreased equatorial diameter are secondary phenomena due to abnormal laxity of the crystalline lens zonules. Although we did not observe broken or missing zonules, we assume that there might have been significant zonular elongation, particularly in those individuals with significant lens displacement or clear visualization of the whole equator of the lens after dilation.
In summary, we have identified null mutations in ADAMTS17 as a cause of an autosomal-recessive WMS-like syndrome. We have confirmed abnormalities in ADAMTS10 in typical cases of WMS. Lenticular myopia, ectopia lentis, glaucoma (secondary to anterior lens subluxation and angle closure), spherophakia, and short stature were common denominators in patients with complete and incomplete manifestations of WMS. In our cohort of patients with these manifestations, ADAMTS17 and ADAMTS10 mutations were present in more than 80% of cases. Further work involving other ethnic groups and ectopia lentis patients might clarify the role that different mutations play in the pathogenesis of this condition.
Supplemental Data
Supplemental data include one figure and four tables and can be found with this article online at http://www.cell.com/AJHG/.
Supplemental Data
Web Resources
The URLs for data presented herein are as follows:
GenBank, http://www.ncbi.nlm.nih.gov/Genbank
Ensembl project genome database, http://www.ensembl.org/index.html
Online Mendelian Inheritance in Man (OMIM), http://www.ncbi.nlm.nih.gov/omim
Single Nucleotide Polymorphism database, http://www.ncbi.nlm.nih.gov/SNP/
UCSC Genome Browser, http://genome.ucsc.edu/
Accession Numbers
Submitter SNP (ss) numbers for variants identified in this study are ss161151830, ss161151831, ss161151832, ss161151833.
Acknowledgments
The authors would like to thank Valorie Balde, Zakia Shinwari, Ayodele. A. Alaiya, and Tarfa Al Sheddi for all their technical assistance; Faisal Al Hamidi for his help in patient recruitment; members of the photography department of the King Khaled Eye Specialist Hospital (KKESH) for their detailed documentation of the clinical signs; Mohamad Rajab, Shamsa Al Enazi, and Dorota Moniez for their help with DNA Sequencing; Batoul Baz and Salma Wakil of the genotyping core facility for their help in processing the 10K affymetrix chip; and Futwan Al Mohanna for his help and efforts. This work was approved and funded by King Faisal Specialist Hospital and Research Centre (RAC# 2070008). and KKESH (RP# 0664-P).
References
- 1.Wright K.W., Chrousos G.A. Weill-Marchesani syndrome with bilateral angle-closure glaucoma. J. Pediatr. Ophthalmol. Strabismus. 1985;22:129–132. doi: 10.3928/0191-3913-19850701-04. [DOI] [PubMed] [Google Scholar]
- 2.Chu B.S. Weill-Marchesani syndrome and secondary glaucoma associated with ectopia lentis. Clin. Exp. Optom. 2006;89:95–99. doi: 10.1111/j.1444-0938.2006.00014.x. [DOI] [PubMed] [Google Scholar]
- 3.Giordano N., Senesi M., Battisti E., Mattii G., Gennari C. Weill-Marchesani syndrome: Report of an unusual case. Calcif. Tissue Int. 1997;60:358–360. doi: 10.1007/s002239900243. [DOI] [PubMed] [Google Scholar]
- 4.Rennert O.M. The Marchesani syndrome. A brief review. Am. J. Dis. Child. 1969;117:703–705. doi: 10.1001/archpedi.1969.02100030705016. [DOI] [PubMed] [Google Scholar]
- 5.Kojuri J., Razeghinejad M.R., Aslani A. Cardiac findings in Weill-Marchesani syndrome. Am. J. Med. Genet. A. 2007;143A:2062–2064. doi: 10.1002/ajmg.a.31861. [DOI] [PubMed] [Google Scholar]
- 6.Herrera J., Morales M. Weill-Marchesani syndrome. Rev. Chil. Pediatr. 1986;57:571–572. [PubMed] [Google Scholar]
- 7.Faivre L., Gorlin R.J., Wirtz M.K., Godfrey M., Dagoneau N., Samples J.R., Le Merrer M., Collod-Beroud G., Boileau C., Munnich A. In frame fibrillin-1 gene deletion in autosomal dominant Weill-Marchesani syndrome. J. Med. Genet. 2003;40:34–36. doi: 10.1136/jmg.40.1.34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dagoneau N., Benoist-Lasselin C., Huber C., Faivre L., Megarbane A., Alswaid A., Dollfus H., Alembik Y., Munnich A., Legeai-Mallet L. ADAMTS10 mutations in autosomal recessive Weill-Marchesani syndrome. Am. J. Hum. Genet. 2004;75:801–806. doi: 10.1086/425231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Young I.D., Fielder A.R., Casey T.A. Weill-Marchesani syndrome in mother and son. Clin. Genet. 1986;30:475–480. doi: 10.1111/j.1399-0004.1986.tb01914.x. [DOI] [PubMed] [Google Scholar]
- 10.Evereklioglu C., Hepsen I.F., Er H. Weill-Marchesani syndrome in three generations. Eye. 1999;13:773–777. doi: 10.1038/eye.1999.226. [DOI] [PubMed] [Google Scholar]
- 11.Kutz W.E., Wang L.W., Dagoneau N., Odrcic K.J., Cormier-Daire V., Traboulsi E.I., Apte S.S. Functional analysis of an ADAMTS10 signal peptide mutation in Weill-Marchesani syndrome demonstrates a long-range effect on secretion of the full-length enzyme. Hum. Mutat. 2008;29:1425–1434. doi: 10.1002/humu.20797. [DOI] [PubMed] [Google Scholar]
- 12.Dietz H.C., Saraiva J.M., Pyeritz R.E., Cutting G.R., Francomano C.A. Clustering of fibrillin (FBN1) missense mutations in Marfan syndrome patients at cysteine residues in EGF-like domains. Hum. Mutat. 1992;1:366–374. doi: 10.1002/humu.1380010504. [DOI] [PubMed] [Google Scholar]
- 13.Dietz H.C., Pyeritz R.E. Mutations in the human gene for fibrillin-1 (FBN1) in the Marfan syndrome and related disorders. Hum. Mol. Genet. 1995;4:1799–1809. doi: 10.1093/hmg/4.suppl_1.1799. [DOI] [PubMed] [Google Scholar]
- 14.El Mouzan M.I., Al Herbish A.S., Al Salloum A.A., Foster P.J., Al Omar A.A., Qurachi M.M., Kecojevic T. Comparison of the 2005 growth charts for Saudi children and adolescents to the 2000 CDC growth charts. Ann. Saudi Med. 2008;28:334–340. doi: 10.5144/0256-4947.2008.334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lindner T.H., Hoffmann K. easyLINKAGE: A PERL script for easy and automated two-/multi-point linkage analyses. Bioinformatics. 2005;21:405–407. doi: 10.1093/bioinformatics/bti009. [DOI] [PubMed] [Google Scholar]
- 16.Bavi P., Abubaker J., Hussain A., Sultana M., Al-Dayel F., Uddin S., Al-Kuraya K.S. Reduced or absent cyclin H expression is an independent prognostic marker for poor outcome in diffuse large B-cell lymphoma. Hum. Pathol. 2008;39:885–894. doi: 10.1016/j.humpath.2007.10.014. [DOI] [PubMed] [Google Scholar]
- 17.Porter S., Clark I.M., Kevorkian L., Edwards D.R. The ADAMTS metalloproteinases. Biochem. J. 2005;386:15–27. doi: 10.1042/BJ20040424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wride M.A., Geatrell J., Guggenheim J.A. Proteases in eye development and disease. Birth Defects Res. C Embryo Today. 2006;78:90–105. doi: 10.1002/bdrc.20063. [DOI] [PubMed] [Google Scholar]
- 19.Cal S., Obaya A.J., Llamazares M., Garabaya C., Quesada V., Lopez-Otin C. Cloning, expression analysis, and structural characterization of seven novel human ADAMTSs, a family of metalloproteinases with disintegrin and thrombospondin-1 domains. Gene. 2002;283:49–62. doi: 10.1016/s0378-1119(01)00861-7. [DOI] [PubMed] [Google Scholar]
- 20.Gao G., Westling J., Thompson V.P., Howell T.D., Gottschall P.E., Sandy J.D. Activation of the proteolytic activity of ADAMTS4 (aggrecanase-1) by C-terminal truncation. J. Biol. Chem. 2002;277:11034–11041. doi: 10.1074/jbc.M107443200. [DOI] [PubMed] [Google Scholar]
- 21.Colige A., Sieron A.L., Li S.W., Schwarze U., Petty E., Wertelecki W., Wilcox W., Krakow D., Cohn D.H., Reardon W. Human Ehlers-Danlos syndrome type VII C and bovine dermatosparaxis are caused by mutations in the procollagen I N-proteinase gene. Am. J. Hum. Genet. 1999;65:308–317. doi: 10.1086/302504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ahram D., Sato T.S., Kohilan A., Tayeh M., Chen S., Leal S., Al-Salem M., El-Shanti H. A homozygous mutation in ADAMTSL4 causes autosomal-recessive isolated ectopia lentis. Am. J. Hum. Genet. 2009;84:274–278. doi: 10.1016/j.ajhg.2009.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Faivre L., Dollfus H., Lyonnet S., Alembik Y., Megarbane A., Samples J., Gorlin R.J., Alswaid A., Feingold J., Le Merrer M. Clinical homogeneity and genetic heterogeneity in Weill-Marchesani syndrome. Am. J. Med. Genet. A. 2003;123A:204–207. doi: 10.1002/ajmg.a.20289. [DOI] [PubMed] [Google Scholar]
- 24.Huxley-Jones J., Apte S.S., Robertson D.L., Boot-Handford R.P. The characterisation of six ADAMTS proteases in the basal chordate Ciona intestinalis provides new insights into the vertebrate ADAMTS family. Int. J. Biochem. Cell Biol. 2005;37:1838–1845. doi: 10.1016/j.biocel.2005.03.009. [DOI] [PubMed] [Google Scholar]
- 25.Verloes A., Hermia J.P., Galand A., Koulischer L., Dodinval P. Glaucoma-lens ectopia-microspherophakia-stiffness-shortness (GEMSS) syndrome: A dominant disease with manifestations of Weill-Marchesani syndromes. Am. J. Med. Genet. 1992;44:48–51. doi: 10.1002/ajmg.1320440112. [DOI] [PubMed] [Google Scholar]
- 26.De Paepe A., Devereux R.B., Dietz H.C., Hennekam R.C., Pyeritz R.E. Revised diagnostic criteria for the Marfan syndrome. Am. J. Med. Genet. 1996;62:417–426. doi: 10.1002/(SICI)1096-8628(19960424)62:4<417::AID-AJMG15>3.0.CO;2-R. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.