

Abstract
We present a model of cultural evolution in which an individual's propensity to engage in social learning is affected by social learning itself. We assume that individuals observe cultural traits displayed by others and decide whether to copy them based on their overall preference for the displayed traits. Preferences, too, can be transmitted between individuals. Our results show that such cultural dynamics tends to produce conservative individuals, i.e., individuals who are reluctant to copy new traits. Openness to new information, however, can be maintained when individuals need significant time to acquire the cultural traits that make them effective cultural models. We show that a gradual enculturation of young individuals by many models and a larger cultural repertoire to be acquired are favorable circumstances for the long-term maintenance of openness in individuals and groups. Our results agree with data about lifetime personality change, showing that openness to new information decreases with age. Our results show that cultural remodeling of cultural transmission is a powerful force in cultural evolution, i.e., that cultural evolution can change its own dynamics.
Keywords: mathematical model, personality, social learning
Because cultural transmission is a crucial element of cultural evolution, researchers typically assume that individuals in cultural species have an inherent tendency to seek social information, according to fixed rules that are determined genetically (1–4). Examples of such rules are to adopt a trait if it is used by the majority of individuals or by successful individuals (3, 5–7). It is likely, however, that the rules of cultural transmission can be modified by social learning. For example, individuals might learn from others whether or not to rely on social information. Cultural modifications of cultural transmission have no analogue in genetic evolution and, thus, are difficult to study with models from evolutionary biology [a major influence on cultural evolutionary theory (1, 8)]. Genes, in fact, are acquired only once in an individual's lifetime, following “fair” rules that typically do not change allele and genotype frequencies (9, 10). In contrast, cultural traits are acquired (and lost) continuously throughout life, according to rules that may depend on the traits themselves (11).
Ghirlanda et al. (12) studied a model in which social learning was allowed to influence an individual's propensity to seek social information. The result was that individuals rapidly learned to be very conservative, i.e., to disregard social information. The reason is that cultural traits that make individuals more conservative are less likely to be relinquished in favor of other traits, for the very reason that they inhibit social learning. Thus, with repeated social interactions, such traits tend to accumulate at the expense of traits that promote social learning. This result shows that cultural evolution can exert a powerful influence on its own dynamics and on characteristics of individuals that are typically studied within personality psychology. Psychologists, in fact, consider openness to new experiences a fundamental personality trait, although often assumed to be genetically determined (13, 14). Empirical studies show that people become more conservative with age (13, 15) and are therefore consistent qualitatively with Ghirlanda et al.'s predictions. The latter, however, appear too extreme: People do not come to flatly reject all new information as their model suggests. Here we investigate whether purely cultural dynamics can maintain openness to new information despite an inherent tendency to favor conservatism. We explore the idea that openness can be maintained because individuals need to acquire culture to be effective cultural models, i.e., to transmit their traits to others. Thus, if an individual becomes very conservative early in life, she may not be able to learn traits that make her a good cultural model. Therefore her traits, among which those that favor conservatism, will not be transmitted to others.
Models
Model 1.
We begin our analysis with a simple model showing why conservatism is favored in cultural evolution. Individuals are characterized by two cultural traits. The first determines whether the individual is conservative (C) or open (O). Conservative individuals never learn from others, whereas open individuals are always willing to learn. The second trait determines whether the individual is either effective (E) or ineffective (I) at transmitting her traits. Thus, cultural transmission is possible only when an open individual observes an effective model. When this happens, social learning succeeds with probability padopt, in which case the observer adopts one trait of the model, chosen at random. We call padopt the “efficiency of cultural transmission.” Fig. 1 shows the possible transitions between cultural types. Assuming that individuals are born open and that effectiveness must be learned, Fig. 1 also represents possible life histories. A young individual may learn directly to be conservative (lower branch) by copying a CE model (CI individuals cannot transmit their traits). Alternatively (upper branch), the individual may first become effective by copying an effective model (either CE or OE), then become conservative by copying a CE individual. In this model, only half of individuals learn to be effective cultural models (CE or OE), and only a fraction 1/τpadopt of these are open (OE), where τ is the expected number of social interactions in a lifetime [see supporting information (SI) Appendix]. If, for example, τ = 100 and padopt = 0.5, at cultural evolutionary equilibrium we have 46% of CE individuals and only 4% of OE individuals. The reason why openness cannot be maintained is that individuals in the OE state quickly transit into the CE state, where they stay until they die (for example, permanence in the CE state is ≈12 times longer than in the OE state if τ = 100 and padopt = 0.5) (see SI Appendix). Hence, OE individuals have fewer chances to enculturate young, compared with CE individuals.
Fig. 1.

Possible transitions between cultural types in a simple model of cultural transmission. O, individual open to social learning; C, conservative individual; E, individual effective as cultural model; I, ineffective individual. Assuming that individuals are born in the OI state, the figure also represents possible developmental pathways, with CI and CE as possible end states.
Model 2.
We consider now a model in which openness and effectiveness depend on many cultural traits, rather than just one. We expect individuals that stay open for longer to become better cultural models, because they can acquire more effective traits. Effective individuals would thus transmit not only traits that favor effectiveness but also traits that favor openness.
Each individual is described by a number of trait pairs, each pair consisting of a feature and a preference for that feature. Features may describe, for instance, aspects of clothing, opinions, ways of playing music, and so on. Our core assumption is that the probability that an observer copies a model depends on the observer's preferences for the features possessed by the model. Thus, features determine an individual's effectiveness as cultural model, and preferences determine how conservative the individual is. Cultural transmission is most likely when the model displays many features for which the observer has a strong preference and unlikely when few or no preferred features are displayed. Being a successful model requires having learned many cultural features that are preferred by others. This can be described assuming that the probability Pom that an observer copies a model is
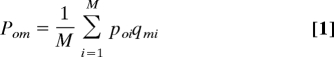 |
where poi is the ith preference of the observer, qmi is the ith feature of the model, and M is the number of feature–preference pairs. We assume in the following discussion that both feature and preference values range between 0 and 1. We have also simulated a model with binary features (qoi ∈ {0,1}), obtaining the same qualitative results (note also that setting M = 1 and binary p and q yields back Model 1).
Eq. 1 is formally identical to simple neural network models of behavior (16), as well as to models of binary choice in economic decision theory (17). Both kinds of models are widely used to describe decisions in which internal factors (here, the observer's preferences) determine how external information (the model's features) is used to reach a decision. In other words, we are assuming that decision-making about whether to copy others works in a similar way as decision-making in other domains. This mechanism allows individuals to copy traits for which they do not have a strong preference, given that a model displays enough features that are preferred. How a pop star dresses, for example, may not be particularly appealing per se, but it is often imitated because the pop star displays other preferred features, such as musical ability, success, and so on. This phenomenon is often observed in studies of human social learning (18, 19) and is related to the finding that people tend to evaluate others as a whole, without analytically distinguishing between different features (20).
Specifically, we assume that, when an observer copies a model, both features and preferences can be copied (we assume that preferences can be made explicit by, e.g., voicing or otherwise expressing appreciation or dislike for particular features). The probability that a trait is successfully adopted by an observer is padopt. Similarly to Model 1, this parameter describes the efficiency of cultural transmission. When an observer copies a model, in fact, an average of Mpadopt preferences and Madopt features are transmitted. Thus, padopt sets an upper limit to the transfer of culture in a single interaction. Such transfer can be substantial if padopt is high, but if padopt is low culture is transferred gradually in many interactions, even if the observer is open and the model is effective.
We simulate a population of N = 100 individuals with overlapping generations for T = 104 discrete time steps. At each time step, all individuals observe a randomly chosen model, and cultural transmission occurs according to the rules above. Feature and preference values also change by innovation, i.e., there is a probability pinnovate = 0.1 per time step that each feature and preference is altered by a random amount in the range [−0.1,0.1]. Lastly, individuals die with a probability pdeath = 0.01 per time step, resulting in an average lifetime of τ = 100 time steps (100 interactions with others). Individuals who die are replaced by “newborns,” i.e., individuals who are culturally naïve and minimally conservative: qi = 0 and pi = 1 for all i = 1,…, M.
We study how individuals' openness to social learning depends on the number, M, of features and preferences, and on the efficiency of cultural transmission, padopt. We define the openness of individual o as the probability that she copies a randomly chosen model, Po = [1/(N − 1)]Σm≠o1,NPom, and the openness of the population, P̄, as the average individual openness. Table 1 provides a symbol legend.
Table 1.
Symbol legend for Model 2
| Symbol | Value | Definition |
|---|---|---|
| N | 100 | Population size |
| M | 1–50 | No. of cultural trait pairs |
| (qij, pij) par | [0,1] × [0,1] | Feature-preference pair j of individual i |
| padopt: | [0,1] | Probability that a trait is successfully adopted when an observer copies a model |
| pinnovate | 0.1 | Probability of adding a uniform random deviate in [−0.1,0.1] to each trait |
| pdeath | 0.01 | Probability of death per time step |
| [0,1] | Probability that individual o copies m | |
| [0,1] | Individual openness (probability that individual o copies a randomly chosen model) | |
| [0,1] | Individual effectiveness (probability that individual m is copied by a randomly chosen model) | |
| [0,1] | Population openness (probability that a randomly chosen observer copies a randomly chosen model) |
Results
Cultural Evolution of Openness and Conservatism.
Fig. 2 shows the time course of population openness for three values of the efficiency of transmission, padopt, and for a population in which individuals have 10 feature–preference pairs. When the efficiency of transmission is either low (padopt = 0.05) or high (padopt = 0.5), the population evolves quickly toward conservatism. For intermediate transmission efficiency, however, population openness is maintained. Fig. 3 summarizes results from more simulations, where we varied the number of feature–preference pairs as well as transmission efficiency. Fig. 3 Left shows steady-state population openness. Fig. 3 Right shows the steady-state probability that a trait (feature or preference) is actually copied in an interaction. When individuals have only one feature–preference pair (blue lines), openness can be maintained only when the efficiency of cultural transmission is very low, resulting in very little cultural transmission. As the number of traits increases, however, it becomes possible to maintain substantial openness for a wider range of values of transmission efficiency, and cultural transmission occurs relatively often. To understand these results we need to consider how individuals acquire and transmit their traits as a function of cultural repertoire size and efficiency of cultural transmission, focusing on what combinations of traits are favored under different circumstances.
Fig. 2.

Time course of population openness in a population with M = 10 feature–preference pairs and for three different values of the efficiency of transmission, padopt. Each line is the average of 25 simulations. Standard deviations are of the order of 10−4 for padopt = 0.05, 10−1 for padopt = 0.25, and 10−2for padopt = 0.5.
Fig. 3.

Steady-state population openness and probability of cultural transmission. (Left) Population openness vs. efficiency of transmission in populations with different numbers of feature–preferences pairs. Blue line, M = 1; green line, M = 5; red line, M = 10; cyan line, M = 20; magenta line, M = 50. Each line is the average of 100 simulations. Standard deviations vary between 10−4 and 10−1 and are higher when population openness is higher. (Right) Average probability that each trait (feature or preference) is actually copied in an interaction (P̄ padopt) vs. efficiency of transmission. The same conditions as for Left apply.
Note first that successful cultural models, i.e., individuals who transmit their traits many times, should possess two characteristics. First, they should have high feature values to promote being copied by others. Second, they should have low preference values so that their traits are stable in time, meaning that other individuals can observe them repeatedly (11, 21). These requirements, however, are in conflict, because to acquire high feature values individuals must be open to copying others. In particular, newborn individuals will not be copied by others unless they first acquire a sufficient number of non-zero feature values. Thus, a successful model is an individual who first learns how to be an efficient model (by acquiring high feature values) and then becomes conservative (by acquiring low preference values).
When only one trait pair is culturally transmitted, naïve individuals can acquire a high feature value in just a few interactions. Further interactions will tend to lower the individual's preference for the feature, because it is more likely that a less conservative model copies a more conservative one than vice versa (12). Thus, newborns quickly become conservative. In larger cultures, more learning attempts are needed to acquire enough traits to be effective cultural models. On one hand, when padopt is very small (e.g., in a species inept at social learning), newborns have little chance of acquiring traits at all; thus, individuals remain essentially naïve throughout their lives. In such circumstances, individuals are potentially willing to copy, but there is little to copy (leftmost part of the curves in Fig. 3). On the other hand, if padopt is very high, a newborn who copies an older individual acquires a large proportion of the latter's traits, i.e., both preferences and features at the same time. Thus, newborns can acquire in few interactions both high feature values and low preference values from older individuals, resulting in a population of conservative individuals in which cultural transmission seldom occurs.
For intermediate transmission efficiency, newborns need many interactions to become effective models. Individuals that remain open have more occasions to acquire new features and thus to become effective models. This process boosts population openness, because the most effective models will have (and therefore, will transmit) not only high features values but also relatively high preferences values. When M = 50, for instance, openness can be maintained even when cultural transmission is extremely efficient. This is a condition in which the cultural repertoire is so large that, to become effective models, individuals must remain open for a significant fraction of their lifetime.
We are not aware of data that bear directly on these predictions, which are summarized in Table 2. We may note, however, that both theoretical models and archaeological evidence show that large cultural repertoires can be maintained only by large groups (22–25). Combining this result with ours, we expect smaller societies to be more conservative, which agrees with the observation that traditional societies (whose very name implies conservatism) tend to be small.*
Table 2.
Culturally evolved characteristics of individuals in Model 2, as a function of size of cultural repertoire and efficiency of cultural transmission
| Efficiency of cultural transmission, padopt | Size of cultural repertoire, M |
||
|---|---|---|---|
| Small | Medium | Large | |
| Low | Open, ineffective | Open, ineffective | Open, ineffective |
| Medium | Conservative, effective | Open, effective | Open, effective |
| High | Conservative, effective | Conservative, effective | Open, effective |
Boldface type indicates cases for which efficient cultural transmission between individuals can occur.
Life History of Conservatism and Effectiveness.
In our model, older individuals are typically more conservative than younger ones (Fig. 4Left). When population openness is high, however, old individuals can be almost as open as young ones. This result agrees with long-term studies of personality change, showing that older individuals typically score lower on the Openness dimension of “Five Factor” personality inventories (refs. 15 and 26–28; see also ref. 29 for a similar finding in chimpanzees). This measure indicates the extent to which individuals are open to new experiences and correlates with such behavior as seeking out information (30, 31), adopting Western traits among Hong Kong youth (32), and ascribing to political liberalism (33).
Fig. 4.

Change of individual openness and effectiveness with age. (Left) Average change of individual openness with age. (Right) Average change of individual effectiveness with age. Each line is the average of 25 simulations with M = 10; other values of M yield similar results. Standard deviations are typically of order 10−2.
The model also predicts change in personality to be slower in old age (Fig. 4) because, as an individual becomes more conservative, she becomes less likely to adopt traits that may affect her personality further. Personality changes, indeed, appear to be larger between age 30 and 50 than between age 50 and 70 (28, 34). A similar pattern is observed for vocabulary size (a proxy of acquired information, to be compared with feature acquisition in Fig. 4 Right), which seems to increase until age 50–55, after which it changes little (35). Importantly, these findings suggest that decrease in openness with age is not a simple by-product of impaired information processing in old age. Indicators of brain function, such as working memory capacity, speed of pattern matching, and reasoning speed, in fact, decrease with age at an approximately constant rate (35) rather than slowing down after age 50.
Another prediction, which demonstrates how openness and effectiveness can interact in cultural transmission, is that open individuals accumulate a larger cultural repertoire (Fig. 5; this is not the case when the population is conservative, because the only open individuals are young who have not acquired any traits). This agrees with evidence that, among individuals of similar age, more open individuals (sensu Five Factor model) have higher levels of education (36, 37). Lastly, the model suggests that, when openness is maintained, older individuals should be more effective cultural models because they have had more opportunities to acquire effective features (Fig. 4 Right). Studies of social learning in children and adolescents often find that older models are more effective (38, 39), and ethnographic data suggest that older individuals are often more effective cultural models (reviewed in ref. 6).
Fig. 5.

Correlation between individual openness and number of learned cultural features vs. efficiency of cultural transmission. The correlation is assessed at three moments during the life span (see key below graphs). Average life span is 100. Each line is the average of 25 simulations with M = 10 cultural traits; other values of M yield similar results.
Discussion
Despite simple assumptions on individual interactions (Eq. 1), our model exhibits a very rich dynamics. In particular, the model shows that cultural evolution can maintain openness to new information as well as effectiveness at cultural transmission. Ghirlanda et al. (12) only observed extreme conservatism, despite also using Eq. 1, because they either considered a single feature–preference pair (M = 1) and efficient transmission (padopt = 1) or assumed that all individual had the same, fixed effectiveness from birth.
The model also suggests a causal mechanism for an observed systematic change in personality. Personality psychologists are divided regarding the relative importance of genetic vs. experiential determinants of personality change (13, 14), partly because it is difficult to disentangle genetic and experiential influences on such complex traits (28, 40). Additionally, most arguments about personality change make only weak predictions about the direction and pattern of change. For instance, the fact that adults need to behave differently from youth [e.g., because of their role as parents (15)] does not directly explain why they are less open. Likewise, the argument that older individuals should invest less in learning because they have less time to amortize the costs of learning does not necessarily predict that change in openness should slow down with age.† Thus, understanding personality change requires not only more empirical work but also more specific theoretical predictions.
Our model neglects many potential influences on the openness of individuals and on their effectiveness as cultural models: genetic influences on personality (41, 42), including potential biases in cultural transmission (3, 6); perceived or actual costs and benefits of different cultural traits (1, 3, 43, 44); and nonrandom social networks (45). Costs and benefits of traits, for instance, can influence the decision of whether to adopt traits both implicitly (e.g., through differential reinforcement of behavior) and explicitly (through rational cost–benefit analysis), and may have such far-reaching consequences as the restructuring of social networks (45). We do not mean to imply that these factors are unimportant, and we have neglected them only to isolate the potential effects of culturally evolving cultural transmission. As recalled earlier, such effects have been largely neglected in theoretical studies. Our results show that cultural remodeling of cultural transmission rules can be a potent force in cultural evolution.
Supplementary Material
Acknowledgments.
We thank Pontus Strimling, Jonas Sjöstrand, Cecilia M. Lipira, and members of the Laland Laboratory at the University of St. Andrews for helpful comments. This work was supported by European Commission Contract FP6-NEST-2004-043434 (CULTAPTATION).
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
This article contains supporting information online at www.pnas.org/cgi/content/full/0908889106/DCSupplemental.
Observation from Kevin Laland.
An anonymous reviewer suggested this point.
References
- 1.Cavalli-Sforza L, Feldman M. Cultural Transmission and Evolution. Princeton: Princeton Univ Press; 1981. [PubMed] [Google Scholar]
- 2.Lumsden CJ, Wilson EO. Genes, Mind, and Culture: The Coevolutionary Process. Cambridge, MA: Harvard Univ Press; 1981. [Google Scholar]
- 3.Boyd R, Richerson PJ. Culture and the Evolutionary Process. Chicago: Univ of Chicago Press; 1985. [Google Scholar]
- 4.Takahasi K. Evolution of transmission bias in cultural inheritance. J Theor Biol. 1998;190:147–159. doi: 10.1006/jtbi.1997.0541. [DOI] [PubMed] [Google Scholar]
- 5.Henrich J, Boyd R. The evolution of conformist transmission and the emergence of between-group differences. Evol Human Behav. 1998;19:215–241. [Google Scholar]
- 6.Henrich J, Gil-White F. The evolution of prestige: Freely conferred deference as a mechanism for enhancing the benefits of cultural transmission. Evol Human Behav. 2001;22:65–96. doi: 10.1016/s1090-5138(00)00071-4. [DOI] [PubMed] [Google Scholar]
- 7.Eriksson K, Enquist M, Ghirlanda S. Critical points in current theory of conformist social learning. J Evol Psychol. 2007;5:67–88. [Google Scholar]
- 8.Mesoudi A, Whiten A, Laland KN. Towards a unified science of cultural evolution. Behav Brain Sci. 2006;29:329–383. doi: 10.1017/S0140525X06009083. [DOI] [PubMed] [Google Scholar]
- 9.Crow JF, Kimura M. An Introduction to Population Genetics Theory. London: Harper & Row; 1970. [Google Scholar]
- 10.Futuyma DJ. Evolutionary biology. Sunderland, MA: Sinauer; 1998. [Google Scholar]
- 11.Strimling P, Eriksson K, Enquist M. Repeated learning makes cultural evolution unique. Proc Natl Acad Sci USA. 2009;106:13870–13874. doi: 10.1073/pnas.0903180106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ghirlanda S, Enquist M, Nakamaru M. Cultural evolution develops its own rules. The rise of conservatism and persuasion. Curr Anthrop. 2006;47:1027–1034. [Google Scholar]
- 13.Helson R, Kwan VSY, John OP, Jones C. The growing evidence for personality change in adulthood: Findings from research with personality inventories. J Res Person. 2002;36:287–306. [Google Scholar]
- 14.Costa Paul TJ, McCrae RR. Age changes in personality and their origins: Comment on roberts, walton, and viechtbauer (2006) Psychol Bull. 2006;132:26–28. doi: 10.1037/0033-2909.132.1.26. [DOI] [PubMed] [Google Scholar]
- 15.McCrae RR, et al. Age differences in personality across the adult life span: Parallels in five cultures. Dev Psychol. 1999;35:466–477. doi: 10.1037//0012-1649.35.2.466. [DOI] [PubMed] [Google Scholar]
- 16.Enquist M, Ghirlanda S. Neural Networks and Animal Behavior. Princeton: Princeton Univ Press; 2005. [Google Scholar]
- 17.Train K. Discrete Choice Models with Simulation. Cambridge, MA: Cambridge Univ Press; 2003. [Google Scholar]
- 18.Bandura A. Social Learning Theory. Englewood Cliffs, NJ: Prentice–Hall; 1977. [Google Scholar]
- 19.Dalton MA, et al. Effect of viewing smoking in movies on adolescent smoking initiation: a cohort study. Lancet. 2003;362:281–285. doi: 10.1016/S0140-6736(03)13970-0. [DOI] [PubMed] [Google Scholar]
- 20.Cooper WH. Ubiquitous halo. Psycholog Bull. 1981;90:212–244. [Google Scholar]
- 21.Tanaka MM, Kendal JL, Laland KN. From traditional medicine to witchcraft: Why medical treatments are not always efficacious. PLOS One. 2009;4:e5192. doi: 10.1371/journal.pone.0005192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Henrich J. Demography and cultural evolution: How adaptive cultural processes can produce maladaptive losses—The Tasmanian case. Am Antiq. 2004;69:197–214. [Google Scholar]
- 23.Enquist M, Ghirlanda S. Evolution of social learning does not explain the origin of human cumulative culture. J Theor Biol. 2007;246:129–135. doi: 10.1016/j.jtbi.2006.12.022. [DOI] [PubMed] [Google Scholar]
- 24.Strimling P, Sjöstrand J, Enquist M, Eriksson K. Accumulation of independent cultural traits. Theor Popul Biol. 2009;78:77–83. doi: 10.1016/j.tpb.2009.04.006. [DOI] [PubMed] [Google Scholar]
- 25.Shennan SJ. Demography and cultural innovation: A model and its implications for the emergence of modern human culture. Camb Archaeol J. 2001;11:5–16. [Google Scholar]
- 26.Labouvie-Vief G, Diehl M, Tarnowski A, Shen J. Age differences in adult personality: Findings from the United States and China. J Gerontol Ser B. 2000;55:4–17. doi: 10.1093/geronb/55.1.p4. [DOI] [PubMed] [Google Scholar]
- 27.Srivastava S, Oliver PJ, Gosling SD, Potter J. Development of personality in early and middle adulthood: Set like plaster or persistent change? J Person Soc Psychol. 2003;84:1041–1053. doi: 10.1037/0022-3514.84.5.1041. [DOI] [PubMed] [Google Scholar]
- 28.Caspi A, Roberts BR, Shiner RL. Personality development: Stability and change. Ann Rev Psychol. 2005;56:453–484. doi: 10.1146/annurev.psych.55.090902.141913. [DOI] [PubMed] [Google Scholar]
- 29.King JE, Weiss A, Farmer K. A chimpanzee (Pan troglodytes) analogue of cross-national generalization of personality structure: Zoological parks and an African sanctuary. J Person. 2005;73:389–410. doi: 10.1111/j.1467-6494.2005.00313.x. [DOI] [PubMed] [Google Scholar]
- 30.Heinström J. Five personality dimensions and their influence on information behaviour. Inf Res. 2003;9:165. [Google Scholar]
- 31.Duff A, Boyle E, Dunleavy K, Ferguson J. The relationship between personality, approach to learning and academic performance. Person Individual Differ. 2004;36:1907–1920. [Google Scholar]
- 32.Yik MSM, Bond MH. Exploring the dimensions of Chinese person perception with indigeneous and imported constructs: Creating a culturally balanced scale. Int J Psychol. 1993;28:75–95. [Google Scholar]
- 33.McCrae RR. Social consequences of experiential openness. Psychol Bull. 1996;120:323–337. doi: 10.1037/0033-2909.120.3.323. [DOI] [PubMed] [Google Scholar]
- 34.Roberts BW, Del Vecchio WF. The rank-order consistency of personality traits from childhood to old age: A quantitative review of longitudinal studies. Psychol Bull. 2000;126:3–25. doi: 10.1037/0033-2909.126.1.3. [DOI] [PubMed] [Google Scholar]
- 35.Salthouse TA. What and when of cognitive aging. Curr Directions Psychol Sci. 2004;13:140–144. [Google Scholar]
- 36.Costa PTJ, McCrae RR. In: Comprehensive handbook of personality and psychopathology. Hersen M, Thomas JC, editors. Vol. 1. Hoboken, NJ: Wiley; 2006. [Google Scholar]
- 37.Graziano WG, Jensen Campbell LA, Finch JF. The self as mediator between personality and adjustment. J Person Soc Psychol. 1997;73:392–404. doi: 10.1037//0022-3514.73.2.392. [DOI] [PubMed] [Google Scholar]
- 38.Brody GH, Stoneman Z. Selective imitation of same-age, older, and younger peer models 52. Child Dev. 1981;52:717–720. [Google Scholar]
- 39.Brody GH, Stoneman Z. Peer imitation: An examination of status and competence hypotheses. J Genet Psychol. 1985;146:161–170. [Google Scholar]
- 40.Allik J, McCrae RR. Toward a geography of personality traits: Patterns of profiles across 36 cultures. J Cross-Cult Psychol. 2004;35:13–28. [Google Scholar]
- 41.Plomin R, Defries JC, McClearn GE, McGuffin P. Behavioral Genetics. 4 Ed. New York: Worth; 2000. [Google Scholar]
- 42.Laland KN. Animal cultures. Curr Biol. 2008;18:R366–R370. doi: 10.1016/j.cub.2008.02.049. [DOI] [PubMed] [Google Scholar]
- 43.Bisin A, Verdier T. The economics of cultural transmission and the evolution of preferences. J Econ Theor. 2001;97:298–319. [Google Scholar]
- 44.Fernandez R. Culture as learning: The evolution of female labor force participation over a century. NBER Work Pap. 2007:w13373. [Google Scholar]
- 45.Skyrms B, Pemantle R. In: The Place of Probability in Science. Eells E, Fetzer J, editors. Chicago: Open Court; 2004. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.