

Abstract
Pollen allergy is characterized by a TH2-biased immune response to pollen-derived allergens. However, pollen-exposed epithelia do not encounter pure allergen but rather a plethora of protein and non-protein substances. We demonstrated that pollen liberate lipids with chemical and functional similarities to leukotriens and prostaglandins - the pollen associated lipid mediators (PALMs). To date, two main groups of PALMs have been characterized: The immunostimulatory PALMs activating innate immune cells such as neutrophils and eosinophils, and the immunomodulatory E1-phytoprostanes blocking IL-12 production of dendritic cells, resulting in the preferential induction of TH2 responses. This article reviews our work in the field of PALMs and their effects on cells of the innate and adoptive immune system. From recent results a general picture starts to emerge in which PALMs (and possibly other pollen-associated substances) may - independently from protein allergens - propagate an overall TH2 favoring micromilieu in pollen exposed tissue of predisposed individuals.
Background
Atopic diseases are characterized by a predominance of T helper cell type 2 (TH2) biased immune responses to environmental allergens. It is well established that allergen specific TH2 cells are the key orchestrators of allergic reactions, initiating and propagating inflammation through the release of a number of TH2 cytokines. While the importance of TH2 cells in allergy is well accepted, little is known about the mechanisms that control the initial TH2 polarization in response to exogenous allergens. While for some aeroallergens, foremost house dust mite Der p 1, several intrinsic TH2 adjuvant effects have been reported [1-3], most major pollen allergens seem to lack such characteristics.
A hallmark in the elucidation of adjuvant factors from pollen was the discovery that pollen release NADPH oxidases which increase reactive oxygen species in lung epithelium thereby promoting neutrophil recruitment and boosting allergic airway inflammation. In contrast, challenge with Amb a 1, the major ragweed allergen alone, did not result in robust airway inflammation [4].
As link between innate and adaptive immune system, dendritic cells (DCs) play a pivotal role in sensing environmental danger signals such as bacterial or viral products, and in mounting a T cell-mediated immune response against those potentially harmful invaders [5]. As professional antigen-presenting cells DCs reside in the periphery in an immature state, where they take up pathogens or allergens. Upon maturation, the cells undergo a series of phenotypic changes: while their capability to phagocytose antigen decreases, intracellular protein processing and presentation, as well as the expression of co-stimulatory markers are enhanced. The DCs acquire a migratory phenotype, serving their mission to transport the sampled antigen to the secondary lymphoid tissues. The trafficking of immature DCs to sites of inflammation and of mature DCs to the T cell area of secondary lymphoid organs is regulated by the expression of different chemokines and chemokine receptors [6].
In the defense against intracellular microbes or tumors, the key cytokine secreted by DCs is IL-12 [7], which skews T cell responses in the direction of TH1 [8]. IL-12 is induced by pathogen associated molecular patterns such as LPS or by T-cell derived signals such as IL-4 or CD40 ligation [9]. However, simultaneous presence of endogenous signals such as IL-10, TGF-β, corticosteroids, vitamin D3, or PGE2 can convert DC from TH1- to TH2-skewing antigen presenting cells [10]. Recent studies demonstrate that also exogenous factors such as lipids produced by parasites can modulate DC function for the purposes of evading host immunity [11].
Besides their well established role in host defence, DCs are also involved in hypersensitivity reactions against harmless environmental antigens, the allergens [12]. Indeed, evidence emerges that DCs are not only key players in allergic sensitization [13,14] but possibly even contribute to maintaining and shaping the immune response to allergens in already sensitized individuals [15,16]. Understanding the role of DCs in allergic sensitization has been hampered, however, by the fact that to date only very few signals have been identified that actively lead to a TH2 promoting DC phenotype [17,18].
We recently demonstrated that pollen, under physiologial exposure conditions, release not only allergens but also bioactive lipids. Among these are monohydroxylated derivatives of linoleic and linolenic acid [19] that resemble human Leucotriens and activate human neutrophils and eosinophils in vitro. We then extended these data on the impact of pollen associated lipid mediators on dendritic cell function. In brief, dinor isoprostanes (phytoprostanes) released from pollen grains under physiological conditions are able to inhibit the DC's production of IL-12 p70, and DC stimulated with aqueous pollen extracts or E1-phytoprostanes become TH2 skewing in mixed lymphocyte reaction. Additionally, DCs matured in the presence of aqueous pollen extracts respond by releasing TH2 attracting chemokines and aquire a distinct migratory phenotype. Finally, we could show that in a murine sensitization model, nasal instillation of OVA together with aqueous pollen extracts lead to a TH2 shift in draining lymph node T cells. Taken together, multiple lines of evidence imply that by modulating functions of the innate and adaptive immune system, PALMs add to creating a TH2 favoring, pro-allergic micromilieu.
Pollen release lipid mediators - the PALMs
It is commonly accepted that in susceptible individuals, allergic sensitization results after allergens have been taken up by antigen-presenting cells residing in the barrier-forming epithelia like skin or airway mucosa. When investigating this allergic sensitization phase, most studies use purified allergen or allergen-extracts. Under physiological exposure conditions, however, pollen-derived allergens are not released alone, but rather in conjunction with pollen granules, starch grains and other, non-protein substances. One major constituent of pollen excine and exsudate are lipids which are essential in the plant fertilization process as they help the pollen tube to penetrate the stigma [20]. This prompted us to investigate the impact of the whole pollen grain on the human immune system. We recently demonstrated that upon hydration, pollen grains very rapidly release significant amounts of lipids- the so-called pollen-associated lipid mediators (PALMs) - that show structural and functional homology to eicosanoids [21]. Since arachidonic acid metabolites are well known to affect human innate and adaptive immune responses we were prompted to further investigate the effects of aqueous pollen extracts and their constituents.
PALMs potently attract and activate PMN and eosinophils
The finding that pollen grains interact with cells of the human immune system was made by Siegel and Sherman as early as the seventies [22]. We were able to extend these observations by investigating the outcome of granulocyte - pollen interactions. Our data show that pollen grains (birch and grass) attract and activate neutrophils [23] and eosinophils [24] leading to the release of myeloperoxidase and eosinophilic cationic protein, respectively. Chemotactic activity seemed to be independent of protein allergen and could be demonstrated in aqueous pollen extracts (APE) as well as in total lipid extracts (Hexane-isopropanol extracts, HIP) and reverse phase extracts of HIP, enriched for mono-hydroxylated products of linoleic acid. Chemotaxis of Eosinophils was blocked by the LTB4 receptor antagonist LY293111, whilst APE-induced calcium influx in PMN was inhibited by pre-treatment with LTB4 and vice versa in cross-sensitization experiments. Interestingly, these effects seemed to be independent of the sensitization status of the donor and thus might occur in allergic and non-allergic individuals, further arguing for allergen-independent effects. Taken together, these findings indicate that, alongsinde the adaptive immune system, innate mechanisms may also contribute to the recognition of allergens within the respiratory tract.
PALMs confer a TH2 promoting phenotype on DCs
Apart from their effects on neutrophils and eosinophils we investigated the impact of PALMs on human dendritic cells - the initiators of T cell responses. As model, we focused on human monocyte-derived dendritic cells (moDCs). Interestingly, exposure of moDCs with LPS-depleted aqueous birch pollen extracts (Bet.-APE) resulted in a selective upregulation of HLA-DR surface expression, while other maturation markers such as CD80, CD86, CD40 and CD83 were not modulated. On LPS-matured moDCs, Bet.-APE synergized with LPS in the up-regulation of all maturation markers tested. At a functional level, Bet.-APE stimulation of moDC resulted in an enhanced allostimmulatory activity as demonstrated by enhanced proliferative responses of naive allogeneic CD4+ T cells. Importantly, Bet.-APE treatment of moDCs induced a dose dependent inhibition of the LPS or CD40L induced IL-12 p70 production, while IL-6, IL-10 and TNF-α production were not impaired. Thus, water soluble factors released from pollen grains are capable to selectively modulate various DC functions, including the inhibition of the key TH1 cytokine IL-12 p70 [25].
By means of gas chromatography-mass spectometry analysis of Bet.-APE, we demonstrate the presence of E1-, F1-, A1/B1-phytoprostanes in aqueous pollen extracts (see table 1) and show that E1-phytoprostanes - similar to Bet.-APE - dose-dependently inhibit the IL-12 p70 production while not affecting IL-6 production. Like in the case of Bet.-APE, pre-treatment of moDC with E1-phytoprostanes results in an increased IL-4/IFN-γ ratio in CD4+ T cells after allogenic mixed lymphocyte reaction. Thus, PPE1 could be identified as one of the substances contained in Bet.-APE which mediate the TH2 polarizing capacity of moDCs [25].
Table 1.
Concentrations of phytoprostanes in aqueous birch pollen extracts (modified from [25]).
| Concentration in Bet.-APE (10 mg/mL) (nM) | Concentration (μg/g pollen) | |
| PPE1 | 543.6 +/- 41.1 | 17.72 +/- 1.34 |
| PPF1 | 68.6 +/- 1.5 | 2.25 +/- 0.05 |
| PPA1/B1 | 23.8.6 +/- 3.5 | 0.74 +/- 0.11 |
Aqueous pollen extracts modulate chemokine/chemokine receptor expression and migratory capacity of DCs
Maturation of DCs results in substantial changes in the surface expression of T cell costimulatory molecules like HLA-DR, CD40, CD86 and CD80. Concomitantly, maturing DCs undergo distinct changes in the expression of chemokine receptors, licensing them to migrate towards chemokine gradients [26]. In a more recent study we therefore examined the effects of aqueous birch pollen extracts (Bet.-APE) on chemokine production, chemokine receptor expression and migratory capacity of moDCs [27]. Here we found that on immature DCs, Bet.-APE induced expression and function of CXCR4, which might be critical for directing DCs to lymphoid organs during allergic inflammation. Concomitantly, Bet.-APE reduced surface expression of CCR1 and CCR5, reflecting DC maturation and acquisition of a "pro-inflammatory" phenotype [26]. In addition, maturation of DCs with LPS in the presence of Bet.-APE impaired the LPS-induced production of the TH1 attracting chemokines CXCL10 and CCL5. Instead, the cells show an enhanced release of the "TH2" chemokine CCL22. The release of CCL17, a chemokine enhanced in atopic ekzema, was not significantly changed as compared to LPS treatment alone. At a functional level, Bet.-APE increased the capacity of LPS-matured DCs to migrate towards CXCL12 - as reflected by the enhanced expression of CXCR4 - and towards the lymph node homing chemokines CCL19 and CCL21. These effects of Bet.-APE depended on adenylyl cyclase and cAMP induction and strongly mimicked some key characteristics of PGE2 [28,29]. Finally, culture supernatants of DCs matured in the presence of LPS and Bet.-APE attracted TH2 cells in transwell chamber migration assays, while the capacity to recruit TH1 cells was reduced. This might imply that pollen-exposed DCs favor the maintainance of already established TH2 immune responses. Importantly, all effects summarized above were observed in DCs derived from monocytes of non-atopic donors. In our view this supports the concept of allergen-independent adjuvant effects of pollen shifting the primary immune response towards TH2 in susceptible individuals (see table 2).
Table 2.
Summary of effects of PALMs on cells of the innate and adoptive immune system
| PMN | Eosinophils | Dendritic cells |
| Chemotaxis* | Chemotaxis# | IL-12¶↓ |
| Calcium influx* | TH2 bias in MLR¶ | |
| CD11b*↑ | CD11b#↑ | cAMPi§↑ |
| Release of MPO*↑ | Release of ECP↑# | CXCR4§↑ |
| CCR5, CCR1§↓ | ||
| CCL22§↑ | ||
| CXCL10, CCL5↓§ |
Arrows indicate an increase (↑) or decrease (↓).
References: *: Traidl-Hoffmann C et al., J Allergy Clin Immunol (2002); #: Plötz SG et al., J Allergy Clin Immunol (2004); ¶: Traidl-Hoffmann C et al., J Exp Med (2005); §: Mariani V et al., J Immunol (2007)
cAMPi = intracellular cyclic 5'-adenosine monophosphate; ECP = eosinophil cationic protein; MLR = mixed lymphocyte reaction.
Factors from pollen lead to a preferential induction of TH2 responses in vivo
Only recently we were able to undermine our in vitro data by studies in a murine sensitization model [30]. OVA-specific CD4+ T cells were adoptively transferred into BALB/c mice. Twenty-four hours later, mice were challenged by means of intranasal application of OVA in the absence or presence of Bet.-APE or phytoprostanes -E1 or -F1. Polarization of T-cell responses in vivo was analyzed in draining lymph node T cells. While intranasal instillation of phytoprostanes down-regulated both TH1 and TH2 cytokines, inhalation of Bet.-APE lead to a selective down-regulation of IFN-γ and an up-regulation of the TH2 cytokines IL-4, IL-5 and IL-13. This implies that water-soluble factors released from pollen might confer a TH2 polarizing capacity independently from phytoprostanes. The identification of those water-soluble substance(s) and dissecting their respective contributions to allergic sensitization or exacerbation should add to our general understanding of the mechanisms of pollen-induced allergy and might ultimately lead to the development of new therapeutic strategies.
In summary, pollen release regulatory mediators which might add to the generation of an overall TH2 promoting micro milieu. First, pollen provide signals for DCs to mature and acquire a migratory phenotype, preferentially priming type 2 T helper cell responses. The latter effect is partly mediated by E1-phytoprostanes, but other substances are likely to play a role. Secondly, PALMs might help to maintain an established TH2 response by preferential recruitment of TH2 cells and other inflammatory cells (neutrophils, eosinophils) to the site of pollen exposure (Figure 1) (see also table 2).
Figure 1.
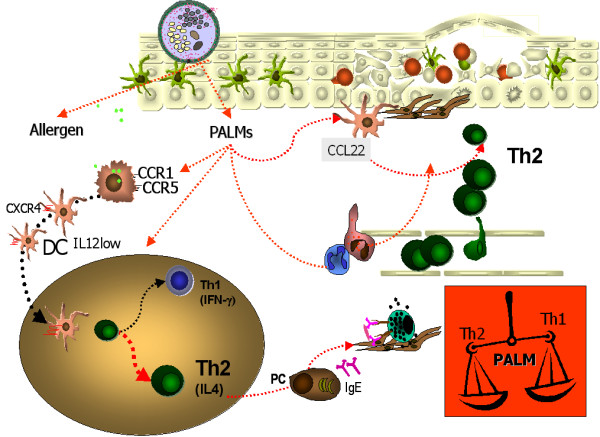
Hypothetical model of a TH2 dominated adoptive immune response and local TH2 promoting micromilieu induced by pollen-associated lipid mediators. When pollen grains are hydrated on the respiratory epithelia, they release allergens and eicosanoid lipids, the so-called pollen-associated lipid mediators (PALMs). Leucotrien-like PALMs have the potential to attract and activate innate cells like neutrophils and eosinophils, while prostaglandin-like PALMs, the phytoprostanes, and possibly other pollen-derived factors, can modulate the migratory and T helper cell polarizing capacities of resident dendritic cells. In addition, DCs exposed to PALMs might be induced to secrete chemokines which preferentially recruit further TH2 cells to the site of pollen exposure. Taken together, the possible effects of PALMs on both cells of the innate and the adoptive immune system might lead to a local microenvironment favoring TH2 responses. I FN-γ = interferon-γ; IL = interleukin; PC = plasma cell
Abbreviations
APE: aqueous pollen extract(s); Bet.-APE: aqueous birch pollen extracts; DC: dendritic cell; MoDC: monocyte-derived dendritic cell; OVA: ovalbumin; PALM: pollen-associated lipid mediator
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
All authors contributed equally to the manuscript. All authors have read and approved the final manuscript.
Acknowledgments
Acknowledgements
The study was supported by a Bundesministerium für Bildung und Forschung (BMBF) grant to T.J. and C.T.-H, V.M. was supported by a research fellowship from the Bayerische Forschungsstiftung, C.T.-H. was a recipient of the Bayerische Habilitationsförderpreis.
Contributor Information
Stefanie Gilles, Email: Stefanie.Gilles@lrz.tum.de.
Valentina Mariani, Email: vamentina@yahoo.it.
Martina Bryce, Email: martinabryce@yahoo.de.
Martin J Mueller, Email: Martin.Mueller@biozentrum.uni-wuerzburg.de.
Johannes Ring, Email: Johannes.Ring@lrz.tum.de.
Heidrun Behrendt, Email: Heidrun.Behrendt@lrz.tum.de.
Thilo Jakob, Email: thilo.jakob@uniklinik-freiburg.de.
Claudia Traidl-Hoffmann, Email: Claudia.Traidl-Hoffmann@rz.tum.de.
References
- Chapman MD, Wunschmann S, Pomes A. Proteases as Th2 adjuvants. Curr Allergy Asthma Rep. 2007;7:363–367. doi: 10.1007/s11882-007-0055-6. [DOI] [PubMed] [Google Scholar]
- Comoy EE, Pestel J, Duez C, Stewart GA, Vendeville C, Fournier C, Finkelman F, Capron A, Thyphronitis G. The house dust mite allergen, Dermatophagoides pteronyssinus, promotes type 2 responses by modulating the balance between IL-4 and IFN-gamma. J Immunol. 1998;160:2456–2462. [PubMed] [Google Scholar]
- Traidl-Hoffmann C, Jakob T, Behrendt H. Determinants of allergenicity. J Allergy Clin Immunol. 2008;123:558–566. doi: 10.1016/j.jaci.2008.12.003. [DOI] [PubMed] [Google Scholar]
- Boldogh I, Bacsi A, Choudhury BK, Dharajiya N, Alam R, Hazra TK, Mitra S, Goldblum RM, Sur S. ROS generated by pollen NADPH oxidase provide a signal that augments antigen-induced allergic airway inflammation. J Clin Invest. 2005;115:2169–2179. doi: 10.1172/JCI24422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, Pulendran B, Palucka K. Immunobiology of dendritic cells. Annu Rev Immunol. 2000;18:767–811. doi: 10.1146/annurev.immunol.18.1.767. [DOI] [PubMed] [Google Scholar]
- Dieu MC, Vanbervliet B, Vicari A, Bridon JM, Oldham E, Ait-Yahia S, Briere F, Zlotnik A, Lebecque S, Caux C. Selective recruitment of immature and mature dendritic cells by distinct chemokines expressed in different anatomic sites. J Exp Med. 1998;188:373–386. doi: 10.1084/jem.188.2.373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trinchieri G, Scott P. Interleukin-12: a proinflammatory cytokine with immunoregulatory functions. Res Immunol. 1995;146:423–431. doi: 10.1016/0923-2494(96)83011-2. [DOI] [PubMed] [Google Scholar]
- Macatonia SE, Hosken NA, Litton M, Vieira P, Hsieh CS, Culpepper JA, Wysocka M, Trinchieri G, Murphy KM, O'Garra A. Dendritic cells produce IL-12 and direct the development of Th1 cells from naive CD4+ T cells. J Immunol. 1995;154:5071–5079. [PubMed] [Google Scholar]
- Cella M, Scheidegger D, Palmer-Lehmann K, Lane P, Lanzavecchia A, Alber G. Ligation of CD40 on dendritic cells triggers production of high levels of interleukin-12 and enhances T cell stimulatory capacity: T-T help via APC activation. J Exp Med. 1996;184:747–752. doi: 10.1084/jem.184.2.747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalinski P, Hilkens CM, Wierenga EA, Kapsenberg ML. T-cell priming by type-1 and type-2 polarized dendritic cells: the concept of a third signal. Immunol Today. 1999;20:561–567. doi: 10.1016/S0167-5699(99)01547-9. [DOI] [PubMed] [Google Scholar]
- Angeli V, Faveeuw C, Roye O, Fontaine J, Teissier E, Capron A, Wolowczuk I, Capron M, Trottein F. Role of the parasite-derived prostaglandin D2 in the inhibition of epidermal Langerhans cell migration during schistosomiasis infection. J Exp Med. 2001;193:1135–1147. doi: 10.1084/jem.193.10.1135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jakob T, Traidl-Hoffmann C, Behrendt H. Dendritic cells--the link between innate and adaptive immunity in allergy. Curr Allergy Asthma Rep. 2002;2:93–95. doi: 10.1007/s11882-002-0001-6. [DOI] [PubMed] [Google Scholar]
- Eisenbarth SC, Piggott DA, Bottomly K. The master regulators of allergic inflammation: dendritic cells in Th2 sensitization. Curr Opin Immunol. 2003;15:620–626. doi: 10.1016/j.coi.2003.09.003. [DOI] [PubMed] [Google Scholar]
- Lambrecht BN. Dendritic cells and the regulation of the allergic immune response. Allergy. 2005;60:271–282. doi: 10.1111/j.1398-9995.2005.00708.x. [DOI] [PubMed] [Google Scholar]
- Novak N, Bieber T. The role of dendritic cell subtypes in the pathophysiology of atopic dermatitis. J Am Acad Dermatol. 2005;53(2 Suppl 2):S171–176. doi: 10.1016/j.jaad.2005.04.060. [DOI] [PubMed] [Google Scholar]
- Novak N, Valenta R, Bohle B, Laffer S, Haberstok J, Kraft S, Bieber T. FcepsilonRI engagement of Langerhans cell-like dendritic cells and inflammatory dendritic epidermal cell-like dendritic cells induces chemotactic signals and different T-cell phenotypes in vitro. J Allergy Clin Immunol. 2004;113:949–957. doi: 10.1016/j.jaci.2004.02.005. [DOI] [PubMed] [Google Scholar]
- Kapsenberg ML, Hilkens CM, Wierenga EA, Kalinski P. The paradigm of type 1 and type 2 antigen-presenting cells. Implications for atopic allergy. Clin Exp Allergy. 1999;2:33–36. [PubMed] [Google Scholar]
- Moore ML, Peebles RS., Jr Update on the role of prostaglandins in allergic lung inflammation: separating friends from foes, harder than you might think. J Allergy Clin Immunol. 2006;117:1036–1039. doi: 10.1016/j.jaci.2005.12.1314. [DOI] [PubMed] [Google Scholar]
- Thoma I, Krischke M, Loeffler C, Mueller MJ. The isoprostanoid pathway in plants. Chem Phys Lipids. 2004;128:135–148. doi: 10.1016/j.chemphyslip.2003.10.002. [DOI] [PubMed] [Google Scholar]
- Wolters-Arts M, Lush WM, Mariani C. Lipids are required for directional pollen-tube growth. Nature. 1998;392:818–821. doi: 10.1038/33929. [DOI] [PubMed] [Google Scholar]
- Behrendt H, Kasche A, Ebner von Eschenbach C, Risse U, Huss-Marp J, Ring J. Secretion of proinflammatory eicosanoid-like substances precedes allergen release from pollen grains in the initiation of allergic sensitization. Int Arch Allergy Immunol. 2001;124:121–125. doi: 10.1159/000053688. [DOI] [PubMed] [Google Scholar]
- Siegel I, Sherman WB. Pollen-white cell interactions. J Allergy. 1970;45:133–145. doi: 10.1016/0021-8707(70)90123-1. [DOI] [PubMed] [Google Scholar]
- Traidl-Hoffmann C, Kasche A, Jakob T, Huger M, Plotz S, Feussner I, Ring J, Behrendt H. Lipid mediators from pollen act as chemoattractants and activators of polymorphonuclear granulocytes. J Allergy Clin Immunol. 2002;109:831–838. doi: 10.1067/mai.2002.124655. [DOI] [PubMed] [Google Scholar]
- Plotz SG, Traidl-Hoffmann C, Feussner I, Kasche A, Feser A, Ring J, Jakob T, Behrendt H. Chemotaxis and activation of human peripheral blood eosinophils induced by pollen-associated lipid mediators. J Allergy Clin Immunol. 2004;113:1152–1160. doi: 10.1016/j.jaci.2004.03.011. [DOI] [PubMed] [Google Scholar]
- Traidl-Hoffmann C, Mariani V, Hochrein H, Karg K, Wagner H, Ring J, Mueller MJ, Jakob T, Behrendt H. Pollen-associated phytoprostanes inhibit dendritic cell interleukin-12 production and augment T helper type 2 cell polarization. J Exp Med. 2005;201:627–636. doi: 10.1084/jem.20041065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sallusto F, Schaerli P, Loetscher P, Schaniel C, Lenig D, Mackay CR, Qin S, Lanzavecchia A. Rapid and coordinated switch in chemokine receptor expression during dendritic cell maturation. Eur J Immunol. 1998;28:2760–2769. doi: 10.1002/(SICI)1521-4141(199809)28:09<2760::AID-IMMU2760>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- Mariani V, Gilles S, Jakob T, Thiel M, Mueller MJ, Ring J, Behrendt H, Traidl-Hoffmann C. Immunomodulatory mediators from pollen enhance the migratory capacity of dendritic cells and license them for Th2 attraction. J Immunol. 2007;178:7623–7631. doi: 10.4049/jimmunol.178.12.7623. [DOI] [PubMed] [Google Scholar]
- Legler DF, Krause P, Scandella E, Singer E, Groettrup M. Prostaglandin E2 is generally required for human dendritic cell migration and exerts its effect via EP2 and EP4 receptors. J Immunol. 2006;176:966–973. doi: 10.4049/jimmunol.176.2.966. [DOI] [PubMed] [Google Scholar]
- Scandella E, Men Y, Legler DF, Gillessen S, Prikler L, Ludewig B, Groettrup M. CCL19/CCL21-triggered signal transduction and migration of dendritic cells requires prostaglandin E2. Blood. 2004;103:1595–1601. doi: 10.1182/blood-2003-05-1643. [DOI] [PubMed] [Google Scholar]
- Gutermuth J, Bewersdorff M, Traidl-Hoffmann C, Ring J, Mueller MJ, Behrendt H, Jakob T. Immunomodulatory effects of aqueous birch pollen extracts and phytoprostanes on primary immune responses in vivo. J Allergy Clin Immunol. 2007;120:293–299. doi: 10.1016/j.jaci.2007.03.017. [DOI] [PubMed] [Google Scholar]