

Abstract
We examine the evidence for the possibility that 21st-century climate change may cause a large-scale “dieback” or degradation of Amazonian rainforest. We employ a new framework for evaluating the rainfall regime of tropical forests and from this deduce precipitation-based boundaries for current forest viability. We then examine climate simulations by 19 global climate models (GCMs) in this context and find that most tend to underestimate current rainfall. GCMs also vary greatly in their projections of future climate change in Amazonia. We attempt to take into account the differences between GCM-simulated and observed rainfall regimes in the 20th century. Our analysis suggests that dry-season water stress is likely to increase in E. Amazonia over the 21st century, but the region tends toward a climate more appropriate to seasonal forest than to savanna. These seasonal forests may be resilient to seasonal drought but are likely to face intensified water stress caused by higher temperatures and to be vulnerable to fires, which are at present naturally rare in much of Amazonia. The spread of fire ignition associated with advancing deforestation, logging, and fragmentation may act as nucleation points that trigger the transition of these seasonal forests into fire-dominated, low biomass forests. Conversely, deliberate limitation of deforestation and fire may be an effective intervention to maintain Amazonian forest resilience in the face of imposed 21st-century climate change. Such intervention may be enough to navigate E. Amazonia away from a possible “tipping point,” beyond which extensive rainforest would become unsustainable.
Keywords: carbon dioxide, drought, fire, tropical forests, adaptation
The response of components of the Earth system to increasing levels of anthropogenic greenhouse-gas forcing is unlikely to be continuous and gradual; instead there may be “tipping elements” in the system (1). Among the most iconic of these is the Amazon rainforest, with some projections suggesting the possibility of substantial and rapid “dieback” (2–4). The Amazon forest biome is biologically the richest region on Earth, hosting ≈25% of global biodiversity, and is a major contributor to the biogeochemical functioning of the Earth system (3). Its large-scale degradation would leave an enduring legacy on the functioning and diversity of the biosphere. We review the evidence for such a tipping element in Amazonia and examine climate model projections in the context of rainforest viability considering direct human pressures on the forest system.
There is clear and ongoing change in the physical environment of Amazonia, whether through increasing atmospheric carbon dioxide concentrations, associated imposed climate change, or more direct intervention because of the spread of settlement, deforestation, forest timber extraction, or any related fire initiation. Such perturbations are certain to persist on policy-relevant timescales. The challenge is to identify and characterize system nonlinearities, thresholds, and feedbacks and determine which components of this system are open to manipulation and management in a manner beneficial to the long-term sustainability of the Amazonian social ecological system.
In this paper, we ask 3 questions. First, what are the climatic thresholds that favor the current presence of a forest as opposed to savanna? Second, what do climate models say about the likely direction of expected climate change in Amazonia and any associated likelihood of large regions of Amazonia crossing thresholds of forest viability during the 21st century? Third, to what extent does direct human pressure (deforestation, fragmentation, and fires) influence the transition?
Current Climate and Vegetation in Amazonia
The lowland forests of Amazonia have a mean annual temperature of 26 °C, with very little spatial variability, and a mean annual precipitation of ≈2400 mm, ranging from >3000 mm in North West Amazonia to <1500 mm at the forest–savanna transition zones (5).
Two relevant features of the rainfall regime are (i) the intensity and duration of the dry season and (ii) the overall water supply. To describe the accumulated water stress that occurs across a dry season, we employ the maximum climatological water deficit (MCWD) (6).
MCWD is defined as the most negative value of climatological water deficit (CWD), attained over a year, where the monthly change in water deficit is precipitation (P) (mm/month) − evapotranspiration (E) (mm/month). For month n,
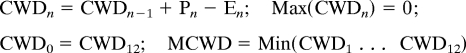 |
At the wettest time of the year, we assume the soil is saturated (i.e., set CWD = 0) and start the 12-month cycle of calculation from this wet phase. For any multiyear period, we apply this calculation to the mean annual cycle of precipitation (rather than calculating for each year and then taking the mean MCWD). We do not attempt to model E but fix it at 3.33 mm/day, ≈100 mm/month. Hence, the CWD is only an approximate indicator of actual soil water deficit because it does not account for seasonal variation in E (driven mainly by radiation and phenology) (7) or spatial variation in E related to soil and root properties. In addition, actual transpiration rates may increase with rising temperatures; the limitations of our assumption of fixed E are discussed in Influences of Temperature and CO2 Change. A fixed E is also inappropriate outside of lowland tropical regions, where lower energy supply may result in lower E.
As a metric of overall water supply, we utilize annual precipitation (AP) (mm); for a given MCWD, higher values of AP reflect higher rainfall during the wet season. AP and MCWD are mapped in Fig. S1 A and B. For spatial analysis, these data are derived from NASA's Tropical Rainfall Monitoring Mission (TRMM) satellite for the period 1998–2005. TRMM has the advantage of complete coverage across otherwise data-poor areas, although it may underestimate wet and dry extremes in rainfall (6).
Fig. S2 plots the annual observed values of AP and MCWD for the period 1970–1999, using the Climate Research Unit (CRU) observational dataset (8). We employed CRU rather than TRMM for time series analysis and model validation because of its longer duration. The northern and southern limits of Amazonia were approximated as 3°N and 12°S. Throughout this paper, W. Amazonia is taken to extend from 72°W to 60°W and E. Amazonia from 60°W to 48°W.
E. Amazonia is drier and more seasonal than W. Amazonia. Within E. Amazonia, in El-Niño-associated dry years (e.g., 1982, 1983, and 1997) (Fig. S2), regional MCWD values approach −200 mm, close to savanna transition thresholds if such dry conditions were to persist for the long term (see Hydrological Thresholds on Current Forest Extent). In W. Amazonia, there is no or very little seasonal water stress even in dry years. This crude East–West division masks within- region variability, e.g., SW Amazonia is as dry and seasonal as much of E. Amazonia (5). There are no significant trends in AP or MCWD during this period (1970–1999).
Hydrological Thresholds on Current Forest Extent
The current biogeography of Amazonia can give some indication of current climatic constraints on forest cover. Here, we explore these constraints by using combined satellite-derived maps of forest extent and precipitation regime. For a vegetation cover classification, we employ the Global Land Cover 2000 map (9), distinguishing between evergreen forest, deciduous forests, grasslands, and shrublands (the latter two we ascribe to savanna) Fig. S1C. Deforested areas are excluded from the analysis.
Fig. 1 plots the distribution of these vegetation types in the rainfall regime space defined by AP and MCWD. The analysis focuses on an area between 45°W and 70°W and 0° and 20°S, covering both forest and savanna. It is evident that there are no sharp vegetation thresholds in the rainfall regime space (MCWD, AP); some savanna pixels are found in predominantly forest climates, and some evergreen forests are found in dry climates. The diffuseness of the boundary reflects variation in local surface hydrology and soil properties, e.g., seasonal flooding can favor savanna in dry forest climates, shallow water tables can allow gallery or riverine forest to persist in dry savanna climates, or more fertile soil may favor trees over grasses. The fuzziness in the thresholds may also reflect errors in vegetation or phenological classification or in the rainfall map. Nevertheless, it is possible to identify broad climatic thresholds consistent with the definition of tipping points. Evergreen forests tend to predominate if the dry season is weak (MCWD > −200 mm). If AP > 1500 mm, forests tend to predominate, intuitively becoming increasingly seasonal and deciduous for more negative values of MCWD (the land-use classification cannot distinguish evergreen from semideciduous forests). If AP < 1500 mm, savannas predominate if MCWD < −400 mm, and there is a broad transition zone for −400 mm < MCWD < −200 mm, where there is a gradual shift in the relative abundance of savanna relative to forest. For analyses later in this paper, we ascribe the following terms to bioclimatic spaces (not necessarily the vegetation types): (i) “rainforest”: MCWD > −200 mm; (ii) “seasonal forest”: MCWD < −200 mm and rainfall >1500 mm (the transition between rainforest and seasonal forest climates is in reality a continuum); (iii) “savanna”: MCWD < −300 mm (midpoint of the transition range) and rainfall < = 1500 mm (latter shaded in Fig. 1).
Fig. 1.

The relationship between vegetation type and rainfall regime. Rainfall regime is derived from TRMM for the period 1998–2006. Our suggested savanna zone is shaded (MCWD < −300 mm, AP < 1500 mm).
Climate and Atmospheric Change in Amazonia
Recently, temperatures in lowland tropical regions worldwide have been increasing at ≈0.25 °C per decade (5) and are projected to rise by 3–8 °C (mean 5 °C) over the 21st century under the A2 emissions scenario (Fig. S3), with the higher values more likely if forest dieback induces strong local biophysical feedbacks (10). Precipitation trends are more difficult to elucidate. There has been no overall trend in region-wide annual mean precipitation in recent decades, but evidence of increasing frequency of dry events in southern Amazonia over the period 1970–1999 has been found (11).
Projections of rainfall change over the 21st century remain a major challenge for climate models and, for that reason, are the focus of particular scrutiny in this paper. In discussions of Amazonian dieback, there has been a tendency to either use one model (2, 4, 12), perturbed physics ensembles of a single model (13), or to employ a number of models but without a critical examination of their respective outputs (2, 14).
Evaluating Model Projections of Rainfall Change in the 21st Century
We examine all 19 models used in the Intergovernmental Panel on Climate Change AR4 Global Climate Models that provide runs over the 20th and 21st centuries under the medium-high range Special Report on Emissions Scenarios A2 (15). We examine model simulations of rainfall regime for both E. and W. Amazonia for the period 1970–1999 (“the late 20th century”) and the period 2070–2099 (“the late 21st century”). Tables S1 and S2 list the models and details of their rainfall regime. Here, none of these simulations incorporate feedbacks of vegetation and soil change because the global carbon cycle is not modeled.
Fig. 2A plots the late 20th-century and late 21st-century precipitation projections for each GCM for E. Amazonia onto the rainfall regime space (MCWD and AP) (a similar plot for W. Amazonia is presented in Fig. S4). The mean CRU-derived rainfall regime for 1970–1999 for each region is also plotted. A very large variation among models is apparent, both in current climates and future trends. In E. Amazonia, almost all models (18/19) substantially underestimate rainfall relative to the CRU or TRMM data. In W. Amazonia, 17/19 models substantially underestimate. The mean late 20th-century state of E. Amazonia shows no substantial shift over the 21st century, although individual models can show large shifts in either direction.
Fig. 2.

An evaluation of GCM simulations in change of rainfall regime in E. Amazonia. (A) The rainfall regime is simulated for the late 20th century (base of arrow) and the late 21st century (tip of arrow) for 19 GCMs under the A2 emissions scenario. The TRMM-derived range of rainfall regime is indicated with gray pixels, the observed spatially averaged CRU climate for 1970–1999 indicated with blue stars, and the region with a suggested savanna-favoring rainfall regime is shaded. (B) The trajectories of changes in GCM rainfall regime when recalculated as relative changes forced to start from the CRU observed climatology. The tip of the arrow indicates the late 21st-century rainfall regime. Our suggested savanna zone is shaded (MCWD < −300 mm, AP < 1500 mm).
Using the bioclimatic thresholds outlined above (MCWD > −300 mm or rainfall >1500 mm), two-thirds of the models (13/19) fail to simulate a late 20th-century rainfall regime sufficient to support widespread rainforest in E. Amazonia and 5/19 fail to do so in W. Amazonia. In E. Amazonia, 14/19 models have a mean state drier than the actual Amazonian rainfall regime in El Niño years (Fig. S2). If we select only the models that simulate a sufficient late 20th rainfall regime, again, there appears little trend in the mean rainfall regime over the 21st century.
The mismatch between observed climate and GCM-simulated climate renders the interpretation of GCM scenarios in Amazonia challenging. For the next step of our analysis, we take the relative change in monthly rainfall predicted by each GCM but offset these changes to the observed climate for the late 20th century rather than the GCM-simulated climate. For the mean late 21st century-rainfall in month n in GCM i:
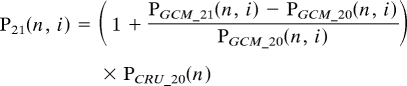 |
where PGCM_20 and PGCM_21 are the GCM-simulated rainfall for the late 20th and 21st centuries, respectively, PCRU_20 is the observed monthly climate from the CRU climatology, and P21 is the revised estimate of rainfall for the late 21st century. The late 21st rainfall regime (MCWD and AP) is then recalculated from this revised prediction of monthly data, P21, and plotted for E. Amazonia in Fig. 2B (and for W. Amazonia in Fig. S4).Our implicit assumption is that GCM simulations better capture future seasonal changes in the relative intensity of precipitation, despite their current tendency to underestimate absolute values of precipitation.
With this revised calculation, many GCMs show a strong tendency for increased seasonality over the 21st century (Fig. 2B). This tendency is driven by decreased rainfall in the dry-season period (July–October in most of Amazonia), which tends to occur irrespective of the present-day seasonality simulated by the GCM. Renormalization to observed precipitation stretches this tendency. All but 2 GCMs show more negative MCWD over the 21st century, with 10/19 models passing our approximate bioclimatic threshold from rainforest to seasonal forest (MCWD < −200 mm). However, the high value of rainfall in the observed climatology ensures that the general tendency is toward a seasonal forest climate rather than savanna climate; wet-season rainfall remains high enough to allow recharge of any dry-season water deficit. One model (HadCM3) dries sufficiently to clearly enter the savanna rainfall regime, and 4 other models approach the seasonal forest–savanna threshold. If we regard all models as independent and equally probable, we can use the fraction of models that indicate a certain result as a crude metric of probability. Hence, there is a high probability (10/19 models, 53%) of transition to a rainfall regime more suitable for seasonal forest and a substantial probability (5/19 models, 26%) of approaching a rainfall regime more appropriate for savanna. These probabilities are broad indicators ascribed to rainfall regime, not to vegetation type; how the vegetation will respond to such a change in rainfall depends on multiple factors discussed under Vegetation Response to a Drying Amazon. Applying a selection criterion (MCWD > −200 mm and AP > 1500 mm, prenormalization) gives 6 models that simulate a climate regime appropriate for rainforest or seasonal forest in the late 20th century (and hence require a smaller perturbation to their simulated rainfall to match the CRU observed rainfall). Considering only the selected models, the decrease in MCWD over the 21st century is still substantial but smaller, with 2 models (2/6, 33%) crossing the threshold into seasonal forest and 1 model (1/6, 17%) approaching the savanna threshold. In reality, all models are not equivalent, and the more extreme outcomes cannot be dismissed as outliers. For example, the model with the most severe change in rainfall regime (HadCM3) captures many aspects of the coupling between Atlantic sea surface temperatures (SSTs) and Amazonian drought (12), and a different model weighting based on capturing interannual variability would suggest this more extreme result is more likely.
Why do GCMs tend to underestimate rainfall in Amazonia, and why does our analysis suggest increased seasonality over the 21st century? The underestimation of rainfall is likely a result of the coarse grid scale of GCMs, resulting in poor representation of finer-scale meteorological processes that are known to intensify precipitation. Some of these features (e.g., intense localized convection in squall lines) are common to all tropical regions but others are peculiar to South America. Many GCMs simulate that the wet-season rains (the “South American monsoon”) penetrate much further south than they actually do. In practice, their southward passage is blocked by 2 main phenomena: the high-elevation “ventilation” of cool, westerly air over the Andes that dissipate the convective energy of the moist Amazonian air masses and the Rossby wave-induced subsidence of air over Bolivia (16). Both of these mechanisms limit the southernly penetration of rains and simultaneously enhance rainfall over the Amazonia convective zone. They are fundamental features of regional atmospheric circulation that are challenging for any coarse-scale GCM to capture and are unlikely to change fundamentally in character over the 21st century, hence, it is likely the GCM underestimation of rainfall will persist into the late 21st-century projections. Finer-scale regional climate models may better capture absolute rainfall.
The 21st-century intensification of dry seasons suggested by our analysis is probably partially driven by the general intensification of tropical circulation caused by increased temperatures and tropospheric moisture content (17). This intensification increases precipitation within the convective zone but also causes narrowing of the convective zone and suppression of convection in the neighboring air subsidence zones. Hence, wet seasons intensify, but dry seasons also lengthen and intensify. Therefore, MCWD becomes more negative, but AP may not necessarily decrease. In addition to this intensification of the existing circulation, precipitation regimes are also affected by shifts in SST patterns. In particular, enhanced relative warming of the tropical north Atlantic is associated with intensification of dry seasons in S. and E. Amazonia. In at least one model (HadCM3), drought becomes more frequent and persistent as the northern tropical Atlantic warms disproportionately over the 21st century (12).
We interpret our findings as suggesting that (i) there is a high probability of some increase of dry-season intensity in E. Amazonia, with a medium probability (≈30–50%) that this will result in a climate more appropriate for seasonal forest; (ii) because GCMs tend to underestimate wet-season rainfall, the real-world probability of future rainfall regimes more appropriate for savanna is smaller (0–25%) but certainly not negligible; and (iii) W. Amazonia, as a whole, is likely to stay in a rainforest-favoring climate (although the drier northern and southern margins may not), but there is a possibility (near 10%) of shifting from a generally aseasonal moisture regime to a seasonally dry regime. These percentage probabilities should be taken as merely indicative, but our basic conclusion remains that the rainfall regime of E. Amazonia is likely to shift over the 21st century in a direction that favors more seasonal forests rather than savanna.
Vegetation Response to a Drying Amazon
We now consider the vegetation response to this drying: Will Amazonian forests simply and rapidly move to be in equilibrium with any new rainfall regime, or are there additional factors to consider?
Three features make 21st century-climate change unique when compared with past climate change. (i) Any change in rainfall regime will occur concurrently with changes in other atmospheric variables (in particular temperature and carbon dioxide concentration) that are being pushed upwards outside of envelopes of natural variability experienced over the last tens of millions of years. These variables also influence vegetation functioning and, thus, mediate the direct response to rainfall regime alone. (ii) The rates of change projected over this century are rapid compared with past transitions. For instance, the postglacial warming in Amazonia (driven predominantly by radiative forcing by CO2 as opposed to global ice-albedo feedbacks) was just 0.1 °C per century (18) compared with 3–5 °C this century. Current rates of change are so large that they are significant over the lifetime of individual trees, making consideration of transient responses and lags in the vegetation system important. (iii) The atmospheric change is also accompanied by an unprecedented intensity of direct pressure on the tropical forests through logging, deforestation, fragmentation, and fire use. This direct pressure is likely to influence the vulnerability or resilience of the biome to climatic change. We discuss each of the above factors in turn, drawing on insights from field and experimental studies.
Influences of CO2 and Temperature Change.
Under the A2 emissions scenario, atmospheric CO2 rises to 730-1020 ppmv by 2100 (19). In addition to its greenhouse-gas role, increased CO2 will potentially directly stimulate plant photosynthesis by increasing the rate of CO2 diffusion into leaves and directly enhancing the rate of carboxylation (20). This may cause a transient increase in biomass production rates and a net biomass carbon sink (21), but it may also accelerate mortality and turnover rates, leading to a more dynamic and potentially lower biomass forest. This “CO2 fertilization effect” is likely to saturate over time because of ecophysiological saturation, nutrient supply limitation, or ecological feedbacks such as increased abundance of high turnover species (22). CO2 fertilization favors plants that utilize the C3 photosynthetic pathway (most trees). However, many savanna grasses use instead the C4 pathway, potentially shifting the balance in favor of forest in forest–savanna transition zones for a given rainfall regime. Moreover, high CO2 increases water use efficiency (leaf stomata need to open less and allow less water to escape for the same uptake of CO2), and this will partially mitigate the effect of increased dry-season intensity.
In contrast, increases in atmospheric temperature will cause higher leaf-to-air vapor pressure deficit and thus enhance transpiration, causing actual soil water deficits to become increasingly more severe than MCWD. Hence, even without change in rainfall regime, warming can be expected to increase plant water stress (14). Increasing temperatures may also cause increases in plant metabolic activity and respiration, but it seems likely that in the longer term, these processes will acclimate to increased temperature (23). Photosynthesis rates may reduce slightly under rising temperatures because tropical forest leaves currently operate near their temperature optimum, although this potential reduction will probably be less than the concurrent gain from higher CO2 concentrations (20).
In conclusion, the 21st-century rise in CO2 may to some extent mitigate the effects of enhanced seasonality in rainfall and lessen the likelihood of forest loss. However, the concurrent rise in temperature could increase the risk of a vegetation transition. Which effect is more important? For a tentative assessment of the relative magnitude of these 2 effects, we tested the sensitivity of transpiration in the MOSES-TRIFFID vegetation model to increases in temperature and CO2 (see SI Text and Fig. S5). The imposed increases in Amazon surface temperature (+4.7 °C) and CO2 (to 850 ppm) are typical of GCM simulations for 2100 under the A2 emissions scenario. Under these conditions, and with adequate water supply, the model simulates the higher temperatures to increase evapotranspiration by ≈55%, whereas high CO2 decreases evapotranspiration by ≈17%. The net effect is a 31% increase of evapotranspiration to 4.7 mm/day or 140 mm/month. This sensitivity analysis is tentative and warrants deeper exploration of model assumptions. Seasonally dry systems may limit water loss through increased stomatal closure, losing productivity to lessen water loss. The implications of such an increase in evapotranspiration are 2-fold: First, the length of the dry season (period of negative water balance) is extended, and the MCWD becomes more negative. Second, the increased annual water demand increases the threshold annual rainfall required to maintain seasonal forest instead of savanna. In terms of Fig. 2B, the end point of each arrow shifts to the left, and the line between savanna and seasonal forest may shift upward, though by how much also depends on other factors such as CO2 fertilization. These factors in combination would substantially increase the risk of transition from forest to savanna.
Transient Responses to Drought.
We turn next to consideration of the transient response to rapid climate change. Forest trees can be long-lived organisms, and it is possible that “demographic inertia” (inertia caused by long lifespans and slow community turnover) in the forest system may delay any forest dieback. Moreover, the presence of forest may modify local microclimate (evapotranspiration, rainfall generation, exclusion of invading grasses, shading of soil surface, and seedlings) sufficiently to favor the persistence of forest (“microclimatic inertia”). Once established, closed-canopy, deep-rooted forest may persist even if the local climate has shifted to savanna conditions. A useful analogy may be drawn from physics. In a phase transition from liquid to solid, a liquid may persist in a supercooled state beyond the equilibrium threshold for solidification if there are insufficient nucleation points (e.g., impurities) to induce solidification. By analogy, a forest may persist in the drying-induced transition to savanna if there are insufficient nucleation points (most likely fire ignition points, see The Role of Fire, below) to break open the forest and trigger the transformation.
It is conceivable that any such ecosystem inertia could be sufficient to buffer the presence of Amazonian forest into the 22nd century, by when, under optimistic scenarios, policy and technological solutions may stabilize or even draw down atmospheric CO2 concentrations.
Two recent manipulation experiments have attempted to address these questions by excluding a fraction of the incoming throughfall of precipitation to the soil in 1-ha rainforest plots in E. Amazonia (24–26). Both experiments shifted a rainfall regime typical of evergreen forest into one near to or within the savanna bioclimatic zone (Fig. 3). These experiments induced reductions in transpiration, leaf area, reproductive output, and photosynthesis in the first 2 years (26) but there was a lag of ≈3 years between the start of the exclusion and the onset of increased tree mortality (25) and the beginning of the break-up of the forest canopy. These experiments suggest that there may be some inherent resilience to reduced rainfall, which is linked to the existence of deep rooting systems that can exploit substantial soil moisture reserves and, hence, buffer the rainforest against seasonal drought. However, a threshold exists for rainforest maintenance when wet-season rain does not completely recharge dry-season soil-water depletion (Fig. 3). After a few years, soil water reserves deplete, and the forest begins to witness enhanced tree mortality, reducing both demographic and microclimatic ecosystem inertia.
Fig. 3.

Drought experiments and field observations of forest fire in the context of the mean of GCM projections in change of rainfall regime in E. and W. Amazonia. For the GCMs, the arrow base indicates late 20th-century rainfall regime whereas the arrow tip indicates late 21st-century rainfall regime. For the drought experiments, the arrow base indicates control rainfall regime whereas the arrow tip indicates the estimated rainfall regime. Diamonds indicate field studies of flammable forests (see Table S3), with rainfall regimes derived from TRMM (1998–2006). Our suggested savanna zone is shaded (MCWD < −300 mm, AP < 1500 mm).
Interaction Between Drought and Deforestation.
Amazonia, and in particular its drier margins, is the scene for intense human pressure on the forest through logging, deforestation, and expansion of fire use. In this context, consideration of the ecophysiological and ecological responses of the natural system to climate change gives only a partial picture of the future of Amazonia, and it is important to consider the impacts of this direct pressure and its interactions with atmospheric change. Under business-as-usual scenarios of deforestation, it is likely that significant areas of E. Amazonia would be directly deforested, although there remains the possibility of greatly improved governance and maintenance of forest area (2, 27). Nevertheless, it is likely that agricultural frontiers will spread further into the region, providing the threads of a web of ignition points ready to ignite Amazonian forests in the event of a shift to a drier and/or more seasonal climate (28). Pressures come from the spread of road infrastructure through the region, coupled with increased regional and global demand for Amazonian beef and soya, and the emerging global demand for biofuels (28, 29)
This pressure will influence the response of forests in a number of ways by (i) directly removing forest cover and being an independent agent of Amazon dieback; (ii) directly modifying local climate, surface temperatures, and rainfall regime, thus, contributing to regional climate change; and (iii) increasing the presence of, and vulnerability to, fire.
Deforestation may directly affect local climate by reducing local recycling of soil water through deep roots into forest transpiration and consequently into precipitation, although this seems to depend on the scale and location of deforestation. Deforestation in E. Amazonia may reduce rainfall downwind in W. Amazonia (30), a region that otherwise seems less vulnerable to change in rainfall regime. In addition, lost forest transpiration results in decreased surface cooling and thereby an increase regional air temperatures, evaporative demand, and water stress in remaining forests. Land-use change and fire also affect the rainfall regime by greatly increasing the aerosol content of the atmosphere through smoke and dust. High aerosol content favors less frequent but more intensive convective rain and possible suppression of rain in the dry season. A retreat of Amazonian forest (whether caused by deforestation or severe drying) would therefore further exacerbate regional climate change by altering local water recycling and other biophysical properties (10).
The Role of Fire.
We have argued that forests may have some resilience to intensification of the dry season. However, this may break down when the presence of fire is considered. A number of field studies have reported that seasonal tropical forests do become temporarily flammable, but the lack of natural ignition points in Amazonia inhibits the amount of natural fire. Fig. 3 plots the rainfall regime for a selection of studies that have demonstrated forest flammability. Surrounding the savanna bioclimate zone, there is a wider zone of potential forest flammability where fires are possible but usually do not occur because of the lack of ignition sources. However, the spread of human settlement, forest fragmentation, and logging has resulted in the expansion of ignition sources, with 28% of Brazilian Amazonia facing incipient fire pressure, now being within 10 km of an ignition source (31). The potentially critical role of fire was apparent during recent droughts in Amazonia, with extensive fires leaking from agricultural zones into flammable forests during the droughts of 1997, 1998, 2005, and 2007 (6, 32).
Over the last decade, a number of field studies have explored the changes in primary rainforests exposed to individual or repeated fire events (33, 34). Most rainforest trees are poorly adapted to fire stress, and even low-intensity forest wildfires can lead to extensive tree death. These studies paint a compelling picture of how fire-intolerant rainforests may decline and break open under scenarios of drying and increasing fire frequency, an early peek into the likely ecology of an actual Amazonian dieback. They document sharply increased mortality of both small and large trees, a rapid collapse of biomass canopy structure, a decline in large vertebrates and tree species dependent on these for dispersal, spread of fast growing, wind-dispersed species more characteristic of secondary forest, and accompanying substantial decline in plant and vertebrate biodiversity (33).
Conclusion: Navigating Away from a Tipping Point?
Our analysis concluded that, under mid-high-range emissions scenarios, there is a high probability of intensified dry seasons in E. Amazonia and a medium probability that the rainfall regime will shift sufficiently to a climate state where seasonal forest is more viable than rainforest. Rising temperatures and transpiration rates, widespread deforestation, and climate-change-induced forest retreat may further contribute to intensified seasonal water stress. Forests at the dry margins or on shallow or infertile soils may be most vulnerable. The extent of forest die-back may be reduced by the ecophysiological response to rising CO2 concentrations and ecosystem acclimation to rising temperature and possibly (temporarily) by inherent ecosystem inertia in the response of forest ecosystems to climatic change. However, the regional surface warming caused by substantial forest loss and the rapid proliferation of human pressure points (in particular forest edges and fire ignition points) may substantially undermine this resilience. In such an event, it could be considered that E. Amazonia had passed a tipping point in ecosystem structure and function.
Just as human activity and the spread of fire may be critical in triggering a breakdown of forest resilience and consequent dieback, direct intervention to maintain forest area and limit the spread of fire offers the potential to maintain forest resilience and avoid any such tipping point (2, 28). Recent developments in expansion of protected areas, reduction of deforestation rate, and pioneering schemes for local payment for Amazonian ecosystem services suggest that this is not impossible (2, 35). Efforts have been further bolstered by the recently stated commitment of Brazil to greatly reduce net deforestation and by the inclusion of Reduced Emissions from Deforestation and Degradation (REDD) as a mechanism for mitigating climate change within United Nations climate-change agreements, which greatly increases the potential funds available for such an enterprise. Maintaining forest cover would not only be a strategy for climate-change mitigation, regional development, and biodiversity conservation but also a potential strategy for adaptation as the climate of E. Amazonia tends toward one of intensified seasonality.
The dieback of the forests of E. Amazonia in the 21st century is far from inevitable but remains a distinct possibility. The first priority (and ultimate responsibility) in minimizing the risk of this dieback is reducing global greenhouse-gas emissions. However, appropriate adaptation measures and forest management with E. Amazonia could play a major role in minimizing the prospects of large-scale forest degradation while also contributing to the global mitigation effort. Even with sufficient funds and willpower, the implementation of biosphere management on such a scale will be a substantial challenge and understanding of the social, political, and economic context will be critically important (36). The prospect of navigating much of Amazonia away from a tipping point makes this a challenge worth facing up to.
Supplementary Material
Acknowledgments.
We thank P. Harris, M. New, R. Betts, O. Phillips, D. Cameron, and P. Brando for helpful comments. We also acknowledge the modeling groups, the Program for Climate Model Diagnosis and Intercomparison, and the World Climate Research Programme (WCRP) Working Group on Coupled Modeling for their roles in making available the WCRP Coupled Model Intercomparison Project 3 multimodel dataset. Y.M. is supported by the Jackson Foundation, and L.E.O.C., D.G., C.H., and R.F. are supported by the Natural Environment Research Council.
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
This article contains supporting information online at www.pnas.org/cgi/content/full/0804619106/DCSupplemental.
References
- 1.Lenton TM, et al. Tipping elements in the Earth's climate system. Proc Natl Acad Sci USA. 2008;105:1786–1793. doi: 10.1073/pnas.0705414105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cox PM, et al. Amazonian forest dieback under climate-carbon cycle projections for the 21st century. Theor Appl Climatol. 2004;78:137–156. [Google Scholar]
- 3.Malhi Y, et al. Climate change, deforestation, and the fate of the Amazon. Science. 2008;319:169–172. doi: 10.1126/science.1146961. [DOI] [PubMed] [Google Scholar]
- 4.Oyama MD, Nobre CA. A new climate-vegetation equilibrium state for tropical South America. Geophys Res Lett. 2003;30:2199. [Google Scholar]
- 5.Malhi Y, Wright J. Spatial patterns and recent trends in the climate of tropical rainforest regions. Philos Trans R Soc London Ser B. 2004;359:311–329. doi: 10.1098/rstb.2003.1433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Aragão LEOC, et al. Spatial patterns and fire response of recent Amazonian droughts. Geophys Res Lett. 2007;34:L07701. [Google Scholar]
- 7.Hasler N, Avissar R. What controls evapotranspiration in the Amazon basin? J Hydrometeor. 2007;8:380–395. [Google Scholar]
- 8.New M, Hulme M, Jones P. Representing twentieth-century space-time climate variability. Part II: Development of 1901–96 monthly grids of terrestrial surface climate. J Climate. 2000;13:2217–2238. [Google Scholar]
- 9.Eva HD, et al. A land map of South America. Global Change Biol. 2004;10:731–744. [Google Scholar]
- 10.Betts RA, et al. The role of ecosystem-atmosphere interactions in simulated Amazonian precipitation decrease and forest dieback under global climate warming. Theor Appl Climatol. 2004;78:157–175. [Google Scholar]
- 11.Li WH, Fu R, Juarez RIN, Fernandes K. Observed change of the standardized precipitation index, its potential cause and implications to future climate change in the Amazon region. Philos Trans R Soc London Ser B. 2008;363:1767–1772. doi: 10.1098/rstb.2007.0022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cox PM, et al. Increasing risk of Amazonian drought due to decreasing aerosol pollution. Nature. 2008;453:212–215. doi: 10.1038/nature06960. [DOI] [PubMed] [Google Scholar]
- 13.Huntingford C, et al. Towards quantifying uncertainty in predictions of Amazon “dieback.”. Philos Trans R Soc London Ser B. 2008;363:1857–1864. doi: 10.1098/rstb.2007.0028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Salazar LF, Nobre CA, Oyama MD. Climate change consequences on the biome distribution in tropical South America. Geophys Res Lett. 2007;34:L09708. [Google Scholar]
- 15.Nakicenovic N, et al. IPCC Special Report on Emission Scenarios. UK: Cambridge Univ Press; 2000. 599 pp. [Google Scholar]
- 16.Chou C, Neelin JD. Mechanisms limiting the southward extent of the South American summer monsoon. Geophys Res Lett. 2001;28:2433–2436. [Google Scholar]
- 17.Chou C, Neelin JD. Mechanisms of global warming impacts on regional tropical precipitation. J Climate. 2004;17:2688–2701. [Google Scholar]
- 18.Bush MB, Silman MR, Urrego DH. 48,000 years of climate and forest change in a biodiversity hot spot. Science. 2004;303:827–829. doi: 10.1126/science.1090795. [DOI] [PubMed] [Google Scholar]
- 19.Friedlingstein P, et al. Climate-carbon cycle feedback analysis, results from the C4MIP model intercomparison. J Climate. 2006;19:3337–3353. [Google Scholar]
- 20.Lloyd J, Farquhar GD. Effects of rising temperatures and [CO2] on the physiology of tropical forest trees. Philos Trans R Soc London Ser B. 2008;363:1811–1817. doi: 10.1098/rstb.2007.0032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Phillips OL, Lewis SL, Baker TR, Chao KJ, Higuchi N. The changing Amazon forest. Philos Trans R Soc London Ser B. 2008;363:1819–1827. doi: 10.1098/rstb.2007.0033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Phillips OL, et al. Increasing dominance of large lianas in Amazonian forests. Nature. 2002;418:770–774. doi: 10.1038/nature00926. [DOI] [PubMed] [Google Scholar]
- 23.Atkin OK, Bruhn D, Hurry VM, Tjoelker MG. The hot and the cold: Unravelling the variable response of plant respiration to temperature. Funct Plant Biol. 2005;32:87–105. doi: 10.1071/FP03176. [DOI] [PubMed] [Google Scholar]
- 24.Brando PM, et al. Drought effects on litterfall, wood production and belowground carbon cycling in an Amazon forest: Results of a throughfall reduction experiment. Philos Trans R Soc London Ser B. 2008;363:1839–1848. doi: 10.1098/rstb.2007.0031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nepstad DC, Tohver IM, Ray D, Moutinho P, Cardinot G. Mortality of large trees and lianas following experimental drought in an amazon forest. Ecology. 2007;88:2259–2269. doi: 10.1890/06-1046.1. [DOI] [PubMed] [Google Scholar]
- 26.Fisher RA, et al. The response of an E. Amazonian rain forest to drought stress: Results and modelling analyses from a throughfall exclusion experiment. Global Change Biol. 2007;13:2361–2378. [Google Scholar]
- 27.Soares BS, et al. Modelling conservation in the Amazon basin. Nature. 2006;440:520–523. doi: 10.1038/nature04389. [DOI] [PubMed] [Google Scholar]
- 28.Nepstad DC, Stickler CM, Soares B, Merry F. Interactions among Amazon land use, forests and climate: Prospects for a near-term forest tipping point. Philos Trans R Soc London Ser B. 2008;363:1737–1746. doi: 10.1098/rstb.2007.0036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sawyer D. Climate change, biofuels and eco-social impacts in the Brazilian Amazon and Cerrado. Philos Trans R Soc London Ser B. 2008;363:1747–1752. doi: 10.1098/rstb.2007.0030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ramos da Silva R, Werth D, Avissar R. Regional impacts of future land-cover changes on the Amazon Basin wet-season climate. J Climate. 2008;21:1153–1170. [Google Scholar]
- 31.Barreto P, Souza CJ, Nogueron R, Anderson A, Salomao R. Human Pressure on the Brazilian Amazon Forests. Washington, DC: World Resources Institute; 2006. [Google Scholar]
- 32.Alencar AAC, Solorzano LA, Nepstad DC. Modeling forest understory fires in an Eastern Amazonian landscape. Ecol Appl. 2004;14:S139–S149. [Google Scholar]
- 33.Barlow J, Peres CA. In: Emerging Threats to Tropical Forests. Laurance WF, Peres CA, editors. Chicago: Chicago Univ Press; 2006. pp. 225–240. [Google Scholar]
- 34.Cochrane MA, Schulze MD. Fire as a recurrent event in tropical forests of the Eastern Amazon: Effects on forest structure, biomass, and species composition. Biotropica. 1999;31:2–16. [Google Scholar]
- 35.Nepstad D, et al. Inhibition of Amazon deforestation and fire by parks and indigenous lands. Conserv Biol. 2006;20:65–73. doi: 10.1111/j.1523-1739.2006.00351.x. [DOI] [PubMed] [Google Scholar]
- 36.Betts RA, Malhi Y, Roberts JT. The future of the Amazon: New perspectives from climate, ecosystem and social sciences. Philos Trans R Soc London Ser B. 2008;363:1729–1735. doi: 10.1098/rstb.2008.0011. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.