

Abstract
Rickettsia peacockii, also known as the East Side Agent, is a non-pathogenic obligate intracellular bacterium found as an endosymbiont in Dermacentor andersoni ticks in the western USA and Canada. Its presence in ticks is correlated with reduced prevalence of Rickettsia rickettsii, the agent of Rocky Mountain Spotted Fever. It has been proposed that a virulent SFG rickettsia underwent changes to become the East Side Agent. We determined the genome sequence of R. peacockii and provide a comparison to a closely related virulent R. rickettsii. The presence of 42 chromosomal copies of the ISRpe1 transposon in the genome of R. peacockii is associated with a lack of synteny with the genome of R. rickettsii and numerous deletions via recombination between transposon copies. The plasmid contains a number of genes from distantly related organisms, such as part of the glycosylation island of Pseudomonas aeruginosa. Genes deleted or mutated in R. peacockii which may relate to loss of virulence include those coding for an ankyrin repeat containing protein, DsbA, RickA, protease II, OmpA, ScaI, and a putative phosphoethanolamine transferase. The gene coding for the ankyrin repeat containing protein is especially implicated as it is mutated in R. rickettsii strain Iowa, which has attenuated virulence. Presence of numerous copies of the ISRpe1 transposon, likely acquired by lateral transfer from a Cardinium species, are associated with extensive genomic reorganization and deletions. The deletion and mutation of genes possibly involved in loss of virulence have been identified by this genomic comparison. It also illustrates that the introduction of a transposon into the genome can have varied effects; either correlating with an increase in pathogenicity as in Francisella tularensis or a loss of pathogenicity as in R. peacockii and the recombination enabled by multiple transposon copies can cause significant deletions in some genomes while not in others.
Introduction
Rickettsia peacockii is an obligate intracellular bacterium identified in Rocky Mountain wood ticks (Dermacentor andersoni) from Montana, USA [1]. It is of interest to rickettsiologists due to its co-localization on the eastern side of the Bitterroot Valley with a much reduced prevalence of D. andersoni infected with Rickettsia rickettsii, while spotted fever ravaged the west side of the valley [2] [3]. Thus began the study of a phenomenon considered as evidence for interference, where the presence of R. peacockii in D. andersoni ticks may prevent the transovarial transmission of R. rickettsii and therefore limit its spread in the tick population. It is not clear whether this interference is an active process or simply a case in which ticks carrying R. peacockii have a reproductive advantage because they do not suffer the reduced fecundity associated with R. rickettsii infection [4]. Surveys of rickettsiae in tick populations around the western US and Canada have shown that R. peacockii is widespread in Dermacentor ticks and R. rickettsii is relatively rare [1], [5]. While R. peacockii is closely related to R. rickettsii, it is not a pathogen of mammals and not deleterious to ticks.
The purpose of this work was to determine the sequence of the R. peacockii genome and compare it to the genome of it's nearest pathogen relative, R. rickettsii. Although the R. peacockii genome is similar in size (1.29Mb) to those of other spotted fever group (SFG) rickettsiae and there is high homology between many of their genes, the genome of R. peacockii has several gene deletions and mutations that may account for its lack of pathogenicity. In addition to the genome sequence of the Iowa strain of R. rickettsii [6] it represents a valuable set of data for comparison with the genomes of related pathogenic rickettsiae. The most dramatic difference between the genomes of R. peacockii and R. rickettsii is the presence of the ISRpe1 transposon and the effects multiple transposon copies have had as a point of homology for recombination, resulting in numerous deletions and genome shuffling (this manuscript). In contrast, genome comparisons of virulent and non-virulent Francisella tularensis showed that transposon mediated recombination and shuffling of gene order occurred in the pathogenic strains rather than the non-pathogenic strain [7]. R. peacockii is also among the growing list of rickettsiae harboring plasmids which are apparently lacking in R. rickettsii.
Results and Discussion
R. peacockii accession numbers, chromosome GenBank:CP001227,plasmid GenBank:CP001228.
The genome sequence of R. peacockii strain Rustic was determined; the size of the circular chromosome (RPR) is 1,288,492 bp and the size of the circular plasmid (pRPR) is 26,406 bp. The gene sequences of R. peacockii were found to be most similar to those of virulent R. rickettsii Sheila Smith (SS) and avirulent R. rickettsii Iowa. The genome of the non-pathogen R. peacockii was compared to the genome of its closest pathogenic neighbor R. rickettsii SS in order to identify differences that may relate to pathogenicity. Presence of laterally transferred DNA in the genome of R. peacockii is the most striking difference between them; including a plasmid, the ISRpe1 transposon and three chromosomal regions of Rickettsia bellii-like DNA containing Tra genes. The locations of chromosomal DNA sequences present in R. peacockii and lacking in R. rickettsii SS are shown in Text S1.
Impact of ISRpe1 Transposons on the R. peacockii Genome
There are 40 copies of the transposon and 2 transposon fragments on the chromosome and 2 copies of the transposon on the plasmid. Most ISRpe1 transposons contain an intact transposase coding sequence (31 of 42) while 11 contain frameshift mutations or internal stop codons. There are no other types of transposases annotated as genes on the chromosome and only three other types of transposon pseudogenes are found on the chromosome, two of which are also found in R. rickettsii and one within a fragment of the tra cluster.
Recombination between the transposons has resulted in a dramatic shuffling of gene order between the R. peacockii and R. rickettsii genomes (Figure 1). A dot plot comparison of the two genomes is shown in Figure S1. By comparing the two genomes using Mauve [8] we found that an ISRpe1 transposon in R. peacockii co-localized to 31 of 37 junctions between syntenic blocks. Numerous deletions co-localized to copies of the transposon as well, suggesting the deletions occurred during this recombination. To determine the extent of correlation between deletions and the presence of transposons, the backbone file from the Mauve comparison, Artemis [9] and blastn was used to locate copies of ISRpe1 within 5 bp of the deletion junctions. All deletions over 100 bp in size were examined. There are transposons not associated with gdeletions or changes to synteny, transposons that locate to changes in synteny with or without deletion, and transposons that locate to a point of deletion but do not affect synteny. Deletions of the latter variety likely occurred when two transposons integrated near each other followed by recombination between them and deletion of the intervening DNA. Text S2 shows that for all deletions greater than 100 bp in size, 71.4% of deletions (25 out of 35) are flanked by one or two ISRpe1 transposons. There are also 3 smaller deletions flanked by transposons. It is possible that some of the small deletions resulting in frameshift mutations or split genes are the result of inexact DNA repair following excision of the transposon as it moved to a new location. In contrast to this study, genome comparisons of virulent and non-virulent Francisella tularensis showed that transposon mediated recombination and shuffling of gene order occurred in the pathogenic strains rather than the non-pathogenic strain [7], also they did not find deletions associated with transposon mediated recombination as seen in R. peacockii.
Figure 1. Alignment of Rickettsia rickettsii and Rickettsia peacockii genomes.

The alignment of the Rickettsia rickettsii SS and Rickettsia peacockii genomes using progressive Mauve with default parameters shows the lack of synteny between the genomes of these closely related organisms. The breakpoints of the syntenic blocks in R. peacockii are largely associated (31 of 37) with the ISRpe1 transposon, indicated with black arrows. The genome on top is that of R. rickettsii SS (reference genome) and that below is R. peacockii.
The transposon ISRpe1, originally identified in R. peacockii, [10] is also found twice in the R. massiliae genome along with a gene fragment (E = 0.0; RMA_0538 and RMA_0748) and a frameshifted mutant copy is found on the R. felis plasmid (E = 4e−154; RF_p48). The genomic locations for the copies of ISRpe1 in R. massiliae are different from those found in R. peacockii, indicating the transposition events occurred independently rather than in a common ancestor. Homologs of ISRpe1 are found three times (E = 5e−173; Aasi_0934, Aasi_0956 and Aasi_0884) in the genome of Candidatus Amoebophilus asiaticus, a member of the phylum Cytophaga-Flavobacterium-Bacteroides (CFB) and endosymbiont of Acanthamoeba [11]. The phylum also contains Candidatus Cardinium endosymbionts of arthropods. C. Cardinium spp. are related to C. A. asiaticus and closely related to one another, yet exist in a wide range of arthropod species [12]. This is indicative of horizontal transmission and could put these bacteria in contact with various species of rickettsiae. A phylogenetic tree, Figure 2, shows the close relationship between ISRpe1 and the Candidatus A. asiaticus transposon, but given the presence of this transposon in few rickettsiae, we suggest the transposon was transferred to these few rickettsiae in the recent past rather than from C. A. asiaticus. A possible source of the transposon is a C. Cardinium species from D. andersoni ticks that is similar to an endosymbiont cultured from Ixodes scapularis ticks [13]. To explore this link, genomic DNA from the cultured C. Cardinium spp. was used as template in a PCR reaction with ISRpe1 primers not previously used in the lab and the sequence of the product was determined. The DNA sequence of the 931 bp C. Cardinium PCR product shares 98% identity with the most homologous ISRpe1 copy in R. peacockii. The derived amino acid sequence was added to the phylogenetic tree shown in Figure 2. These results support the hypothesis that the transposon was transferred from a C. Cardinium species to a recent ancestor of R. peacockii.
Figure 2. Phylogenetic analysis of ISRpe1 transposon.

Neighbor joining (NJ) and maximum parsimony (MP) analyses included 14 taxa. Exclusion of gaps left 309 amino acids for the analyses; 105 amino acids were constant, 36 of the variable amino acids were parsimony uninformative and 168 of the variable amino acids were parsimony informative. Bootstrap analysis involved 2,000 replicates: top number is NJ bootstrap value and bottom number the MP bootstrap value. Genbank references for the proteins found in Text S7.
Tra Gene Cluster of R. peacockii
The three regions of R. bellii-like DNA in the chromosome are located at nucleotides 142335–149261 (one end of the tra cluster with genes TraB, TraE, leucine-rich protein gene and a TraV fragment), 806589–813116 (with degraded genes for a permease, TraA and TraD) and 497519–499744 (other end of the tra cluster with U gene). The phenomenon of lateral transfer of the tra cluster was first observed in the R. massiliae genome [14]. An ISRpe1 transposon is present at four of the six junctions of these three regions and the remaining two junctions are the tRNAVal gene and a chimeric tRNA gene. It was difficult to determine the junction at 149261 (the chimeric tRNA gene) as this area has been lost in R. rickettsii. This junction border was chosen due to blastn comparison with R. conorii that shows the highest homology (98%) upstream from nucleotide 149262 and no homology downstream. This correlates well with events of integration at tRNA genes by integrons such as the tra cluster [15]. The leucine-rich protein gene (RPR_00830) in the TraBE region is not found in R. massiliae and only shows homology (97%) to RBE_0439 of R. bellii which is near TraE, suggesting either strong selection for this gene sequence uniquely in R. peacockii and R. bellii or independent introduction of the tra cluster within the rickettsiae. Also, the permease pseudogene in the TraAD region of R. peacockii is not found in R. massiliae, but found in the tra cluster of R. canadensis (A1E_02610 and A1E_02615). In R. peacockii it appears the tra cluster integration preceded the arrival of the ISRpe1 transposon which then transposed into the tra cluster and split it into the three remaining regions by recombination and presumable deletion of the bulk of the tra cluster. The genome of R. rickettsii SS does not contain this tra cluster but may have a remnant 227 bp fragment located at nucleotides 7243640–724132 near the tRNAVal gene [14]. When this 227 bp region from R. peacockii is compared with other rickettsiae using blastn, R. rickettsii shows 68% identity (E = 6e−14) while R. bellii shows 88% identity (E = 1e−73) and R. massiliae shows 85% identity (E = 4e−67). If the 227 bp region of R. rickettsii shared ancestry with the tra cluster of closely related R. peacockii, one would expect the percent identity to R. rickettsii to be higher than that of R. bellii and R. massiliae.
Features of R. peacockii Plasmid
The 26 kb plasmid of R. peacockii (pRPR) contains 20 putative genes (Table 1), two of which are involved in plasmid maintenance and replication, ParA and DnaA. The ParA gene RPR_p01 and the two neighboring genes RPR_p02 and RPR_p03 are most closely related to RF_p23, RF_p22 and RF_p21 of R. felis and flanked by 56 bp inverted repeats, whereas the R. massiliae plasmid has an unrelated ParA gene. A phylogenetic tree was made comparing rickettsial plasmid borne parA proteins with their closest blast hits (Figure 3). It is interesting that the ParA genes on rickettsial plasmids fall into diverse groups suggesting foreign plasmids have periodically entered rickettsiae. ParA is the likely determinant of compatibility so entrance of a new parA gene enables a second plasmid to be maintained. The C-terminal domain of DnaA is similar to the plasmid-borne DnaA-like proteins of R. massiliae RMA_p01, R. felis RF_p05 and also the smaller version in R. felis RF_p19. The N-terminal domain is similar to the DnaA-like protein of R. monacensis. The plasmid contains five genes, RPR_p06 – RPR_p10, most closely related to orfs B, C, D, F, G found in a region termed the glycosylation island in Pseudomonas aeruginosa, shown to be involved (orfs A, N and E) in flagellar glycosylation [16]. The gene order is mostly maintained in this gene cluster on pRPR with only orf E deleted and orfB flipped between the repeats shown in the annotation. The GC content of this region is 48.4% vs. 34.7% for the entire plasmid and 32.6% for the R. peacockii chromosome, another indication beyond the homology for lateral transfer. The function of these five genes in R. peacockii is unknown but by homology they appear to be involved in phospholipid biosynthesis and may be maintained to increase the flow of glycerol-3-phosphate into the phospholipid biosynthesis pathway given that R. peacockii has a frameshift mutation in the glycerol-3-phosphate dehydrogenase gene (Table 2, location 96554..97530). This gene is necessary to make glycerol-3-phosphate from dihydroxyacetone phosphate (DHAP) in rickettsiae. Due to the absence of the glycolytic pathway, rickettsiae are unable to synthesize DHAP from fructose-1,6-diphosphate and must import it from the host cell [17]. Rickettsiae commonly have a glycerol-3-phosphate transporter as well as a DHAP transporter so can obtain glycerol-3-phosphate from the host cell directly or indirectly, while only R. peacockii has a mutant copy of the glycerol-3-phosphate dehydrogenase gene, so is dependent on import of glycerol-3-phosphate alone. This mutation in R. peacockii may limit the amount of glycerol-3-phosphate available for phospholipid biosynthesis and the presence of these 5 genes on the plasmid may alleviate this problem.
Table 1. Comparison of plasmid sequence to Genbank using blastP, or blastX for frameshifts and fragments.
| Location on pRPR | Top blast hit |
| RPR_p01 | Rickettsia felis RF_p23 Plasmid stability protein ParA2, E = 2e−92, Positives = 194/211 (91%) Not found in other rickettsiae. |
| RPR_p02 | Rickettsia felis RF_p22 Hypothetical protein, E = 3e−25, Positives = 69/82 (84%) |
| RPR_p03 | Rickettsia felis RF_p21 Hypothetical protein, E = 3e−29, Positives = 90/130 (69%) |
| RPR_p04 | Rickettsia massiliae RMA_p01 DnaA-like replication initiator protein E = 2e−123, Positives = 435/798 (54%) |
| 5154..6508 | Rickettsia bellii RBE_0057 Type I restriction-modification system methyltransferase subunit, pseudogene E = 0.0, Positives = 392/450 (87%) |
| RPR_p05 | Rickettsia massiliae RMA_0538 Transposase, E = 0.0, Positives = 360/364 (98%) |
| RPR_p06 | Pseudomonas aeruginosa PA2G_00065 UDP-3-O-[3-hydroxymyristoyl] glucosamine N-acyltransferase, E = 6e-72, Positives = 165/205 (80%) |
| RPR_p07 | Pseudomonas aeruginosa PA2G_00064 Phenylpropionate dioxygenase and related ring-hydroxylating dioxygenases, E = 3e−164, Positives = 315/375 (84%) |
| RPR_p08 | Pseudomonas aeruginosa PaerPA_01001561 (PA2G_00062) FabG, 3-ketoacyl-(acyl-carrier-protein) reductase, E = 6e−101, Positives = 221/255 (86%) |
| RPR_p09 | Pseudomonas aeruginosa PA2G_00061 FabH, 3-oxoacyl-[acyl-carrier-protein], E = 3e−158, Positives = 291/319 (91%) |
| RPR_p10 | Pseudomonas aeruginosa PA2G_00060 Acyl carrier protein, E = 1e−25, Positives = 65/72 (90%) |
| RPR_p11 | Candidatus Amoebophilus asiaticus Aasi_0982 ABC-type transport system, ATPase and permease components, E = 0.0, Positives = 442/575 (76%) |
| 14357..15476 | Rickettsia massiliae RMA_0746 Transposase, pseudogene E = 3e−140, Positives = 257/272 (94%) Not found in other rickettsiae. |
| RPR_p12 | Rickettsia monacensis RM-p7 Hsp1, Molecular chaperone (small heat shock protein), E = 1e−56, Positives = 139/176 (78%) |
| RPR_p13 | Rickettsia monacensis RM-p6 Hsp2, Molecular chaperone (small heat shock protein), E = 1e−73, Positives = 146/154 (94%) |
| RPR_p14 | Orientia tsutsugamushi OTT_1892 Transposase-like, E = 5e−104, Positives = 241/303 (79%) |
| RPR_p15 | Rickettsia massiliae RMA_0538 Transposase, E = 0.0, Positives = 358/364 (98%) |
| 19828..20073 | Wolbachia WD_0253 Transposase fragment, E = 1e−27, Positives = 66/77 (85%) |
| 20088..21168 | Rickettsia felis RF_p41 Transposase, pseudogene E = 3e−164, Positives = 330/356 (93%) |
| RPR_p16 | Erwinia tasmaniensis ETA_pET460420 Putative lipoprotein, E = 2e−13, Positives = 67/121 (55%) |
| RPR_p17 | Rickettsia bellii RBE_0152 SMR-type multi-drug efflux transporter, E = 2e−41, Positives = 94/103 (91%) Not found in other Rickettsiae. |
| RPR_p18 | Methanosarcina barkeri Mbar_A2604 TPR repeat-containing protein, E = 8e−13, Positives = 64/116 (55%) |
| RPR_p19 | First half of protein is fragment of Rickettsia conorii protein PS 120 (Sca4) E = 2e−68, Positives = 164/223 (73%). Second half of protein yields no homologies. Apparent chimeric protein. |
| RPR_p20 | Rickettsia akari A1C_03990 hypothetical protein, partial homology, E = 8e−92, Positives = 228/323 (70%) |
Figure 3. Maximum parsimony analysis of 27 parA proteins.

Exclusion of gaps left 166 amino acids for the analysis; 8 amino acids were constant, 2 of the variable amino acids were parsimony uninformative and 156 of the variable amino acids were parsimony informative. Bootstrap analysis involved 1,000 replicates: numbers at selected branches are the NJ (MP/NJ) bootstrap values that were ≥50%. The top two or three blastP hits to rickettsial plasmid parA's with E = >1e−30 were chosen for the analysis. The parA from the Trichoplax adhaerens genome project is likely from a bacterium associated with this simplest of eukaryotes as several contigs have homology to Rickettsiales. The parA proteins from the rickettsial endosymbiont of Ixodes scapularis (REIS) were added to the analysis following PCR and sequencing to confirm the presence of the genes in our REIS isolate (Baldridge et. al., in preparation). Maximum parsimony and neighbor joining (not shown) phylograms were congruent. Genbank references for the proteins found in Text S7.
Table 2. Nonsense mutations and split genes in R. peacockii relative to R. rickettsii Sheila Smith.
| Location of gene in R. peacockii | Gene and type of mutation |
| 10935..10988 and 151409..151545 | Small hypothetical gene A1G_03990 split by recombination |
| 15676..16998 | Methyltransferase A1G_03950, premature stop codon |
| around 188490 | Recombination near this point (219340 in SS) split hypothetical gene A1G_01175 |
| 29106..29608 | BioY family protein RrIowa_0811 (not annotated in SS), frameshift |
| 96554..97530 | NAD(P)H-dependent glycerol-3-phosphate dehydrogenase A1G_03470, frameshift |
| 186175..186749 | Cytochrome oxidase biogenesis protein A1G_00265, frameshift |
| 298139..299185 | AFG1-like ATPase A1G_01615, premature stop codon |
| 441508..442904 | NAD/NADP transhydrogenase beta subunit A1G_00635, N-terminal truncation of 82 AA's due to frameshift. |
| 462858..463848 | SAM-dependent methyltransferase domain A1G_00720, frameshifts, results in lack of TPR domain in RPR_02715 |
| 559562..560115 | (di)nucleoside polyphosphate hydrolase A1G_06270, frameshift and internal stop codon |
| 617196..618666 | Patatin b1 precursor A1G_05085, frameshift, C-terminal truncation but contains the complete patatin superfamily domain. |
| 630337..631282 | Hypothetical protein (conserved rickettsial) A1G_04725, frameshift |
| 884549..885426 | Acyltransferase COG1835 A1G_07015 (likely pseudogene), frameshift |
| 888608..892941 | OmpA A1G_06990, three frameshifts |
| 917962..918565 | Ankyrin repeat-containing protein A1G_04305, frameshift |
| 998131..999374 | AmpG A1G_03035, frameshift |
| 1007548..1009092 | Cytochrome c oxidase, subunit I, A1G_02985, frameshifts |
| 1139934..1140955 | Hypothetical protein A1G_02790, two frameshifts |
| 1171010..1172275 | Conserved hypothetical protein A1G_02605, two frameshifts |
| 1176591..1178159 | Putative phosphoethanolamine transferase A1G_02570, two frameshifts |
Also found on the plasmid are two small heat shock genes, one (RPR_p13) appears to be a common feature present on rickettsial plasmids [18]. RPR_p13 homologs are not represented on the chromosomes of other rickettsiae except R. felis (RF_1004) but this is an unusual case in that the other R. felis chromosomal copy (RF_1005) has a frameshift mutation and RF_1004 may have arisen via recombination with the plasmid copy. RPR_p12 is more similar to small heat shock proteins found on the chromosome of all rickettsiae but phylogenetic analysis shows this plasmid copy falls into a third group of rickettsial small heat shock proteins (Text S3). A comparison of the three small heat shock proteins of R. peacockii (including chromosomal copy RPR_2300) using Kyte-Doolittle plots shows the degree of N-terminal hydrophobicity varies from high to low between the three proteins with the two plasmid copies having the greatest difference (Text S4). In yeast the strength of this N-terminal hydrophobicity determines the strength of interaction of these chaperones with their target proteins [19], [20]. A family of these chaperones with a range of strengths of interaction may well help rickettsiae survive changing environmental temperatures during their life cycles in arthropods. It is also possible that individual target proteins or membranes benefit from a specialized chaperone. The plasmid contains genes for two transporters; RPR_p11 is an ABC type with ATPase and permease domains with strong homology (E = 0.0) to Aasi_0982 from Candidatus A. asiaticus and no homology to known rickettsial genes. The second transporter RPR_p17 is an SMR-type multi-drug efflux transporter and the only other rickettsia to have a homolog of RPR_p17 is R. bellii, while next closest relatives are in CFB group of bacteria. Other genes on the plasmid code for various transposases, a putative lipoprotein, a TPR repeat-containing protein and an apparent chimeric protein. All the plasmid genes with rickettsial chromosomal homologs have far lower homology to those of R. peacockii or R. rickettsii and higher homology to other more distantly related rickettsiae, which is an indication of horizontal gene transfer to the plasmid, as seen in pRF of R. felis [21]. The exceptions are the two ISRpe1 transposon copies on the plasmid.
Deletions and Mutations in R. peacockii vs. R. rickettsii Strain Sheila Smith
Deletions in R. peacockii vs R. rickettsii SS are shown in Table 3. Deletions greater than 100 bases were examined as well as smaller deletions that disrupted genes or were within 5 bases of ISRpe1 transposons. Nonsense mutations resulting from premature stop codons and small deletions or insertions causing frameshifts in R. peacockii vs R. rickettsii SS are shown in Table 2. It appears that some deletions and mutations in R. peacockii may be responsible for its lack of pathogenicity and are focused upon in this section. Possible candidate genes include those coding for an ankyrin repeat containing protein, DsbA, RickA, Protease II, OmpA, Sca1, and a putative phosphoethanolamine transferase. The deletion located at SS coordinates 869412..871928 (Table 3) was likely deleted in R. peacockii during recombination between ISRpe1 transposons and contains a gene coding for one of the two larger ankyrin repeat containing proteins in R. rickettsii SS (A1G_05165). Ankyrin repeat proteins have been shown to be effector proteins or virulence factors in several pathogens [22]. In another member of the order Rickettsiales, AnkA is rapidly translocated to the host cell and phosphorylated by host cell kinases upon Anaplasma phagocytophilum binding to the host cell [23]. AnkA also binds DNA and alters transcription of defense related genes in HL-60 cells infected with A. phagocytophilum or transfected with an AnkA expression plasmid [24]. Transcript levels for ankA (APH_0740) were shown to be 2.3 to 3 fold higher in A. phagocytophilum grown in mammalian cells versus tick cells [25] and in R. rickettsii transcript levels of A1G_05165 were 3.2 fold higher in mammalian cells versus tick cells as well as being one of few differentially expressed genes detected [26]. Moreover, this gene that is deleted in R. peacockii is mutated in R. rickettsii Iowa, which has attenuated virulence compared to R. rickettsii SS [6]. The deletion in the Iowa gene (RrIowa_1113) removes three of the four ankyrin repeats from the protein. Strengthening the case for this as a virulence factor is the observation that this gene has been deleted in Rickettsia monacensis (R. Felsheim, unpublished) and is not found in R. bellii, both non-pathogenic for humans. Recently the putative genome sequence from the non-pathogenic rickettsial endosymbiont of Ixodes scapularis (REIS) has been released and this ank gene also appears to have been deleted in a similar manner to that seen in R. monacensis. A remnant of the gene found at nucleotides 53520–53589 of REIS contig ACLC01000066.1 are homologous to the 3′ end of the ank gene. There is about a 3.7 kb deletion in this region of the REIS genome compared to R. rickettsii. All of the rickettsial pathogens for which genome sequence is available have this gene, except Rickettsia akari.
Table 3. Deletions in R. peacockii relative to R. rickettsii Sheila Smith.
| Location in R. rickettsii SS | Size of deletion | Genes or gene products contained in deletion |
| 14498..15082 | 583 | A1G_00085 truncated, antitoxin A1G_00090 lost and A1G_00095 truncated |
| 19653..25701 | 6047 | A1G_00130, Cell surface antigen Sca1-like |
| 36902..37126 | 223 | A1G_00215, Part of dihydrofolate reductase gene |
| 48049..52257 | 4207 | Region of gene fragments resulting from gene reduction |
| 219917..220001 | 83 | A1G_01175, 3′ end of hypothetical gene, also split by rearrangement at bp 219337 |
| 231815..231898 | 82 | A1G_01245, radical SAM family enzyme gene, results in frameshift |
| 232280..232351 | 70 | A1G_01245, 3′ end of radical SAM family enzyme gene |
| 258009..266392 | 8382 | Gene fragments for penicillin binding protein and fragments of Sca8 resulting from gene reduction |
| 333287..333616 | 328 | A1G_01880, hypothetical gene |
| 372621..372708 | 86 | Small intergenic deletion |
| 381886..381961 | 74 | A1G_02165, Protease II, results in frameshift |
| 444166..444318 | 151 | A1G_02530, small hypothetical gene |
| 494539..498554 | 4014 | A1G_02820, 02825, 02830, ABC transporter component genes |
| 506267..506762 | 494 | Gene fragments resulting from gene reduction |
| 561101..563976 | 2874 | Gene fragments of RND efflux transporter gene |
| 588183..589234 | 1050 | A1G_03355, Protein-disulfide isomerase, DsbA gene |
| 614711..615355 | 643 | Missing part of A1G_03530 at point of transposon insertion |
| 619349..619541 | 191 | Small intergenic deletion |
| 687771..689062 | 1290 | Gene fragments resulting from gene reduction |
| 691605..692087 | 481 | A1G_04035 hypothetical gene |
| 708170..708343 | 172 | Small intergenic deletion |
| 715158..716455 | 1296 | A1G_04170 hypothetical gene |
| 723952..728058 | 4105 | Region of gene fragments resulting from gene reduction |
| 730884..731135 | 250 | A1G_4290, Gene fragment |
| 741167..741466 | 298 | A1G_4355, Gene fragment |
| 742387..742859 | 471 | A1G_4365, Gene fragment |
| 747947..753101 | 5153 | Region of gene fragments resulting from gene reduction |
| 780788..783183 | 2394 | A1G_04605, YhbC and A1G_04620, transcriptional regulator rirA and gene fragments resulting from gene reduction |
| 787638..788105 | 466 | A1G_04660 truncated (S4 ribosomal and related proteins) |
| 807386..807708 | 321 | A1G_04775 hypothetical |
| 813412..813703 | 290 | Gene fragments resulting from gene reduction |
| 839333..839525 | 191 | A1G_04970 truncated, hypothetical gene |
| 841975..846686 | 4710 | A1G_05015 RickA gene, A1G_04995 to A1G_05010 Succinyl-CoA:3-ketoacid-coenzyme A transferase subunits A and B and gene fragments |
| 856816..859634 | 2817 | Gene fragments resulting from gene reduction |
| 869412..871928 | 2515 | A1G_05165 ankyrin repeat protein gene, also mutated in R. rickettsii Iowa (RrIowa_1113) |
| 913278..917419 | 4140 | Region of gene fragments resulting from gene reductions |
| 975118..975936 | 817 | A1G_05855, two copies of LPS biosynthesis protein gene recombined into one |
| 1000150..1000436 | 285 | Gene fragments resulting from gene reduction |
| 1139429..1140228 | 798 | Gene fragments of COG2602 Beta-lactamase class D |
| 1145752..1146794 | 1041 | Gene fragment resulting from gene reduction |
While most bacteria have a single DsbA gene, R. rickettsii SS and other rickettsiae have two DsbA genes, one (A1G_03355) is deleted in R. peacockii due to recombination between two ISRpe1 transposons (Table 3, location 588183..589234). DsbA codes for a protein-disulfide oxidoreductase which catalyzes disulfide-bond formation in the periplasm during the folding of secreted proteins. Both rickettsial DsbA proteins are predicted to be anchored into the membrane, one via a transmembrane domain and one as a lipoprotein. In pathogenic Vibrio cholerae, the DsbA homolog (TcpG) is responsible for the folding, maturation and secretion of virulence factors [27]. The importance of DsbA for virulence has been demonstrated in a variety of organisms [28], [29], [30], [31], [32], [33], [34]. Compensation for this deletion by the other copy of DsbA is likely for some functions, but in Neisseria meningitides which has three DsbA homologs, they vary in functional activity in complementation assays [35].
RickA was previously shown to be truncated by the ISRpe1 transposon in R. peacockii and interaction of R. peacockii with actin was found to be lacking [10]. We show that most of the RickA gene has been deleted along with the neighboring succinyl-CoA:3-ketoacid-coenzyme A transferase genes during recombination between transposons (Table 3, location 841975..846686). Actin based motility is thought to mediate intracellular and cell-to-cell movement of rickettsiae. Time-lapse photography of our GFP expressing R. peacockii shows them to be non-motile compared to other rickettsiae observed (unpublished data). The protease II gene in R. peacockii (Table 3, location 381886..381961) has a 74 bp deletion causing a frameshift in the middle of the gene. Proteases have been shown to be important for binding and entry in a wide variety of organisms and protease II is an S9A type protease, the type shown to be necessary for entry of Trypanosoma cruzi into host cells [36]. The three frameshift mutations in the OmpA gene of R. peacockii have previously been discussed [37]. In the avirulent strain R. rickettsia Iowa, OmpA is also truncated as a result of a frameshift mutation [6] that differs from those in the R. peacockii OmpA gene. Sca1, which is in the same superfamily of autotransported surface proteins as OmpA, is deleted in R. peacockii via transposon insertion upstream of the Sca1 gene and also near the stop codon (Table 3, location 19653..25701), followed by recombination and deletion of a 6kb DNA fragment containing the Sca1 gene. Sca1 is present in all other Rickettsia spp. and the type of selection pressure on the N-terminal passenger domain implicates the N-terminal domain of this autotransported protein in interactions with the host cell [38]. Outer membrane proteins implicated in binding of rickettsiae to host cells, include OmpA [39], OmpB [40], RP828, RC1281, and the autotransporter domain of OmpB [41]. Of these only OmpA is defective in R. peacockii. It is possible that rickettsiae use different members of this Omp family to bind to different host cell types or cells of different species.
Since the N-terminal domain of OmpA-B family members is likely extended from the membrane through the slime layer, it is possible that proper configuration of the surface lipooligosaccharides is important for proper arrangement of these surface proteins. While the slime layer is characteristic of the SFG rickettsiae, in R. peacockii the slime layer is thin and not always discernable [42]. The increase in the thickness of the slime layer in R. rickettsii upon tick feeding correlates with the restoration of virulence [43]. Genes for a sugar reductase and sugar epimerases including CapD, predicted to be involved in slime layer biosynthesis [44] are located very close to a mutant putative phosphoethanolamine transferase gene in R. peacockii. The putative phosphoethanolamine transferase gene is found between nucleotides 1176591..1178159 (Table 2) with two frameshift mutations close together that introduce a stop codon truncating the transferase domain. This enzyme is required for the correct structure of surface lipooligosaccharides of Neisseria meningitides and mutation of phosphoethanolamine transferase decreases bacterial binding to endothelial cells 10 fold [45], [46]. Our experience with R. peacockii in culture is that it binds very poorly to host cells and extracellular R. peacockii are observed more abundantly in vitro than other rickettsial species maintained in our laboratory [47]. The protein sequences of phosphoethanolamine transferases are not well conserved among bacteria except around the transferase domain, which is where significant homology exists to this rickettsial protein. This putative phosphoethanolamine transferase shares the same 5-transmembrane structure with others as well (Text S5). The locus tag in R. rickettsii SS is A1G_02570 and closely related genes are found in all other Rickettsia spp.
Another deletion via recombination between transposons removes A1G_04605 (YhbC) and A1G_04620 (transcriptional regulator of the RirA / Rrf-2 superfamily). YhbC was picked up in a mutant screen for virulence factors of Salmonella enteritidis due to its effect on the growth rate of the bacteria, making the mutant a potential live vaccine candidate [48]. The growth rate of R. peacockii in culture is slower than other rickettsiae grown in our lab. Transcription factors of the rirA type are repressors containing an iron-sulfur cluster, and thus can sense iron concentrations as well as nitric oxide which dissociates the cluster and alters DNA binding. The lack of iron or presence of nitric oxide leads to derepression of genes regulated by rirA, so lack of rirA protein results in an increase in expression of these regulated genes [49], [50] [51]. In the rirA mutant Sinorhizobium meliloti a toxic amount of iron builds up and leads to a hypersensitivity to H2O2 [52]. R. peacockii are not found in hemocytes while R. rickettsii are commonly found in hemocytes and this deletion of rirA may contribute to this observation, given that it has been shown that reactive oxygen species are produced in cattle tick hemocytes [53] and presumably in other tick species as well.
R. peacockii also has other deletions and mutations, notably genes that are conserved in other rickettsiae like the methyltransferase A1G_03950 (Table 2) and hypothetical gene A1G_03530 (Table 3). Nonsense mutations in R. rickettsii SS vs R. peacockii are shown in Text S6. DNA sequence found in R. peacockii and not in R. rickettsii SS is shown in Text S1 and includes mainly the ISRpe1 transposons, the three fragments of the tra cluster, the 10.5 kb fragment present in R. rickettsii Iowa vs. R. rickettsii SS [6] and a tandem gene duplication of A1G_02330 (RPR_04375 and RPR_04376).
Our results support the speculation that in the past, a virulent SFG rickettsia underwent changes to become the East Side Agent (Rickettsia peacockii) [2]. Gene reduction in rickettsiae and some other bacteria correlates with an increase in virulence [54], [55], [56] but our analysis of gene loss in R. peacockii suggests that transposon mediated gene reduction is responsible for avirulence in this case.
R. peacockii has a dynamic genome that has been and is likely still being shaped by ISRpe1 activity. The changes have resulted in a dramatic lack of synteny with R. rickettsii and likely contributed to rendering it non-pathogenic for vertebrates, restricting it to the tick host. R. peacockii has lost several genes that appear important in the transmission of pathogenic rickettsiae to a vertebrate host. At the same time it has retained a gene repertoire that enables it to survive and grow in the tick and to be transmitted transovarially to the tick's progeny. The extensive remodeling of the genome makes reversion to pathogenicity unlikely unless new virulence genes are imported. Ticks encounter and interact with many bacteria during their life cycle, some of which can invade the ovaries and cohabit the same cell (e.g. the Francisella-like D. andersoni symbiont and C. Cardinium spp.). We propose that symbionts such as R. peacockii could conceivably acquire novel genes via lateral gene transfer through their interactions with a range of bacteria including pathogens acquired by the tick during its blood meal [57]. Ticks have mechanisms for excluding foreign bacteria during the internalization of the blood meal but feeding on a heavily infected mammal may provide a challenge to this system. The acquisition of DNA from P. aeruginosa onto the R. peacockii plasmid may relate to the fact that ticks absorb cells and large macromolecules intracellularly for digestion [57] and P. aeruginosa is known to secrete large amounts of genomic DNA [58]. R. peacockii and its acquisition of mobile DNA is a good example of the ‘intracellular arena’ hypothesis at work, in that obligate intracellular bacteria more readily share genetic material if they cohabit the same cells [59]. Obligate intracellular bacteria like rickettsiae that live in arthropods which feed on mammals also increase their rate of exposure to novel gene pools [60]. We see the plasmid as the only recent recipient of foreign DNA, other than the ISRpe1 transposon, in the genome of R. peacockii.
Materials and Methods
Rickettsia peacockii Rustic [42] was grown in Ixodes scapularis cell line ISE6 [47], [61] for eight in vitro passages. Genomic DNA was prepared from rickettsiae released from infected cells by forcing suspended cells five times through a 25 G needle attached to a 5 ml syringe. The resulting lysate was centrifuged at 270 rcf for 5 min to remove whole cells and the supernatant filtered through a 1.2 µm syringe filter (Whatman Puradisc FP30; Sigma-Aldrich St. Louis, MO). Rickettsiae were recovered from the filtrate by centrifugation (18,400 rcf, 5 min 4°C), resuspended in Dulbecco's Phosphate Buffered Saline (PBS) containing calcium and magnesium (Mediatech, Inc. Herndon, VA) and DNase I (15 µg/ml; from bovine pancreas Type II-S, Sigma-Aldrich), and incubated at room temperature for 30 min. After DNase I treatment to remove contaminating Ixodes DNA rickettsiae were centrifuged again (18,400 rcf, 5 min, 4°C) and genomic DNA was prepared using the Puregene kit (Gentra Systems, Minneapolis, MN) following the protocol for Gram negative bacteria. The C. Cardinium spp. isolate [13] was grown and DNA isolated in the same manner as above.
DNA was pyrosequenced on a 454FLX machine (454-Roche, Branford, CT) (226,040 reads, >30X coverage) at the BioMedical Genomics Center, U of M, St. Paul, MN and assembled using Newbler (454-Roche) requiring 99% homology; 56 contigs 500 bases or larger were obtained. To determine if transposons occupied the gaps, 50–100 basepairs from each end of the ISRpe1 transposon were used to recover 454 traces using Blastn and assembled using Sequencher (Gene Codes, Ann Arbor, MI) requiring 100% homology. These contigs were then assembled onto the ends of the original 454 generated contigs. The ISRpe1 transposons are too similar to one another to be assembled from 454 traces and mapping the contigs to the R. rickettsii genome followed by PCR across the gaps yielded artifactual results, again due to the similarity between individual transposons and their interaction during PCR. Therefore, contigs were extended using GenomeWalker (Clontech, Mountain View, CA) ligation mediated PCR using a single gene specific primer for each contig end. (nested PCR was unnecessary). Four GenomeWalker libraries were made using EcoRV, HaeIII, PvuII and HpaI which do not cut in the transposon. The last gap could not be filled this way, nor with standard PCR and it appeared that two transposons were present here. All contigs from the 454 assembly were then assembled onto the linear genome contig, and with the exception of contigs from Ixodes scapularis mitochondrial and genome sequence, only one remained unassembled. This contig contained the junction of a transposon and a transposon fragment. This junction sequence was used to retrieve 454 traces using blastn that were then assembled using Sequencher, requiring 100% homology. Primers were designed to bind to unique sequence within this junction for use in PCR with gene specific primers from the ends of the large contig. A primer was designed from a region of the transposon not found in the transposon fragment to confirm the sequence at the junction. The GenomeWalker PCR products were sequenced with transposon specific primers. AccuTaq DNA polymerase (Sigma-Aldrich) was used throughout. The coverage layout along the contigs was calculated with 454 de novo assembler software (version 2.0.00.20) using the derived file 454AlignmentInfo.tsv in 100 nucleotide scale and visually scanned for anomalies. One contig had twice the normal number of traces per unit contig length and this region was investigated with PCR and was found to contain a gene duplication (RPR_04375 and RPR_04376) that was originally assembled into one gene. The results are a circular chromosome of 1,288,492 bp and a circular plasmid of 26,406 bp.
Annotation and Analysis
The genome was annotated using PGAAP at NCBI (http://www.ncbi.nlm.nih.gov/genomes/static/Pipeline.html). Apparent frameshifts were examined manually by recovering 454 traces from each area using Blastn and assembling them using Sequencher requiring 90% homology to determine the validity of the sequence. In ten of the areas PCR and sequencing was carried out to validate the sequence. The sequence was subjected to manual annotation by viewing the .gbf file using Artemis [9] and editing of the .sqn file. Gene fragments from apparent gene reduction auto-annotated as orfs by PGAAP were extended by blast analysis and re-annotated as misc_features (152) or removed.
Artemis Comparison Tool (ACT) (The Sanger Institute, Cambridge, UK) was used to compare the R. peacockii genome to that of R. rickettsii SS. Unique DNA sequence of each was extracted as a text file and examined using blast analysis. The level of synteny (or lack thereof) between the two genomes was examined by using Mauve [8] (Figure 1), ACT and a dotplot comparison (Figure S1). Mauve and Artemis were used to determine where the presence of a transposon coincided with a change in synteny between R. peacockii and R. rickettsii SS. Blastn and the backbone file from Mauve were used to examine all deletions in R. peacockii over 100 bp in size, to find which deletion junctions in R. peacockii were found within 5 bp of an ISRpe1 transposon (Text S2).
Supporting Information
A dot plot comparison of the R. peacockii and R. rickettsii genomes.
(0.01 MB PNG)
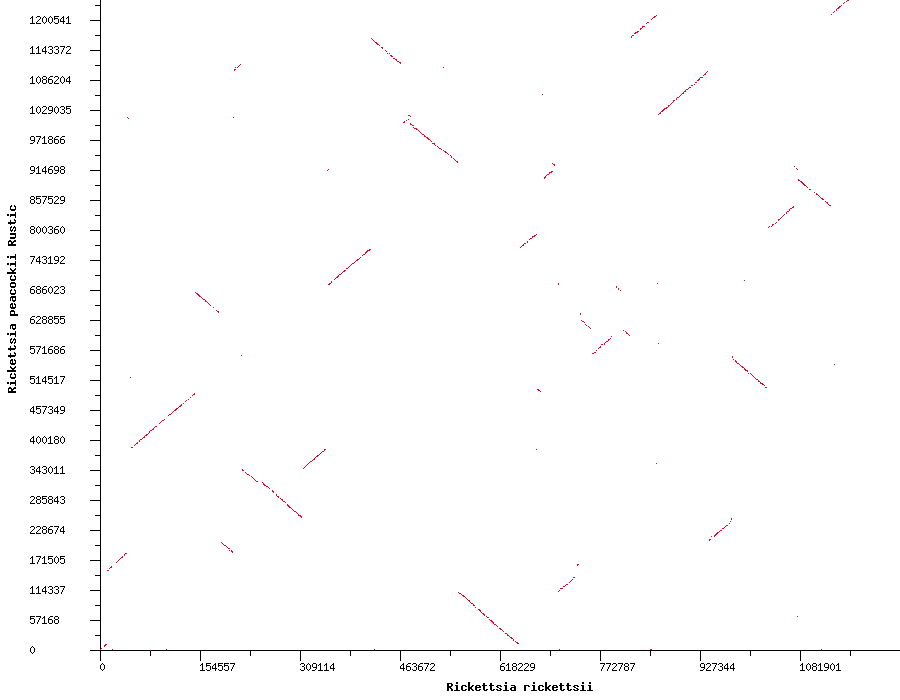{kind=link}
DNA sequence found in R. peacockii and not in R. rickettsii SS.
(0.07 MB DOC)
Deletions in R. peacockii and their association with ISRpe1 transposons.
(0.26 MB DOC)
Phylogenetic analysis of the rickettsial small hsp proteins.
(0.15 MB DOC)
Kyte-Doolittle plots of hydrophobicity for the three small hsp/chaperone proteins from R. peacockii.
(0.04 MB DOC)
Support for R. rickettsii A1G_02570 as phosphoethanolamine transferase.
(0.05 MB DOC)
Nonsense mutations in R. rickettsii Sheila Smith relative to R. peacockii.
(0.04 MB DOC)
Acknowledgments
The authors would like to thank Zheng Jin Tu at the University of Minnesota Supercomputing Institute for help with UNIX.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: This research was supported by National Institutes of Health (NIH) grant 2RO1 AI49424-06A2 to UGM. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Niebylski ML, Schrumpf ME, Burgdorfer W, Fischer ER, Gage KL, et al. Rickettsia peacockii sp. nov., a new species infecting wood ticks, Dermacentor andersoni, in western Montana. Int J Syst Bacteriol. 1997;47:446–452. doi: 10.1099/00207713-47-2-446. [DOI] [PubMed] [Google Scholar]
- 2.Burgdorfer W, Hayes SF, Mavros AJ. Nonpathogenic rickettsiae in Dermacentor andersoni: a limiting factor for the distribution of Rickettsia rickettsii. In: Burgdorfer W, Anacker RL, editors. Rickettsiae and rickettsial diseases. New York, N.Y.: Academic Press; 1981. pp. 585–594. [Google Scholar]
- 3.Philip RN, Casper EA. Serotypes of spotted fever group rickettsiae isolated from Dermacentor andersoni (Stiles) ticks in western Montana. Am J Trop Med Hyg. 1981;30:230–238. doi: 10.4269/ajtmh.1981.30.230. [DOI] [PubMed] [Google Scholar]
- 4.Niebylski ML, Peacock MG, Schwan TG. Lethal effect of Rickettsia rickettsii on its tick vector (Dermacentor andersoni). Appl Environ Microbiol. 1999;65:773–778. doi: 10.1128/aem.65.2.773-778.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dergousoff SJ, Gajadhar AJ, Chilton NB. Prevalence of Rickettsia in Canadian populations of the ticks Dermacentor andersoni and D. variabilis. Appl Environ Microbiol. 2009 doi: 10.1128/AEM.02554-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ellison DW, Clark TR, Sturdevant DE, Virtaneva K, Porcella SF, et al. Genomic comparison of virulent Rickettsia rickettsii Sheila Smith and avirulent Rickettsia rickettsii Iowa. Infect Immun. 2008;76:542–550. doi: 10.1128/IAI.00952-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rohmer L, Fong C, Abmayr S, Wasnick M, Larson Freeman TJ, et al. Comparison of Francisella tularensis genomes reveals evolutionary events associated with the emergence of human pathogenic strains. Genome Biol. 2007;8:R102. doi: 10.1186/gb-2007-8-6-r102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Darling AC, Mau B, Blattner FR, Perna NT. Mauve: multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004;14:1394–1403. doi: 10.1101/gr.2289704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rutherford K, Parkhill J, Crook J, Horsnell T, Rice P, et al. Artemis: sequence visualization and annotation. Bioinformatics. 2000;16:944–945. doi: 10.1093/bioinformatics/16.10.944. [DOI] [PubMed] [Google Scholar]
- 10.Simser JA, Rahman MS, Dreher-Lesnick SM, Azad AF. A novel and naturally occurring transposon, ISRpe1 in the Rickettsia peacockii genome disrupting the rickA gene involved in actin-based motility. Mol Microbiol. 2005;58:71–79. doi: 10.1111/j.1365-2958.2005.04806.x. [DOI] [PubMed] [Google Scholar]
- 11.Horn M, Harzenetter MD, Linner T, Schmid EN, Muller KD, et al. Members of the Cytophaga-Flavobacterium-Bacteroides phylum as intracellular bacteria of acanthamoebae: proposal of ‘Candidatus Amoebophilus asiaticus’. Environ Microbiol. 2001;3:440–449. doi: 10.1046/j.1462-2920.2001.00210.x. [DOI] [PubMed] [Google Scholar]
- 12.Zchori-Fein E, Perlman SJ. Distribution of the bacterial symbiont Cardinium in arthropods. Mol Ecol. 2004;13:2009–2016. doi: 10.1111/j.1365-294X.2004.02203.x. [DOI] [PubMed] [Google Scholar]
- 13.Kurtti TJ, Munderloh UG, Andreadis TG, Magnarelli LA, Mather TN. Tick cell culture isolation of an intracellular prokaryote from the tick Ixodes scapularis. J Invertebr Pathol. 1996;67:318–321. doi: 10.1006/jipa.1996.0050. [DOI] [PubMed] [Google Scholar]
- 14.Blanc G, Ogata H, Robert C, Audic S, Claverie JM, et al. Lateral gene transfer between obligate intracellular bacteria: evidence from the Rickettsia massiliae genome. Genome Res. 2007;17:1657–1664. doi: 10.1101/gr.6742107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Williams KP. Integration sites for genetic elements in prokaryotic tRNA and tmRNA genes: sublocation preference of integrase subfamilies. Nucleic Acids Res. 2002;30:866–875. doi: 10.1093/nar/30.4.866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Arora SK, Bangera M, Lory S, Ramphal R. A genomic island in Pseudomonas aeruginosa carries the determinants of flagellin glycosylation. Proc Natl Acad Sci U S A. 2001;98:9342–9347. doi: 10.1073/pnas.161249198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fuxelius HH, Darby A, Min CK, Cho NH, Andersson SG. The genomic and metabolic diversity of Rickettsia. Res Microbiol. 2007;158:745–753. doi: 10.1016/j.resmic.2007.09.008. [DOI] [PubMed] [Google Scholar]
- 18.Baldridge GD, Burkhardt NY, Felsheim RF, Kurtti TJ, Munderloh UG. Plasmids of the pRM/pRF family occur in diverse Rickettsia species. Appl Environ Microbiol. 2008;74:645–652. doi: 10.1128/AEM.02262-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Haslbeck M, Ignatiou A, Saibil H, Helmich S, Frenzl E, et al. A domain in the N-terminal part of Hsp26 is essential for chaperone function and oligomerization. J Mol Biol. 2004;343:445–455. doi: 10.1016/j.jmb.2004.08.048. [DOI] [PubMed] [Google Scholar]
- 20.Stromer T, Fischer E, Richter K, Haslbeck M, Buchner J. Analysis of the regulation of the molecular chaperone Hsp26 by temperature-induced dissociation: the N-terminal domail is important for oligomer assembly and the binding of unfolding proteins. J Biol Chem. 2004;279:11222–11228. doi: 10.1074/jbc.M310149200. [DOI] [PubMed] [Google Scholar]
- 21.Gillespie JJ, Beier MS, Rahman MS, Ammerman NC, Shallom JM, et al. Plasmids and rickettsial evolution: insight from Rickettsia felis. PLoS ONE. 2007;2:e266. doi: 10.1371/journal.pone.0000266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Pan X, Luhrmann A, Satoh A, Laskowski-Arce MA, Roy CR. Ankyrin repeat proteins comprise a diverse family of bacterial type IV effectors. Science. 2008;320:1651–1654. doi: 10.1126/science.1158160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.IJdo JW, Carlson AC, Kennedy EL. Anaplasma phagocytophilum AnkA is tyrosine-phosphorylated at EPIYA motifs and recruits SHP-1 during early infection. Cell Microbiol. 2007;9:1284–1296. doi: 10.1111/j.1462-5822.2006.00871.x. [DOI] [PubMed] [Google Scholar]
- 24.Garcia-Garcia JC, Rennoll-Bankert KE, Pelly S, Milstone AM, Dumler JS. Silencing of Host Cell CYBB Gene Expression by the Nuclear Effector AnkA of the Intracellular Pathogen Anaplasma phagocytophilum. Infect Immun. 2009 doi: 10.1128/IAI.00023-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nelson CM, Herron MJ, Felsheim RF, Schloeder BR, Grindle SM, et al. Whole genome transcription profiling of Anaplasma phagocytophilum in human and tick host cells by tiling array analysis. BMC Genomics. 2008;9:364. doi: 10.1186/1471-2164-9-364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ellison DW, Clark TR, Sturdevant DE, Virtaneva K, Hackstadt T. Limited transcriptional responses of Rickettsia rickettsii exposed to environmental stimuli. PLoS One. 2009;4:e5612. doi: 10.1371/journal.pone.0005612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Peek JA, Taylor RK. Characterization of a periplasmic thiol:disulfide interchange protein required for the functional maturation of secreted virulence factors of Vibrio cholerae. Proc Natl Acad Sci U S A. 1992;89:6210–6214. doi: 10.1073/pnas.89.13.6210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Coulthurst SJ, Lilley KS, Hedley PE, Liu H, Toth IK, et al. DsbA plays a critical and multifaceted role in the production of secreted virulence factors by the phytopathogen Erwinia carotovora subsp. atroseptica. J Biol Chem. 2008;283:23739–23753. doi: 10.1074/jbc.M801829200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ha UH, Wang Y, Jin S. DsbA of Pseudomonas aeruginosa is essential for multiple virulence factors. Infect Immun. 2003;71:1590–1595. doi: 10.1128/IAI.71.3.1590-1595.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lee Y, Kim Y, Yeom S, Kim S, Park S, et al. The role of disulfide bond isomerase A (DsbA) of Escherichia coli O157:H7 in biofilm formation and virulence. FEMS Microbiol Lett. 2008;278:213–222. doi: 10.1111/j.1574-6968.2007.00993.x. [DOI] [PubMed] [Google Scholar]
- 31.Rosadini CV, Wong SM, Akerley BJ. The periplasmic disulfide oxidoreductase DsbA contributes to Haemophilus influenzae pathogenesis. Infect Immun. 2008;76:1498–1508. doi: 10.1128/IAI.01378-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yu J. Inactivation of DsbA, but not DsbC and DsbD, affects the intracellular survival and virulence of Shigella flexneri. Infect Immun. 1998;66:3909–3917. doi: 10.1128/iai.66.8.3909-3917.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yu J, Kroll JS. DsbA: a protein-folding catalyst contributing to bacterial virulence. Microbes Infect. 1999;1:1221–1228. doi: 10.1016/s1286-4579(99)00239-7. [DOI] [PubMed] [Google Scholar]
- 34.Yu J, Oragui EE, Stephens A, Kroll JS, Venkatesan MM. Inactivation of DsbA alters the behaviour of Shigella flexneri towards murine and human-derived macrophage-like cells. FEMS Microbiol Lett. 2001;204:81–88. doi: 10.1111/j.1574-6968.2001.tb10867.x. [DOI] [PubMed] [Google Scholar]
- 35.Sinha S, Langford PR, Kroll JS. Functional diversity of three different DsbA proteins from Neisseria meningitidis. Microbiology. 2004;150:2993–3000. doi: 10.1099/mic.0.27216-0. [DOI] [PubMed] [Google Scholar]
- 36.Bastos IM, Grellier P, Martins NF, Cadavid-Restrepo G, de Souza-Ault MR, et al. Molecular, functional and structural properties of the prolyl oligopeptidase of Trypanosoma cruzi (POP Tc80), which is required for parasite entry into mammalian cells. Biochem J. 2005;388:29–38. doi: 10.1042/BJ20041049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Baldridge GD, Burkhardt NY, Simser JA, Kurtti TJ, Munderloh UG. Sequence and expression analysis of the ompA gene of Rickettsia peacockii, an endosymbiont of the Rocky Mountain wood tick, Dermacentor andersoni. Appl Environ Microbiol. 2004;70:6628–6636. doi: 10.1128/AEM.70.11.6628-6636.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ngwamidiba M, Blanc G, Raoult D, Fournier PE. Sca1, a previously undescribed paralog from autotransporter protein-encoding genes in Rickettsia species. BMC Microbiol. 2006;6:12. doi: 10.1186/1471-2180-6-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Li H, Walker DH. rOmpA is a critical protein for the adhesion of Rickettsia rickettsii to host cells. Microb Pathog. 1998;24:289–298. doi: 10.1006/mpat.1997.0197. [DOI] [PubMed] [Google Scholar]
- 40.Uchiyama T, Kawano H, Kusuhara Y. The major outer membrane protein rOmpB of spotted fever group rickettsiae functions in the rickettsial adherence to and invasion of Vero cells. Microbes Infect. 2006;8:801–809. doi: 10.1016/j.micinf.2005.10.003. [DOI] [PubMed] [Google Scholar]
- 41.Renesto P, Samson L, Ogata H, Azza S, Fourquet P, et al. Identification of two putative rickettsial adhesins by proteomic analysis. Res Microbiol. 2006;157:605–612. doi: 10.1016/j.resmic.2006.02.002. [DOI] [PubMed] [Google Scholar]
- 42.Simser JA, Palmer AT, Munderloh UG, Kurtti TJ. Isolation of a spotted fever group Rickettsia, Rickettsia peacockii, in a Rocky Mountain wood tick, Dermacentor andersoni, cell line. Appl Environ Microbiol. 2001;67:546–552. doi: 10.1128/AEM.67.2.546-552.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hayes SF, Burgdorfer W. Reactivation of Rickettsia rickettsii in Dermacentor andersoni ticks: an ultrastructural analysis. Infect Immun. 1982;37:779–785. doi: 10.1128/iai.37.2.779-785.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Santhanagopalan V, Coker C, Radulovic S. Characterization of RP 333, a gene encoding CapD of Rickettsia prowazekii with UDP-glucose 4-epimerase activity. Gene. 2006;369:119–125. doi: 10.1016/j.gene.2005.10.030. [DOI] [PubMed] [Google Scholar]
- 45.Takahashi H, Carlson RW, Muszynski A, Choudhury B, Kim KS, et al. Modification of lipooligosaccharide with phosphoethanolamine by LptA in Neisseria meningitidis enhances meningococcal adhesion to human endothelial and epithelial cells. Infect Immun. 2008;76:5777–5789. doi: 10.1128/IAI.00676-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Cox AD, Wright JC, Li J, Hood DW, Moxon ER, et al. Phosphorylation of the lipid A region of meningococcal lipopolysaccharide: identification of a family of transferases that add phosphoethanolamine to lipopolysaccharide. J Bacteriol. 2003;185:3270–3277. doi: 10.1128/JB.185.11.3270-3277.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kurtti TJ, Simser JA, Baldridge GD, Palmer AT, Munderloh UG. Factors influencing in vitro infectivity and growth of Rickettsia peacockii (Rickettsiales: Rickettsiaceae), an endosymbiont of the Rocky Mountain wood tick, Dermacentor andersoni (Acari, Ixodidae). J Invertebr Pathol. 2005;90:177–186. doi: 10.1016/j.jip.2005.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chang J, Pang E, He H, Kwang J. Identification of novel attenuated Salmonella Enteritidis mutants. FEMS Immunol Med Microbiol. 2008;53:26–34. doi: 10.1111/j.1574-695X.2008.00394.x. [DOI] [PubMed] [Google Scholar]
- 49.Heier K, Thomson MJ, Aziz N, Moir JW. The nitric oxide (NO)-sensing repressor NsrR of Neisseria meningitidis has a compact regulon of genes involved in NO synthesis and detoxification. J Bacteriol. 2008;190:2488–2495. doi: 10.1128/JB.01869-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Isabella VM, Lapek JD, Jr, Kennedy EM, Clark VL. Functional analysis of NsrR, a nitric oxide-sensing Rrf2 repressor in Neisseria gonorrhoeae. Mol Microbiol. 2009;71:227–239. doi: 10.1111/j.1365-2958.2008.06522.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ngok-Ngam P, Ruangkiattikul N, Mahavihakanont A, Virgem SS, Sukchawalit R, et al. Roles of Agrobacterium tumefaciens RirA in iron regulation, oxidative stress response and virulence. J Bacteriol. 2009 doi: 10.1128/JB.01380-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Chao TC, Buhrmester J, Hansmeier N, Puhler A, Weidner S. Role of the regulatory gene rirA in the transcriptional response of Sinorhizobium meliloti to iron limitation. Appl Environ Microbiol. 2005;71:5969–5982. doi: 10.1128/AEM.71.10.5969-5982.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Pereira LS, Oliveira PL, Barja-Fidalgo C, Daffre S. Production of reactive oxygen species by hemocytes from the cattle tick Boophilus microplus. Exp Parasitol. 2001;99:66–72. doi: 10.1006/expr.2001.4657. [DOI] [PubMed] [Google Scholar]
- 54.Pallen MJ, Wren BW. Bacterial pathogenomics. Nature. 2007;449:835–842. doi: 10.1038/nature06248. [DOI] [PubMed] [Google Scholar]
- 55.Darby AC, Cho NH, Fuxelius HH, Westberg J, Andersson SG. Intracellular pathogens go extreme: genome evolution in the Rickettsiales. Trends Genet. 2007;23:511–520. doi: 10.1016/j.tig.2007.08.002. [DOI] [PubMed] [Google Scholar]
- 56.Fournier PE, El Karkouri K, Leroy Q, Robert C, Giumelli B, et al. Analysis of the Rickettsia africae genome reveals that virulence acquisition in Rickettsia species may be explained by genome reduction. BMC Genomics. 2009;10:166. doi: 10.1186/1471-2164-10-166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Munderloh UG, Jauron SD, Kurtti TJ. The Tick: a Different Kind of Host for Human Pathogens. In: Goodman JL, Dennis DT, Sonenshine DE, editors. Tick-Borne Diseases of Humans. Washington, D.C.: ASM Press; 2005. pp. 37–64. [Google Scholar]
- 58.Allesen-Holm M, Barken KB, Yang L, Klausen M, Webb JS, et al. A characterization of DNA release in Pseudomonas aeruginosa cultures and biofilms. Mol Microbiol. 2006;59:1114–1128. doi: 10.1111/j.1365-2958.2005.05008.x. [DOI] [PubMed] [Google Scholar]
- 59.Bordenstein SR, Wernegreen JJ. Bacteriophage flux in endosymbionts (Wolbachia): infection frequency, lateral transfer, and recombination rates. Mol Biol Evol. 2004;21:1981–1991. doi: 10.1093/molbev/msh211. [DOI] [PubMed] [Google Scholar]
- 60.Bordenstein SR, Reznikoff WS. Mobile DNA in obligate intracellular bacteria. Nat Rev Microbiol. 2005;3:688–699. doi: 10.1038/nrmicro1233. [DOI] [PubMed] [Google Scholar]
- 61.Munderloh UG, Jauron SD, Fingerle V, Leitritz L, Hayes SF, et al. Invasion and intracellular development of the human granulocytic ehrlichiosis agent in tick cell culture. J Clin Microbiol. 1999;37:2518–2524. doi: 10.1128/jcm.37.8.2518-2524.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
A dot plot comparison of the R. peacockii and R. rickettsii genomes.
(0.01 MB PNG)
DNA sequence found in R. peacockii and not in R. rickettsii SS.
(0.07 MB DOC)
Deletions in R. peacockii and their association with ISRpe1 transposons.
(0.26 MB DOC)
Phylogenetic analysis of the rickettsial small hsp proteins.
(0.15 MB DOC)
Kyte-Doolittle plots of hydrophobicity for the three small hsp/chaperone proteins from R. peacockii.
(0.04 MB DOC)
Support for R. rickettsii A1G_02570 as phosphoethanolamine transferase.
(0.05 MB DOC)
Nonsense mutations in R. rickettsii Sheila Smith relative to R. peacockii.
(0.04 MB DOC)