

Abstract
Lipid droplets are the least characterized of cellular organelles. Long considered simple lipid storage depots, these dynamic and remarkable organelles have recently been implicated in many biological processes, and we are only now beginning to gain insights into their fascinating lives in cells. Here we examine what we know of the life of lipid droplets. We review emerging data concerning their cellular biology and present our thoughts on some of the most salient questions for investigation.
Keywords: Adiposome, Triglyceride, Steryl ester, Fat storage, Organelle dynamic
The compartmentalization of biochemical reactions is fundamental to living organisms. At the cellular level, this is achieved by separating different functions, such as gene regulation, protein secretion and degradation, energy generation, and storage of macromolecules, into organelles. Most organelles comprise one or more aqueous compartments that are bounded by a lipid bilayer, which defines biochemically distinct spaces.
Notable exceptions to this paradigm are lipid droplets (also called lipid bodies, fat bodies, or adiposomes). Lipid droplets have a unique physical structure: they are delimited by a single phospholipid monolayer that separates the aqueous and hydrophobic phases within the cell. The hydrophobic core contains neutral lipids, most notably triacylglycerols (TG) and sterol esters. These are crucial substances for cells. TG is the major form of energy storage, and both TG and sterol esters serve as reservoirs of membrane lipid components.
Most eukaryotic cells have lipid droplets (examples in Fig. 1). Indeed, in many cells they occupy a considerable portion of cell volume and mass. In plants and microorganisms such as algae, lipid droplets contain oils that are of great importance for the food and energy industries. Lipid droplets are also prominent in many types of mammalian cells, with adipocytes being the most highly specialized for lipid and energy storage. In fact, white adipocytes are so filled with one or more lipid droplets that other organelles are not easily seen in micrographs. Obesity, in which the number and size of adipocytes are excessive, is essentially a disease of lipid droplet excess. In addition, hepatocytes, heart and skeletal myocytes, adrenocortical cells, enterocytes, and macrophages all may contain large amounts of lipid droplets. Lipid droplets are found in cells from yeast to humans, and the molecular processes that govern droplet biology are likely to be conserved.
Fig. 1.
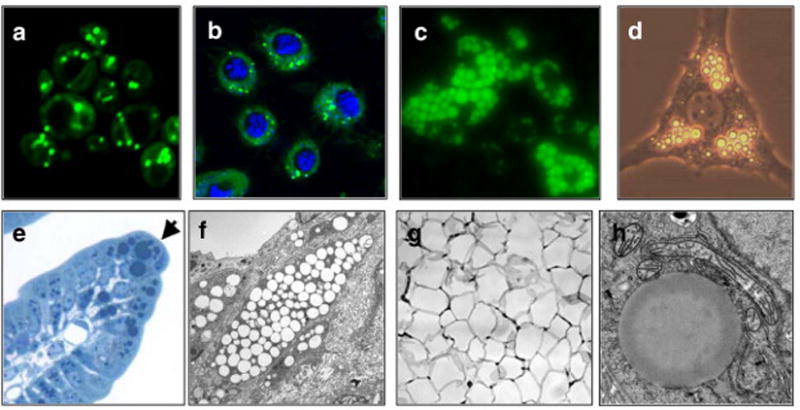
Examples of lipid droplets in eukaryotic cells. Lipid droplets stained with BODIPY (green) in (a) S. cerevisiae, (b) Drosophila S2 cells [nuclei are stained blue with 4′,6-diamidino-2-phenylindole (DAPI)], and (c) murine adipocytes derived from embryonic fibroblasts (image courtesy of R. Streeper). (d) Lipid droplets (bright round organelles) in a single adipocyte derived from OP9 cells (phase contrast, image courtesy of C. Harris). (e) Lipid droplets stained with osmium tetroxide in intestinal enterocytes (arrow indicates an enterocyte that is filled with numerous dark-stained lipid droplets). (f) Electron micrograph of macrophage foam cells in a murine atherosclerotic lesion. Lipid droplets appear as round, empty objects. (g) Section of murine white adipose tissue showing large unilocular lipid droplets (empty spaces) that occupy most of the cytoplasm in white adipocytes. (h) High magnification electron micrograph of a single lipid droplet (large amorphous sphere) in a rat hepatoma cell (image courtesy of S. Stone and J. Wong).
Despite their importance in energy metabolism and disease, lipid droplets have been grossly understudied and are probably the least understood cellular organelle. This reflects in part an assumption that they are relatively inert organelles, serving simply as lipid storage depots. This notion has been reinforced by the term lipid droplet, which suggests a static and passive collection of oil in the aqueous cellular environment. Interestingly, for many years these organelles were called “liposomes,” a more dynamic name that was usurped by artificial liposomes in 1968 [1]. Recent functional analyses of droplets [2], [3] and [4] and detailed analyses of their protein composition [summarized in [5]] suggest a radically different view. Indeed, lipid droplets are emerging as highly dynamic organelles that play crucial roles in cellular energy homeostasis and lipid metabolism. Here we examine the life of lipid droplets, focusing on their newly discovered properties and highlighting fundamental unanswered questions about their biology.
1. Vital statistics
1.1. Lipid composition
Lipid droplets are bounded by a protein-decorated phospholipid monolayer that envelopes a neutral lipid core. In mammalian cells, phosphatidylcholine is the most abundant phospholipid, followed by phosphatidylethanolamine and phosphatidylinositol [6]. Compared with total membranes, droplet phospholipids contain more lysophospholipids but less sphingomyelin and phosphatidic acid. The fatty acid composition of phospholipids in lipid droplets of human hepatoma cells differs from those of the endoplasmic reticulum (ER), and they contain relatively more free cholesterol [7]. Lipid droplets vary dramatically in size, from a diameter of 0.1 μm in yeast to over 100 μm in a white adipocyte. This translates to a staggering range of nine orders of magnitude in volume of neutral lipids that can occupy the core. The hydrophobic interior of lipid droplets comprises mainly TG and sterol esters, although in some cell types retinyl esters, waxes, or diacylglycerols are found. Lipid droplets in yeast contain 50% TG and 50% sterol esters, whereas adipocytes contain primarily TG, and macrophage “foam cells” contain mostly sterol esters. Ether lipids, such as monoalk(en)yl diacylglycerol, make up 10–20% of lipids in some mammalian cells [6].
1.2. Protein composition
The unique structure–an organic lipid phase surrounded by a single layer of polar lipids–poses many interesting and so far unanswered questions concerning lipid droplet-associated proteins. For example, how are proteins, such as those that maintain droplet structure, traffic lipids, or regulate lipolysis, targeted to the surfaces of droplets? The normal conformation of transmembrane proteins, with water-soluble domains on both sides of a membrane bilayer, should not allow targeting of these proteins to the monolayer droplet surface. One solution to this problem is suggested by the multifunctional caveolin protein, which has been localized to both the plasma membrane and lipid droplets (for reviews, see [8], [9], [10] and [11]). Caveolin possesses an unusually long central hydrophobic membrane helix that is embedded in the membrane, with both of its termini exposed to the cytoplasm [12] and [13]. Acyl CoA:diacylglycerol acyltransferase 2 (DGAT2), the major enzyme catalyzing TG synthesis, also localizes to both the ER membrane and lipid droplets [14] and has a similar topology as caveolin [15]. This type of structure (i.e., a long internal hydrophobic stretch) may enable such proteins to be embedded in either bilayers or monolayers.
1.3. PAT proteins
The PAT proteins (perilipin, adipophilin/adipose differentiation-related protein, TIP47, and other related proteins) [16], which have structural and regulatory functions, also target to lipid droplets, likely through different mechanisms. Some PAT proteins localize exclusively to droplets (e.g., adipophilin). Others are found both in the cytosol and on lipid droplets. Clues to the mechanism of this complex targeting come from a recent crystal structure of the C-terminal half of TIP47 [17]. This structure revealed a four-helix bundle with great similarity to the N-terminal domain of apolipoprotein (apo) E [18]. Upon binding to lipids, the apoE four-helix bundle opens to expose amphipathic helixes that can bind the monolayer surfaces of lipoproteins [19]. In an analogous manner, PAT proteins may bind to lipid droplets by embedding hydrophobic helices into the droplet surface. PAT proteins also share a common structural element: N-terminal 11-mer repeats that have an amphipathic helical structure [17]. Although the structure of these domains has not been determined, they likely also bind to lipids at the surfaces of lipid droplets. In adipocytes, TIP47 and S3-12 are found on smaller droplets, and adipophilin and perilipin are found on larger droplets [20]. It is not clear why different PAT proteins distribute to different sized lipid droplets. Possible explanations include that different PAT proteins are best suited to different degrees of membrane curvature (i.e., corresponding to droplet size) or that they bind to different droplet-associated proteins or to different polar head groups of monolayer phospholipids.
The structural properties underlying the binding of PAT proteins to lipid droplets offer many possibilities for regulating this localization. Indeed, the localization of several proteins, such as TIP47, to lipid droplets is highly regulated and can be induced by adding fatty acids to the medium to trigger droplet formation (reviewed in [21]). The mechanism of this regulation remains largely unclear. One possibility is that a posttranslational modification alters the conformation of a lipid-binding domain. Indeed, the targeting of Nir2, a peripheral Golgi protein, to lipid droplets appears to be regulated by phosphorylation [22].
In addition to the few lipid droplet-specific proteins, such as the PAT proteins, many cellular proteins with known functions have been identified in lipid droplet fractions by proteomic studies [21], [23], [24] and [25]. Some of these are involved in dynamic cellular processes, suggesting that they play similar roles in lipid droplet biology. However, both the localization of most of these proteins to droplets and their functional roles remain to be validated.
2. Birth of a lipid droplet
Relatively little is known about the most basic biological features of lipid droplets. The molecular processes that govern their formation, growth, movement, fusion, fission, interactions with other organelles, and utilization are mostly unclear. First, we consider the birth of lipid droplets.
2.1. Clues to lipid-droplet origins
Where do lipid droplets come from? Are they autonomously replicating organelles (like mitochondria) or are they derived from another organelle? In prokaryotes, lipid droplets appear to be formed from the plasma membrane [26]. In eukaryotes, evidence suggests that lipid droplets originate from the ER. Electron micrographs from a variety of cell types show that lipid droplets are often tightly associated and sometimes appear to be connected with the ER. In some instances, sheets of ER membrane even partially surround the droplet, cradling it like an eggcup [27]. In addition, the enzymes [28] that catalyze the synthesis of neutral lipids in the droplet core are localized predominantly to the ER. In fact, DGAT2, the major enzyme catalyzing TG synthesis [29] and [30], localizes to areas where droplets are tightly associated with ER under conditions that promote droplet formation [14]. This suggests that the ER–lipid droplet association facilitates the transfer of TG and other newly synthesized lipids to the droplet core. Consistent with the notion that such contact zones are sites of active lipid synthesis, mitochondria are often localized near lipid droplets (for examples see [3], [31] and [32]), as are specialized compartments of the ER called mitochondria-associated membranes (MAM), which are thought to be centers of lipid synthesis [4]. MAM and mitochondria may provide energy and NADPH that facilitate TG synthesis and lipid droplet biogenesis. Thus, although the bulk of evidence suggests that the ER gives birth to droplets, other organelles may contribute to their maturation.
2.2. Theories of lipid-droplet biogenesis
If we assume that droplets are derived from the ER or a similar bilayer, an obvious question arises: how do bilayers give rise to organelles enclosed by monolayers? The answer is unknown, but several models for the formation of lipid droplets have been proposed (Fig. 2).
Fig. 2.
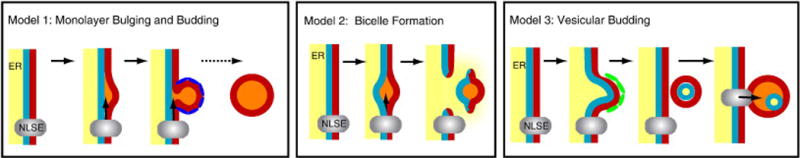
Models of lipid-droplet biogenesis. (Left) Model 1: Lipid droplet biogenesis by ER budding. Neutral lipids (orange) are synthesized by neutral lipid-synthesizing enzymes (NLSE) and bulge from the outer leaflet of the ER membrane (red). The nascent droplet may be coated by proteins (dark blue) that facilitate the budding process. (Middle) Model 2: Bilayer excision. Newly synthesized neutral lipids accumulate between the inner (blue) and outer (red) leaflets of the ER membrane and cause bulging. This entire lipid lens is then excised from the ER, leaving a transient hole in the membrane. ER contents (yellow) might leak through this hole into the cytosol. (Right) Model 3: Vesicular budding. A vesicle containing both leaflets of the ER membrane (red and blue) and a lumen (yellow) is formed by the vesicular budding machinery (green) at the ER membrane. The vesicle is subsequently tethered to the ER, where NLSEs (grey) fill the intramembranous space with neutral lipids (orange). The luminal space (yellow) is compressed, and its contents may leak into the cytosol. This process may trap luminal proteins within a compartment of the lipid droplet.
The most widely cited model posits that lipid droplets form in the ER membrane as a result of an accumulation of neutral lipids within the bilayer that drives a “budding” of a nascent droplet from the cytosolic leaflet of the membrane (Fig. 2, Model 1). This process may be driven by neutral lipids that accumulate and bulge out of the membrane or by cytosolic factors that bind to the ER leaflet and facilitate the budding reaction by acting as coatamer-like proteins. Such factors may be PAT proteins [16], which could mediate budding in specialized domains of the ER. How the neutral lipid molecules are channeled into the droplet and prevented from diffusing through the hydrophobic phase in the membrane bilayer and the mechanism of budding is unclear. In a variation of this model, the droplets may never be completely severed from the ER but might remain connected to the outer leaflet of the ER membrane [33].
An alternative model was recently put forth in which lipid droplets are excised from both leaflets of the ER membrane bilayer as a bicelle (Fig. 2, Model 2) [34]. An attractive feature of this model is that the excision process could mediate the extrusion of proteins destined for ER-associated degradation (ERAD) of proteins by the proteosome [34], implicating a functional link between lipid-droplet formation and protein degradation. An excision mechanism could also help to explain how viruses, such as polyoma and simian virus 40, escape from the ER lumen to the cytosol as large assembled particles. A caveat of this model is that the droplet-excision process would violate the integrity of the ER membrane, effectively creating a transient hole during droplet formation. In this case, calcium ions otherwise used as second messengers could leak out of the ER lumen and the unique redox environment of the ER could locally collapse. It is also unclear how excised droplets would be directed toward the cytosol rather than the ER lumen.
In a third model that we call vesicular budding (Fig. 2, Model 3), droplets are initially formed within small bilayer vesicles, utilizing the machinery of vesicle formation of the secretory pathway. This might occur in a specialized domain of ER that is distinct from vesicular transport and dedicated to lipid synthesis. In this scenario, a nascent vesicle is formed and remains tethered in close apposition to the ER membrane. Neutral lipids could fill the vesicle while it is still continuous with the ER membrane or after vesicle budding, via a shuttling mechanism to a vesicle that is tethered to the cytoplasmic side of the ER. Either process would gradually fill the intermembrane space to the point where the vesicular lumen is either obliterated or only makes up a small part of the lipid droplet. Any remaining lumen of the vesicle could then either fuse with the outer leaflet of the droplet or remain as an inclusion of aqueous material within the droplet. The latter possibility could explain why hydrophilic proteins within the lipid droplet core have been identified by electron microscopy and freeze-facture techniques [35]. Indeed, to explain such findings, a model that posits a similar manner of lipid filling has been proposed [36]. The transfer of lipids to growing droplets is likely, since at least DGAT2 is localized to zones near droplets, and triglycerides synthesized by DGAT2 can directly be incorporated into droplets [14]. As the droplets enlarge, coatamer proteins would presumably dissociate from the droplet surface, possibly being replaced by PAT proteins, which would afford structural stability.
The involvement of coatamer proteins in this model is speculative, but there is evidence to support their role in droplet formation. Knockdown of the COPI/Arf1 vesicular transport machinery by siRNAi interferes with droplet formation [2], and prolonged incubation of cells with brefeldin A, which inhibits Arf1 GTPases, eventually depletes Drosophila cells of lipid droplets (Guo Y, Walther T, Farese R, unpublished observations) and inhibits droplet formation in NIH 3T3-L1 adipocytes [37]. Also, the vesicular budding model predicts features of lipid-droplet dynamics that could be variations of ER dynamics. For example, lipid-droplet fusion could occur through an ER fusion-like reaction. According to the most accepted model, when two bilayer membranes fuse, SNARE proteins bring the two bilayers in close proximity, leading to mixing of the two outer leaflets of the membranes. This results in a hemifusion intermediate, which is resolved for bilayers by generation of a pore through the fusion of the inner leaflets. For monolayer-bound droplets, the first step in this process, perhaps mediated by similar SNARE machinery, would theoretically suffice to mediate the fusion of lipid droplets. This model of droplet fusion is supported by the large overlap in the machinery required for lipid droplet fusion and the ER fusion machinery [38].
2.3. Growth of lipid droplets involvement of multiple organelles?
Once droplets are formed, how do they accumulate more neutral lipids in their cores and phospholipids at their surfaces? Evidence supports a role for the ER, as well as additional organelles, in this process.
2.3.1. Endoplasmic reticulum
As the presumed major location for neutral lipid synthesis, the ER almost certainly plays a role in droplet growth. The enzymes that catalyze this synthesis are integral membrane proteins, which presumably encounter the substrates for TG synthesis, such as diacylglycerol and fatty acyl CoA, at the cytosolic surface of the ER. These enzymes are members of two families of proteins. One is the membrane-bound O-acyltransferases [39], which include DGAT1 and the acyl CoA:cholesterol acyltransferases. The latter enzymes catalyze the synthesis of TG and cholesterol esters, respectively [40]. The yeast orthologs of these enzymes, Are1 and Are2, catalyze the synthesis of ergosterol esters [41]. These enzymes have multiple hydrophobic regions and are localized primarily to the ER. The other family of enzymes is related to DGAT2 [29] and [30] and includes acyl CoA:monoacylglycerol acyltransferases (MGAT) and wax synthases. These proteins are less hydrophobic. DGAT2 has one long hydrophobic domain that appears to mediate embedding of the protein in the ER bilayer [15]. DGAT2 localizes primarily to the ER but, when cells are treated with oleic acid, also localizes to the surfaces of lipid droplets [[14]; Stone S, Farese R, unpublished data]. Similarly, activity for the yeast ortholog of DGAT2, Dga1, has been found in both the ER and lipid droplets [42]. Intriguingly, overexpression of DGAT2 in cultured mammalian cells yields droplets that are much larger and closer to the nucleus than those obtained by overexpression of DGAT1 [30]. A possible explanation for larger droplets is that the substantial increase in TG mass catalyzed by DGAT2 diverts diacylglycerol toward neutral lipids and away from phospholipid synthesis. This would favor an increase in core lipids (volume) over surface lipids and possibly promote the fusion of droplets [2].
A major question is how lipids are transferred from the site of synthesis to lipid droplets. In the simplest model, the droplets remain tethered to the ER, and the monolayer surrounding them is continuous with the ER bilayer. In this case, lipids could diffuse laterally into the droplet. However, if lipid droplets accumulate neutral lipids after detaching from the ER, these lipids must either be synthesized at the droplet surface or be transferred to droplets by a dedicated mechanism that might involve specific transfer proteins, analogous to ceramide transfer proteins, glycerolipid transfer proteins, or oxysterol binding proteins [43], [44], [45] and [46]. Proteins mediating the cytosolic transfer of neutral lipids have not been reported, although the microsomal triglyceride transfer protein mediates transfer of neutral lipids within the ER lumen. Notably, neither lipid-transfer proteins nor enzymes that synthesize neutral lipids have been detected in lipid-droplet proteomes. Another possibility is that, after detaching, lipid droplets grow exclusively through fusion.
ER involvement in lipid-droplet maturation is also supported by recent evidence from screens for genes that regulate lipid droplets in yeast [47] and [48]. Two independent screens of the yeast deletion collection identified numerous genes involved in droplet biology, and both found that the ER protein seipin (also called FLD1) plays a role in droplet formation; in seipin-deficient cells, droplets are abnormal, appearing “immature” and clustered.
2.3.2. Plasma membrane
Other data suggest that lipid-droplet biogenesis or growth might occur at the plasma membrane. In macrophages, for example, the PAT proteins are integral to the plasma membrane and form specialized domains that are associated with lipid droplets [27]. Similarly, a subclass of caveolae, flask-shaped invaginations of the plasma membrane that are involved in endocytosis, has been implicated in the synthesis of TG and lipid-droplet formation in adipocytes [49]. Moreover, both caveolins and the related flotillins/reggie-proteins that are involved in plasma membrane organization and endocytosis, have been detected on lipid droplets [23], [50] and [51]. This localization of caveolin is likely to be functionally important because embryonic fibroblasts from caveolin knockout mice show impaired lipid droplet formation [3]. In addition, these knockout mice exhibit defects in lipolysis [3] and greatly reduced lipid storage in hepatocytes during liver regeneration [52]. How caveolin participates in droplet formation is unknown. One possibility is by coupling the uptake of fatty acids with their storage in lipid droplets, since either expression of a dominant negative mutant of caveolin-3 or disassembly of caveolae by incubation with the sterol-binding drug β-cyclodextrin inhibits uptake of long chain fatty acids into cells [53].
A connection between lipid droplets and the plasma membrane might be an ancient feature of lipid-droplet biology. Bacteria lack an ER, and in certain bacteria that accumulate neutral lipids, lipid droplets appear to form at the plasma membrane [26]. What purpose would lipid-droplet formation at mammalian plasma membranes serve? One possibility is to deal with large amounts of exogenous fatty acids that many cells take up from the extracellular space. These fatty acids must be rapidly converted to their CoA forms, esterified, and stored to prevent lipotoxic effects.
3. A dynamic life
3.1. Trafficking and interactions with other organelles
Recent discoveries indicate that the dynamics of lipid droplets are complex (Fig. 3). For example, endosomes [54] and peroxisomes [55] apparently interact with lipid droplets. At least for endosomes, Rab GTPases may be important for these interactions. Nine “tethering” Rab GTPases were identified in the lipid-droplet proteome [23]. Of these, Rab5 mediates interactions with endosomes and early endosome effectors such as EEA1; Rab11 establishes the identity of recycling endosomes and modulates cholesterol transport [56], and Rab18 may regulate the interaction of lipid droplets with the ER [57]. Whether Rab-mediated interactions are crucial for lipid-droplet trafficking or formation is so far unclear.
Fig. 3.
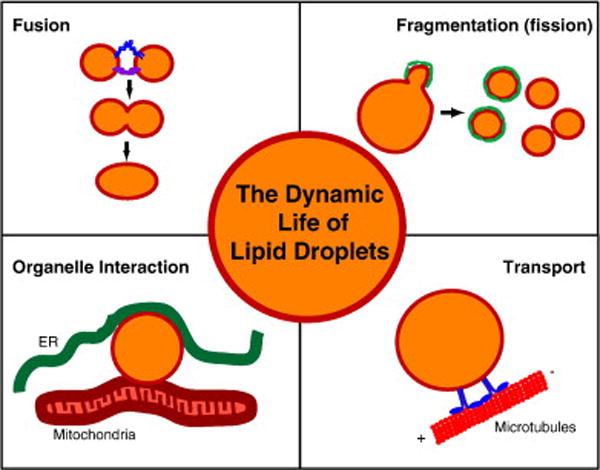
Dynamic processes linked to lipid-droplet biology. Different events in the dynamic life of a lipid droplet (orange interior representing the neutral lipids and a red line indicating the bounding monolayer of phospholipids) are shown in the four panels. In the “fusion” panel, blue and purple protein complexes represent SNARE proteins and tethering complexes, respectively. In the “fragmentation” panel, green proteins covering the lipid droplet during fragmentation indicate COPI/Arf1 coat proteins. During lipid droplet transport, motor proteins (blue) mediate the movement of droplets along microtubules (red).
A role for Arf1-type GTPases in lipid-droplet dynamics is also emerging. Arf1 was identified in a lipid droplet proteome [5] and localizes in part to the surfaces of lipid droplets in Drosophila cells [2]. RNAi-mediated knockdown of the Arf1-COPI machinery, which forms coated vesicles at Golgi and endosomal membranes but not of other transport machinery proteins, leads to larger and more dispersed lipid droplets and a defect in lipolysis [2]. It is tempting to speculate that active Arf1-GTP is recruited to the surfaces of lipid droplets and, via recruitment of COPI machinery, mediates budding of smaller lipid droplets with increased surface area, thereby facilitating hydrolysis of lipids by lipases. Impairing this mechanism through knockdowns would diminish lipolysis and shift the equilibrium between lipid deposition and mobilization such that individual droplets would grow larger. Activated Arf proteins recruit phospholipase D (PLD) [37], which generates phosphatidic acid from phospholipids that could alter membrane curvature. Studies of lipid droplets in a cell-free system suggest that PLD is important in droplet formation [58]. Another report suggested that PLD is activated by Arf1-GDP at lipid droplets [37]. However, knockdown of PLD has no effect on lipid droplets in Drosophila cells (Guo Y, Walther T, unpublished observations), and activity of the GDP-bound form of a small GTPase is unprecedented.
Two important points emerge from these early glimpses into droplet dynamics. First, it is crucial to make a clear distinction between de novo formation of lipid droplets and lipid-droplet dynamics and trafficking. Assays that enable the separate study of these processes would be highly valuable. Second, lipid droplets are likely to be heterogeneous in composition and function in different cell types and within individual cells. This point is underscored by the observation of differential localization of PAT proteins to oleate-loaded 3T3-L1 adipocytes, with Tip47 and S3-12 localized to the smallest droplets, adipophilin to intermediate size droplets, and perilipin to the largest droplets [20]. To better analyze the composition and function of different droplets, a purification protocol that separates droplets into different classes, similar to that used for plasma lipoproteins, is needed.
3.2. Movement
Imaging of droplets in living cells often shows them as relatively immobile organelles that move rapidly within small areas. However, in some instances, their movements are clearly regulated. For example, in Drosophila embryos, droplets move coordinately to and from the periphery of the embryo in a manner that is dependent on microtubules and dynein [59]. This transport is regulated by the phosphorylation of Lsd2, a Drosophila ortholog of perilipin [60]. The loading of Drosophila cells with oleic acid provides another example of droplet movement [2]. In this paradigm, droplets initially proliferate, grow larger, and then move to grape-like clusters, which are often located near the nucleus. The basis for this movement of droplets is not understood. However, knockdowns of the Arf1/COPI machinery components prevent this clustering, and dynein appears to be required for droplet growth. Finally, regulated droplet movement was reported in axons of cultured giant cortical neurons of the sea slug Aplysia californica [61]. Droplets formed within axons, often in tandem arrays of 3–15 droplets, and moved unidirectionally in a retrograde manner toward the cell body. Since lipid droplet transport occurs in a variety of organisms, it may be a universal feature in the regulation of lipid-droplet biogenesis and utilization.
3.3. Fusion and fission
Lipid droplets not only interact dynamically with other organelles but also appear to undergo fusion and fission with other droplets. Reports suggest that droplet fusion is dependent on microtubules, the motor protein dynein, and the SNARE fusion machinery [38] and [62]. In an in vitro system, ERK2-mediated phosphorylation of dynein was implicated in droplet fusion [58] and [63]. Under specific conditions, up to 15% of droplets might be involved in fusion at any given time [62]. Fusion of lipid droplets was also observed in Drosophila cells with a knockdown of a phospholipid synthesis gene CCT1 [2] and in yeast lacking seipin [48].
Other data indicate that droplets undergo fission. The induction of lipolysis in 3T3-L1 adipocytes causes dramatic fission of lipid droplets [64]. Such fragmentation increases the surface area of droplets available for lipolysis and is predicted to require an increase in phospholipid availability to coat the surfaces of the droplets. Whether droplets participate in complex, regulated fusion and fission events, similar to those for mitochondria, remains to be determined. If so, the Arf1/COPI machinery may participate [2], as discussed above.
3.4. Lipid mobilization and utilization
The balance of lipids stored within lipid droplets is controlled by the net cycle of neutral lipid synthesis and degradation. Neutral lipids in the core of the droplet are hydrolyzed by intracellular lipases [65], yielding fatty acids for energy generation and anabolism of membrane phospholipids. The liberated fatty acids may be oxidized in mitochondria in mammalian cells or in peroxisomes in yeast [55] to generate ATP. Cycles of esterification and lipolysis are likely to be continuously active in smaller droplets of most cells. The PAT proteins on smaller droplets may afford some protection from lipolysis [66], but basal levels of lipolysis are usually detectable. On the other hand, droplets in adipose tissue or in the fat bodies of flies are major energy reserves for the organism. Therefore, the storage and utilization of lipids likely involve more complex regulation.
For hydrolysis of TG in lipid droplets of adipocytes, this regulation and the molecular processes of lipolysis are becoming clearer. Although lipolysis proceeds at basal levels, the rate can be dramatically stimulated by adrenergic hormones by activation of PKA. PKA phosphorylates perilipin and hormone-sensitive lipase (HSL) and causes a complex set of events leading to TG hydrolysis. Adipose tissue TG lipase first acts on TG to hydrolyze a fatty acyl chain [67]. Then HSL hydrolyzes a second acyl side chain of diacylglycerol. Finally, monoacylglycerol lipase hydrolyzes the last acyl side chain. The net result is that fatty acids and glycerol are liberated and released from the cell. During this process, significant portions of the diacylglycerol and monoacylglycerol intermediates are likely re-esterified by DGAT and MGAT enzymes, respectively [68].
How lipases access neutral lipids within droplets is unknown but presents a fascinating biophysical problem. The lipase must gain access to substrates by extending a portion of the protein through the phospholipid monolayer, or a microenvironment must be created that allows a neutral lipid to “pop up” through the monolayer to gain access to the lipase. The access of lipases to the droplet core is regulated in part by PAT proteins, and indeed, the phosphorylation of perilipin may be crucial for allowing HSL to gain access to its substrates [69]. Initial models suggested that PAT proteins cover the lipid droplet, thereby sterically hindering the access of lipases. However, these proteins are estimated to only cover roughly 15–20% of the droplet surface [70].
The hydrolysis and mobilization of sterol esters is less well understood. In Saccharomyces cerevisiae, sterol esters are hydrolyzed by Yehl, Yeh2, and Tgl1 [71], and TG is hydrolyzed by Tgl3, Tgl4, and Tgl5 [72]. These enzymes localize to the surfaces of lipid droplets, but little is known about how their localization and activities are regulated. In mammalian cells, cholesterol esters are hydrolyzed by HSL and neutral cholesterol esterases [65].
4. Renaissance organelles of many talents?
4.1. Assisted replication of intracellular pathogens
A novel function for lipid droplets emerged with the discovery that the core protein of the hepatitis C virus is associated with droplets [73] and [74]. Lipid droplets function in the assembly of the virus, and the efficiency of that process correlated with the ability of the core protein D2 to bind to lipid droplets [75]. These findings raise the intriguing possibility that treatments that diminish droplets might interfere with replication of the hepatitis C virus.
More recently, lipid droplets have been linked to the intracellular replication of Chlamydia trachomatis [76]. Intriguingly, the bacteria-containing vacuole where replication occurs appears to interact in a highly specific manner with lipid droplets, perhaps to provide lipids for replication of the microorganisms.
4.2. Safe haven for proteins?
The protein composition of lipid droplets suggests that they perform functions besides the storage of neutral lipids. Particularly surprising is the identification of histones as major lipid-droplet proteins in Drosophila embryos. High concentrations of these highly basic proteins were associated with lipid droplets of Drosophila oocytes [25]. This pool appears to serve an important storage function, since the histones are subsequently incorporated into chromatin during development, and this process requires large amounts of histones for the rapid cell divisions that occur during early embryogenesis. How these highly basic proteins are localized to and released from lipid droplets is not known.
In addition to histones, other proteins have unexpectedly been found as components of droplets. Prp19, a component of the NineTeen complex that has a canonical role in the activation of the spliceosome was found on lipid droplets, and its knockdown decreased lipid-droplet formation in cultured adipocytes [77]. Consistent with this finding, spliceosomal factors were prominent among genes involved in lipid-droplet formation that were identified by knockdown studies in Drosophila cells [2]. An important question is whether the splicing machinery participates directly in lipid-droplet formation or is indirectly involved, perhaps through regulated splicing of mRNAs encoding proteins required for droplet formation.
Finding diverse classes of proteins that interact with lipid droplets has led to the hypothesis that these organelles provide a depot for proteins, either to store them for future use or to sequester them until they can be degraded [21]. Intriguingly, in overexpression studies, α-synuclein increases the number of lipid droplets in yeast [78] and localizes to the droplet surface in mammalian cells [79], perhaps as a means of detoxifying the overexpressed protein.
5. Integration into the larger, multicellular world
5.1. Obesity and diseases of lipid storage
An understanding of the cell biology of lipid droplets is likely to have implications for other related fields of biology. Several human diseases are particularly relevant. The excessive accumulation of lipids in droplets is a hallmark of obesity, type 2 diabetes, hepatic steatosis, atherosclerosis, and other metabolic diseases that are prevalent worldwide. As pathways and functions of key molecules in lipid-droplet biology are identified, some will emerge as therapeutic targets. Promising examples include the DGAT enzymes [80] and [81], which catalyze triglyceride synthesis, and perilipin [82], the key structural protein in adipocytes. Also, as seen for hepatitis C virus and Chlamydia, some intracellular pathogens may require lipid droplets for replication, and elucidations of related mechanisms may suggest novel therapeutic strategies.
5.2. Oil production and biofuels
Lipid-droplet biology is also relevant to the industrial production of oils for food consumption and biofuels. In the agricultural industry, there is great interest in boosting oil production or altering the fatty acid composition of seed oils. An understanding of the molecular processes governing oil storage in lipid droplets promises novel approaches to engineering crops for this purpose. In fact, a polymorphism in DGAT was recently identified as the cause of a quantitative trait locus for controlling high levels of oil production in maize [83]. In this species, a single amino-acid substitution boosts oil content by up to 41% and oleic-acid content to 107%. DGAT polymorphisms are also determinants of milk-fat content [84] and [85], which originates through a poorly understood process in which lipid droplets are extruded from mammary epithelial cells into ducts of the mammary glands [86]. There is also great interest in maximizing oil production in unicellular organisms, such as yeast or algae, and plants for the generation of biofuels, including biodiesel. Like fossil-derived hydrocarbons, TG oils are highly concentrated stores of saturated hydrocarbons that can be oxidized for energy production. One can imagine using a toolbox of genetic manipulations to engineer these organisms to maximize cellular lipid storage and therefore oil production per cell.
6. An evolving biography
We see from this brief examination that the lives of lipid droplets are rich and complex. Mysteries remain concerning their birth, dynamic functions in cells, and connections with a multitude of intracellular processes. After languishing in relative obscurity for many years, lipid droplets have emerged as a fascinating and important field of study, and knowledge of their functions will undoubtedly grow rapidly in coming years. Future reviews will therefore be rich with new insights into the biology of lipid droplets and establish their rightful place as key cellular organelles.
Acknowledgments
We thank members of the Walther and Farese laboratories, Carole Sztalryd, and Dawn Brasaemle for their helpful discussions; Erin Currie, Yi Guo, Charlie Harris, Natalie Krahmer, Caroline Mrejen and Heather Newman for their comments on the manuscript; Daryl Jones for the manuscript preparation; and Gary Howard and Stephen Ordway for their editorial assistance. This work was supported by NIH grant R21-DK78254 (to R.F.), an International Human Frontier Science Program Organization career development award (to T.W.), the J. David Gladstone Institutes (R.F.), and the Max Planck Society (T.W.).
References
- 1.Sessa G, Weissmann G. Phospholipid spherules (liposomes) as a model for biological membranes. J Lipid Res. 1968;9:310–318. [PubMed] [Google Scholar]
- 2.Guo Y, Walther T, Rao M, Stuurman N, Goshima G, Terayama K, Wong J, Vale R, Walter P, Farese RV., Jr Functional genomic screen reveals genes involved in lipid droplet formation and utilization. Nature. 2008 May 29;453(7195):657–661. doi: 10.1038/nature06928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cohen A, Razani B, Schubert W, Williams T, Wang X, Iyengar P, Brasaemle D, Scherer P, Lisanti M. Role of caveolin-1 in the modulation of lipolysis and lipid droplet formation. Diabetes. 2004;53:1261–1270. doi: 10.2337/diabetes.53.5.1261. [DOI] [PubMed] [Google Scholar]
- 4.Vance J. Molecular and cell biology of phosphatidylserine and phosphatidylethanolamine metabolism. Prog Nucleic Acid Res Mol Biol. 2003;75:69–111. doi: 10.1016/s0079-6603(03)75003-x. [DOI] [PubMed] [Google Scholar]
- 5.Bartz R, Zehmer J, Zhu M, Chen Y, Serrero G, Zhao Y, Liu P. Dynamic activity of lipid droplets: protein phosphorylation and GTP-mediated protein translocation. J Proteome Res. 2007;6:3256–3265. doi: 10.1021/pr070158j. [DOI] [PubMed] [Google Scholar]
- 6.Bartz R, Li WH, Venables B, Zehmer JK, Roth MR, Welti R, Anderson RG, Liu P, Chapman KD. Lipidomics reveals that adiposomes store ether lipids and mediate phospholipid traffic. J Lipid Res. 2007;48:837–847. doi: 10.1194/jlr.M600413-JLR200. [DOI] [PubMed] [Google Scholar]
- 7.Tauchi-Sato K, Ozeki S, Houjou T, Tagushi R, Fujimoto T. The surface of lipid droplets is a phospholipid monolayer with a unique fatty acid composition. J Biol Chem. 2002;277:44507–44512. doi: 10.1074/jbc.M207712200. [DOI] [PubMed] [Google Scholar]
- 8.Parton R, Simons K. The multiple faces of caveolae. Nat Rev Mol Cell Biol. 2007;8:185–194. doi: 10.1038/nrm2122. [DOI] [PubMed] [Google Scholar]
- 9.Ostermeyer A, Paci J, Zeng Y, Lublin D, Munro S, Brown D. Accumulation of caveolin in the endoplasmic reticulum redirects the protein to lipid storage droplets. J Cell Biol. 2001;152:1071–1078. doi: 10.1083/jcb.152.5.1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pol A, Luetterforst R, Lindsay M, Heinoo S, Ikonen E, Parton R. A caveolin dominant negative mutant associates with lipid bodies and induces intracellular cholesterol imbalance. J Cell Biol. 2001;152:1057–1070. doi: 10.1083/jcb.152.5.1057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Fujimoto T, Kogo H, Ishiguro K, Tauchi K, Nomura R. Caveolin-2 is targeted to lipid droplets, a new “membrane domain” in the cell. J Cell Biol. 2001;152:1079–1085. doi: 10.1083/jcb.152.5.1079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Glenney J, Soppet D. Sequence and expression of caveolin, a protein component of caveolae plasma membrane domains phosphorylated on tyrosine in Rous sarcoma virus-transformed fibroblasts. Proc Natl Acad Sci U S A. 1992;89:10517–10521. doi: 10.1073/pnas.89.21.10517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ostermeyer A, Ramcharan L, Zeng Y, Lublin D, Brown D. Role of the hydrophobic domain in targeting caveolin-1 to lipid droplets. J Cell Biol. 2004;164:69–78. doi: 10.1083/jcb.200303037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kuerschner L, Moessinger C, Thiele C. Imaging of lipid biosynthesis: how a neutral lipid enters lipid droplets. Traffic. 2008;9:338–352. doi: 10.1111/j.1600-0854.2007.00689.x. [DOI] [PubMed] [Google Scholar]
- 15.Stone S, Levin M, Farese RV., Jr Membrane topology and identification of key functional amino acid residues of murine acyl-CoA:diacylglycerol acyltransferase-2. J Biol Chem. 2006;281:40273–40282. doi: 10.1074/jbc.M607986200. [DOI] [PubMed] [Google Scholar]
- 16.Londos C, Sztalryd C, Tansey J, Kimmel A. Role of PAT proteins in lipid metabolism. Biochimie. 2005;87:45–49. doi: 10.1016/j.biochi.2004.12.010. [DOI] [PubMed] [Google Scholar]
- 17.Hickenbottom S, Kimmel A, Londos C, Hurley J. Structure of a lipid droplet protein; the PAT family member TIP47. Structure. 2004;12:1199–1207. doi: 10.1016/j.str.2004.04.021. [DOI] [PubMed] [Google Scholar]
- 18.Wilson C, Wardell MR, Weisgraber KH, Mahley RW, Agard DA. Three-dimensional structure of the LDL receptor-binding domain of human apolipoprotein E. Science. 1991;252:1817–1822. doi: 10.1126/science.2063194. [DOI] [PubMed] [Google Scholar]
- 19.Lu B, Morrow JA, Weisgraber KH. Conformational reorganization of the four-helix bundle of human apolipoprotein E in binding to phospholipids. J Biol Chem. 2000;275:20775–20781. doi: 10.1074/jbc.M003508200. [DOI] [PubMed] [Google Scholar]
- 20.Wolins N, Quaynor B, Skinner J, Schoenfish M, Tzekov A, Bickel P. S3-12, adipophilin, and TIP47 package lipid in adipocytes. J Biol Chem. 2005;280:19146–19155. doi: 10.1074/jbc.M500978200. [DOI] [PubMed] [Google Scholar]
- 21.Welte M. Proteins under new management: lipid droplets deliver. Trends Cell Biol. 2007;17:363–369. doi: 10.1016/j.tcb.2007.06.004. [DOI] [PubMed] [Google Scholar]
- 22.Litvak V, Shaul Y, Shulewitz M, Amarilio R, Carmon S, Lev S. Targeting of Nir2 to lipid droplets is regulated by a specific threonine residue within its PI-transfer domain. Curr Biol. 2002;12:1513–1518. doi: 10.1016/s0960-9822(02)01107-7. [DOI] [PubMed] [Google Scholar]
- 23.Liu P, Ying Y, Zhao Y, Mundy DI, Zhu M, Anderson RG. Chinese hamster ovary K2 cell lipid droplets appear to be metabolic organelles involved in membrane traffic. J Biol Chem. 2004;279:3787–3792. doi: 10.1074/jbc.M311945200. [DOI] [PubMed] [Google Scholar]
- 24.Beller M, Riedel D, Jansch L, Dieterich G, Wehland J, Jackle H, Kuhnlein RP. Characterization of the Drosophila lipid droplet subproteome. Mol Cell Proteomics. 2006;5:1082–1094. doi: 10.1074/mcp.M600011-MCP200. [DOI] [PubMed] [Google Scholar]
- 25.Cermelli S, Guo Y, Gross S, Welte M. The lipid-droplet proteome reveals that droplets are a protein-storage depot. Curr Biol. 2006;16:1783–1795. doi: 10.1016/j.cub.2006.07.062. [DOI] [PubMed] [Google Scholar]
- 26.Wältermann M, Hinz A, Robenek H, Troyer D, Reichelt R, Malkus U, Galla H, Kalscheuer R, Stöveken T, Landenberg PV, Steinbüchel A. Mechanism of lipid-body formation in prokaryotes: how bacteria fatten up. Mol Microbiol. 2005;55:750–763. doi: 10.1111/j.1365-2958.2004.04441.x. [DOI] [PubMed] [Google Scholar]
- 27.Robenek H, Hofnagel O, Buers I, Robenek MJ, Troyer D, Severs NJ. Adipophilin-enriched domains in the ER membrane are sites of lipid droplet biogenesis. J Cell Sci. 2006;119:4215–4224. doi: 10.1242/jcs.03191. [DOI] [PubMed] [Google Scholar]
- 28.Buhman KK, Chen HC, Farese RV., Jr The enzymes of neutral lipid synthesis. J Biol Chem. 2001;276:40369–40372. doi: 10.1074/jbc.R100050200. [DOI] [PubMed] [Google Scholar]
- 29.Lardizabal KD, Mai JT, Wagner NW, Wyrick A, Voelker T, Hawkins DJ. DGAT2 is a new diacylglycerol acyltransferase gene family. Purification, cloning, and expression in insect cells of two polypeptides from Mortierella ramanniana with diacylglycerol acyltransferase activity. J Biol Chem. 2001;276:38862–38869. doi: 10.1074/jbc.M106168200. [DOI] [PubMed] [Google Scholar]
- 30.Cases S, Stone SJ, Zhou P, Yen E, Tow B, Lardizabal KD, Voelker T, Farese RV., Jr Cloning of DGAT2, a second mammalian diacylglycerol acyltransferase, and related family members. J Biol Chem. 2001;276:38870–38876. doi: 10.1074/jbc.M106219200. [DOI] [PubMed] [Google Scholar]
- 31.Sturmey R, O’Toole P, Leese H. Fluorescence resonance energy transfer analysis of mitochondrial:lipid association in the porcine oocyte. Reproduction. 2006;132:829–837. doi: 10.1530/REP-06-0073. [DOI] [PubMed] [Google Scholar]
- 32.Blanchette-Mackie E, Dwyer N, Barber T, Coxey R, Takeda T, Rondinone C, Theodorakis J, Greenberg A, Londos C. Perilipin is located on the surface layer of intracellular lipid droplets in adipocytes. J Lipid Res. 1995;36:1211–1226. [PubMed] [Google Scholar]
- 33.Goodman JM. The gregarious lipid droplet. J Biol Chem. 2008 Oct 17;283(42):28005–28009. doi: 10.1074/jbc.R800042200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ploegh H. A lipid-based model for the creation of an escape hatch from the endoplasmic reticulum. Nature. 2007;448:435–438. doi: 10.1038/nature06004. [DOI] [PubMed] [Google Scholar]
- 35.Robenek H, Robenek M, Troyer D. PAT family proteins pervade lipid droplet cores. J Lipid Res. 2005;46:1331–1338. doi: 10.1194/jlr.M400323-JLR200. [DOI] [PubMed] [Google Scholar]
- 36.Robenek MJ, Severs NJ, Schlattmann K, Plenz G, Zimmer KP, Troyer D, Robenek H. Lipids partition caveolin-1 from ER membranes into lipid droplets: Updating the model of lipid droplet biogenesis. Faseb J. 2004;18:866–868. doi: 10.1096/fj.03-0782fje. [DOI] [PubMed] [Google Scholar]
- 37.Nakamura N, Banno Y, Tamiya-Koizumi K. Arf1-dependent PLD1 is localized to oleic acid-induced lipid droplets in NIH3T3 cells. Biochem Biophys Res Commun. 2005;335:117–123. doi: 10.1016/j.bbrc.2005.07.050. [DOI] [PubMed] [Google Scholar]
- 38.Boström P, Andersson L, Rutberg M, Perman J, Lidberg U, Johansson B, Fernandez-Rodriguez J, Ericson J, Nilsson T, Borén J, Olofsson S. SNARE proteins mediate fusion between cytosolic lipid droplets and are implicated in insulin sensitivity. Nat Cell Biol. 2007;9:1286–1293. doi: 10.1038/ncb1648. [DOI] [PubMed] [Google Scholar]
- 39.Hofmann K. A superfamily of membrane-bound O-acyltransferases with implications for wnt signaling. Trends Biochem Sci. 2000;25:111–112. doi: 10.1016/s0968-0004(99)01539-x. [DOI] [PubMed] [Google Scholar]
- 40.Buhman KF, Accad M, Farese RV., Jr Mammalian acyl-CoA:cholesterol acyltransferases. Biochim Biophys Acta. 2000;1529:142–154. doi: 10.1016/s1388-1981(00)00144-x. [DOI] [PubMed] [Google Scholar]
- 41.Yang H, Bard M, Bruner DA, Gleeson A, Deckelbaum RJ, Aljinovic G, Pohl TM, Rothstein R, Sturley SL. Sterol esterification in yeast: a two-gene process. Science. 1996;272:1353–1356. doi: 10.1126/science.272.5266.1353. [DOI] [PubMed] [Google Scholar]
- 42.Sorger D, Daum G. Synthesis of triacylglycerols by the acyl-coenzyme A:diacyl-glycerol acyltransferase Dga1p in lipid particles of the yeast Saccharomyces cerevisiae. J Bacteriol. 2002;184:519–524. doi: 10.1128/JB.184.2.519-524.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Levine T, Loewen C. Inter-organelle membrane contact sites: through a glass, darkly. Curr Opin Cell Biol. 2006;18:371–378. doi: 10.1016/j.ceb.2006.06.011. [DOI] [PubMed] [Google Scholar]
- 44.Schulz T, Prinz W. Sterol transport in yeast and the oxysterol binding protein homologue (OSH) family. Biochim Biophys Acta. 2007;1771:769–780. doi: 10.1016/j.bbalip.2007.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hanada K, Kumagai K, Tomishige N, Kawano M. CERT and intracellular trafficking of ceramide. Biochim Biophys Acta. 2007;1771:644–653. doi: 10.1016/j.bbalip.2007.01.009. [DOI] [PubMed] [Google Scholar]
- 46.Brown R, Mattjus P. Glycolipid transfer proteins. Biochim Biophys Acta. 2007;1771:746–760. doi: 10.1016/j.bbalip.2007.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Szymanski K, Binns D, Bartz R, Grishin N, Li W, Agarwal A, Garg A, Anderson R, Goodman J. The lipodystrophy protein seipin is found at endoplasmic reticulum lipid droplet junctions and is important for droplet morphology. Proc Natl Acad Sci U S A. 2007;104:20890–20895. doi: 10.1073/pnas.0704154104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Fei W, Shui G, Gaeta B, Du X, Kuerschner L, Li P, Brown A, Wenk M, Parton R, Yang H. Fld1p, a functional homologue of human seipin, regulates the size of lipid droplets in yeast. J Cell Biol. 2008;180:473–482. doi: 10.1083/jcb.200711136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ost A, Ortegren U, Gustavsson J, Nystrom FH, Stralfors P. Triacylglycerol is synthesized in a specific subclass of caveolae in primary adipocytes. J Biol Chem. 2005;280:5–8. doi: 10.1074/jbc.C400429200. [DOI] [PubMed] [Google Scholar]
- 50.Brasaemle DL, Dolios G, Shapiro L, Wang R. Proteomic analysis of proteins associated with lipid droplets of basal and lipolytically stimulated 3T3-L1 adipocytes. J Biol Chem. 2004;279:46835–46842. doi: 10.1074/jbc.M409340200. [DOI] [PubMed] [Google Scholar]
- 51.Rajendran L, LeLay S, Illges H. Raft association and lipid droplet targeting of flotillins are independent of caveolin. Biol Chem. 2007;388:307–314. doi: 10.1515/BC.2007.034. [DOI] [PubMed] [Google Scholar]
- 52.Fernández M, Albor C, Ingelmo-Torres M, Nixon S, Ferguson C, Kurzchalia T, Tebar F, Enrich C, Parton R, Pol A. Caveolin-1 is essential for liver regeneration. Science. 2006;313:1628–1632. doi: 10.1126/science.1130773. [DOI] [PubMed] [Google Scholar]
- 53.Pohl J, Ring A, Stremmel W. Uptake of long-chain fatty acids in HepG2 cells involves caveolae: analysis of a novel pathway. J Lipid Res. 2002;43:1390–1399. doi: 10.1194/jlr.m100404-jlr200. [DOI] [PubMed] [Google Scholar]
- 54.Liu P, Bartz R, Zehmer J, Ying Y, Zhu M, Serrero G, Anderson R. Rab-regulated interaction of early endosomes with lipid droplets. Biochim Biophys Acta. 2007;1773:784–793. doi: 10.1016/j.bbamcr.2007.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Binns D, Januszewski T, Chen Y, Hill J, Markin V, Zhao Y, Gilpin C, Chapman K, Anderson R, Goodman J. An intimate collaboration between peroxisomes and lipid bodies. J Cell Biol. 2006;173:719–731. doi: 10.1083/jcb.200511125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hölttä-Vuori M, Tanhuanpää K, Möbius W, Somerharju P, Ikonen E. Modulation of cellular cholesterol transport and homeostasis by Rab11. Mol Biol Cell. 2002;13:3107–3122. doi: 10.1091/mbc.E02-01-0025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ozeki S, Cheng J, Tauchi-Sato K, Hatano N, Taniguchi H, Fujimoto T. Rab18 localizes to lipid droplets and induces their close apposition to the endoplasmic reticulum-derived membrane. J Cell Sci. 2005;118:2601–2611. doi: 10.1242/jcs.02401. [DOI] [PubMed] [Google Scholar]
- 58.Andersson L, Boström P, Ericson J, Rutberg M, Magnusson B, Marchesan D, Ruiz M, Asp L, Huang P, Frohman M, Borén J, Olofsson S. PLD1 and ERK2 regulate cytosolic lipid droplet formation. J Cell Sci. 2006;119:2246–2257. doi: 10.1242/jcs.02941. [DOI] [PubMed] [Google Scholar]
- 59.Guo Y, Jangi S, Welte M. Organelle-specific control of intracellular transport: distinctly targeted isoforms of the regulator Klar. Mol Biol Cell. 2005;16:1406–1416. doi: 10.1091/mbc.E04-10-0920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Welte MA, Cermelli S, Griner J, Viera A, Guo Y, Kim DH, Gindhart JG, Gross SP. Regulation of lipid-droplet transport by the perilipin homolog LSD2. Curr Biol. 2005;15:1266–1275. doi: 10.1016/j.cub.2005.06.062. [DOI] [PubMed] [Google Scholar]
- 61.Savage M, Goldberg D, Schacher S. Absolute specificity for retrograde fast axonal transport displayed by lipid droplets originating in the axon of an identified Aplysia neuron in vitro. Brain Res. 1987;406:215–223. doi: 10.1016/0006-8993(87)90785-2. [DOI] [PubMed] [Google Scholar]
- 62.Boström P, Rutberg M, Ericsson J, Holmdahl P, Andersson L, Frohman M, Borén J, Olofsson S. Cytosolic lipid droplets increase in size by microtubule-dependent complex formation. Arterioscler Thromb Vasc Biol. 2005;25:1945–1951. doi: 10.1161/01.ATV.0000179676.41064.d4. [DOI] [PubMed] [Google Scholar]
- 63.Marchesan D, Rutberg M, Andersson L, Asp L, Larsson T, Borén J, Johansson B, Olofsson S. A phospholipase D-dependent process forms lipid droplets containing caveolin, adipocyte differentiation-related protein, and vimentin in a cell-free system. J Biol Chem. 2003;278:27293–27300. doi: 10.1074/jbc.M301430200. [DOI] [PubMed] [Google Scholar]
- 64.Marcinkiewicz A, Gauthier D, Garcia A, Brasaemle DL. The phosphorylation of serine 492 of perilipin a directs lipid droplet fragmentation and dispersion. J Biol Chem. 2006;281:11901–11909. doi: 10.1074/jbc.M600171200. [DOI] [PubMed] [Google Scholar]
- 65.Zechner R, Strauss JG, Haemmerle G, Lass A, Zimmermann R. Lipolysis: pathway under construction. Curr Opin Lipidol. 2005;16:333–340. doi: 10.1097/01.mol.0000169354.20395.1c. [DOI] [PubMed] [Google Scholar]
- 66.Sztalryd C, Bell M, Lu X, Mertz P, Hickenbottom S, Chang B, Chan L, Kimmel A, Londos C. Functional compensation for adipose differentiation-related protein (ADFP) by Tip47 in an ADFP null embryonic cell line. J Biol Chem. 2006;281:34341–34348. doi: 10.1074/jbc.M602497200. [DOI] [PubMed] [Google Scholar]
- 67.Zimmermann R, Strauss JG, Haemmerle G, Schoiswohl G, Birner-Gruenberger R, Riederer M, Lass A, Neuberger G, Eisenhaber F, Hermetter A, Zechner R. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science. 2004;306:1383–1386. doi: 10.1126/science.1100747. [DOI] [PubMed] [Google Scholar]
- 68.Salter A, Wiggins D, Sessions V, Gibbons G. The intracellular triacylglycerol/fatty acid cycle: a comparison of its activity in hepatocytes which secrete exclusively apolipoprotein (apo) B100 very-low-density lipoprotein (VLDL) and in those which secrete predominantly apoB48 VLDL. Biochem J. 1998;332:667–672. doi: 10.1042/bj3320667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sztalryd C, Xu G, Dorward H, Tansey J, Contreras J, Kimmel A, Londos C. Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation. J Cell Biol. 2003;161:1093–1103. doi: 10.1083/jcb.200210169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Londos C, Brasaemle D, Schultz C, Segrest J, Kimmel A, Perilipins ADRP, and other proteins that associate with intracellular neutral lipid droplets in animal cells. Semin Cell Dev Biol. 1999;10:51–58. doi: 10.1006/scdb.1998.0275. [DOI] [PubMed] [Google Scholar]
- 71.Köffel R, Tiwari R, Falquet L, Schneiter R. The Saccharomyces cerevisiae YLL012/YEH1, YLR020/YEH2, and TGL1 genes encode a novel family of membrane-anchored lipases that are required for steryl ester hydrolysis. Mol Cell Biol. 2005;25:1655–1668. doi: 10.1128/MCB.25.5.1655-1668.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Daum G, Wagner A, Czabany T, Athenstaedt K. Dynamics of neutral lipid storage and mobilization in yeast. Biochimie. 2007;89:243–248. doi: 10.1016/j.biochi.2006.06.018. [DOI] [PubMed] [Google Scholar]
- 73.Hope R, McLauchlan J. Sequence motifs required for lipid droplet association and protein stability are unique to the hepatitis C virus core protein. J Gen Virol. 2000;81:1913–1925. doi: 10.1099/0022-1317-81-8-1913. [DOI] [PubMed] [Google Scholar]
- 74.Miyanari Y, Atsuzawa K, Usuda N, Watashi K, Hishiki T, Zayas M, Bartenschlager R, Wakita T, Hijikata M, Shimotohno K. The lipid droplet is an important organelle for hepatitis C virus production. Nat Cell Biol. 2007;9:1089–1097. doi: 10.1038/ncb1631. [DOI] [PubMed] [Google Scholar]
- 75.Shavinskaya A, Boulant S, Penin F, McLauchlan J, Bartenschlager R. The lipid droplet binding domain of hepatitis C virus core protein is a major determinant for efficient virus assembly. J Biol Chem. 2007;282:37158–37169. doi: 10.1074/jbc.M707329200. [DOI] [PubMed] [Google Scholar]
- 76.Kumar Y, Cocchiaro J, Valdivia RH. The obligate intracellular pathogen Chlamydia trachomatis targets host lipid droplets. Curr Biol. 2006;16:1646–1651. doi: 10.1016/j.cub.2006.06.060. [DOI] [PubMed] [Google Scholar]
- 77.Cho S, Shin E, Park P, Shin D, Chang H, Kim D, Lee H, Lee J, Kim S, Song M, Chang I, Lee O, Lee T. Identification of mouse Prp19p as a lipid droplet-associated protein and its possible involvement in the biogenesis of lipid droplets. J Biol Chem. 2007;282:2456–2465. doi: 10.1074/jbc.M608042200. [DOI] [PubMed] [Google Scholar]
- 78.Outeiro T, Lindquist S. Yeast cells provide insight into alpha-synuclein biology and pathobiology. Science. 2003;302:1772–1775. doi: 10.1126/science.1090439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Cole N, Murphy D, Grider T, Rueter S, Brasaemle D, Nussbaum R. Lipid droplet binding and oligomerization properties of the Parkinson’s disease protein alpha-synuclein. J Biol Chem. 2002;277:6344–6352. doi: 10.1074/jbc.M108414200. [DOI] [PubMed] [Google Scholar]
- 80.Matsuda D, Tomoda H. DGAT inhibitors for obesity. Curr Opin Investig Drugs. 2007;8:836–841. [PubMed] [Google Scholar]
- 81.Zhao G, Souers A, Voorbach M, Falls H, Droz B, Brodjian S, Lau Y, Iyengar R, Gao J, Judd A, Wagaw S, Ravn M, Engstrom K, Lynch J, Mulhern M, Freeman J, Dayton BD, Wang X, Grihalde N, Fry D, Beno D, Marsh K, Su Z, Diaz G, Collins C, Sham H, Reilly R, Brune M, Kym P. Validation of diacyl glycerolacyltransferase I as a novel target for the treatment of obesity and dyslipidemia using a potent and selective small molecule inhibitor. J Med Chem. 2008;51:380–383. doi: 10.1021/jm7013887. [DOI] [PubMed] [Google Scholar]
- 82.Tansey JT, Sztalryd C, Gruia-Gray J, Roush DL, Zee JV, Gavrilova O, Reitman ML, Deng CX, Li C, Kimmel AR, Londos C. Perilipin ablation results in a lean mouse with aberrant adipocyte lipolysis, enhanced leptin production, and resistance to diet-induced obesity. Proc Natl Acad Sci U S A. 2001;98:6494–6499. doi: 10.1073/pnas.101042998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Zheng P, Allen W, Roesler K, Williams M, Zhang S, Li J, Glassman K, Ranch J, Nubel D, Solawetz W, Bhattramakki D, Llaca V, Deschamps S, Zhong G, Tarczynski M, Shen B. A phenylalanine in DGAT is a key determinant of oil content and composition in mice. Nat Genet. 2008;40:367–372. doi: 10.1038/ng.85. [DOI] [PubMed] [Google Scholar]
- 84.Winter A, Kramer W, Werner FA, Kollers S, Kata S, Durstewitz G, Buitkamp J, Womack JE, Thaller G, Fries R. Association of a lysine-232/alanine polymorphism in a bovine gene encoding acyl-CoA:diacylglycerol acyltransferase (DGAT1) with variation at a quantitative trait locus for milk fat content. Proc Natl Acad Sci U S A. 2002;99:9300–9305. doi: 10.1073/pnas.142293799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Schennink A, Stoop W, Visker M, Heck J, Bovenhuis H, Poel Jvd, Valenberg Hv, Arendonk Jv. DGAT1 underlies large genetic variation in milk-fat composition of dairy cows. Anim Genet. 2007;38:467–473. doi: 10.1111/j.1365-2052.2007.01635.x. [DOI] [PubMed] [Google Scholar]
- 86.Heid H, Keenan T. Intracellular origin and secretion of milk fat globules. Eur J Cell Biol. 2005;84:245–258. doi: 10.1016/j.ejcb.2004.12.002. [DOI] [PubMed] [Google Scholar]