

Abstract
We summarize a series of bidirectional research studies that demonstrate the persistence-strengthening effects of differential reinforcement of alternative behavior on problem behavior. We model a possible solution to this problem with rats followed by replication with a human clinical population. The importance of coordinated basic and applied research to stimulate new behavioral technologies is emphasized.
Keywords: translational research, behavioral momentum, differential reinforcement of alternative behavior, resistance to change
The past three decades have seen growing recognition of the importance of linking basic and applied research to develop more effective behavioral technologies and establish the broad generality of basic behavioral relations (e.g., Dietz, 1978; Hake, 1982; Lerman, 2003; Mace, 1994; Mace & Wacker, 1994). This coordinated effort has come to be known as translational research. The most common approach is to translate basic research findings into behavioral technologies that are often novel and better suited to complex human behavior in natural settings. However, translational research can also be bidirectional when human behavioral problems stimulate new basic research aimed at elucidating fundamental behavioral relations that have clear implications for technology development (e.g., Mace, Mauro, Boyajian, & Eckert, 1997).
We will illustrate this approach to bidirectional translational research with a summary of three studies related to behavioral momentum. The first is a clinical study that demonstrates that although differential reinforcement of alternative behavior (DRA) can reduce the frequency of problem behavior, it can also inadvertently increase the persistence of problem behavior. The second study is a rat model of one possible solution to the persistence-strengthening effects of DRA that could be translated into a novel clinical intervention for problem behavior. Finally, the procedures employed in the rat model are translated into a clinical intervention, and its effectiveness is evaluated.
Implications of Behavioral Momentum Theory for Clinical Research and Practice
Nevin and his colleagues have described two independent aspects of the discriminated operant: (a) baseline response rate and (b) the resistance of that response rate to change when disrupted by various operations such as extinction, satiation, punishment, alternative reinforcement, and distraction (Nevin, 1992; Nevin & Grace, 2000; Nevin, Tota, Torquato, & Shull, 1990). These two aspects of the discriminated operant are a function of two different types of contingencies. It is well known that baseline response rate is a function of the contingency between the response and the reinforcer that maintains it (i.e., response–reinforcer contingencies). However, the resistance to change of the baseline response rate is dependent on the total reinforcement correlated with the context in which the discriminated operant has occurred and been reinforced (i.e., context–reinforcer contingencies). This distinction suggests the possibility that contingencies capable of reducing response rate, such as DRA, can have the opposite effect on the persistence of the discriminated operant.
This situation is illustrated in Experiment 2 of Nevin et al. (1990) with pigeon subjects. Figure 1 diagrams the baseline three-component multiple concurrent schedule of reinforcement used in this experiment. In all components, the left key represents a concurrently available response and the right key denotes the target response. Condition A is a model of DRA in which a relatively high rate of reinforcement is arranged for alternative behavior (45 reinforcers per hour) and substantially less reinforcement is available for the target behavior (15 per hour). Although the target behavior in Condition A is reinforced at a relatively low rate, the total reinforcement correlated with the condition, or green-key context, is relatively high (60 per hour). Conditions B and C represent reinforcement of the target behavior only at rates of 15 per hour and 60 per hour, respectively. Following stable baseline performance, responding was alternately disrupted with extinction and satiation. Resistance of the target behavior (right key) to extinction and satiation was greatest and comparable in Conditions A and C that had 60 reinforcers correlated with their key-color contexts. This pigeon model suggests that clinical applications of DRA may inadvertently increase the persistence of problem behavior, even though the intervention reduces its frequency. This becomes clinically relevant when lapses in treatment integrity degrade the DRA intervention and the possibility of a resurgence of problem behavior exists.
Figure 1.
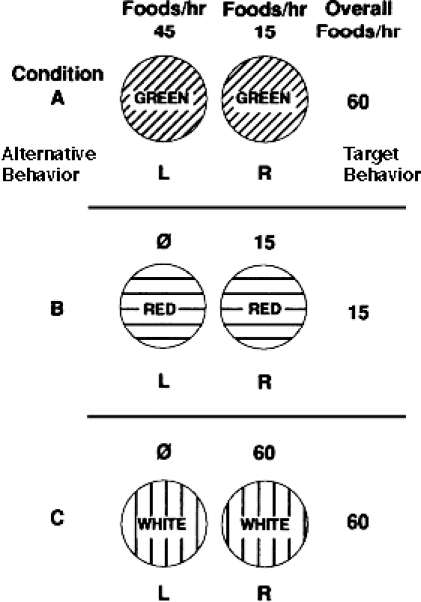
Diagram of the three-component multiple concurrent schedule of reinforcement from Experiment 2 of Nevin et al. (1990). Condition A (green-key condition) models DRA in which reinforcement on the alternative left key is three times greater than reinforcement of the target right key. Adapted from Nevin et al. (1990) and reprinted by permission.
Clinical Demonstration of the Persistence-Strengthening Effects of DRA
This study tested the hypothesis that clinically relevant problem behavior would be more resistant to extinction following high-rate DRA reinforcement than comparatively low-rate baseline reinforcement. Three children with mental retardation and problem behaviors (hair pulling, food stealing, and aggression) participated in the study. Prestudy functional analyses identified the reinforcer that maintained the target behaviors for all 3 participants, and these were arranged in ratio or interval schedules during the baseline condition (BL). In the DRA condition, the reinforcer that maintained the problem behavior was delivered contingent on either a communication response (functional communication training) or appropriate eating. DRA reinforcement rates were between 165% to 195% of baseline rates of reinforcement across participants, and baseline contingencies for target behaviors remained in effect in order to closely replicate Condition A of Nevin et al. (1990). The extinction condition (EXT) consisted of the therapist blocking the target behavior with his or her hand or arm while providing no other responses to the participants. These three conditions were arranged in one of two sequences across participants: (a) BL-DRA-EXT-BL-EXT or (b) BL-EXT-BL-DRA-EXT.
Figure 2 summarizes the results of this study. For all 3 participants, occurrences of problem behavior, expressed as the proportion of the preceding baseline response rates, were 76.9% to 462.2% higher following baseline reinforcement than DRA reinforcement. This clinical finding is entirely consistent with and predicted by Nevin et al.'s (1990) Experiment 2 with pigeons, and provides preliminary support for consideration of the persistence-strengthening effects of DRA in clinical applications.
Figure 2.

Mean proportion of baseline response rate of disruptive behavior during DRA, extinction following baseline reinforcement (BL → EXT), and extinction following DRA reinforcement (DRA → EXT) for the 3 participants.
Rat Model of a Possible Solution to the Persistence-Strengthening Effects of DRA
Resurgence of problem behavior following lapses in treatment integrity or discontinuation of treatment is commonly reported (Osnes & Lieblein, 2003). Given the basic research and the clinical study reported above, it is possible that resurgence of problem behavior is at least partly due to the added reinforcement that comes with the introduction of DRA to the context in which problem behavior occurs and has been reinforced. We will now summarize a study that modeled a possible solution to this problem. We chose to model a solution with nonhuman subjects because placing a prosocial alternative behavior on extinction with humans requires, in our view, a basis for predicting that the solution developed would be effective with humans in a clinical context.
Four experimentally naive rats were presented with the baseline three-component multiple concurrent schedule diagrammed in the top panel of Figure 3. Each component arranged food pellets on variable-interval (VI) schedules and was correlated with a different light flicker rate located above the response levers; extinction in baseline was correlated with a dark light above the lever. In this model, the right lever represents the target problem behavior, and the left lever denotes a prosocial alternative to problem behavior. Component 1 with a slow flicker rate (1 flick per second) arranged 24 pellets per hour for the target behavior, with no reinforcement for alternative behavior on the left lever. This component modeled reinforcement of problem behavior alone. Component 2, with a fast flicker rate (5 flicks per second), modeled high-rate DRA for prosocial behavior on the left lever (96 per hour) in the same context in which problem behavior was also reinforced at a rate of 24 per hour, for a total of 120 per hour correlated with the component context. Component 3 represented high-rate reinforcement of alternative behavior alone (96 per hour) in the presence of a constant light above the lever without reinforcing problem behavior on the right lever (dark light above the lever). That is, this component modeled DRA reinforcement in a context in which problem behavior had no history of reinforcement.
Figure 3.
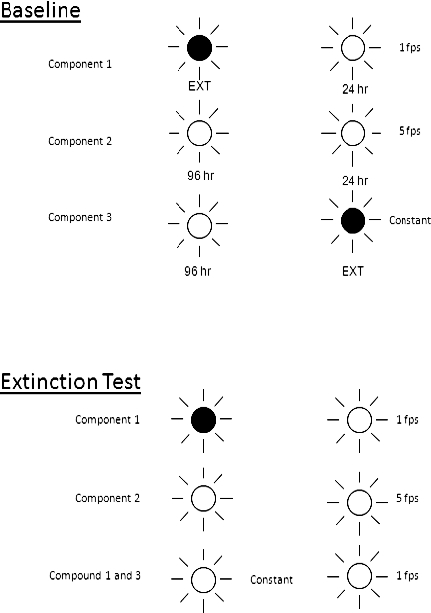
Diagram of the discriminative stimuli and reinforcement rates in the three-component multiple concurrent schedule of the rat model of a possible solution to the persistence-strengthening effects of DRA. The top panel depicts the baseline arrangement, and the bottom panel represents arrangements during the extinction tests.
Extinction tests that followed baseline are diagrammed in the lower panel of Figure 3. The discriminative stimuli in Components 1 and 2 were identical to those in baseline. However, the third component during extinction was comprised of a compound of the discriminative stimuli correlated with DRA in a separate context (constant light) and the one correlated with baseline reinforcement of problem behavior only (1 flick per second). This compound stimulus modeled the clinical situation in which a prosocial alternative behavior (e.g., a communication response) is established in a context that has not been correlated with reinforcement of problem behavior. It represented a possible solution to the persistence-strengthening effects of DRA, namely, to teach alternative behavior in a separate setting to avoid adding reinforcement to the setting correlated with reinforcement of problem behavior. If successful, resistance to extinction during the compound stimulus would be substantially lower than during the stimulus correlated with DRA and reinforcement of problem behavior in the same context (Component 2). Figure 4 summarizes the results of the extinction tests. For all subjects, resistance to extinction was greatest in Component 2, and for 3 subjects (MV-54, MV-55, and MV-56) resistance during the compound stimulus was comparable to or lower than in Component 1, in which DRA had never been introduced. This rat model suggests that training an alternative behavior with DRA in a setting separate from the one in which problem behavior has a history of reinforcement may be a possible solution to the persistence-strengthening effects of DRA.
Figure 4.

Mean proportion of baseline log response rate on the target right lever across blocks of extinction during Components 1 and 2 and compound of Baseline Components 1 and 3 for the 4 subjects.
Clinical Demonstration of the Avoidance of the Persistence-Strengthening Effects of DRA
Our third study is a translation of the rat model into a clinical intervention aimed at avoiding the persistence-strengthening effects of DRA. Two children and 1 adult with developmental disabilities, who displayed a variety of disruptive behaviors, were participants. Prestudy functional analyses found that escape from task demands maintained all 3 participants' disruptive behavior. All participants were presented with the following baseline multiple concurrent schedule in which the left schedule consisted of reinforcement of mands for escape from task demands and the right schedule consisted of reinforcement of disruptive behavior: Component 1 (yellow: EXT, VI 75 s), Component 2 (blue: fixed-interval [FI] 20 s, VI 75 s), and Component 3 (white: FI 20 s, EXT). Therapists presented instructional demands to participants using a three-step prompt hierarchy and wore different-colored hospital gowns in each of the components, as designated above. As with the rat model, Component 1 represented reinforcement of disruptive behavior without reinforcement of mands for escape. Component 2 represented high-rate reinforcement of mands for escape plus comparatively low-rate reinforcement of disruption. Component 3 was conducted in a novel room that the participants had never been in and, hence, had no history of reinforcement for disruptive behavior. This component reinforced mands for escape and did not reinforce disruptive behavior.
The top panel of Figure 5 shows the median disruptions per hour during the three baseline components. Rates of disruptive behavior were greatest in Component 1 and lowest in Component 3 for all participants. The addition of DRA to the context in which disruptive behavior continued to be reinforced resulted in substantial reductions in disruptive behavior for 2 participants (Mickey and Paul) and moderate reductions for Terry.
Figure 5.

Median disruptions per hour during Baseline Components 1, 2, and 3 (top panel) and during extinction tests in Components 1 and 2 and compound of Baseline Components 1 and 3 (bottom panel) for the 3 participants.
Following the baseline phase, disruptive behavior and mands were placed on extinction in order to assess the extent to which conducting mand training in a setting with no history of reinforcement for disruptive behavior would reduce the persistence-strengthening effects of DRA. Extinction consisted of continuation of instruction at a constant pace that was not altered by disruptive behavior. During extinction, the discriminative stimuli (therapists, gown colors, and room) in Components 1 and 2 were identical to baseline. In a third component, a stimulus compound was created that combined the baseline discriminative stimuli from Component 1 (yellow; reinforcement of disruptive behavior only) and Component 3 (white; reinforcement of mands in a novel setting). This stimulus compound comprised the same total rate of reinforcement present in Component 2. However, because the DRA discriminative stimuli in Baseline Component 3 were distinct from those in Baseline Component 1, we anticipated that, as in the rat model, introducing a therapist wearing a white gown into the setting in which the therapist wore the yellow gown would not increase the resistance to extinction of disruptive behavior in that setting.
The lower panel of Figure 5 presents the median disruptions per hour during the three components under extinction. For all 3 participants, resistance to extinction was greatest in Component 2 and substantially less in Component 1 and the compound of Baseline Components 1 and 3. These data support the proposition that the persistence-strengthening effects of DRA can be significantly lessened or avoided by teaching a prosocial alternative behavior in a setting with no history of reinforcement for problem behavior before introducing the discriminative stimuli correlated with prosocial behavior into the context correlated with reinforcement of problem behavior.
Conclusion
DRA has been a mainstay intervention for applied behavior analysis since the inception of the field. That this important approach to treatment could have the unintended side effect of strengthening the persistence of the very problem behavior it successfully reduces could not have been anticipated without basic research on the nature of resistance to change. One of Nevin et al.'s (1990) experiments modeled a typical DRA intervention and showed that DRA could strengthen the persistence of problem behavior. This research inspired a series of bidirectional studies aimed at demonstrating the problem in clinical populations and finding a possible solution to it. In our view, this type of research would not be possible without the close connection between basic and applied research with bidirectional influences on each sector.
Acknowledgments
This article is a summary of a symposium presentation at the 30th annual convention of the Association of Behavior Analysis International in Phoenix, Arizona (May, 2009), which has been submitted for publication in the Journal of the Experimental Analysis of Behavior. We thank Tim Hackenberg for inviting this presentation and chairing the symposium.
REFERENCES
- Dietz S.M. Current status of applied behavior analysis: Science versus technology. American Psychologist. 1978;33:805–814. [Google Scholar]
- Hake D.F. The basic-applied continuum and the possible evolution of human operant social and verbal research. The Behavior Analyst. 1982;5:21–28. doi: 10.1007/BF03393137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerman D.C. From the laboratory to community application: Translational research in behavior analysis. Journal of Applied Behavior Analysis. 2003;36:415–419. doi: 10.1901/jaba.2003.36-415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mace F.C. Basic research needed for stimulating the development of behavioral technologies. Journal of the Experimental Analysis of Behavior. 1994;61:529–550. doi: 10.1901/jeab.1994.61-529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mace F.C, Lalli J.S, Shea M.C, Lalli E.P, West B.J, Roberts M, et al. The momentum of human behavior in a natural setting. Journal of the Experimental Analysis of Behavior. 1990;54:163–172. doi: 10.1901/jeab.1990.54-163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mace F.C, Mauro B.C, Boyajian A.E, Eckert T.L. Effects of reinforcer quality on behavioral momentum: Coordinated applied and basic research. Journal of Applied Behavior Analysis. 1997;30:1–20. doi: 10.1901/jaba.1997.30-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mace F.C, Wacker D.P. Toward greater integration of basic and applied behavioral research: An introduction. Journal of Applied Behavior Analysis. 1994;27:569–574. doi: 10.1901/jaba.1994.27-569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nevin J.A. An integrative model for the study of behavioral momentum. Journal of the Experimental Analysis of Behavior. 1992;57:301–16. doi: 10.1901/jeab.1992.57-301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nevin J.A, Grace R.C. Behavioral momentum and the law of effect. Behavioral and Brain Sciences. 2000;23:73–130. doi: 10.1017/s0140525x00002405. [DOI] [PubMed] [Google Scholar]
- Nevin J.A, Tota M.E, Torquato R.D, Shull R.L. Alternative reinforcement increases resistance to change: Pavlovian or operant contingencies? Journal of the Experimental Analysis of Behavior. 1990;53:359–379. doi: 10.1901/jeab.1990.53-359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osnes P.G, Lieblein T. An explicit technology of generalization. The Behavior Analyst Today. 2003;3:364–374. [Google Scholar]