

Abstract
In order to investigate the replication timing properties of PCDH11X and PCDH11Y, a pair of protocadherin genes located in the hominid-specific non-pseudoautosomal homologous region Xq21.3/Yp11.2, we conducted a FISH-based comparative study in different human and non-human primate (Gorilla gorilla) cell types. The replication profiles of three genes from different regions of chromosome X (ZFX, XIST and ATRX) were used as terms of reference. Particular emphasis was given to the evaluation of allelic replication asynchrony in relation to the inactivation status of each gene. The human cell types analysed include neuronal cells and ICF syndrome cells, considered to be a model system for the study of X inactivation. PCDH11 appeared to be generally characterized by replication asynchrony in both male and female cells, and no significant differences were observed between human and gorilla, in which this gene lacks X-Y homologous status. However, in differentiated human neuroblastoma and cerebral cortical cells PCDH11X replication profile showed a significant shift towards allelic synchrony. Our data are relevant to the complex relationship between X-inactivation, as a chromosome-wide phenomenon, and asynchrony of replication and expression status of single genes on chromosome X.
Key words: PCDH11X/Y, replication asynchrony, replication timing, X-inactivation, Xq21.3/Yp11.2 homology block
Introduction
The non-pseudoautosomal X-Y homologous region (Xq21.3/Yp11.2) is the largest region (∼4 Mb) shared by the human sex chromosomes. It was generated approximately 6 million years ago, close to the divergence of hominid and chimpanzee lineages, by a duplicative transposition from the X to the Y chromosome, followed by a paracentric inversion of the whole block and deletion of the DXS214 subinterval on the Y chromosome (Schwartz et al. 1998, Ross et al. 2005, Williams et al. 2006). So far, only three genes have been described in this region: TGIFLX/Y, a testis-specific transcription factor from the homeobox gene family (Blanco-Arias et al. 2002); PABPC5, a poly-A binding protein, whose homologue has been lost from the Y chromosome during human evolution (Blanco et al. 2001); and PCDH11X/Y, a gene pair coding for protocadherin X and protocadherin Y, two cell-surface adhesion molecules expressed predominantly in brain (Yoshida & Sugano 1999, Blanco et al. 2000).
In female mammals most genes on one X chromosome are silenced as a result of X-chromosome inactivation, a silencing mechanism which evolved to restore equal gene expression between males and females (Lyon 1961). However, pseudoautosomal genes and X-linked genes with Y homology or functional Y orthologues tend to escape X-inactivation and are usually expressed from both the active and inactive X chromosome (reviewed in (Disteche et al. 2002, Brown & Greally 2003).
Accordingly, it is reasonable to envisage that PCDH11X would escape from inactivation. The proposed role for protocadherin X and protocadherin Y in language development, handedness, and brain asymmetry (Crow 2002), combined with the lack of X-Y homologous status in non-human primates (Wilson et al. 2006), suggest the possibility of a sexlinked differential expression and gene dosage differences between human and primates. Indication of a possible escape of PCDH11X from X-inactivation was initially provided by results obtained with a human X chromosome-specific cDNA microarray showing elevated expression of PCDH11X in cells with multiple X chromosomes (Sudbrak et al. 2001). Recently Lopes and collaborators (2006) demonstrated that the CpG islands of PCDH11X/Y were not methylated in human and chimpanzee, and that there was a two-fold higher expression of PCDH11X in brain tissue in human females than in males. However, whether PCDH11X escapes from inactivation remains to be conclusively proved.
X-inactivation is associated with a series of epigenetic modifications (Reviewed in Chow et al. 2005). Besides undergoing changes in DNA methylation and histone modifications, the inactive X chromosome condenses into the Barr body and becomes late replicating in S phase. As a result, X chromosomes in female cells replicate asynchronously, with genes on the inactive X replicating later than their active counterparts.
Replication banding and fluorescence in situ hybridization (FISH) in its different guises have been repeatedly and successfully used as reliable cytological assays for the study of replication properties of chromosomes and genes (Dutrillaux et al. 1976, Vogel et al. 1989, Drouin et al. 1990, Haaf 1997, Takebayashi et al. 2005). In particular, the ‘ISHbased Fdot counting’ method (Selig et al. 1992), used in this investigation, allows one to visualize on cell nuclei the unreplicated DNA segments as single hybridization signals (‘singlets’) and the replicated loci as double signals (‘doublets’). In a random cell population of an unsynchronized culture, genes which replicate early in the cell cycle will show a high percentage of doublets, while late-replicating genes will show mostly singlets.
The validity of this approach was initially tested by analysing genes from the cystic fibrosis (CF) region on human chromosome 7, the replication profiling of which had been previously determined in expressing and non-expressing cells by molecular methods. By ‘dot-counting’ on unsynchronized cell cultures the authors confirmed the feasibility of the method to infer replication timing properties of genes (Selig et al. 1992). In the same study the conclusion was reached that the replication timing of autosomal genes is normally highly synchronous, with 90% of cell nuclei showing an equivalent hybridization pattern on both homologues (singlet-singlet/SS or doublet-doublet/DD) and only 10% of nuclei showing an asynchronous pattern (singlet-doublet/SD).
However, subsequent studies on the replication timing patterns of murine chromosomal regions containing four imprinted genes (Igf2, Igf2r, H19 and Snrpn) suggested that asynchronous replication — inferred from the high percentage of SD signals in a random population of unsynchronized cells — might be a characteristic of imprinted genetic regions (Kitsberg et al. 1993), consistent with earlier replication banding studies on the chromosomal region 15q11.2 (Izumikawa et al. 1991).
A ‘dot-counting’ comparison of the replication timing of genes on the human chromosome X that are known to escape inactivation (pseudoautosomal) with those that are inactivated (Boggs & Chinault 1994), as well as confirming the correlation between expression and relative time of replication of individual genes, established that genes subject to inactivation replicate asynchronously while genes that escape inactivation replicate synchronously, thus validating replication asynchrony as an indicator of an allele-specific mode of expression.
As part of a coordinated research effort aimed at evaluating the genetic and epigenetic status of PCDH11X/Y in human and non-human primates, we conducted a study on the replication timing of this gene pair carrying out a FISH-based comparative investigation in different human and gorilla (Gorilla gorilla) cell types, using as terms of reference three genes from chromosome X (ZFX, XIST and ATRX), previously assessed for their replication timing by means of similar and/or alternative analytical methods. In particular, in the attempt to infer information on PCDH11X possible escape from inactivation in human cells, our analysis focused on comparing the allelic replication synchrony profile of this gene to those of the other X-linked genes under investigation, the inactivation status of which is known.
Materials and methods
Cell culture
Chromosome preparations were obtained from: (1) human male and female peripheral blood lymphocytes; (2) EBV-transformed human lymphoblastoid cell lines from male (COX, ECACC No. 85102902) and female (DO208915, ECACC No. 95010407); (3) EBV-transformed lymphoblastoid cell line from a female immunodeficiency, centromeric instability, facial anomalies (ICF) syndrome patient (GM08714, Coriell Cell Repositories); (4) EBV-transformed lymphoblastoid cell lines from female gorilla (Gorilla gorilla) (Machi cell line provide by Professor A. Rickinson, University of Birmingham); (5) human female neuroblastoma cell line SH-SY5Y (ECACC No. 94030304) before and after differentiation with retinoic acid; and (6) a cerebral cortical cell line developed from a female patient with Rasmussen encephalitis HCN-2 (ATCC No. CRL-10742). PHA-stimulated peripheral blood lymphocytes and B-lymphoblast cells were cultured in RPMI-1640 supplemented with 10% foetal bovine serum (FBS) and 1% l-glutamine (Sigma). Prior to hypotonic treatment, one of the two cultures of the human female B-Cell line (identified in the text as ‘line 1’), was treated with thymidine (300 µg/ml) for 17 h and then washed and left in complete medium for 7 h, before adding colcemid (0.05 µg/ml). The cells were then treated with hypotonic solution (0.0075 M KCl) for 20 min at 37°C and fixed in three changes of Carnoy's fixative (3:1 methanol:acetic acid). Slides were prepared following standard procedures and stored at −20°C. The karyotype of each cell line (apart from the cortical cell line from which it had not been possible to obtain metaphase preparations due to the extremely slow growth rate) was checked for chromosome X numerical or structural abnormalities.
Neuroblastoma cell line differentiation
SH-SY5Y cells were grown in flasks in DMEM: Ham's F12 (1:1) complete medium. After trypsinization the cells were plated onto adherent substrate-coated (Collagen 1) dishes at low density (35-5×103 cells/cm2) and allowed to attach overnight. The following day trans-retinoic acid (RA) was added to the medium at a final concentration of 10 mM. After 5 days the medium was changed to NeuroBasal (NB) supplemented with B-27 (1×), Glutamax I (2 mM), KCl (20 mM), dibutyril cyclic AMP (2 mM) and rhBDNF (50 ng/ml). The cells were kept in this medium for 5 days before proceeding with the chromosome harvest protocol as above.
Probes
The following BAC probes were used in the in situ hybridization experiments: RP1-290C9 for ZFX (Sanger Institute), RP11-42M11 for ATRX (Sanger Institute), RP13-216E22 for XIST (Sanger Institute), and RP11-187C6 for PCDH11 (provided by Dr Carole Sargent, University of Cambridge). BAC RP11-17E6 from chromosome 2q23.3 (Sanger Institute) was used as an autosomal control.
Fluorescence in-situ Hybridization (FISH)
The hybridization procedure followed a standard protocol. The probes were labelled with either Biotin-16-dUTP (Roche) or Digoxigenin-11-dUTP (Roche) by nick-translation. Following labelling, the probes were ethanol precipitated in a mix of salmon testis DNA (Gibco BRL), Escherichia coli tRNA (Boehringer) and 3 M sodium acetate. They were then dried on a heating block at 60°C with a 50× excess of human Cot-1 DNA and resuspended at 20 ng/µl in hybridization solution (50% formamide, 10% dextran sulphate, 2× SSC). The probes were denatured at 72°C for 5 min and pre-annealed at 37°C for 15 min, before being applied to the denatured slides. Prior to replication timing analysis on interphase nuclei, in order to detect the possible presence of numerical and/ or structural abnormalities involving chromosome X and in particular the genes of interest, each BAC probe was co-hybridized with an FITC-directly labelled chromosome X paint (Cambio) and checked on metaphase chromosomes (in all cell lines, apart from the cortical cell line from which it had not been possible to obtain metaphase preparations due to the extremely slow growth rate). Before proceeding with the hybridization, the slides were denatured in 70% formamide at 70°C for 2 min, quenched in 2× SSC at 4°C and then dehydrated in an ethanol series. Following hybridization, the slides were washed in 50% formamide at 42°C for 10 min and 2× SSC at 42°C for 5 min. The biotinylated probes were detected with Texas red-Conjugated streptavidin (Molecular Probes), followed by a layer of biotinylated anti-streptavidin (Vector Laboratories) and a final layer of Texas redconjugated streptavidin. The digoxigenin probes were detected using mouse anti-digoxigenin antibody (Roche) and goat anti-mouse Alexa-488 (Molecular Probes). The slides were mounted with Vectashield (Vector Laboratories) containing 4′,6-diamidino-2-phenylindole (DAPI) for chromosomecounterstaining.
BrdU pulse-labelling
The cells were incubated at 37°C with 10 µM 5′-Bromo-2′-deoxyuridine (BrdU) (Sigma) for 30 min before cell fixation. Prior to FISH, the BrdU pulse-labelled cells were pretreated by permeabilization in 0.5% Triton-X for 10 min and then transferred to 0.1 M HCl for 10 min at room temperature. The slides were then washed in 2×SSC for 5 min and equilibrated in 50% formamide/2×SSC for at least 15 min before denaturation. After hybridization the BrdU labelling was detected using a mouse anti-BrdU antibody (Roche) followed by a goat antimouse Alexa 488 antibody (Invitrogen).
Microscopy analysis
The slides were examined using an Olympus BX-51 epifluorescence microscope coupled to a Sensys charge-coupled device (CCD) camera (Photometrics). A minimum of 100 nuclei were analysed for each hybridization experiment. Texas red, Alexa-488 and DAPI fluorescence images were taken as separate grey-scale images using specific filter combinations and then pseudocoloured and merged using the software package Genus (Applied Imaging International).
Statistical analysis
RT-PCR
Detection of gene expression was performed using RT-PCR. RNA samples were extracted from four cell lines (female B-cell line DO208915, female ICF B-cell line GM08714, SH-SY5Y female neuroblastoma cell line before and after differentiation with retinoic acid) and a female brain control using QIAGEN RNeasy Mini Kit according to the manufacturer's protocol. Samples were treated with DNAseI prior to reverse transcription according to the manufacturer's the protocol (Invitrogen), with 1 µg of RNA treated with DNAseI. cDNA was prepared using SuperScriptIII (Invitrogen) according to standard procedure. PCR reactions used the following primers:
- Xist, product size 80 bp:
- Forward: 5′-AACCAGGAAAGAGCTAGTAT GAGGAA-3′;
- Reverse: 5′-TTCGCAAGCTGAATTAAGTG-3′.
- ATRX, product size 79 bp:
- Forward: 5′-CATCATCTAGTTGAACTTTGG CATTT-3′;
- Reverse: 5′-CAATAAACGGCCAGAATTTC CA-3′.
- ZFX, product size 80 bp:
- Forward: 5′-CGAATATAGGTCTGCAGAC TC-3′;
- Reverse: 5′-CACACTTGAACGGCATCTCT-3′.
- PCDH11, product size 133 bp:
- Forward: 5′-CCCTTTCGTTTACCTCTTCATTC-3′;
- Reverse: 5′-CACAAGATTACAAAGTTGAGAG CAG-3′.
RT-PCR reactions were performed using RedTaq (Sigma) according to the manufacturer's protocol, using a 2.5 mM final concentration of MgCl2. Cycling parameters were as follows: 94°C-4 min, followed by 35 cycles of 94°C for 30 s, 55°C for 30 s and 72°C for 30 s, followed by a final 5 min at 72°C. Products were resolved on a 2% agarose gel using Hyper-LadderV (Bioline).
Quantitative RT-PCR
Quantitation of PCDH11 expression was performed by real-time RT-PCR using the above primers. The results were normalized to the control gene RPII, product size 267 bp: forward: 5′-GCAC CACGTCCAATGACAT-3′; reverse 5′-GTGCGGC TGCTTCCATAA-3′. Quantitation was performed using SYBR green PCR master mix (Applied Biosystems) according to the manufacturer℉s protocol, on an iCyler real-time PCR machine (BioRad). Analysis was performed using the standard curve method and normalization to expression of the RPII control, using the iCyler software.
Results
We compared the replication timing properties of PCDH11X to its homologue on the Y chromosome (PCDH11Y), and three other genes from chromosome X (ZFX, XIST and ATRX) by conducting a FISH-based ‘dot-counting’ assay on a range of different human cell types and a gorilla (Gorilla gorilla) lymphoblastoid cell line.
The percentage of different replication patterns — single-single (SS), double-double (DD) and single-double (SD) fluorescent signals — for each probe in interphase was determined by randomly scoring at least 100 nuclei for each hybridization experiment (examples in Figure 1). In agreement with the analytical parameters most recently applied in similar assays (Singh et al. 2003, Gribnau et al. 2005, Gimelbrant & Chess 2006), a value of %SD ≥ 30% was considered indicative of replication asynchrony. As single signals in interphase can occur as a result of inefficient hybridization, the efficiency of hybridization was checked in parallel on metaphase chromosomes on the same slide. Experiments in which the hybridization efficiency was considered not satisfactory were omitted from the analysis.
Figure 1.
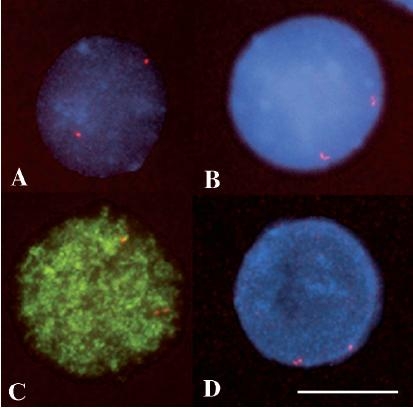
Fluorescence in situ hybridization (FISH) allows visualization of replicated and unreplicated DNA fragments in the cell nuclear context. Examples of interphase nuclei showing in red: (A) a single-single (SS) hybridization pattern (both DNA segments have not replicated yet); (B) a double-double (DD) hybridization pattern (both DNA segments have already replicated); (C) and (D) single-double (SD) hybridization patterns (one fragment has replicated, but its homologue has not replicated yet). In A, B and D the nuclei are counterstained with Dapi (blue). In C hybridization was performed on BrdU pulse-labelled cells. The anti-BrdU antibody (green) identifies nuclei in S phase. (Scale bar = 10 µm.).
The replication profile or frequency of different replication patterns for PCDH11X was initially assessed by hybridizing and analysing chromosome preparations obtained from a human female peripheral blood culture and from two different cultures of a human female B-lymphoblastoid cell line, which hereafter will be referred to as line 1 and line 2. The replication profile of PCDH11X was also analysed in a B-lymphoblastoid cell line obtained from a female ICF patient. As shown in Table 1, replication asynchrony between the two alleles was observed for the PCDH11X locus in all of these cell types, including the ICF cells, with the percentage of hemizygous replication patterns or %SD (one singlet and one doublet in the same nucleus) consistently within a range of 36–39%, and no statistically significant difference observed when comparing the PCDH11X replication profile between any of cell types analysed (chi-squared test of no differences between blood, line 1, line 2 and ICF cell line p < 0.172). A hemizygous pattern of replication — with a %SD value (37%) within the same range observed above — was also observed in gorilla female cells. When the replication profile of PCDH11X was compared between human and gorilla B cells there was no statistically significant difference between gorilla and line 2, and only a marginal difference between gorilla and line 1 (p = 0.03) was observed, with the difference between line 1 and 2 probably due to the slightly different culture conditions.
Table 1.
Summary of the ‘dot-counting’ replication timing assay carried out on five different genes (PCDH11, ATRX, XIST, ZFX and autosomal control probe) in different human and gorilla cell types
| SS | DD | SD | SD% | N | Singlets | Doublets | |
|---|---|---|---|---|---|---|---|
| PCDH | |||||||
| ♀ blood | 48 | 16 | 37 | 36% | 101 | 66% | 34% |
| ♀ line 1 | 58 | 11 | 40 | 37% | 109 | 72% | 28% |
| ♀ line 2 | 40 | 25 | 39 | 37% | 104 | 57% | 45% |
| ♀ line ICF | 45 | 16 | 39 | 39% | 100 | 65% | 35% |
| ♀ Gorilla | 41 | 23 | 38 | 37% | 102 | 59% | 41% |
| ♀ neurobl. | 40 | 26 | 40 | 38% | 106 | 57% | 43% |
| ♀ neurobl.dif. | 93 | 21 | 44 | 28% | 158 | 73% | 27% |
| ♀ cortical | 61 | 11 | 30 | 29% | 102 | 75% | 25% |
| ♂ blood | 66 | 19 | 36 | 30% | 121 | 69% | 31% |
| ♂ line | 51 | 10 | 43 | 41% | 104 | 70% | 30% |
| ATRX | |||||||
| ♀ blood | 50 | 11 | 45 | 42% | 106 | 68% | 32% |
| ♀ line 1 | 62 | 20 | 48 | 37% | 130 | 66% | 34% |
| ♀ line 2 | 52 | 19 | 38 | 35% | 109 | 65% | 35% |
| ♀ line ICF | 51 | 15 | 35 | 35% | 101 | 68% | 32% |
| ♀ Gorilla | 49 | 14 | 39 | 38% | 102 | 67% | 33% |
| ♀ neurobl. | 62 | 12 | 46 | 38% | 120 | 71% | 29% |
| ♀ neurobl.dif. | 48 | 17 | 44 | 40% | 109 | 64% | 36% |
| XIST | |||||||
| ♀ blood | 67 | 8 | 34 | 31% | 109 | 77% | 23% |
| ♀ line 1 | 49 | 13 | 41 | 40% | 103 | 67% | 33% |
| ♀ line 2 | 61 | 14 | 34 | 31% | 109 | 72% | 28% |
| ♀ Gorilla | 53 | 10 | 43 | 40% | 106 | 70% | 30% |
| ♀ neurobl. | 66 | 20 | 39 | 31% | 125 | 68% | 32% |
| ♀ neurobl.dif. | 63 | 15 | 49 | 38% | 127 | 69% | 31% |
| ZFX | |||||||
| ♀ blood | 55 | 19 | 37 | 33% | 111 | 66% | 34% |
| ♀ line 1 | 56 | 21 | 35 | 31% | 112 | 66% | 34% |
| ♀ line 2 | 61 | 13 | 40 | 35% | 114 | 71% | 29% |
| ♀ Gorilla | 48 | 21 | 36 | 34% | 105 | 63% | 37% |
| ♀ neurobl. | 66 | 21 | 34 | 28% | 121 | 69% | 31% |
| ♀ neurobl.dif. | 77 | 13 | 30 | 25% | 120 | 77% | 23% |
| Autosom. | |||||||
| ♀ blood | 84 | 6 | 25 | 22% | 115 | 84% | 16% |
| ♀ neurobl.dif. | 77 | 7 | 33 | 28% | 117 | 80% | 20% |
| ♂ blood | 86 | 2 | 28 | 24% | 116 | 86% | 14% |
| BrdU-Lab. | |||||||
| PCDH | 47 | 8 | 54 | 50% | 109 | n.a. | n.a. |
| ATRX | 40 | 12 | 51 | 49% | 103 | n.a. | n.a. |
| Xist | 41 | 18 | 47 | 44% | 106 | n.a. | n.a. |
| ZFX | 73 | 36 | 100 | 48% | 209 | n.a. | n.a. |
| autosom. | 44 | 19 | 48 | 43% | 111 | n.a. | n.a. |
‘SS’ is the number of single-single hybridization patterns observed in each experiment, ‘DD’ is the number of double-double hybridization patterns observed in each experiment and ‘D’ is the number of single-double hybridization patterns observed in each experiment (also shown as a percentage), while ‘N’ is the total number of nuclei analysed for each experiment. ‘BrdU-labelled’ indicates the observations carried out on BrdU-positive nuclei. Percentages of total number of single signals (‘Singlets’) and double signals (‘Doublets’) are also included.
In order to compare PCDH11X to its homologue on chromosome Y, the ‘dot-counting’ assay with the PCDH11 probe was then carried out on chromosome preparations obtained from a human male peripheral blood culture and a human male B-lymphoblastoid cell line. Findings of SD values equivalent to 30% in male blood and 41% in male B cells confirmed that — similarly to the two alleles on the X chromosomes in female cells — in male cells the X and Y homologue genes replicate asynchronously. Statistical comparison of the gene pair replication profile in male vs female blood or male vs female B cells also showed no difference.
We then compared PCDH11X to ATRX (Xq13.3) and XIST (Xq13.2), two X-linked genes known to undergo inactivation (reviewed in Carrel & Willard 2005). The range of SD pattern frequency in the different human and gorilla cell types was 35–42% for ATRX and 31–40% for XIST, in both cases indicative of allelic asynchrony. There were no significant differences between PCDH11X and ATRX replication profiles in line 1, line 2, blood and gorilla. There was a significant difference between PCDH11X and XIST in blood cells (p < 0.06) and in line 2 (p < 0.02) but not in line 1 (p < 0.68). However, when account is taken of the number of tests performed in this analysis the significance is only marginal. As well as comparable levels of asynchrony, no significant changes in the replication timing profile of PCDH11X and ATRX were also observed in the ICF cells, with the percentages of SS (single-single) and DD (double-double), as well as the total percentages of ‘singlets’ and ‘doublets’, being very similar in ICF and controls for both genes.
Comparisons were then carried out between PCDH11X and ZFX (Xp22.2-p21.3), a gene known to escape inactivation and previously reported by FISH analysis to replicate synchronously (Boggs & Chinault 1994). Although a relatively lower percentage of hemizygous replication patterns was consistently observed for ZFX in each of the cell types analysed, no statistically significant differences were found when the replication profiles for PCDH11X and ZFX were compared in either blood, line 1 or gorilla, and only a marginally significant difference was observed in line 2 (p < 0.02).
As a further control we compared the replication asynchrony of the four X-linked genes with that of a randomly chosen autosomal probe, mapping on chromosome 2. The percentages of hemizygous patterns observed when hybridizing this autosomal probe to chromosome preparations from male (%SD = 24%) and female (%SD = 22%) peripheral blood cells were significantly lower than those observed for all the other genes in male and female blood (p < 0.0001), consistent with a synchronous replication pattern expected for autosomal genes.
Since the percentage of SD can sometimes be overestimated because of doublet artefacts arising from the separation of denatured chromatin strands in G1 and G2, we carried out a further set of experiments in which bromodeoxyuridine (BrdU) was incorporated during cell culture in order to identify and assay cells in S phase exclusively. Observations conducted on the human female B-lymphoblastoid cell line after treatment with BrdU and hybridization showed — as expected — an increase in the percentage of SD signals for PCDH11X (50%) and similarly raised values for the other three X-linked genes used as controls (ZFX = 48%, XIST = 44% and ATRX = 49%), further confirming the finding of an asynchronous replication pattern for all the X-linked genes analysed — irrespective of their inactivation status — as previously shown by our experiments on unsynchronized B cells. Unexpectedly, a substantial increase in replication asynchrony after BrdU treatment was also observed for the autosomal control. We suspect that its distinctive late-replicating nature when compared to the other loci — suggested by the relative percentages of total singlets and doublets (Table 1) — might have slightly biased the analysis in synchronized conditions.
Next we investigated the replication timing properties of PCDH11X in human neuronal cells, carrying out fluorescent in situ hybridization experiments on chromosome preparations obtained from the SH-SY5Y neuroblastoma cell line, before and after differentiation with retinoic acid, and from the HCN-2 cerebral cortical cell line. The latter are cells of neuroepithelial origin and express neuronal markers even in the undifferentiated state. In both cases a statistically significant (p<0.02) reduction in the asynchrony of replication was observed for PCDH11X with a 29% of SD patterns in HCN-2, and 28% of SD patterns in the differentiated SH-SY5Y, with the same neuroblastoma cell line showing a higher percentage of asynchrony (38%) before differentiation. Corresponding values for ATRX and XIST in the neuroblastoma cells both before and after differentiation were asynchronous, while ZFX in both non-differentiated and differentiated neuroblastoma cells presented with a more synchronous pattern. In the differentiated neuroblastoma cells the %SD value for PCDH11X was found to be significantly different from ATRX (p < 0.05), marginally different from XIST (p = 0.05), but not significantly different from ZFX and the autosomal control.
Finally, we tested the hypothesis of no differences in replication profile between all four X-linked genes in females within each cell type for blood, line 1, line 2, differentiated neuroblastoma and gorilla (Figure 2). Only differentiated neuroblastoma cells showed significant differences between the genes (p = 0.01 when the SD values were compared to SS and DD values pooled together and p = 0.02 when the SS and DD values were not pooled together).
Figure 2.
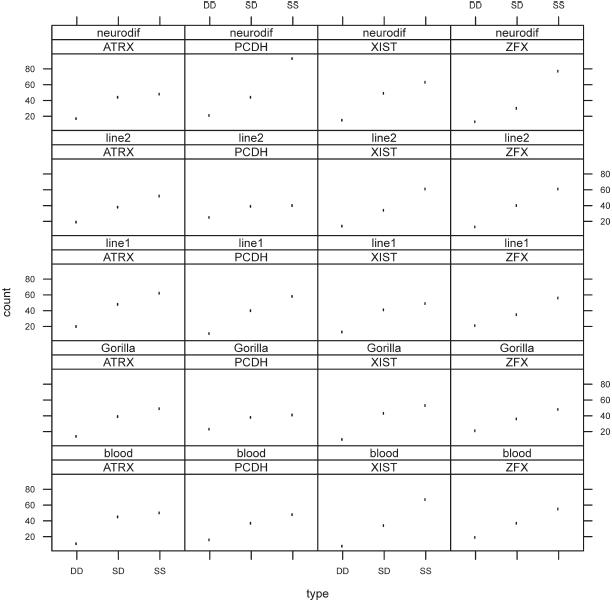
Graphic representation of the test carried out on the hypothesis of no differences in replication profile between all four X-linked genes (ATRX, PCDH11X, XIST and ZFX) in female human blood cells, female human B cells (line 1 and line 2), female gorilla B cells and female human differentiated neuroblastoma cells. Only differentiated neuroblastoma cells showed significant differences between the genes.
Expression of PCDH11 was not detected by RT-PCR in any of the cell lines analysed, but only in female brain cells used as control (Figure 3). On the contrary, expression of ATRX, XIST and ZFX was detected in all samples. Since the lack of expression of PCDH11 in cells expressing neuronal markers was unexpected, we decided to conduct further investigations. To confirm that there was no low-level expression of PCDH11 in the cell lines analysed we performed quantitative RT-PCR using SYBR green. The results in Table 2 reveal that the level of PCDH11 expression in the neuroblastoma samples before and after differentiation is negligible.
Figure 3.
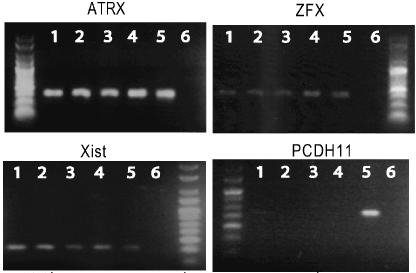
Expression of X-linked genes in cell lines. Samples are as follows: 1: DO208915 (female B-cell line), 2: GM08714 (female ICF B-cell line), 3: SHYSY5Y (female neuroblastoma cell line), 4: differentiated SHYSY5Y, 5: female brain control, 6: negative control. Molecular weight marker is HyperLadderV (bioline). Expression of control genes can be observed in all samples and all products are appropriate size. Expression of PCDH11 is observed only in female brain control sample.
Table 2.
Results of quantitative RT-PCR on cell line samples relative to female brain control; the expression level is negligible in the cell lines tested
| Sample | Relative expression |
|---|---|
| Neuroblastoma | |
| Differentiated neuroblastoma | 0.00001776 |
| Undifferentiated female brain | 0.00001874 |
| 1.00000000 |
Discussion
This is the first investigation on the replication timing properties of a gene pair (PCDH11X/Y) belonging to the non-pseudoautosomal X-Y homologous block (Xq21.3/Yp11.2), the largest shared region among sex chromosomes generated by a duplicative transposition from the X to the Y chromosome after the divergence of hominid and chimpanzee lineages.
By means of a comprehensive and statistically validated FISH assay we show that PCDH11 replicates asynchronously in both male and female human cells, as well as gorilla female cells. Replication asynchrony is usually associated with an allele-specific mode of expression, normally brought about by imprinting, X chromosome inactivation, or other mechanisms leading to monoallelic expression. Previous investigations on the replication timing of the human X chromosome, although leading in few circumstances to apparent discrepancies, have generally agreed in concluding that genes known to escape X-inactivation — either because pseudoautosomal or X-linked with Y homology or functional Y orthologues — replicate synchronously, while genes known to be inactivated replicate asynchronously (Boggs & Chinault 1994, Torchia et al. 1994, Hansen et al. 1995, 1996, Xiong et al. 1998, Yeshaya et al. 1999). Our observations showing homologous and allelic replication asynchrony for PCDH11 seem not to support the prediction, based on the X/Y homologous status of this gene in human, that PCDH11X might escape inactivation.
One possible explanation is that in fact, against the expectations, in human — as well as non-human primates — PCDH11X does not escape inactivation. Interestingly, autosomal monoalleleic expression has been reported for three other Protocadherin gene families (Pcdha, Pcdhb and Pcdhc), which reside in a single large cluster located on chromosome 5q, with their stochastic regulation believed to help specifying neuronal cell identity (Esumi et al. 2005).
It has recently become clear that escape from inactivation is a phenomenon more common than initially envisaged. A comprehensive X-inactivation study has shown that more than 15% of X-linked genes escape inactivation to some degree, with the proportion of genes escaping inactivation differing between different regions of the X chromosome, this occurrence reflecting the evolutionary history of the sex chromosomes (Carrel & Willard 2005). However, PCDH11X would not be the first case of an X-linked gene expected to escape that does not in fact conform. For instance, unlike other genes present in the Xq28 pseudoautosomal region (PAR2), both SYBL1, a synaptobrevin-like gene, and SPRY3, a human homologue of sprouty in Drosophila, have been reported to undergo inactivation on both X and Y (D'Esposito et al. 1996, Matarazzo et al. 2002, De Bonis et al. 2006).
An alternative interpretation of our results is that, notwithstanding its asynchronous replication profile, PCDH11X does escape inactivation, as recently published expression data would also suggest (Sudbrak et al. 2001, Lopes et al. 2006). Replication asynchrony for genes known to escape inactivation has been reported earlier. In particular, female cell lines with or without TIMP1 expression from the inactive X showed a similar extent of replication asynchrony for the two alleles of this gene (Anderson & Brown 2005). Also, in a study on the role of replication timing in X inactivation choice, all loci tested in undifferentiated ES cells replicated asynchronously, including the Smcx gene, which is known to escape X inactivation (Gribnau et al. 2005). Asynchronous replication of an X-linked pseudoautosomal locus was also reported by (Vorsanova et al. 2001).
A comprehensive study on the replication timing of the human genome carried out by microarray analysis has confirmed the existence of a significant positive correlation between the probability of gene expression and replication timing (Woodfine et al. 2004). However, from a cytological perspective the correspondence between expression of individual genes or alleles and replication timing, especially on chromosome X, appears less straightforward than generally believed, with different aspects of chromatin structure and intranuclear organization emerging, from a series of original investigations, closely intertwined with replication timing. First, the initial prediction that asynchronous replication might represent a general characteristic of imprinted genetic regions (Kitsberg et al. 1993) was subsequently challenged by Kawame et al. (1995), who, by using a bromodeoxyuridine method to detect replicated DNA by flow cytometry, studied the allele-specific replication for several sites within the human Prader-Willi/Angelman and IGF2/H19 imprinted regions in mouse, and showed that no obvious allele-specific differences in time of replication were detected at most loci previously reported to replicate asynchronously in the same cell type as determined by a FISH-based replication assay (Knoll et al. 1994, LaSalle & Lalande 1995). The authors suggested that FISH data implying replication asynchrony in nonexpressing cells reflect structural differences between the maternal and paternal alleles rather than differences in replication timing. Similar conclusions on replication imprinting reflecting differences in chromatin structure between homologues were also reached by Bickmore & Carothers (1995). More recently, asynchronous replication patterns of imprinted loci were found to be consistent with their differential localization to nuclear compartments with different replication characteristics (Gribnau et al. 2003).
Particularly significant evidence pointing at chromosome-pair non-equivalence and coordination of replication at the whole chromosome level was provided by a study by Ensminger & Chess (2004) on the replication timing of mono-allelically expressed genes along human autosomes and on chromosome X, confirming uniform late replication of loci on the inactive X. Gartler et al. (1999) had previously demonstrated that the silent XIST gene on the active chromosome X does replicate before the expressed allele on the inactive X, their results advocating for the first time the view that the replication timing of a gene is determined by the multi-replicon domain in which the gene resides, and not necessarily its expression status. It was recently demonstrated that asynchrony is present before initiation of X-inactivation and replication timing appears to be coordinated along the X chromosome over relatively large distances around the Xist locus (Gribnau et al. 2005).
Azuara et al. (2003) also conducted a study on leuckocyte-specific genes aimed at examining the relationship between gene silencing, chromatin structure and DNA replication. Their observations on integration of transgenes within heterochromatin provide further evidence for the importance of local chromosome environment in determining the timing of replication, and show that retarded sister-chromatid resolution is a feature of inactive chromatin.
Our findings on PCDH11X/Y and other X-linked genes are consistent with a model according to which the replication timing properties of X-linked genes are related principally to the epigenetic characteristics of the chromosomal environment in which the alleles reside. The four genes analysed in this study — two of them known to undergo inactivation (ATRX and XIST), one (ZFX) known to escape inactivation and PCDH11, the gene at the centre of our investigation, the inactivation status of which remains to be conclusively proved — show no significant differences when their replication profiles are compared in the majority of cell types analysed, and they consistently show similar levels of replication asynchrony in human and gorilla female cells. In our opinion this is to be attributed primarily to the ‘nonequivalence’ and temporal shift in replication between the active and inactive X chromosome, rather than to the expression status of each single locus, with the slightly variable levels of asynchrony between the genes reflecting uneven epigenetic changes along the inactive X. This conclusion seems to be substantiated by our observations that similar levels of asynchrony were observed when comparing the replication profile of PCDH11X to PCDH11Y — its homologue on chromosome Y — in human male cells, and also when carrying out the replication assay for PCDH11X in gorilla, in which this gene lacks X-Y homologous status and consequently is very likely to be inactivated. Our results also reconcile with observations by Gribnau et al. (2005) who — in their paper on the replication timing of the X chromosome before the onset of X inactivation — reported all the X-linked genes tested to replicate asynchronously.
The discrepancy between our observations on ZFX (SD > 30% in non-neuronal cells) and the results (SD < 20%) in a previous study on the same and other X-linked genes carried out by Boggs & Chinault (1994) is puzzling, but most probably due to slight differences in the protocols and parameters for the interpretation of the FISH results. Worth mention is that the original set of ‘dot-counting’ assays carried out by different research groups in the early 1990s seem to be all generally characterized by relatively low counts of single-double (SD) hybridization patterns. At the time the conclusion was reached that most of the homologous alleles replicate highly synchronously, with normally less than 10% of nuclei showing an SD pattern, this generally believed to be the result of sub-optimal hybridization conditions rather than an indication of replication asynchrony (Selig et al. 1992, Kitsberg et al. 1993, Boggs & Chinault 1994, Amiel et al. 1997). However, more recently, the perception of allele-specific mode of expression and replication has substantially changed, and it has become clear that asynchronous replication is a more common occurrence than expected, found in association not only with imprinting, but also with random mono-allelic expression, pathologies, tandem duplications as well as encountered in random assays (Chess et al. 1994, Amiel et al. 1998a,b, 2001, Smith & Higgs 1999, Mostoslavsky et al. 2001, Reish et al. 2002, 2003, Gimelbrant et al. 2005, Gimelbrant & Chess 2006). In conclusion, asynchronous replication — as a biological phenomenon — might have been initially underestimated, and relevant experimental evidence misinterpreted and over-cautiously discarded. This is also reflected by the fact that through the course of the years the percentage of SD generally perceived as the threshold for asynchrony has increased from the initial ‘>10%’ to the current ‘≥30%’ (Selig et al. 1992, Kitsberg et al. 1993, Goren & Cedar 2003, Gimelbrant & Chess 2006), with the distinction between asynchronous and synchronous replication appearing much less defined than previously thought.
Our investigations in neuronal markers expressing cell types — the retinoic acid differentiated neuroblastoma cells and the cerebral cortical cells — show an intriguing shift in the replication pattern of PCDH11X towards allelic synchrony, with a statistically significant difference between the SD value for PCDH11X in the neuroblastoma cell line before and after differentiation with retinoic acid, and also between the undifferentiated neuroblastoma cells and the cortical cells. This appears to be independent from the expression status of PCDHX in these cells, as shown by our real-time RT-PCR analysis. A statistically significant reduction in asynchrony was also observed when comparing the SD value for PCDH11X in differentiated neuroblastoma and cortical cells to all the other cell types analysed. Replication asynchrony of PCDH11X in differentiated neuroblastoma was significantly different from ATRX and XIST, but not different from ZFX and the autosomal control. Taken all together, these data seem to indicate a relaxation in the temporal control of allelic replication in neuroblastoma differentiated and cortical cells, probably due to tissue-specific epigenetic adjustments along the inactive X chromosome, although another explanation could be that, upon differentiation, global changes at the cell-cycle level could affect the replication profile of some genes. Also there seems to be no obvious link between replication properties of these X-linked genes and alleles, as inferred cytologically by FISH, and their expression status.
Finally, we observed no significant changes in the replication timing profiles of PCDH11X and ATRX in ICF cells, as demonstrated by similar percentages of SS, SD and DD patterns for both genes in ICF cells and controls. The ICF syndrome was previously reported to be characterized by significant disturbances in the inactive X methylation and by a shift in replication timing of some of the X-linked genes, and deemed to provide an important model system for the study of X chromosome inactivation (reviewed in Gartler & Hansen 2002). We found for PCDH11X and ATRX no cytological evidence of advanced replication timing as that previously shown by molecular methods for other X-linked genes in ICF (Hansen et al. 2000, Tao et al. 2002), and considered the major determinant of escape from inactivation in this syndrome. This is not necessarily unexpected as the phenotypic similarity of affected ICF males and females seems to suggest this to be a significant, but limited, phenomenon, not necessarily affecting all genes on the inactive X. However, it should also be pointed out that discrepancies between cytological and molecular investigations on the replication timing properties of the inactive X in ICF syndrome have been reported earlier (Bourc'his et al. 1999, Hansen et al. 2000).
In conclusion, our study provides novel and controversial data on the replication timing properties of a genomic region of evolutionary interest and aims to contribute to the debate on the complex relationship between X-inactivation, as a chromosome-wide phenomenon, and asynchrony of replication and expression status of single genes and alleles. Our investigations, combined with a critical review of the relevant literature, have allowed us to reassess the validity of the ‘dot-counting’ method as a cytological assay for replication profiling. Whilst confirming the soundness of this FISH-based approach — specifically when supported by a robust statistical analysis Y for a cell-by-cell assessment of replication properties of genes and alleles, our results highlight the potential risks of non-contextual and non-comparative ‘dot-counting’. Finally, by proving the unsuitability of this method to infer the inactivation status of X-linked genes, our analysis reaffirms the need for a combined molecular and cytological approach to reach definitive conclusions on interrelated replication and expression issues.
Acknowledgement
We are grateful to Dr Carole Sargent (Human Molecular Genetics Group, Division of Cellular and Molecular Pathology, University of Cambridge) for the gift of the PCDH11 probe, and to Dr Veronica Buckle (MRC Molecular Haematology Unit, Wheatherall Institute of Molecular Medicine, University of Oxford) for the ATRX probe. We also thank Dr Kostantinos Lymperopoulos-Klitsakis (Department of Human Anatomy and Genetics, University of Oxford) for the protocol for the differentiation of neuroblastoma cells, Professor A. Rickinson (Cancer Research Campaign Laboratory, University of Birmingham) for the gorilla cell line, and Dr Francis Marriott (Department of Statistics, University of Oxford) for helpful discussions on possible alternative approaches to the interpretation of the FISH data. Natalie Wilson and Emanuela Volpi are supported by the Wellcome Trust.
References
- Amiel A, Avivi L, Gaber E, Fejgin MD (1998a) Asynchronous replication of allelic loci in Down syndrome. Eur J Hum Genet6: 359-64. [DOI] [PubMed]
- Amiel A, Kolodizner T, Fishman A et al. (1998b) Replication pattern of the p53 and 21q22 loci in the premalignant and malignant stages of carcinoma of the cervix. Cancer83: 1966-971. [DOI] [PubMed]
- Amiel A, Levi E, Reish O, Sharony R, Fejgin MD (2001) Replication status as a possible marker for genomic instability in cells originating from genotypes with balanced rearrangements. Chromosome Res9: 611-16. [DOI] [PubMed]
- Amiel A, Litmanovich T, Gaber E, Lishner M, Avivi L, Fejgin MD (1997) Asynchronous replication of p53 and 21q22 loci in chronic lymphocytic leukemia. Hum Genet101: 219-22. [DOI] [PubMed]
- Anderson CL, Brown CJ (2005) Epigenetic predisposition to expression of TIMP1 from the human inactive X chromosome. BMC Genet6: 48. [DOI] [PMC free article] [PubMed]
- Azuara V, Brown KE, Williams RR et al. (2003) Heritable gene silencing in lymphocytes delays chromatid resolution without affecting the timing of DNA replication. Nat Cell Biol5: 668-74. [DOI] [PubMed]
- Bickmore WA, Carothers AD (1995) Factors affecting the timing and imprinting of replication on a mammalian chromosome. J Cell Sci108: 2801-809. [DOI] [PubMed]
- Blanco P, Sargent CA, Boucher CA, Howell G, Ross M, Affara NA (2001) A novel poly(A)-binding protein gene (PABPC5) maps to an X-specific subinterval in the Xq21.3/Yp11.2 homology block of the human sex chromosomes. Genomics74: 1-1. [DOI] [PubMed]
- Blanco P, Sargent CA, Boucher CA, Mitchell M, Affara NA (2000) Conservation of PCDHX in mammals; expression of human X/Y genes predominantly in brain. Mamm Genome11: 906-14. [DOI] [PubMed]
- Blanco-Arias P, Sargent CA, Affara NA (2002) The human-specific Yp11.2/Xq21.3 homology block encodes a potentially functional testis-specific TGIF-like retroposon. Mamm Genome13: 463-68. [DOI] [PubMed]
- Boggs BA, Chinault AC (1994) Analysis of replication timing properties of human X-chromosomal loci by fluorescence in situ hybridization. Proc Natl Acad Sci USA91: 6083-087. [DOI] [PMC free article] [PubMed]
- Bourc’his D, Miniou P, Jeanpierre M et al. (1999) Abnormal methylation does not prevent X inactivation in ICF patients. Cytogenet Cell Genet84: 245-52. [DOI] [PubMed]
- Brown CJ, Greally JM (2003) A stain upon the silence: genes escaping X inactivation. Trends Genet19: 432-38. [DOI] [PubMed]
- Carrel L, Willard HF (2005) X-inactivation profile reveals extensive variability in X-linked gene expression in females. Nature434: 400-04. [DOI] [PubMed]
- Chess A, Simon I, Cedar H, Axel R (1994) Allelic inactivation regulates olfactory receptor gene expression. Cell78: 823-34. [DOI] [PubMed]
- Chow JC, Yen Z, Ziesche SM, Brown CJ (2005) Silencing of the mammalian X chromosome. Annu Rev Genomics Hum Genet6: 69-2. [DOI] [PubMed]
- Crow TJ (2002) Handedness, language lateralisation and anatomical asymmetry: relevance of protocadherin XY to hominid speciation and the aetiology of psychosis. Point of view. Br J Psychiatry181: 295-97. [DOI] [PubMed]
- D’Esposito M, Ciccodicola A, Gianfrancesco F et al. (1996) A synaptobrevin-like gene in the Xq28 pseudoautosomal region undergoes X inactivation. Nat Genet13: 227-29. [DOI] [PubMed]
- De Bonis ML, Cerase A, Matarazzo MR et al. (2006) Maintenance of X- and Y-inactivation of the pseudoautosomal (PAR2) gene SPRY3 is independent from DNA methylation and associated to multiple layers of epigenetic modifications. Hum Mol Genet15: 1123-132. [DOI] [PubMed]
- Disteche CM, Filippova GN, Tsuchiya KD (2002) Escape from X inactivation. Cytogenet Genome Res99: 36-3. [DOI] [PubMed]
- Drouin R, Lemieux N, Richer CL (1990) Analysis of DNA replication during S-phase by means of dynamic chromosome banding at high resolution. Chromosoma99: 273-80. [DOI] [PubMed]
- Dutrillaux B, Couturier J, Richer CL, Viegas-Pequignot E (1976) Sequence of DNA replication in 277 R- and Q-bands of human chromosomes using a BrdU treatment. Chromosoma58: 51-1. [DOI] [PubMed]
- Ensminger AW, Chess A (2004) Coordinated replication timing of monoallelically expressed genes along human autosomes. Hum Mol Genet13: 651-58. [DOI] [PubMed]
- Esumi S, Kakazu N, Taguchi Y et al. (2005) Monoallelic yet combinatorial expression of variable exons of the protocadherin-alpha gene cluster in single neurons. Nat Genet37: 171-76. [DOI] [PubMed]
- Gartler SM, Goldstein L, Tyler-Freer SE, Hansen RS (1999) The timing of XIST replication: dominance of the domain. Hum Mol Genet8: 1085-089. [DOI] [PubMed]
- Gartler SM, Hansen RS (2002) ICF syndrome as a model system for studying X chromosome inactivation. Cytogenet Genome Res99: 25-9. [DOI] [PubMed]
- Gimelbrant AA, Chess A (2006) An epigenetic state associated with areas of gene duplication. Genome Res16: 723-29. [DOI] [PMC free article] [PubMed]
- Gimelbrant AA, Ensminger AW, Qi P, Zucker J, Chess A (2005) Monoallelic expression and asynchronous replication of p120 catenin in mouse and human cells. J Biol Chem280: 1354-359. [DOI] [PubMed]
- Goren A, Cedar H (2003) Replicating by the clock. Nat Rev Mol Cell Biol4: 25-2. [DOI] [PubMed]
- Gribnau J, Hochedlinger K, Hata K, Li E, Jaenisch R (2003) Asynchronous replication timing of imprinted loci is independent of DNA methylation, but consistent with differential subnuclear localization. Genes Dev17: 759-73. [DOI] [PMC free article] [PubMed]
- Gribnau J, Luikenhuis S, Hochedlinger K, Monkhorst K, Jaenisch R (2005) X chromosome choice occurs independently of asynchronous replication timing. J Cell Biol168: 365-73. [DOI] [PMC free article] [PubMed]
- Haaf T (1997) Analysis of replication timing of ribosomal RNA genes by fluorescence in situ hybridization. DNA Cell Biol16: 341-45. [DOI] [PubMed]
- Hansen RS, Canfield TK, Fjeld AD, Gartler SM (1996) Role of late replication timing in the silencing of X-linked genes. Hum Mol Genet5: 1345-353. [DOI] [PubMed]
- Hansen RS, Canfield TK, Gartler SM (1995) Reverse replication timing for the XIST gene in human fibroblasts. Hum Mol Genet4: 813-20. [DOI] [PubMed]
- Hansen RS, Stoger R, Wijmenga C et al. (2000) Escape from gene silencing in ICF syndrome: evidence for advanced replication time as a major determinant. Hum Mol Genet9: 2575-587. [DOI] [PubMed]
- Izumikawa Y, Naritomi K, Hirayama K (1991) Replication asynchrony between homologs 15q11.2: cytogenetic evidence for genomic imprinting. Hum Genet87: 1-. [DOI] [PubMed]
- Kawame H, Gartler SM, Hansen RS (1995) Allele-specific replication timing in imprinted domains: absence of asynchrony at several loci. Hum Mol Genet4: 2287-293. [DOI] [PubMed]
- Kitsberg D, Selig S, Brandeis M et al. (1993) Allele-specific replication timing of imprinted gene regions. Nature364: 459-63. [DOI] [PubMed]
- Knoll JH, Cheng SD, Lalande M (1994) Allele specificity of DNA replication timing in the Angelman/Prader-Willi syndrome imprinted chromosomal region. Nat Genet6: 41-6. [DOI] [PubMed]
- LaSalle JM, Lalande M (1995) Domain organization of allele-specific replication within the GABRB3 gene cluster requires a biparental 15q11-13 contribution. Nat Genet9: 386-94. [DOI] [PubMed]
- Lopes AM, Ross N, Close J, Dagnall A, Amorim A, Crow TJ (2006) Inactivation status of PCDH11X: sexual dimorphisms in gene expression levels in brain. Hum Genet119: 267-75. [DOI] [PubMed]
- Lyon MF (1961) Gene action in the X-chromosome of the mouse (Mus musculus L.). Nature190: 372-73. [DOI] [PubMed]
- Matarazzo MR, De Bonis ML, Gregory RI et al. (2002) Allelic inactivation of the pseudoautosomal gene SYBL1 is controlled by epigenetic mechanisms common to the X and Y chromosomes. Hum Mol Genet11: 3191-198. [DOI] [PubMed]
- Mostoslavsky R, Singh N, Tenzen T et al. (2001) Asynchronous replication and allelic exclusion in the immune system. Nature414: 221-25. [DOI] [PubMed]
- Reish O, Gal R, Gaber E, Sher C, Bistritzer T, Amiel A (2002) Asynchronous replication of biallelically expressed loci: a new phenomenon in Turner syndrome. Genet Med4: 439-43. [DOI] [PubMed]
- Reish O, Orlovski A, Mashevitz M et al. (2003) Modified allelic replication in lymphocytes of patients with neurofibromatosis type 1. Cancer Genet Cytogenet143: 133-39. [DOI] [PubMed]
- Ross MT, Grafham DV, Coffey AJ et al. (2005) The DNA sequence of the human X chromosome. Nature434: 325-37. [DOI] [PMC free article] [PubMed]
- Schwartz A, Chan DC, Brown LG et al. (1998) Reconstructing hominid Y evolution: X-homologous block, created by X-Y transposition, was disrupted by Yp inversion through LINE–LINE recombination. Hum Mol Genet7: 1-1. [DOI] [PubMed]
- Selig S, Okumura K, Ward DC, Cedar H (1992) Delineation of DNA replication time zones by fluorescence in situ hybridization. Embo J11: 1217-225. [DOI] [PMC free article] [PubMed]
- Singh N, Ebrahimi FA, Gimelbrant AA et al. (2003) Coordination of the random asynchronous replication of autosomal loci. Nat Genet33: 339-41. [DOI] [PubMed]
- Smith ZE, Higgs DR (1999) The pattern of replication at a human telomeric region (16p13.3): its relationship to chromosome structure and gene expression. Hum Mol Genet8: 1373-386. [DOI] [PubMed]
- Sudbrak R, Wieczorek G, Nuber UA et al. (2001) X chromosome-specific cDNA arrays: identification of genes that escape from X-inactivation and other applications. Hum Mol Genet10: 77-3. [DOI] [PubMed]
- Takebayashi S, Sugimura K, Saito T et al. (2005) Regulation of replication at the R/G chromosomal band boundary and pericentromeric heterochromatin of mammalian cells. Exp Cell Res304: 162-74. [DOI] [PubMed]
- Tao Q, Huang H, Geiman TM et al. (2002) Defective de novo methylation of viral and cellular DNA sequences in ICF syndrome cells. Hum Mol Genet11: 2091-102. [DOI] [PubMed]
- Torchia BS, Call LM, Migeon BR (1994) DNA replication analysis of FMR1, XIST, and factor 8C loci by FISH shows nontranscribed X-linked genes replicate late. Am J Hum Genet55: 96-04. [PMC free article] [PubMed]
- Vogel W, Autenrieth M, Mehnert K (1989) Analysis of chromosome replication by a BrdU antibody technique. Chromosoma98: 335-41. [DOI] [PubMed]
- Vorsanova SG, Yurov YB, Kolotii AD, Soloviev IV (2001) FISH analysis of replication and transcription of chromosome X loci: new approach for genetic analysis of Rett syndrome. Brain Dev23(Suppl. 1): S191-95. [DOI] [PubMed]
- Williams NA, Close JP, Giouzeli M, Crow TJ (2006) Accelerated evolution of protocadherin11x/Y: a candidate gene-pair for cerebral asymmetry and language. Am J Med Genet B Neuropsychiatr Genet141: 623-33. [DOI] [PubMed]
- Wilson ND, Ross LJ, Crow TJ, Volpi EV (2006) PCDH11 is X/Y homologous in Homo sapiens but not in Gorilla gorilla and Pan troglodytes. Cytogenet Genome Res114: 137-39. [DOI] [PubMed]
- Woodfine K, Fiegler H, Beare DM et al. (2004) Replication timing of the human genome. Hum Mol Genet13: 191-02. [DOI] [PubMed]
- Xiong Z, Tsark W, Singer-Sam J, Riggs AD (1998) Differential replication timing of X-linked genes measured by a novel method using single-nucleotide primer extension. Nucleic Acids Res26: 684-86. [DOI] [PMC free article] [PubMed]
- Yeshaya J, Shalgi R, Shohat M, Avivi L (1999) FISH-detected delay in replication timing of mutated FMR1 alleles on both active and inactive X-chromosomes. Hum Genet105: 86-7. [DOI] [PubMed]
- Yoshida K, Sugano S (1999) Identification of a novel protocadherin gene (PCDH11) on the human XY homology region in Xq21.3. Genomics62: 540-43. [DOI] [PubMed]