

Summary
Cancer cells exhibit deregulation in multiple cellular signaling pathways. Therefore, treatments using specific agents that target only one pathway usually fail in cancer therapy. The combination treatments using chemotherapeutic agents with distinct molecular mechanisms are considered more promising for higher efficacy; however, using multiple agents contributes to added toxicity. Emerging evidence has shown that some “natural products” such as isoflavones, indole-3-carbinol (I3C) and its in vivo dimeric product 3,3′-diindolylmethane (DIM), and curcumin among many others, have growth inhibitory and apoptosis inducing effects on human and animal cancer cells mediated by targeting multiple cellular signaling pathways in vitro without causing unwanted toxicity in normal cells. Therefore, these non-toxic “natural products” from natural resources could be useful in combination with conventional chemotherapeutic agents for the treatment of human malignancies with lower toxicity and higher efficacy. In fact, recently increasing evidence from pre-clinical in vivo studies and clinical trials have shown some success in support of the use of rational design of multi-targeted therapies for the treatment of cancers using conventional chemotherapeutic agents in combination with “natural products”. These studies have provided promising results and further opened-up newer avenues for cancer therapy. In this review article, we have succinctly summarized the known effects of “natural products” especially by focusing on isoflavones, indole-3-carbinol (I3C) and its in vivo dimeric product 3,3′-diindolylmethane (DIM), and curcumin, and provided a comprehensive view on the molecular mechanisms underlying the principle of cancer therapy using combination of “natural products” with conventional therapeutics.
Keywords: cancer therapy, natural products, cell signaling
Introduction
Despite enormous efforts made toward the development of cancer therapies over the past several decades, cancer is still a major public health problem in the United States, and accounting for one in 4 deaths in the United States.1 Chemotherapy is one of the important approaches among cancer therapies. Physicians and scientists have vigorously investigated ways to improve the efficacy of chemotherapy by developing new chemotherapeutic agents, refining therapeutic approaches, and optimizing combination treatment strategies. It is now well accepted that cancer cells are highly heterogeneous and they exhibit deregulation in multiple cellular signaling pathways. Therefore, treatments using specific agents or inhibitors that target only one biological event or a single pathway usually fail in cancer therapy. In order to overcome this failure and to achieve higher efficacy, combination treatments using multiple agents with distinct targets have been considered more promising, resulting in better cancer cell killing. Therefore the premise is that by attacking or blocking multiple important pathways that are responsible for promoting cancer cell survival and growth, and thus multi-targeted cancer therapy is likely to be more effective in inhibiting cancer cell growth and inducing apoptotic cell death. However, the combination treatment using conventional chemotherapeutic agents has been shown to be associated with unacceptable dose-related toxicity; therefore, the development of mechanism-based multi-targeted therapeutic strategies in the new era of targeted therapy is urgently needed for improving therapeutic efficacy without added systemic toxicity during the combination treatment. We believe that such a strategy of rational design is likely to be a breakthrough in the treatment of the majority of human malignancies.
In recent years, dietary and natural compounds harvested from nature (natural compounds) have received increased attention primarily because epidemiological studies have shown that the consumption of fruits, soybean and vegetables is associated with reduced risk of several types of cancers.2,3 Emerging evidence has also shown that some “natural products” especially isoflavones, indole-3-carbinol (I3C) and its in vivo dimeric product 3,3′-diindolylmethane (DIM), and curcumin have potent growth inhibitory and apoptosis inducing effects on human and animal cancer cells mediated by targeting multiple cellular signaling pathways in vitro. 4,5 Therefore, these non-toxic “natural products” could be useful in combination with conventional chemotherapeutic agents for the treatment of cancer and such strategies are expected to have lower toxicity but higher efficacy. Recent in vivo pre-clinical studies and clinical trials have provided increasing evidence in support of the development of multi-targeted therapies for cancer treatment using chemotherapeutic agents in combination with “natural products”.6-13 These studies have provided some early promising results and opened-up newer avenues for cancer therapy. In this review article, we have attempted to summarize the known effects of selected “natural products” and provided a comprehensive view on the molecular mechanisms and underlying rationale for the combination of conventional therapeutics with “natural products” for the treatment of human malignancies.
Soy isoflavones including genistein, daidzein, and glycitein are mainly derived from soybean, and genistein has been found to inhibit cancer cell growth in vivo and in vitro.14-16 Whereas I3C and its in vivo dimeric product DIM are produced from naturally occurring glucosinolates found in the family Cruciferae have shown inhibitory effects on cancer cell growth through the modulation of genes that are related to the control of cell proliferation, cell cycle, apoptosis, signal transduction, oncogenesis, and transcription regulation.17,18 Moreover, curcumin is also a natural compound present in turmeric and has been known to possess both anti-inflammatory and antioxidant effects and it has also been studied as a cancer chemopreventive agent in several cancer models.19,20 Thus, it is becoming clear that these “natural products” exerts their pleiotropic effects on cancer cells through targeting multiple cellular signaling pathways including NF-κB, Akt, MAPK, Wnt, Notch, and androgen receptor (AR), suggesting that these “natural products” could be useful either alone or in combination with conventional therapeutics for the prevention of tumor progression and/or treatment of human malignancies.
The roles of isoflavone in cancer therapy
Targeting multiple pathways by Isoflavone
There are growing in vitro and in vivo evidence demonstrating the inhibitory effects of isoflavone genistein on carcinogenesis and cancer progression. These inhibitory effects have been believed to be mediated by multiple signaling pathways, among which NF-κB appears to be most important, which is well known for its central function in promoting human malignancies. We found that isoflavone genistein significantly inhibited the DNA-binding activity of NF-κB in prostate cancer cells 21, suggesting that the inactivation of NF-κB is mechanistically linked with growth inhibitory and apoptosis promoting activity of isoflavone. By immunochemistry and confocal microscopic analysis, we also found that isoflavone genistein inhibited the nuclear translocation of NF-κB 21, suggesting that genistein may reduce the DNA binding activity of NF-κB to its target DNA and thereby inhibit the transcription of its target genes. Similar findings have been reported by other investigators showing that isoflavone could inhibit NF-κB through modulation of IKK and IκB in human lung epithelial cells and myeloid cells 22,23, suggesting that soy isoflavone could exert its cancer chemopreventive activity through the regulation of NF-κB signaling. More importantly, we have found that isoflavone genistein could enhance the anti-tumor activity of chemotherapeutic agents not only via the down-regulation of NF-κB signaling but by also attenuating the activation of NF-κB induced by chemotherapeutic agents.24 These results clearly suggest that isoflavones either alone or with rational combination with conventional therapeutics could be useful for the treatment of human malignancies as schematically demonstrated in Fig 1.
Figure 1.
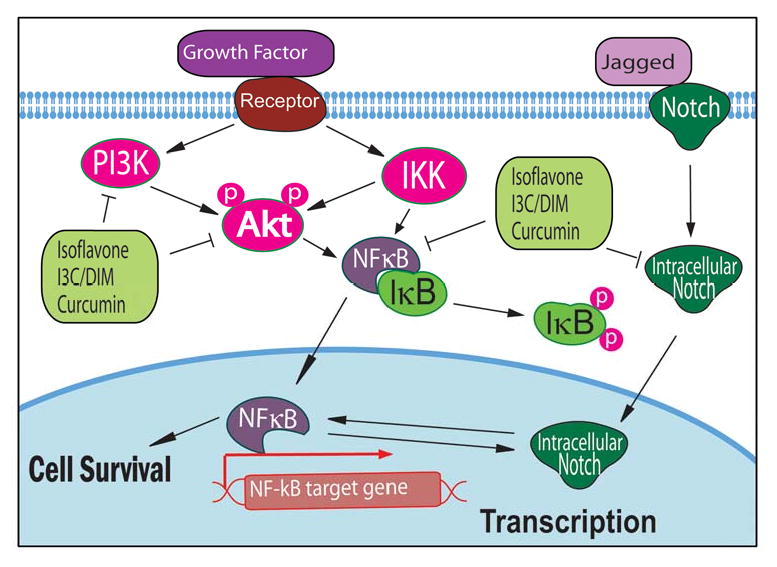
NF-κB, Akt, and Notch signaling pathways altered by isoflavone, I3C, DIM, and curcumin.
Isoflavone genistein also regulates Akt signaling which is another important signaling pathway that is known to be responsible for cancer development and progression. We found that genistein did not alter the level of total Akt protein; however, the phosphorylated Akt protein at Ser473 and the Akt kinase activity were decreased after genistein treatment.25 To further explore the inhibitory mechanism of genistein on the activity of Akt and NF-κB pathways, we have conducted transfection experiments in PC-3 cells using Akt expression vector and NF-κB-Luc reporter construct. Luciferase assay showed an increased luciferase activity in PC-3 cells co-transfected with the constructs. However, genistein inhibited the luciferase activity in co-transfected PC-3 cells. These results were further confirmed by examining the DNA-binding activity of NF-κB in transfected cells using EMSA, and the results clearly suggested that genistein exerts its inhibitory effects on NF-κB pathway mediated in part by targeting Akt signaling pathway. We have also observed similar results in MDA-MB-231 breast cancer cells;26 therefore, isoflavone genistein could down-regulate both Akt and NF-κB signaling, leading to the inhibition of cancer cell growth and the induction of apoptosis (Fig 1). It is also important to note that EGFR could activate PI3K/Akt signaling through induction of Src and that isoflavone is a Src kinase inhibitor.27 Therefore, the inhibition of Akt activity by isoflavone could also be via the down-regulation of Src signaling.
Recently, we have investigated the effects of isoflavone on Wnt signaling. We found that isoflavone up-regulated the expression of GSK-3β, enhanced GSK-3β binding to β-catenin, and increased the phosphorylation of β-catenin, suggesting that isoflavone could inhibit prostate cancer cell growth by the inactivation of Wnt signaling (Fig 2).28 Other investigators have also reported that isoflavone could inhibit the expression of Wnt-5a,29 diminish basal and Wnt-1-induced cell proliferation, and attenuate Wnt-1 targeted c-Myc and Cyclin D1 expressions,30 suggesting the inhibitory effects of isoflavone on Wnt signaling.
Figure 2.
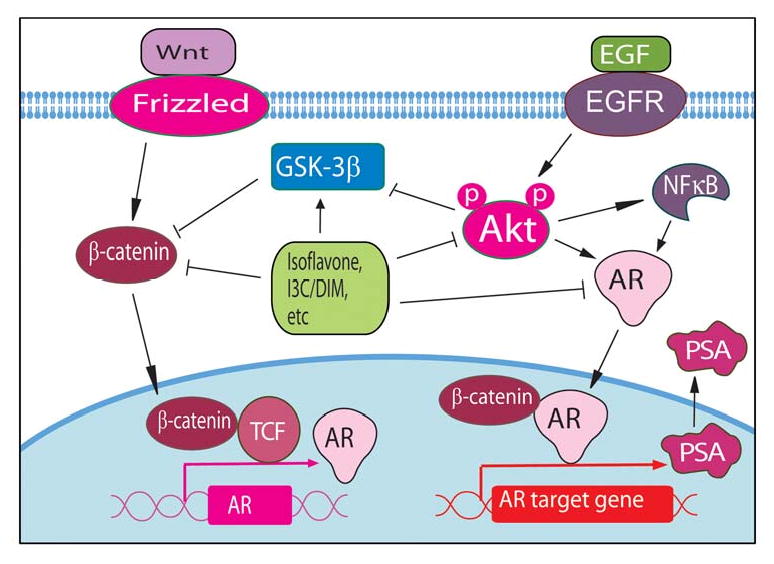
Wnt and AR signaling pathways altered by natural products such as isoflavone, I3C, DIM, etc.
Isoflavone genistein also showed inhibitory effects on Notch signaling, resulting in the inhibition of cancer cell growth. We have found that isoflavone genistein inhibited Notch signaling, leading to the down-regulation of NF-κB activity consistent with the inhibition of cell proliferation, and the induction of apoptosis in pancreatic cancer cells.31,32 Su et al also reported that isoflavone genistein could suppress the expression of Notch-2,29 which is consistent with our findings. These results suggest that isoflavone genistein could inhibit cancer cell growth and induce apoptosis through the down-regulation of Notch and NF-κB signaling (Fig 1).
In an effort to find the effects of isoflavone genistein on androgen receptor (AR) signaling especially because targeting of AR could be useful strategy for the treatment of prostate cancer. We found that genistein transcriptionally down-regulated AR, decreased nuclear AR binding to androgen responsive element (ARE) and, thereby, inhibited the transcription and translation of PSA in androgen-sensitive LNCaP cells 33,34 as schematically summarized in Fig-2. Fritz et al also found that isoflavone genistein down-regulated the expression of AR in the rat prostate at concentrations comparable to those found in humans on a soy diet.35 Moreover, we have found that isoflavone could inhibit cancer cell proliferation and induce apoptosis through the regulation of the Akt/FOXO3a/GSK-3β/AR signaling network.28 The above molecular mechanistic studies from our laboratory and others have clearly suggested that isoflavone regulates multiple cellular signaling pathways in vitro, suggesting that isoflavone could also suppress tumor growth in vivo, which could be due to attenuation of these signaling pathways.
Effects of isoflavone on the inhibition of tumor growth in animal
It has been reported that isoflavone has protective roles against carcinogenesis in animals. In an early study, soy-containing diets has been shown to reduce the severity of prostatitis in rats.36 The diets supplemented with soy isoflavone also prevented the development of adenocarcinomas in the prostate and seminal vesicles in a rat carcinogenesis model.37 Moreover, soy diet reduced growth of transplantable prostate adenocarcinomas and inhibited tumor cell proliferation and angiogenesis of transplantable prostate cancer in immunodeficient mice.38,39 A diet rich in soy also inhibited pulmonary metastasis of melanoma cells in C57Bl/6 mice.40 Furthermore, isoflavone genistein also inhibited the growth of carcinogen-induced cancers in rats and human leukemia cells transplanted into mice.41,42 These in vivo studies provided strong evidence in support of the role of isoflavone genistein in the protection against carcinogenesis in animal models. Importantly, increasing number of human studies and clinical trials has also been conducted in recent years toward molecular investigation and anti-tumor activity of isoflavone on human cancers as summarized in this article.
Effects of isoflavone on the suppression of oxidant stress in human through the inhibition of NF-κB activation
Isoflavone genistein is a well known antioxidant 43 and the treatment of cancer cells with genistein could significantly increase the expression of anti-oxidant proteins,44 boosting the endogenous anti-oxidant pathways. Therefore, soy isoflavone supplementation is expected to inhibit NF-κB activation and, in turn, may reduce the oxidative DNA damage in human cells. We have conducted a human intervention study to investigate the effects of isoflavone supplementation on NF-κB activation in vivo in human volunteers.45 The lymphocytes from healthy male subjects were harvested from peripheral blood and cultured for 24 hours in the absence and presence of isoflavone genistein that was previously shown to protect cells from TNF-α or H2O2 induced NF-κB activation in vitro. We found that isoflavone genistein treatment inhibited basal levels of NF-κB DNA binding activity by 56% and abrogated TNF-α induced NF-κB activity by 50%.45 Furthermore, when human volunteers received isoflavone supplements for three weeks, TNF-α failed to activate NF-κB activity ex-vivo in lymphocytes harvested from these volunteers, while lymphocytes from these volunteers collected prior to soy isoflavone intervention showed activation of NF-κB DNA binding activity upon TNF-α treatment ex-vivo.45 We have further determined whether soy supplementation had any effect on the levels of oxidative DNA damage in human lymphocytes by measuring the levels of 5-OHmdU, which represents oxidative DNA damage, in the blood of the volunteers before and after isoflavone supplementation. The mean value of 5-OHmdU before supplementation was 156.7±25.72 and it was decreased to 60.83±12.61 (p < 0.01) after three weeks of soy supplementation.45 These results clearly demonstrate that soy isoflavone supplementation has a protective effect against NF-κB activation and oxidative DNA damage in humans in vivo.
Effects of isoflavone on the inhibition of serum PSA in patients with prostate cancer
In an in vitro study, we found that isoflavone genistein transcriptionally down-regulated AR and decreased nuclear AR binding to ARE, leading to the inhibition of PSA expression in prostate cancer cells 34 as indicated above. Because PSA is an important biomarker for the diagnosis of prostate cancer and the prediction of prostate cancer progression in humans, we conducted a Phase II clinical trial to investigate the modulation in serum PSA levels in patients diagnosed with prostate cancer by soy isoflavone supplementation. Patients with prostate cancer were enrolled if they had rising PSA levels and were previously untreated (Group I), treated with local therapy (Group II), or treated with hormone therapy (Group III), and had either three successive rising PSA levels or a PSA of > 10 ng/ml at two successive evaluation. Patients received 100 mg Novasoy™ (Archer Daniels midland Company, Decatur, IL, USA) orally twice daily for a minimum of three months in the absence of progression or toxicity. Novasoy™ contains genistein, daidzein, and glycitin at a 1.3:1:0.3 ratios. Serum PSA, IGF-1 and IGFBP-3 levels were measured and toxicity was assessed. There was a decrease in the rate of rise of serum PSA in the whole group (P = 0.01) with rate of rise in PSA was decreased from 14 to 6% in group II (P = 0.21) and from 31 to 9% in group III (P = 0.05) following the soy isoflavone intervention.46 This pilot clinical data demonstrated that soy isoflavone supplementation could decrease the rate of rise in serum PSA levels without any toxicity in prostate cancer patients. The lack of significant side effects of soy isoflavone makes it an ideal agent for patients with advanced prostate cancer, suggesting that further definitive clinical trials are warranted.
Another phase II clinical trial has been conducted to evaluate the efficacy of isoflavone in patients with PSA recurrent prostate cancer after prior therapy.47 Patients received soy milk containing 47 mg of isoflavonoid per 8 oz serving three times per day for 12 months. Serum PSA, testosterone, lipids, and isoflavone levels (genistein, daidzein, and equol) were measured at various time points from 0 to 12 months and PSA outcome was also evaluated. It was found that dietary intervention with isoflavone supplementation decreased the slope of rising PSA, providing beneficial evidence in support of the use of isoflavone supplements against prostate cancer.47 In another clinical trial focusing on determining the biological effects of soy protein isolate (SPI) consumption on circulating hormone profiles and AR expression patterns in men at high risk for developing advanced prostate cancer, the authors used prostate biopsy samples obtained pre- and post-intervention and subjected to immunostaining for assessing the expression of AR and ER-β. The authors have found that consumption of SPI significantly suppressed AR expression but did not alter ER-β expression or circulating hormones, suggesting that isoflavone could be beneficial in preventing prostate cancer by inhibition of AR and PSA expression 48 and these results from clinical studies are consistent with in vitro findings.
In a prospective follow-up study, the effects of a dietary supplementation including isoflavone have been tested in men with isolated high-grade prostatic intraepithelial neoplasia (HGPIN). It has been found that 67.6% of patient showed a stable or decreasing PSA level and had a lower risk (25.5%) of prostate cancer after isoflavone supplementation, suggesting that the supplements could decrease the level of PSA and thereby decreasing the risk of prostate cancer.49 Several other clinical trials have also shown similar results documenting reduced PSA after receiving isoflavone supplements;50,51 however, another clinical trial showed that isoflavone did not alter the level of PSA in healthy men aged 50-80 years,52 suggesting that differential effects of isoflavone may exist between healthy men and patients diagnosed with prostate cancer. Collectively, evidence from both in vitro as well as in vivo clinical trials clearly suggest that soy isoflavone has biological activity against prostate cancer without any known systemic toxicity; therefore in our humble opinion further definitive studies are warranted in order to fully appreciate the chemopreventive and therapeutic effects of soy isoflavone for human prostate cancer.
The effects of isoflavone on breast and other cancers
Since it is known that low concentration of genistein could stimulate the growth of ER-positive MCF-7 breast cancer cells,53 there is a concern whether isoflavones will increase breast cancer risk in healthy women or worsen the prognosis of breast cancer patients although there is no clinical evidence whether isoflavone intake could increase breast tissue density in pre- or postmenopausal women or increase breast cell proliferation in postmenopausal women with or without a history of breast cancer, the known risk factors for the development of breast cancer, and such clinical studies have shown no adverse effects although it was tested in a small number of subjects.54 Moreover, the epidemiological data are consistent with the clinical data, showing no indication of increased risk of breast cancer in the populations consumes isoflavone-rich diets. In a large epidemiological study in women in China, the plasma genistein and daidzein concentrations have been measured and correlated with the risk of breast disease and the authors have found that isoflavone concentrations were inversely associated with the risk of non-proliferative and proliferative fibrocystic conditions and breast cancer.55 Women in the highest quartile of plasma genistein (>76.95 ng/mL) were less likely to have breast cancer or benign fibrocystic conditions compared with women in the lowest quartile (<9.42 ng/mL), suggesting the inhibitory effects of isoflavone on mammary carcinogenesis 55. In another study of 117 case-control pairs of postmenopausal women, the inverse associations between urinary phytoestrogens and breast cancer risk were also found especially among women with a high body mass index.56 Other clinical trials also showed the beneficial effects of isoflavone on breast cancer,57,58 suggesting that the consumption of soy-rich diet could be useful for the prevention of breast cancer although longitudinal studies in a large-population would be required to authenticate the above findings so that the NCI could issue a public health release in support of the role of soy isoflavone in breast cancer prevention.
In addition to breast cancer, a recent study has found that high intake of flavonols and isoflavonoid genistein was associated with decreased risk of advanced colorectal adenoma recurrence, suggesting that a isoflavone-rich diet could also decrease the risk of advanced colorectal adenoma recurrence.59 However, another clinical study on colorectal neoplasia showed that supplementation with soy protein containing isoflavones did not reduce colorectal epithelial cell proliferation or the average height of proliferating cells in the cecum, sigmoid colon, and rectum,60 suggesting that further clinical trials are needed in this area.
New or ongoing clinical trials using isoflavone
Based on the molecular evidence showing that isoflavone targets multiple cellular signaling pathways, more and more clinical trials are being conducted to investigate the value of isoflavone in human cancer (Table 1). These clinical trials are focused on investigating the effects of isoflavone on the prevention of various cancer developments and the combination treatment of various cancers with chemotherapeutic agents or other “natural products”. We believe that the conventional cancer therapeutics combined with isoflavone supplement could be an important novel strategy for the treatment of human cancers and/or the prevention of cancer progression; therefore, further rationally-designed clinical trials are needed in order to fully appreciate the health benefit of soy isoflavones in humans. In support of our belief, we have recently shown that soy isoflavone in conjunction with radiation therapy could reduce urinary, gastrointestinal and sexual adverse effects observed in patients receiving external beam radiation therapy for prostate cancer,61 suggesting the beneficial effects of isoflavone on combination treatment in cancer therapy.
Table 1.
The new or ongoing clinical trials to investigate the value of isoflavone in cancer clinic*.
| NCT ID | Title |
|---|---|
| NCT00617617 | The specific role of isoflavones in reducing prostate cancer risk |
| NCT00204490 | Soy isoflavones and breast cancer risk reduction |
| NCT00243048 | Isoflavones and radiation therapy in treating patients with localized prostate cancer |
| NCT00078923 | Soy isoflavones in treating patients who are undergoing radical prostatectomy for stage I or stage II PCa |
| NCT00200824 | Effects of soy compounds on breast cancer, prostate cancer, and bone health |
| NCT00003100 | Dietary soy isoflavones for the prevention of cancer |
| NCT00354432 | Soy protein/isoflavones and venlafaxine in treating hot flashes in patients receiving hormone therapy for prostate cancer |
| NCT00255125 | Role of soy supplementation in prostate cancer development |
| NCT00513916 | Effect of dietary soy on estrogens in breast fluid, blood, and urine samples from healthy women |
| NCT00499408 | Vitamin d and soy supplements in treating patients with recurrent prostate cancer |
| NCT00555386 | Soy, selenium and breast cancer risk |
| NCT00118040 | Genistein in patients who are undergoing surgery for bladder cancer |
| NCT00345813 | Soy supplements in treating patients undergoing surgery for localized prostate cancer |
| NCT00765479 | Soy protein in preventing recurrent cancer in patients who have undergone surgery for stage II prostate cancer |
| NCT00290758 | Genistein in preventing breast cancer in women at high risk for breast cancer |
| NCT00701584 | The role of diet and lifestyle in breast cancer survival |
| NCT00244933 | Gemcitabine hydrochloride and genistein in treating women with stage iv breast cancer |
| NCT00005827 | Genistein in treating patients with stage ii, stage iii, or stage iv prostate cancer |
| NCT00118040 | Genistein in patients who are undergoing surgery for bladder cancer |
| NCT00276835 | Genistein and interleukin-2 in treating patients with metastatic melanoma or kidney cancer |
| NCT00546039 | Synthetic genistein (bonistein™) in patients who are undergoing surgery for prostate cancer |
| NCT00290758 | Genistein in preventing breast cancer in women at high risk for breast cancer |
| NCT00376948 | Genistein, gemcitabine, and erlotinib in treating patients with locally advanced or metastatic pancreatic cancer |
| NCT00499408 | Vitamin D and soy supplements in treating patients with recurrent |
Information collected from ClinicalTrials.gov
The roles of I3C/DIM in cancer therapy
I3C is produced from naturally occurring glucosinolates contained in a wide variety of plants including members of the family Cruciferae. In the family Cruciferae, vegetables of the genus Brassica contribute most to our intake of glucosinolates found in all kinds of cabbages, broccoli, cauliflower, and Brussels sprouts. I3C is biologically active and it is the immediate precursor of DIM. Under the acidic conditions of the stomach, I3C undergoes extensive and rapid self-condensation reactions to form several derivatives.62 DIM is the major derivative and condensation product of I3C and its formation from I3C has been believed to be a likely prerequisite for I3C-induced anti-carcinogenesis.63
Targeting multiple pathways by I3C and DIM
Both I3C and DIM have been found to regulate NF-κB signaling in various cancer cells in vitro. We have reported that I3C significantly inhibited NF-κB DNA binding activity with the induction of apoptosis in PC-3 prostate cancer cells.17 We have also found that DIM could inhibit NF-κB DNA binding activity in androgen sensitive LNCaP and androgen insensitive PC-3 and C4-2B prostate cancer cells.64,65 These results suggest that inhibition of NF-κB signaling pathway may be one of the molecular mechanisms by which I3C and DIM induce apoptosis and inhibit prostate cancer cell growth (Fig 1).
Both I3C and DIM also regulate Akt signaling transduction and inhibit the activation of Akt in various cancer cells. We have found that the phosphorylated Akt protein was decreased in I3C or DIM treated prostate cancer cells,65,66 suggesting their inhibitory effects on Akt activation. Akt kinase activity in I3C or DIM treated prostate cancer cells was also down-regulated, suggesting the inactivation of Akt after I3C or DIM treatment. From the gene expression profiles of PC-3 cells exposed to I3C, we found down-regulation of PI3K expression, which is consistent with our results showing inactivation of Akt kinase activity by I3C.18 In summary, these results clearly suggest that I3C and DIM inhibits Akt signaling pathway, which may contribute to the inhibition of cell proliferation and the induction of apoptotic cell death (Fig 1).
To further explore the comprehensive molecular effects of I3C and DIM, we have conducted microarray analysis to determine the alternation of gene expression profiles of PC-3 prostate cancer cells exposed to I3C or DIM.18 We found that I3C and DIM treatments down-regulated the expression of MAP2K3, MAP2K4, MAP4K3, and MAPK3, suggesting the inhibitory effects of I3C and DIM on MAPK pathway. Other investigators also reported similar results showing that the effects of DIM were mediated by cross-talk between the protein kinase A and MAPK signaling pathways.67 Therefore, the down-regulation of the important molecules in MAPK pathway by I3C and DIM could be mechanistically responsible for the inhibition of cancer cell survival and growth.
It has been well known that there is a cross-talk between Akt and Wnt signaling pathways through the signal transduction between GSK-3β and β-catenin, two of the important molecules in Akt and Wnt pathways as discussed previously with respect to the biological effects of isoflavone. Since both I3C and DIM inhibits the activation of Akt as described above, it is obviously the DIM that inhibits Wnt activation through the signal transduction between Akt and Wnt signaling pathway because DIM is the in vivo dimeric and biologically active product of I3C. In prostate cancer cells, we have found that DIM significantly increased the phosphorylation of β-catenin and inhibited β-catenin nuclear translocation,68 suggesting that DIM could suppress the activation of Wnt signaling and inhibit the growth of prostate cancer cells (Fig 2).
It has been known that DIM could inhibit the AR nuclear translocation, the PSA expression, and the cell proliferation induced by dihydrotestosterone (DHT) in LNCaP cells.69 We have also found that DIM significantly inhibited Akt activation, NF-κB DNA binding activity, AR phosphorylation, AR nuclear translocation, and the expressions of AR and PSA, suggesting that DIM could inhibit the signal transduction between Akt/NF-κB and AR.64 These results collectively demonstrated that DIM-induced inhibition of cell proliferation and induction of apoptosis are partly mediated through the down-regulation of AR, Akt, and NF-κB signaling. Therefore, we have further investigated the molecular effects of DIM on Akt and AR signaling and we found that DIM significantly inhibited the phosphorylation of Akt and FOXO3a, suppress FOXO3a binding to the promoter of AR, and enhanced FOXO3a binding to the p27KIP1 promoter, resulting in the alteration of AR and p27KIP1 expression, the inhibition of cell proliferation, and the induction of apoptosis in both androgen-sensitive and -insensitive prostate cancer cells.68 These results clearly demonstrated that DIM-induced inhibition of cell proliferation and induction of apoptosis are partly mediated through the regulation of Akt/FOXO3a/AR signaling (Fig 2), and thus we believe that DIM could be useful for the treatment of both androgen-sensitive but most importantly castrate-resistant prostate cancer for which there is no curative therapy.
Inhibition of tumor growth in animal by I3C and DIM
I3C has been shown to inhibit chemically induced tumorigenesis of the liver, mammary gland, colon, and other organs, and suppress spontaneous carcinogenesis of mammary gland and endometrium.62,70,71 It has been reported that treatment of rat with I3C increases CYP1A1, 1A2, 2B, and 3A activity in rat liver.72,73 I3C also up-regulates the level and activity of glutathione S-transferases (GST).72,74 Therefore, the anti-oncogenic activity of I3C administered before or concurrently with a carcinogen is thought to be mediated through alternations in the levels and activities of Phase I (e.g. p450 or CYP) and phase II (e.g. GST) isozymes in the liver and/or extrahepatic tissues, resulting in their increased capacity for detoxification of carcinogens. Recent report showed that I3C exhibited inhibitory and preventive effects on prostate tumors in mice with the induction of apoptosis.75 I3C also inhibited 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone plus benzo(a)pyrene-induced lung tumorigenesis in A/J mice and modulated carcinogen-induced alterations in protein levels;76 however another study has shown tumor promoting activity of I3C,77 suggesting that I3C could have harmful effects in selected tumor model. It is important to note however that DIM do not show any genotoxic adverse effects which is typically seen by I3C,78 suggesting that future clinical trials should be conducted with DIM but not I3C.
It has been reported that DIM inhibited angiogenesis and the growth of transplantable human breast carcinoma in athymic mice with the upregulation of p27Kip1 expression and the down-regulation in the expression of CDK2 and CDK6.79 We have also found that DIM could inhibit prostate cancer cell growth in mice through the down-regulation of uPA, MMP-9, mTOR, PDGF-D, and angiogenesis.80,81 Moreover, we found that DIM could potentiate the anti-tumor activity of erlotinib, leading to a significant inhibition in pancreatic cancer cell growth in vitro and tumor growth in experimental model in vivo in SCID mice.82 Other investigators also reported that DIM significantly inhibited TRAMP-C2 mouse prostate cancer cells growing in C57BL/6 mice with increased apoptosis and decreased cell proliferation compared to controls.83 These evidences all suggest that I3C and DIM could inhibit tumor growth in experimental animals in vivo, suggesting that DIM could be an useful agent for the inhibition of tumor growth in humans.
Clinical trials for cancer therapy using I3C or DIM
Several clinical trials have been conducted to investigate the tolerability and effects of I3C and DIM in human. In a phase I trial of I3C, 17 women from a high-risk breast cancer cohort have been involved. All subjects tolerated chronic doses of 400 mg I3C daily for 4 weeks followed by a 4-week period of 800 mg I3C daily,84 suggesting the safety of I3C at these doses in human. It was found that 400 mg I3C daily elicited a maximal protective effect if the ratio of hydroxylated estrone metabolites were considered as a biomarkers for chemoprevention.84 A phase II clinical trial has been conducted to determine the potential therapeutic benefits of I3C in the management of vulvar intraepithelial neoplasia (VIN). It has been found that there was a significant improvement in symptomatology and vulvoscopic appearance of VIN with I3C therapy.13 In a clinical study using I3C for the treatment of recurrent respiratory papillomatosis (RRP), 33% of patients experienced remission of papillomatous growth and did not require surgery while on I3C intervention and 30% of patients had a reduction in papillomatous growth that resulted in less frequent surgery, suggesting that I3C is a successful treatment option for RRP.85 Several NIH sponsored clinical trials at different phases to test the effects of I3C on the prostatectomy patients with PSA recurrence, the healthy participants for preventing cancer, and the nonsmoking women at high risk of breast cancer for preventing breast cancer have been completed but no results are yet available at this time (ClinicalTrials.gov). More new clinical trials are being conducted to investigate the clinical value of I3C or DIM in cancer patients (Table 2). These clinical trials are focused on investigating the effects of I3C or DIM in the treatment of prostate cancer or cervical dysplasia. We believe that I3C and DIM could be useful and potent agents either alone or combination with conventional therapeutics for the treatment of human malignancies in the future. Because studies have shown that I3C may have adverse effects,77 DIM has been the recent choice for further clinical studies. In a recent study, the authors have shown the phase I data supporting the non-toxic nature of DIM in healthy volunteers.86 Our group has recently shown that a formulated DIM (B-DIM) is biologically active in vitro and in pre-clinical animal model in vivo,64,68 which prompted us to conduct a phase I clinical trials in prostate cancer patients. We found that B-DIM was well tolerated and that pharmacokinetics revealed that drug exposure was dose proportional with measurable levels of B-DIM in the plasma.87 Serum PSA stabilization and partial response were seen at the recommended phase II dose of B-DIM (225 mg twice daily), suggesting the beneficial effects of B-DIM in prostate cancer treatment, which is being tested by a phase II study in out institution.
Table 2.
The new or ongoing clinical trials to investigate the value of I3C or DIM in cancer clinic *.
| NCT ID | Title |
|---|---|
| NCT00607932 | Brassica vegetables or indole-3-carbinol in treating patients with PSA recurrence after surgery for prostate cancer |
| NCT00450229 | Diindolylmethane in treating patients undergoing surgery for stage I or stage II prostate cancer |
| NCT00305747 | Diindolylmethane in treating patients with nonmetastatic prostate cancer that has not responded to previous hormone therapy |
| NCT00462813 | Diindolylmethane in treating patients with abnormal cervical cells |
| NCT00212381 | Oral diindolylmethane (DIM) for the treatment of cervical dysplasia |
| NCT00784394 | Diindolylmethane in healthy nonsmokers |
Information collected from ClinicalTrials.gov
The roles of curcumin in cancer therapy
Targeting multiple pathways by curcumin
Curcumin has been used for the inhibition of inflammation because of its strong inhibitory effect on NF-κB; however, curcumin also inhibits proliferation, angiogenesis, invasion, and metastasis of cancer cells through the regulation of multiple cell signaling pathways.88 It has been reported that curcumin down-regulated IKK and suppressed NF-κB nuclear translocation.89 Recent studies have shown that curcumin inhibited constitutive activation of NF-κB 90 and sensitized human colorectal cancer xenografts in nude mice to γ-radiation by targeting NF-κB-regulated gene products.91 It has also been reported that liposomal formulation of curcumin could cause a dose-dependent growth inhibition of cancer cells and a decreased activation of NF-κB.92 NF-κB target genes including cyclin D1, cyclooxygenase-2, matrix metalloproteinase-9, Bcl-2, Bcl-xL, Mcl-1L, and Mcl-1S were also down-regulated, suggesting the effect of curcumin on the NF-κB signaling pathway. The NF-κB targeted effects of curcumin could also be due to the inhibition of proteasome activity as reported recently by our group.93 Furthermore, clinical study showed that curcumin inhibited the expressions of NF-κB and cyclooxygenase-2 in peripheral blood mononuclear cells in pancreatic cancer patients.12 These results clearly indicate that curcumin could inhibit NF-κB signaling in vitro and in vivo (Fig 1).
Apart from NF-κB, Akt signaling is also a target gene of curcumin. Curcumin inhibited the phosphorylation of Akt, mTOR, and their downstream targets in prostate cancer cells in a dose- and time-dependent manner.94 Curcumin also suppressed the proliferation of cisplatin-resistant ovarian cancer cells through the inhibition of Akt activation.95 In addition, an analogue of curcumin, 4-hydroxy-3-methoxybenzoic acid methyl ester (HMBME), has been found to inhibit the proliferation of cancer cells and induce apoptosis.96 HMBME decreased the level of phosphorylated Akt, inhibited Akt kinase activity, and reduced DNA-binding activity of NF-κB.96 Other investigators also reported that curcumin targeted the molecules within the Akt signaling pathways and that the inhibition of Akt activity by curcumin led to the inhibition of proliferation and induction of apoptosis in cancer cells (Fig 1).97,98 Despite the biological activity of curcumin in vitro and in selected models in vivo, curcumin is perceived as an agent with poor tissue bioavailability.99 Preclinical and clinical pilot studies have shown that concentrations of curcumin achieved in plasma and target tissues are low. It has been reported that the plasma and colorectal tissue concentrations of curcumin in patients receiving 3,600 mg curcumin orally were 11.1 nM 100 and 12.7 nM,101 respectively. Furthermore, the level of curcumin concentration in the liver was below the limit of detection.102 These findings suggest that novel synthetic analogs of curcumin must be developed or the bioavailability of curcumin must be improved by nano-formulation. In our recent study, we have shown that a curcumin analog is biologically active and also inhibit proteasome 103 which is consistent with our previous findings.93 These results are encouraging, and thus further studies are needed for testing whether curcumin analog could show superior bioavailability compared to curcumin and, in turn, could show better anti-tumor activity.
Curcumin also modulates MAPK signaling pathway, resulting in the inhibition of inflammation and cancer cell growth. It has been reported that curcumin inhibited the activation of MAPK and the translocation of NF-κB.104 Curcumin could attenuate experimental colitis through a reduction in the activity of p38 MAPK.105 Gene expression profiles of curcumin treated cells showed that curcumin suppressed the expression of MEKK4, MKK4, and JNK,106 demonstrating its inhibitory effect on MAPK signaling.
Curcumin also exerts its inhibitory effects on Wnt signaling. It has been reported that curcumin suppressed β-catenin response transcription activated by Wnt3a and inhibited the growth of various colon cancer cell lines.107 Curcumin could also induce caspase-3-mediated degradation of β-catenin,108 leading to the reduced binding of β-catenin to TCF and the inactivation of Wnt signaling. Curcumin has been found to down-regulate p300, which is a positive regulator of the Wnt/β-catenin pathway.107 Gene expression profile analysis showed that the expression of Frizzled-1 (Wnt receptor) was strongly attenuated by curcumin.106 These results suggest that curcumin could inhibit cancer cell growth through the inhibition of Wnt signaling pathway (Fig 2).
It has been found that curcumin down-regulated the transactivation and expression of AR and AR-related molecules (AP-1 and NF-κB), and reduced colony formation in soft agar.109 Several curcumin analogues has been evaluated as potential androgen receptor antagonists in the presence of AR and AR coactivator, ARA70.110 The results demonstrated that some curcumin analogs have potent anti-androgenic activities and were superior to hydroxyflutamide, which is one of the currently available anti-androgen for the treatment of prostate cancer. The results from structure-activity relationship studies confirmed that some moieties were important factors related to the anti-androgenic activity.110 These results suggest that the curcumin analogs could serve as a new class of anti-androgenic agents to control AR-mediated prostate cancer growth.
Inhibition of tumor growth in animal by curcumin
The effects of curcumin on tumor formation and growth in animals have been tested. In a study evaluating the effects of curcumin on N-bis(2-hydroxypropyl)nitrosamine (DHPN) induced tumor in BALB/c mice, curcumin has been found to significantly inhibit pulmonary and liver adenoma formation and growth in BALB/c mice.111 The effects of curcumin on 7,12-dimethylbenz[a]anthracene (DMBA)-induced oral carcinogenesis in hamsters has also been investigated.112 It has been found that curcumin significantly decreased the visible tumor incidence and the squamous cell carcinoma incidence of oral cancer.112 Curcumin also decreased the proliferation index in hyperplasia, dysplasia and papillomas and inhibited the angiogenesis in papilloma and squamous cell carcinoma.112 These results suggested that curcumin had inhibitory effects against carcinogenesis. It has been reported that liposome-encapsulated curcumin suppressed growth of head and neck squamous cell carcinoma in xenografts through the inhibition of NF-κB and the NF-κB related molecules including cyclin D1, cyclooxygenase-2, MMP-9, Bcl-2, Bcl-xL, Mcl-1L, and Mcl-1S.92 Curcumin has also been found to sensitize human colorectal cancer xenografts in nude mice to gamma-radiation by targeting NF-κB and NF-κB-regulated molecules, cyclin D1, c-myc, Bcl-2, Bcl-xL, cyclooxygenase-2, MMP-9, and VEGF.91 Curcumin also potentiated antitumor activity of gemcitabine in an orthotopic model of pancreatic cancer through the down-regulation of NF-κB signaling.113 Moreover, curcumin has been found to prevent hematogenous breast cancer metastases in immunodeficient mice with down-regulation of NF-κB, AP-1, and MMP-9. These results clearly suggest that curcumin is a potent agent for the inhibition of carcinogenesis and tumor progression in vivo in animal models, which clearly provided rationale for testing the anti-tumor activity of curcumin in human patients as summarized below.
Clinical trials for cancer therapy using curcumin
In a phase I trial using combination treatment with curcumin and quercetin to regress adenomas in patients with familial adenomatous polyposis (FAP), all 5 patients had a decreased polyp number and size from baseline after 6 months of curcumin and quercetin treatment without appreciable toxicity,114 suggesting that curcumin could inhibit adenoma growth in vivo. Another phase I trial has been conducted to evaluate the dose and the biomarkers in fifteen patients with advanced colorectal cancer refractory to standard chemotherapies. It has been found that a daily dose of 3.6 g curcumin caused 62% decreases in inducible PGE2 production in blood samples, indicating the biological activity in humans.100 The effects of curcumin has also been tested in a phase I trial in patients with high-risk or pre-malignant lesions which include recently resected urinary bladder cancer, arsenic Bowen's disease of the skin, uterine cervical intraepithelial neoplasm, oral leucoplakia, and intestinal metaplasia of the stomach.115 The results showed that histologic improvement of precancerous lesions was seen in 1 out of 2 patients with recently resected bladder cancer, 2 out of 7 patients of oral leucoplakia, 1 out of 6 patients of intestinal metaplasia of the stomach, 1 out of 4 patients with cervical intraepithelial neoplasm and 2 out of 6 patients with Bowen's disease after curcumin treatment. This study also demonstrated that curcumin was non-toxic to humans up to 8,000 mg/day by mouth for 3 months and also suggesting the biologic effect in the chemoprevention of cancer. A phase II trial of curcumin in patients with advanced pancreatic cancer has been reported.12 In this study, twenty-one patients received 8 g curcumin by mouth daily until disease progression. Out of 21 cases, two patients showed clinical evidence for biological activity. Among them, one had ongoing stable disease for more than 18 months and another patient had a significant tumor regression (73%).12 No toxicities were observed in all patients during the trial period. Theses results suggest that oral curcumin is well tolerated and has biological activity in some patients with pancreatic cancer; however further improvements in bioavailability of curcumin or its synthetic analogs are warranted.
More new clinical trials are being conducted to investigate the value of curcumin in cancer patients (Table 3). These clinical trials are ongoing to investigate the efficacy of curcumin in the prevention setting for the development of various cancers, and the combination treatment of cancer with conventional chemotherapeutic agents or other dietary agents. We also believe that cancer chemotherapy combined with curcumin supplement could be an important novel strategy for the treatment of cancers and/or for the prevention of cancer progression of various human cancers.
Table 3.
The new or ongoing clinical trials to investigate the value of curcumin in cancer clinic*.
| NCT ID | Title |
|---|---|
| NCT00745134 | Curcumin with pre-operative capecitabine and radiation therapy followed by surgery for rectal cancer |
| NCT00094445 | Trial of curcumin in advanced pancreatic cancer |
| NCT00365209 | Curcumin in preventing colon cancer in smokers with aberrant crypt foci |
| NCT00896103 | Curcumin and docetaxel in treating patients with metastatic breast |
| NCT00192842 | Gemcitabine with curcumin for pancreatic cancer |
| NCT00295035 | Phase III trial of gemcitabine, curcumin and celebrex in patients with metastatic colon cancer |
| NCT00486460 | Phase III trial of gemcitabine, curcumin and celebrex in patients with advance or inoperable pancreatic cancer |
| NCT00003365 | Sulindac and plant compounds in preventing colon cancer |
| NCT00113841 | Curcumin (diferuloylmethane derivative) with or without bioperine in patients with multiple myeloma |
| NCT00118989 | Curcumin for the chemoprevention of colorectal cancer |
| NCT00542711 | Bio-availability of a new liquid tumeric extract |
| NCT00689195 | Pilot study of curcumin formulation and ashwagandha extract in advanced osteosarcoma |
| NCT00475683 | Curcumin for prevention of oral mucositis in children chemotherapy |
| NCT00768118 | A capsule containing curcumin, green tea extract, polygonum cuspidatum extract, and soybean extract in healthy participants |
| NCT00641147 | Curcumin for treatment of intestinal adenomas in familial adenomatous polyposis (FAP) |
Information collected from ClinicalTrials.gov
Conclusion and perspectives
In recent years, more dietary compounds including isoflavone, I3C, DIM, curcumin, (-)-epigallocatechin-3-gallate, resveratrol, lycopene, etc, have been recognized as cancer chemopreventative agents because of their anti-carcinogenic activity. Among them, isoflavone, I3C, DIM and curcumin, the subject of the current article, have been demonstrated to be more potent in the inhibition of carcinogenesis and cancer progression in vivo in human and in animal studies, which has been clearly supported by various in vitro experiments. These inhibitory effects have been found to be mediated through the regulation of multiple cell signaling pathways including NF-κB, Akt, MAPK, Wnt, Notch, and AR pathways. It is our humble opinion that in order to effectively suppress cancer cell growth and induce apoptosis without causing any systemic toxicity to humans, “natural agents” harvested from the bounties of nature could play an important role toward cancer prevention and therapy by targeting multiple signaling pathways. Therefore, we believe that isoflavone, I3C, DIM and curcumin could be considered multi-targeted agents which are likely to be useful as single agents or in combination with conventional therapeutics for the prevention of tumor progression and/or treatment of human malignancies. However, further in-depth mechanistic in vitro studies, relevant in vivo animal model studies, and rationally designed novel clinical trials are needed to fully appreciate the value of these “natural products” in the management of human cancers.
Acknowledgments
The authors' work cited in this reviews was funded by grants from the National Cancer Institute, NIH (5R01CA108535, 5R01CA083695, and 5R01CA101870 awarded to FHS).
Footnotes
Contributing Author: Yiwei Li, Department of Pathology, Barbara Ann Karmanos Cancer Institute, Wayne State University School of Medicine, 715 Hudson Webber Cancer Research Center, 4100 John R, Detroit, MI 48201, USA. Tel: 313-576-8318; Fax: 313-576-8389; E-mail: yiweili@med.wayne.edu
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Jemal A, Siegel R, Ward E, et al. Cancer statistics, 2008. CA Cancer J Clin. 2008;58:71–96. doi: 10.3322/CA.2007.0010. [DOI] [PubMed] [Google Scholar]
- 2.Lee MM, Gomez SL, Chang JS, Wey M, Wang RT, Hsing AW. Soy and isoflavone consumption in relation to prostate cancer risk in China. Cancer Epidemiol Biomarkers Prev. 2003;12:665–8. [PubMed] [Google Scholar]
- 3.Smith-Warner SA, Spiegelman D, Yaun SS, et al. Fruits, vegetables and lung cancer: a pooled analysis of cohort studies. Int J Cancer. 2003;107:1001–11. doi: 10.1002/ijc.11490. [DOI] [PubMed] [Google Scholar]
- 4.Surh YJ. Cancer chemoprevention with dietary phytochemicals. Nat Rev Cancer. 2003;3:768–80. doi: 10.1038/nrc1189. [DOI] [PubMed] [Google Scholar]
- 5.Khan N, Afaq F, Mukhtar H. Apoptosis by dietary factors: the suicide solution for delaying cancer growth. Carcinogenesis. 2007;28:233–9. doi: 10.1093/carcin/bgl243. [DOI] [PubMed] [Google Scholar]
- 6.Sarkar FH, Li Y, Wang Z, Kong D. Cellular signaling perturbation by natural products. Cell Signal. 2009 doi: 10.1016/j.cellsig.2009.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sarkar FH, Li YW. Targeting multiple signal pathways by chemopreventive agents for cancer prevention and therapy. Acta Pharmacol Sin. 2007;28:1305–15. doi: 10.1111/j.1745-7254.2007.00689.x. [DOI] [PubMed] [Google Scholar]
- 8.Hamilton-Reeves JM, Rebello SA, Thomas W, Slaton JW, Kurzer MS. Isoflavone-rich soy protein isolate suppresses androgen receptor expression without altering estrogen receptor-beta expression or serum hormonal profiles in men at high risk of prostate cancer. J Nutr. 2007;137:1769–75. doi: 10.1093/jn/137.7.1769. [DOI] [PubMed] [Google Scholar]
- 9.Pendleton JM, Tan WW, Anai S, et al. Phase II trial of isoflavone in prostate-specific antigen recurrent prostate cancer after previous local therapy. BMC Cancer. 2008;8:132. doi: 10.1186/1471-2407-8-132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Vaishampayan U, Hussain M, Banerjee M, et al. Lycopene and soy isoflavones in the treatment of prostate cancer. Nutr Cancer. 2007;59:1–7. doi: 10.1080/01635580701413934. [DOI] [PubMed] [Google Scholar]
- 11.Bemis DL, Katz AE, Buttyan R. Clinical trials of natural products as chemopreventive agents for prostate cancer. Expert Opin Investig Drugs. 2006;15:1191–200. doi: 10.1517/13543784.15.10.1191. [DOI] [PubMed] [Google Scholar]
- 12.Dhillon N, Aggarwal BB, Newman RA, et al. Phase II trial of curcumin in patients with advanced pancreatic cancer. Clin Cancer Res. 2008;14:4491–9. doi: 10.1158/1078-0432.CCR-08-0024. [DOI] [PubMed] [Google Scholar]
- 13.Naik R, Nixon S, Lopes A, Godfrey K, Hatem MH, Monaghan JM. A randomized phase II trial of indole-3-carbinol in the treatment of vulvar intraepithelial neoplasia. Int J Gynecol Cancer. 2006;16:786–90. doi: 10.1111/j.1525-1438.2006.00386.x. [DOI] [PubMed] [Google Scholar]
- 14.Barnes S. The chemopreventive properties of soy isoflavonoids in animal models of breast cancer. Breast Cancer Res Treat. 1997;46:169–79. doi: 10.1023/a:1005956326155. [DOI] [PubMed] [Google Scholar]
- 15.Dixon RA, Ferreira D. Genistein. Phytochemistry. 2002;60:205–11. doi: 10.1016/s0031-9422(02)00116-4. [DOI] [PubMed] [Google Scholar]
- 16.Li Y, Sarkar FH. Down-regulation of invasion and angiogenesis-related genes identified by cDNA microarray analysis of PC3 prostate cancer cells treated with genistein. Cancer Lett. 2002;186:157–64. doi: 10.1016/s0304-3835(02)00349-x. [DOI] [PubMed] [Google Scholar]
- 17.Chinni SR, Li Y, Upadhyay S, Koppolu PK, Sarkar FH. Indole-3-carbinol (I3C) induced cell growth inhibition, G1 cell cycle arrest and apoptosis in prostate cancer cells. Oncogene. 2001;20:2927–36. doi: 10.1038/sj.onc.1204365. [DOI] [PubMed] [Google Scholar]
- 18.Li Y, Li X, Sarkar FH. Gene expression profiles of I3C- and DIM-treated PC3 human prostate cancer cells determined by cDNA microarray analysis. J Nutr. 2003;133:1011–9. doi: 10.1093/jn/133.4.1011. [DOI] [PubMed] [Google Scholar]
- 19.Mukhopadhyay A, Bueso-Ramos C, Chatterjee D, Pantazis P, Aggarwal BB. Curcumin downregulates cell survival mechanisms in human prostate cancer cell lines. Oncogene. 2001;20:7597–609. doi: 10.1038/sj.onc.1204997. [DOI] [PubMed] [Google Scholar]
- 20.Shao ZM, Shen ZZ, Liu CH, et al. Curcumin exerts multiple suppressive effects on human breast carcinoma cells. Int J Cancer. 2002;98:234–40. doi: 10.1002/ijc.10183. [DOI] [PubMed] [Google Scholar]
- 21.Davis JN, Kucuk O, Sarkar FH. Genistein inhibits NF-kappa B activation in prostate cancer cells. Nutr Cancer. 1999;35:167–74. doi: 10.1207/S15327914NC352_11. [DOI] [PubMed] [Google Scholar]
- 22.Chen CC, Sun YT, Chen JJ, Chiu KT. TNF-alpha-induced cyclooxygenase-2 expression in human lung epithelial cells: involvement of the phospholipase C-gamma 2, protein kinase C-alpha, tyrosine kinase, NF-kappa B-inducing kinase, and I-kappa B kinase 1/2 pathway. J Immunol. 2000;165:2719–28. doi: 10.4049/jimmunol.165.5.2719. [DOI] [PubMed] [Google Scholar]
- 23.Natarajan K, Manna SK, Chaturvedi MM, Aggarwal BB. Protein tyrosine kinase inhibitors block tumor necrosis factor-induced activation of nuclear factor-kappaB, degradation of IkappaBalpha, nuclear translocation of p65, and subsequent gene expression. Arch Biochem Biophys. 1998;352:59–70. doi: 10.1006/abbi.1998.0576. [DOI] [PubMed] [Google Scholar]
- 24.Li Y, Ahmed F, Ali S, Philip PA, Kucuk O, Sarkar FH. Inactivation of nuclear factor kappaB by soy isoflavone genistein contributes to increased apoptosis induced by chemotherapeutic agents in human cancer cells. Cancer Res. 2005;65:6934–42. doi: 10.1158/0008-5472.CAN-04-4604. [DOI] [PubMed] [Google Scholar]
- 25.Li Y, Sarkar FH. Inhibition of nuclear factor kappaB activation in PC3 cells by genistein is mediated via Akt signaling pathway. Clin Cancer Res. 2002;8:2369–77. [PubMed] [Google Scholar]
- 26.Gong L, Li Y, Nedeljkovic-Kurepa A, Sarkar FH. Inactivation of NF-kappaB by genistein is mediated via Akt signaling pathway in breast cancer cells. Oncogene. 2003;22:4702–9. doi: 10.1038/sj.onc.1206583. [DOI] [PubMed] [Google Scholar]
- 27.Akiyama T, Ishida J, Nakagawa S, et al. Genistein, a specific inhibitor of tyrosine-specific protein kinases. J Biol Chem. 1987;262:5592–5. [PubMed] [Google Scholar]
- 28.Li Y, Wang Z, Kong D, Li R, Sarkar SH, Sarkar FH. Regulation of Akt/FOXO3a/GSK-3beta/AR signaling network by isoflavone in prostate cancer cells. J Biol Chem. 2008;283:27707–16. doi: 10.1074/jbc.M802759200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Su Y, Simmen FA, Xiao R, Simmen RC. Expression profiling of rat mammary epithelial cells reveals candidate signaling pathways in dietary protection from mammary tumors. Physiol Genomics. 2007;30:8–16. doi: 10.1152/physiolgenomics.00023.2007. [DOI] [PubMed] [Google Scholar]
- 30.Su Y, Simmen RC. Soy isoflavone genistein upregulates epithelial adhesion molecule E-cadherin expression and attenuates beta-catenin signaling in mammary epithelial cells. Carcinogenesis. 2009;30:331–9. doi: 10.1093/carcin/bgn279. [DOI] [PubMed] [Google Scholar]
- 31.Wang Z, Zhang Y, Li Y, Banerjee S, Liao J, Sarkar FH. Down-regulation of Notch-1 contributes to cell growth inhibition and apoptosis in pancreatic cancer cells. Mol Cancer Ther. 2006;5:483–93. doi: 10.1158/1535-7163.MCT-05-0299. [DOI] [PubMed] [Google Scholar]
- 32.Wang Z, Zhang Y, Banerjee S, Li Y, Sarkar FH. Inhibition of nuclear factor kappab activity by genistein is mediated via Notch-1 signaling pathway in pancreatic cancer cells. Int J Cancer. 2006;118:1930–6. doi: 10.1002/ijc.21589. [DOI] [PubMed] [Google Scholar]
- 33.Davis JN, Muqim N, Bhuiyan M, Kucuk O, Pienta KJ, Sarkar FH. Inhibition of prostate specific antigen expression by genistein in prostate cancer cells. Int J Oncol. 2000;16:1091–7. doi: 10.3892/ijo.16.6.1091. [DOI] [PubMed] [Google Scholar]
- 34.Davis JN, Kucuk O, Sarkar FH. Expression of prostate-specific antigen is transcriptionally regulated by genistein in prostate cancer cells. Mol Carcinog. 2002;34:91–101. doi: 10.1002/mc.10053. [DOI] [PubMed] [Google Scholar]
- 35.Fritz WA, Wang J, Eltoum IE, Lamartiniere CA. Dietary genistein down-regulates androgen and estrogen receptor expression in the rat prostate. Mol Cell Endocrinol. 2002;186:89–99. doi: 10.1016/s0303-7207(01)00663-3. [DOI] [PubMed] [Google Scholar]
- 36.Sharma OP, Adlercreutz H, Strandberg JD, Zirkin BR, Coffey DS, Ewing LL. Soy of dietary source plays a preventive role against the pathogenesis of prostatitis in rats. J Steroid Biochem Mol Biol. 1992;43:557–64. doi: 10.1016/0960-0760(92)90244-d. [DOI] [PubMed] [Google Scholar]
- 37.Onozawa M, Kawamori T, Baba M, et al. Effects of a soybean isoflavone mixture on carcinogenesis in prostate and seminal vesicles of F344 rats. Jpn J Cancer Res. 1999;90:393–8. doi: 10.1111/j.1349-7006.1999.tb00760.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Landstrom M, Zhang JX, Hallmans G, et al. Inhibitory effects of soy and rye diets on the development of Dunning R3327 prostate adenocarcinoma in rats. Prostate. 1998;36:151–61. doi: 10.1002/(sici)1097-0045(19980801)36:3<151::aid-pros2>3.0.co;2-k. [DOI] [PubMed] [Google Scholar]
- 39.Zhou JR, Gugger ET, Tanaka T, Guo Y, Blackburn GL, Clinton SK. Soybean phytochemicals inhibit the growth of transplantable human prostate carcinoma and tumor angiogenesis in mice. J Nutr. 1999;129:1628–35. doi: 10.1093/jn/129.9.1628. [DOI] [PubMed] [Google Scholar]
- 40.Li D, Yee JA, McGuire MH, Murphy PA, Yan L. Soybean isoflavones reduce experimental metastasis in mice. J Nutr. 1999;129:1075–8. doi: 10.1093/jn/129.5.1075. [DOI] [PubMed] [Google Scholar]
- 41.Lamartiniere CA, Moore JB, Brown NM, Thompson R, Hardin MJ, Barnes S. Genistein suppresses mammary cancer in rats. Carcinogenesis. 1995;16:2833–40. doi: 10.1093/carcin/16.11.2833. [DOI] [PubMed] [Google Scholar]
- 42.Uckun FM, Evans WE, Forsyth CJ, et al. Biotherapy of B-cell precursor leukemia by targeting genistein to CD19-associated tyrosine kinases. Science. 1995;267:886–91. doi: 10.1126/science.7531365. [DOI] [PubMed] [Google Scholar]
- 43.Wei H, Wei L, Frenkel K, Bowen R, Barnes S. Inhibition of tumor promoter-induced hydrogen peroxide formation in vitro and in vivo by genistein. Nutr Cancer. 1993;20:1–12. doi: 10.1080/01635589309514265. [DOI] [PubMed] [Google Scholar]
- 44.Kameoka S, Leavitt P, Chang C, Kuo SM. Expression of antioxidant proteins in human intestinal Caco-2 cells treated with dietary flavonoids. Cancer Lett. 1999;146:161–7. doi: 10.1016/s0304-3835(99)00253-0. [DOI] [PubMed] [Google Scholar]
- 45.Davis JN, Kucuk O, Djuric Z, Sarkar FH. Soy isoflavone supplementation in healthy men prevents NF-kappa B activation by TNF-alpha in blood lymphocytes. Free Radic Biol Med. 2001;30:1293–302. doi: 10.1016/s0891-5849(01)00535-4. [DOI] [PubMed] [Google Scholar]
- 46.Hussain M, Banerjee M, Sarkar FH, et al. Soy isoflavones in the treatment of prostate cancer. Nutr Cancer. 2003;47:111–7. doi: 10.1207/s15327914nc4702_1. [DOI] [PubMed] [Google Scholar]
- 47.Pendleton JM, Tan WW, Anai S, et al. Phase II trial of isoflavone in prostate-specific antigen recurrent prostate cancer after previous local therapy. BMC Cancer. 2008;8:132. doi: 10.1186/1471-2407-8-132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hamilton-Reeves JM, Rebello SA, Thomas W, Slaton JW, Kurzer MS. Isoflavone-rich soy protein isolate suppresses androgen receptor expression without altering estrogen receptor-beta expression or serum hormonal profiles in men at high risk of prostate cancer. J Nutr. 2007;137:1769–75. doi: 10.1093/jn/137.7.1769. [DOI] [PubMed] [Google Scholar]
- 49.Joniau S, Goeman L, Roskams T, Lerut E, Oyen R, Van PH. Effect of nutritional supplement challenge in patients with isolated high-grade prostatic intraepithelial neoplasia. Urology. 2007;69:1102–6. doi: 10.1016/j.urology.2007.02.063. [DOI] [PubMed] [Google Scholar]
- 50.Kumar NB, Cantor A, Allen K, et al. The specific role of isoflavones in reducing prostate cancer risk. Prostate. 2004;59:141–7. doi: 10.1002/pros.10362. [DOI] [PubMed] [Google Scholar]
- 51.Vere White RW, Hackman RM, Soares SE, Beckett LA, Li Y, Sun B. Effects of a genistein-rich extract on PSA levels in men with a history of prostate cancer. Urology. 2004;63:259–63. doi: 10.1016/j.urology.2003.09.061. [DOI] [PubMed] [Google Scholar]
- 52.Adams KF, Chen C, Newton KM, Potter JD, Lampe JW. Soy isoflavones do not modulate prostate-specific antigen concentrations in older men in a randomized controlled trial. Cancer Epidemiol Biomarkers Prev. 2004;13:644–8. [PubMed] [Google Scholar]
- 53.Martin PM, Horwitz KB, Ryan DS, McGuire WL. Phytoestrogen interaction with estrogen receptors in human breast cancer cells. Endocrinology. 1978;103:1860–7. doi: 10.1210/endo-103-5-1860. [DOI] [PubMed] [Google Scholar]
- 54.Messina MJ, Wood CE. Soy isoflavones, estrogen therapy, and breast cancer risk: analysis and commentary. Nutr J. 2008;7:17. doi: 10.1186/1475-2891-7-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lampe JW, Nishino Y, Ray RM, et al. Plasma isoflavones and fibrocystic breast conditions and breast cancer among women in Shanghai, China. Cancer Epidemiol Biomarkers Prev. 2007;16:2579–86. doi: 10.1158/1055-9965.EPI-07-0368. [DOI] [PubMed] [Google Scholar]
- 56.Dai Q, Franke AA, Yu H, et al. Urinary phytoestrogen excretion and breast cancer risk: evaluating potential effect modifiers endogenous estrogens and anthropometrics. Cancer Epidemiol Biomarkers Prev. 2003;12:497–502. [PubMed] [Google Scholar]
- 57.Sartippour MR, Rao JY, Apple S, et al. A pilot clinical study of short-term isoflavone supplements in breast cancer patients. Nutr Cancer. 2004;49:59–65. doi: 10.1207/s15327914nc4901_8. [DOI] [PubMed] [Google Scholar]
- 58.Kumar NB, Cantor A, Allen K, Riccardi D, Cox CE. The specific role of isoflavones on estrogen metabolism in premenopausal women. Cancer. 2002;94:1166–74. doi: 10.1002/cncr.10320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Bobe G, Sansbury LB, Albert PS, et al. Dietary flavonoids and colorectal adenoma recurrence in the Polyp Prevention Trial. Cancer Epidemiol Biomarkers Prev. 2008;17:1344–53. doi: 10.1158/1055-9965.EPI-07-0747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Adams KF, Lampe PD, Newton KM, et al. Soy protein containing isoflavones does not decrease colorectal epithelial cell proliferation in a randomized controlled trial. Am J Clin Nutr. 2005;82:620–6. doi: 10.1093/ajcn.82.3.620. [DOI] [PubMed] [Google Scholar]
- 61.Ahmad IU, Forman JD, Sarkar F, et al. Reduction of adverse events by soy isoflavones in patients undergoing external beam radiation therapy for prostate cancer. Int J Radiat Oncol Biol Phys. 2009;72:S318. [Google Scholar]
- 62.Verhoeven DT, Verhagen H, Goldbohm RA, van den Brandt PA, van PG. A review of mechanisms underlying anticarcinogenicity by brassica vegetables. Chem Biol Interact. 1997;103:79–129. doi: 10.1016/s0009-2797(96)03745-3. [DOI] [PubMed] [Google Scholar]
- 63.Dashwood RH, Fong AT, Arbogast DN, Bjeldanes LF, Hendricks JD, Bailey GS. Anticarcinogenic activity of indole-3-carbinol acid products: ultrasensitive bioassay by trout embryo microinjection. Cancer Res. 1994;54:3617–9. [PubMed] [Google Scholar]
- 64.Bhuiyan MM, Li Y, Banerjee S, et al. Down-regulation of androgen receptor by 3,3′-diindolylmethane contributes to inhibition of cell proliferation and induction of apoptosis in both hormone-sensitive LNCaP and insensitive C4-2B prostate cancer cells. Cancer Res. 2006;66:10064–72. doi: 10.1158/0008-5472.CAN-06-2011. [DOI] [PubMed] [Google Scholar]
- 65.Li Y, Chinni SR, Sarkar FH. Selective growth regulatory and pro-apoptotic effects of DIM is mediated by AKT and NF-kappaB pathways in prostate cancer cells. Front Biosci. 2005;10:236–43. doi: 10.2741/1523. [DOI] [PubMed] [Google Scholar]
- 66.Chinni SR, Sarkar FH. Akt inactivation is a key event in indole-3-carbinol-induced apoptosis in PC-3 cells. Clin Cancer Res. 2002;8:1228–36. [PubMed] [Google Scholar]
- 67.Leong H, Riby JE, Firestone GL, Bjeldanes LF. Potent ligand-independent estrogen receptor activation by 3,3′-diindolylmethane is mediated by cross talk between the protein kinase A and mitogen-activated protein kinase signaling pathways. Mol Endocrinol. 2004;18:291–302. doi: 10.1210/me.2003-0196. [DOI] [PubMed] [Google Scholar]
- 68.Li Y, Wang Z, Kong D, et al. Regulation of FOXO3a/beta-catenin/GSK-3beta signaling by 3,3′-diindolylmethane contributes to inhibition of cell proliferation and induction of apoptosis in prostate cancer cells. J Biol Chem. 2007;282:21542–50. doi: 10.1074/jbc.M701978200. [DOI] [PubMed] [Google Scholar]
- 69.Le HT, Schaldach CM, Firestone GL, Bjeldanes LF. Plant-derived 3,3′-Diindolylmethane is a strong androgen antagonist in human prostate cancer cells. J Biol Chem. 2003;278:21136–45. doi: 10.1074/jbc.M300588200. [DOI] [PubMed] [Google Scholar]
- 70.Bradlow HL, Michnovicz J, Telang NT, Osborne MP. Effects of dietary indole-3-carbinol on estradiol metabolism and spontaneous mammary tumors in mice. Carcinogenesis. 1991;12:1571–4. doi: 10.1093/carcin/12.9.1571. [DOI] [PubMed] [Google Scholar]
- 71.Kojima T, Tanaka T, Mori H. Chemoprevention of spontaneous endometrial cancer in female Donryu rats by dietary indole-3-carbinol. Cancer Res. 1994;54:1446–9. [PubMed] [Google Scholar]
- 72.Manson MM, Hudson EA, Ball HW, et al. Chemoprevention of aflatoxin B1-induced carcinogenesis by indole-3-carbinol in rat liver--predicting the outcome using early biomarkers. Carcinogenesis. 1998;19:1829–36. doi: 10.1093/carcin/19.10.1829. [DOI] [PubMed] [Google Scholar]
- 73.Stresser DM, Bailey GS, Williams DE. Indole-3-carbinol and beta-naphthoflavone induction of aflatoxin B1 metabolism and cytochromes P-450 associated with bioactivation and detoxication of aflatoxin B1 in the rat. Drug Metab Dispos. 1994;22:383–91. [PubMed] [Google Scholar]
- 74.Bradfield CA, Bjeldanes LF. Effect of dietary indole-3-carbinol on intestinal and hepatic monooxygenase, glutathione S-transferase and epoxide hydrolase activities in the rat. Food Chem Toxicol. 1984;22:977–82. doi: 10.1016/0278-6915(84)90147-9. [DOI] [PubMed] [Google Scholar]
- 75.Souli E, Machluf M, Morgenstern A, Sabo E, Yannai S. Indole-3-carbinol (I3C) exhibits inhibitory and preventive effects on prostate tumors in mice. Food Chem Toxicol. 2008;46:863–70. doi: 10.1016/j.fct.2007.10.026. [DOI] [PubMed] [Google Scholar]
- 76.Kassie F, Anderson LB, Scherber R, et al. Indole-3-carbinol inhibits 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone plus benzo(a)pyrene-induced lung tumorigenesis in A/J mice and modulates carcinogen-induced alterations in protein levels. Cancer Res. 2007;67:6502–11. doi: 10.1158/0008-5472.CAN-06-4438. [DOI] [PubMed] [Google Scholar]
- 77.Oganesian A, Hendricks JD, Pereira CB, Orner GA, Bailey GS, Williams DE. Potency of dietary indole-3-carbinol as a promoter of aflatoxin B1-initiated hepatocarcinogenesis: results from a 9000 animal tumor study. Carcinogenesis. 1999;20:453–8. doi: 10.1093/carcin/20.3.453. [DOI] [PubMed] [Google Scholar]
- 78.Doppalapudi RS, Riccio ES, Rausch LL, et al. Evaluation of chemopreventive agents for genotoxic activity. Mutat Res. 2007;629:148–60. doi: 10.1016/j.mrgentox.2007.02.004. [DOI] [PubMed] [Google Scholar]
- 79.Chang X, Tou JC, Hong C, et al. 3,3′-Diindolylmethane inhibits angiogenesis and the growth of transplantable human breast carcinoma in athymic mice. Carcinogenesis. 2005;26:771–8. doi: 10.1093/carcin/bgi018. [DOI] [PubMed] [Google Scholar]
- 80.Kong D, Banerjee S, Huang W, et al. Mammalian target of rapamycin repression by 3,3′-diindolylmethane inhibits invasion and angiogenesis in platelet-derived growth factor-D-overexpressing PC3 cells. Cancer Res. 2008;68:1927–34. doi: 10.1158/0008-5472.CAN-07-3241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Kong D, Li Y, Wang Z, Banerjee S, Sarkar FH. Inhibition of angiogenesis and invasion by 3,3′-diindolylmethane is mediated by the nuclear factor-kappaB downstream target genes MMP-9 and uPA that regulated bioavailability of vascular endothelial growth factor in prostate cancer. Cancer Res. 2007;67:3310–9. doi: 10.1158/0008-5472.CAN-06-4277. [DOI] [PubMed] [Google Scholar]
- 82.Ali S, Banerjee S, Ahmad A, El-Rayes BF, Philip PA, Sarkar FH. Apoptosis-inducing effect of erlotinib is potentiated by 3,3′-diindolylmethane in vitro and in vivo using an orthotopic model of pancreatic cancer. Mol Cancer Ther. 2008;7:1708–19. doi: 10.1158/1535-7163.MCT-08-0354. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 83.Nachshon-Kedmi M, Fares FA, Yannai S. Therapeutic activity of 3,3′-diindolylmethane on prostate cancer in an in vivo model. Prostate. 2004;61:153–60. doi: 10.1002/pros.20092. [DOI] [PubMed] [Google Scholar]
- 84.Reed GA, Peterson KS, Smith HJ, et al. A phase I study of indole-3-carbinol in women: tolerability and effects. Cancer Epidemiol Biomarkers Prev. 2005;14:1953–60. doi: 10.1158/1055-9965.EPI-05-0121. [DOI] [PubMed] [Google Scholar]
- 85.Rosen CA, Bryson PC. Indole-3-carbinol for recurrent respiratory papillomatosis: long-term results. J Voice. 2004;18:248–53. doi: 10.1016/j.jvoice.2003.05.005. [DOI] [PubMed] [Google Scholar]
- 86.Reed GA, Sunega JM, Sullivan DK, et al. Single-dose pharmacokinetics and tolerability of absorption-enhanced 3,3′-diindolylmethane in healthy subjects. Cancer Epidemiol Biomarkers Prev. 2008;17:2619–24. doi: 10.1158/1055-9965.EPI-08-0520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Heath EI, Heilbrun LK, Vaishampayan UN, et al. A phase I dose-escalation study of oral BR-DIM (BioResponse 3,3′- Diindolylmethane) in castrate-resistant, non-metastatic, PSA relapse prostate cancer patients. 2009 Genitourinary Cancers Symposium; 2009; Abstract 188. [PMC free article] [PubMed] [Google Scholar]
- 88.Kunnumakkara AB, Anand P, Aggarwal BB. Curcumin inhibits proliferation, invasion, angiogenesis and metastasis of different cancers through interaction with multiple cell signaling proteins. Cancer Lett. 2008;269:199–225. doi: 10.1016/j.canlet.2008.03.009. [DOI] [PubMed] [Google Scholar]
- 89.Bharti AC, Donato N, Singh S, Aggarwal BB. Curcumin (diferuloylmethane) down-regulates the constitutive activation of nuclear factor-kappa B and IkappaBalpha kinase in human multiple myeloma cells, leading to suppression of proliferation and induction of apoptosis. Blood. 2003;101:1053–62. doi: 10.1182/blood-2002-05-1320. [DOI] [PubMed] [Google Scholar]
- 90.Hussain AR, Ahmed M, Al-Jomah NA, et al. Curcumin suppresses constitutive activation of nuclear factor-kappa B and requires functional Bax to induce apoptosis in Burkitt's lymphoma cell lines. Mol Cancer Ther. 2008;7:3318–29. doi: 10.1158/1535-7163.MCT-08-0541. [DOI] [PubMed] [Google Scholar]
- 91.Kunnumakkara AB, Diagaradjane P, Guha S, et al. Curcumin sensitizes human colorectal cancer xenografts in nude mice to gamma-radiation by targeting nuclear factor-kappaB-regulated gene products. Clin Cancer Res. 2008;14:2128–36. doi: 10.1158/1078-0432.CCR-07-4722. [DOI] [PubMed] [Google Scholar]
- 92.Wang D, Veena MS, Stevenson K, et al. Liposome-encapsulated curcumin suppresses growth of head and neck squamous cell carcinoma in vitro and in xenografts through the inhibition of nuclear factor kappaB by an AKT-independent pathway. Clin Cancer Res. 2008;14:6228–36. doi: 10.1158/1078-0432.CCR-07-5177. [DOI] [PubMed] [Google Scholar]
- 93.Milacic V, Banerjee S, Landis-Piwowar KR, Sarkar FH, Majumdar AP, Dou QP. Curcumin inhibits the proteasome activity in human colon cancer cells in vitro and in vivo. Cancer Res. 2008;68:7283–92. doi: 10.1158/0008-5472.CAN-07-6246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Yu S, Shen G, Khor TO, Kim JH, Kong AN. Curcumin inhibits Akt/mammalian target of rapamycin signaling through protein phosphatase-dependent mechanism. Mol Cancer Ther. 2008;7:2609–20. doi: 10.1158/1535-7163.MCT-07-2400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Weir NM, Selvendiran K, Kutala VK, et al. Curcumin induces G2/M arrest and apoptosis in cisplatin-resistant human ovarian cancer cells by modulating Akt and p38 MAPK. Cancer Biol Ther. 2007;6:178–84. doi: 10.4161/cbt.6.2.3577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Kumar AP, Garcia GE, Ghosh R, Rajnarayanan RV, Alworth WL, Slaga TJ. 4-Hydroxy-3-methoxybenzoic acid methyl ester: a curcumin derivative targets Akt/NF kappa B cell survival signaling pathway: potential for prostate cancer management. Neoplasia. 2003;5:255–66. doi: 10.1016/S1476-5586(03)80057-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Squires MS, Hudson EA, Howells L, et al. Relevance of mitogen activated protein kinase (MAPK) and phosphotidylinositol-3-kinase/protein kinase B (PI3K/PKB) pathways to induction of apoptosis by curcumin in breast cells. Biochem Pharmacol. 2003;65:361–76. doi: 10.1016/s0006-2952(02)01517-4. [DOI] [PubMed] [Google Scholar]
- 98.Woo JH, Kim YH, Choi YJ, et al. Molecular mechanisms of curcumin-induced cytotoxicity: induction of apoptosis through generation of reactive oxygen species, down-regulation of Bcl-XL and IAP, the release of cytochrome c and inhibition of Akt. Carcinogenesis. 2003;24:1199–208. doi: 10.1093/carcin/bgg082. [DOI] [PubMed] [Google Scholar]
- 99.Anand P, Thomas SG, Kunnumakkara AB, et al. Biological activities of curcumin and its analogues (Congeners) made by man and Mother Nature. Biochem Pharmacol. 2008;76:1590–611. doi: 10.1016/j.bcp.2008.08.008. [DOI] [PubMed] [Google Scholar]
- 100.Sharma RA, Euden SA, Platton SL, et al. Phase I clinical trial of oral curcumin: biomarkers of systemic activity and compliance. Clin Cancer Res. 2004;10:6847–54. doi: 10.1158/1078-0432.CCR-04-0744. [DOI] [PubMed] [Google Scholar]
- 101.Garcea G, Berry DP, Jones DJ, et al. Consumption of the putative chemopreventive agent curcumin by cancer patients: assessment of curcumin levels in the colorectum and their pharmacodynamic consequences. Cancer Epidemiol Biomarkers Prev. 2005;14:120–5. [PubMed] [Google Scholar]
- 102.Garcea G, Jones DJ, Singh R, et al. Detection of curcumin and its metabolites in hepatic tissue and portal blood of patients following oral administration. Br J Cancer. 2004;90:1011–5. doi: 10.1038/sj.bjc.6601623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Padhye S, Yang H, Jamadar A, et al. New Difluoro Knoevenagel Condensates of Curcumin, Their Schiff Bases and Copper Complexes as Proteasome Inhibitors and Apoptosis Inducers in Cancer Cells. Pharm Res. 2009 doi: 10.1007/s11095-009-9900-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Kim GY, Kim KH, Lee SH, et al. Curcumin inhibits immunostimulatory function of dendritic cells: MAPKs and translocation of NF-kappa B as potential targets. J Immunol. 2005;174:8116–24. doi: 10.4049/jimmunol.174.12.8116. [DOI] [PubMed] [Google Scholar]
- 105.Salh B, Assi K, Templeman V, et al. Curcumin attenuates DNB-induced murine colitis. Am J Physiol Gastrointest Liver Physiol. 2003;285:G235–G243. doi: 10.1152/ajpgi.00449.2002. [DOI] [PubMed] [Google Scholar]
- 106.Yan C, Jamaluddin MS, Aggarwal B, Myers J, Boyd DD. Gene expression profiling identifies activating transcription factor 3 as a novel contributor to the proapoptotic effect of curcumin. Mol Cancer Ther. 2005;4:233–41. [PubMed] [Google Scholar]
- 107.Ryu MJ, Cho M, Song JY, et al. Natural derivatives of curcumin attenuate the Wnt/beta-catenin pathway through down-regulation of the transcriptional coactivator p300. Biochem Biophys Res Commun. 2008;377:1304–8. doi: 10.1016/j.bbrc.2008.10.171. [DOI] [PubMed] [Google Scholar]
- 108.Jaiswal AS, Marlow BP, Gupta N, Narayan S. Beta-catenin-mediated transactivation and cell-cell adhesion pathways are important in curcumin (diferuylmethane)-induced growth arrest and apoptosis in colon cancer cells. Oncogene. 2002;21:8414–27. doi: 10.1038/sj.onc.1205947. [DOI] [PubMed] [Google Scholar]
- 109.Nakamura K, Yasunaga Y, Segawa T, et al. Curcumin down-regulates AR gene expression and activation in prostate cancer cell lines. Int J Oncol. 2002;21:825–30. [PubMed] [Google Scholar]
- 110.Ohtsu H, Xiao Z, Ishida J, et al. Antitumor agents. 217. Curcumin analogues as novel androgen receptor antagonists with potential as anti-prostate cancer agents. J Med Chem. 2002;45:5037–42. doi: 10.1021/jm020200g. [DOI] [PubMed] [Google Scholar]
- 111.Huang AC, Lin SY, Su CC, et al. Effects of curcumin on N-bis(2-hydroxypropyl) nitrosamine (DHPN)-induced lung and liver tumorigenesis in BALB/c mice in vivo. In Vivo. 2008;22:781–5. [PubMed] [Google Scholar]
- 112.Li N, Chen X, Liao J, et al. Inhibition of 7,12-dimethylbenz[a]anthracene (DMBA)-induced oral carcinogenesis in hamsters by tea and curcumin. Carcinogenesis. 2002;23:1307–13. doi: 10.1093/carcin/23.8.1307. [DOI] [PubMed] [Google Scholar]
- 113.Kunnumakkara AB, Guha S, Krishnan S, Diagaradjane P, Gelovani J, Aggarwal BB. Curcumin potentiates antitumor activity of gemcitabine in an orthotopic model of pancreatic cancer through suppression of proliferation, angiogenesis, and inhibition of nuclear factor-kappaB-regulated gene products. Cancer Res. 2007;67:3853–61. doi: 10.1158/0008-5472.CAN-06-4257. [DOI] [PubMed] [Google Scholar]
- 114.Cruz-Correa M, Shoskes DA, Sanchez P, et al. Combination treatment with curcumin and quercetin of adenomas in familial adenomatous polyposis. Clin Gastroenterol Hepatol. 2006;4:1035–8. doi: 10.1016/j.cgh.2006.03.020. [DOI] [PubMed] [Google Scholar]
- 115.Cheng AL, Hsu CH, Lin JK, et al. Phase I clinical trial of curcumin, a chemopreventive agent, in patients with high-risk or pre-malignant lesions. Anticancer Res. 2001;21:2895–900. [PubMed] [Google Scholar]