

Abstract
Background and purpose:
Large-conductance Ca2+-activated K+ channels (BKCa), located on the arterial and corporal smooth muscle, are potential targets for treatment of erectile dysfunction (ED). This study investigated whether NS11021 (1-(3,5-Bis-trifluoromethyl-phenyl)-3-[4-bromo-2-(1H-tetrazol-5-yl)-phenyl]-thiourea), a novel opener of BKCa channels, relaxes erectile tissue in vitro and enhances erectile responses in intact rats. The effects were compared with sildenafil, an inhibitor of phosphodiesterase type 5.
Experimental approach:
Patch clamp was used to record whole cell current in rat isolated corpus cavernosum smooth muscle cells (SMCs) and human umbilical vein endothelial cells (HUVECs). Isometric tension was measured in intracavernous arterial rings and corpus cavernosum strips isolated from rats and men, and simultaneous measurements of intracellular Ca2+ concentration ([Ca2+]i) and tension were performed in intracavernous arteries. Erectile response was measured in anaesthetized rats.
Key results:
In patch clamp recordings, NS11021 increased currents sensitive to the selective BKCa channel blocker, iberiotoxin (IbTX) in SMCs, but did not modulate K+ current in HUVECs. NS11021 reduced [Ca2+]i and tension in penile arteries. IbTX inhibited the vasorelaxation induced by NS11021 and sildenafil in human erectile tissue. NS11021 and sildenafil but not vehicle increased erectile responses in anaesthetized rats, an effect which was abolished after pretreatment with tetraethylammonium.
Conclusions and implications:
NS11021 leads to relaxation of both intracavernous arteries and corpus cavernosum strips primarily through opening of BKCa channels. It is also effective in facilitating erectile responses in anaesthetized rats. These results suggest a potential for use of BKCa openers in the treatment of ED.
Keywords: penile tissue, BKCa channel, NS11021, sildenafil, vasodilation, erection
Introduction
Male erectile dysfunction (ED) is defined by the inability to attain or maintain penile erection sufficient for satisfactory sexual intercourse. The prevalence of ED is increased in patients with ischaemic heart disease, diabetes and hypertension (Shiri et al., 2003; Montorsi et al., 2004; Vrentzos et al., 2007). One of the underlying causes of ED of vascular origin is reduced nitric oxide (NO) bioavailability resulting in insufficient relaxation of smooth muscle in the penis (Azadzoi et al., 2005; Moore and Wang, 2006). Phosphodiesterase type 5 (PDE5) inhibitors increase cyclic GMP levels in penile smooth muscle followed by smooth muscle relaxation and erection. However, PDE5 inhibitors are less efficient in patients with diabetes and they are contraindicated in all men taking nitrate compounds (Jackson et al., 2006). Therefore, novel pharmacological approaches are required for the treatment of ED in these patients.
Activation of K+ channels followed by hyperpolarization and vasodilatation of penile arteries and relaxation of corporal smooth muscle cells (SMCs) is thought to be an important mechanism in erection (Prieto, 2008). Both ATP-sensitive K+ channels (KATP) and big conductance calcium-activated (BKCa or KCa1.1) channels (nomenclature follows Alexander et al., 2008) are expressed in corporal smooth muscle (Fan et al., 1995; Lee and Kang, 2001; Spektor et al., 2002). The BKCa channels are directly involved in neurogenic NO-mediated relaxations in horse penile arteries (Simonsen et al., 1995), inhibition of myogenic tone of rat penile arteries (Kun et al., 2003) and relaxation to C-type natriuretic peptide in human penile arteries (Kun et al., 2008). Moreover, mice lacking BKCa channels have ED (Werner et al., 2005), and ageing is associated with ED and causes cytoplasmic retention of BKCa channels in rat corporal SMCs (Davies et al., 2007). Intra-cavernosal injection of the hSlo gene, which encodes the BKCa channel, enhances erectile responses in old and diabetic rats (Christ et al., 2004), hence suggesting that activation of BKCa channels may restore erectile function in patients with ED.
Both openers of BKCa channels (Spektor et al., 2002; Hewawasam et al., 2003; 2004; Boy et al., 2004) and of KATP channels (Holmquist et al., 1990; Venkateswarlu et al., 2002) were found to relax corpus cavernosum strips. However, the KATP channel openers, cromakalim and pinacidil, were mainly proposed for intracavernosal injection, since systemic administration was associated with hypotension and reflex tachycardia (Trigo-Rocha et al., 1995; Moon et al., 1999). Moreover, the selectivity of the benzimidazolone, NS1619 for BKCa channels has been questioned, since at high concentrations, NS1619 directly inhibits L-type Ca2+ channels in rat ventricular myocytes (Park et al., 2007), calcium-activated chloride currents (Saleh et al., 2007) and voltage-activated Ca2+, K+ and Na+ channels (Edwards et al., 1994; Holland et al., 1996). Recently, a novel BKCa channel opener NS11021 (1-(3,5-bis-trifluoromethyl-phenyl)-3-[4-bromo-2-(1H-tetrazol-5-yl)-phenyl]-thiourea) was found to be a more potent BKCa channel activator than NS1619 and specific, as it did not affect L- and T-type calcium currents in native guinea pig ventricular cardiomyocytes (Bentzen et al., 2007). Therefore, the present study proposed that NS11021 would relax erectile smooth muscle preparations and enhance erectile function. Thus, in the present study, we investigated the effect of NS11021 on: (i) K+ current in smooth muscle and endothelial cells; (ii) intra-cavernous arteries and corpus cavernosum strips isolated from rat and man in vitro; (iii) smooth muscle intracellular Ca2+ concentration ([Ca2+]i) and tension in intact penile arteries; and (iv) cardiovascular and erectile function in a rat model. The effect of NS11021 was compared to an inhibitor of PDE5, sildenafil. We found that NS11021 induces powerful relaxation in vitro and increases erectile function in vivo.
Methods
Animals
Animal care and experimental procedures were according to Danish Animal Law and regulations (permission 2007/561-1265). Adult male Wistar rats (12–16 weeks old) were kept and cared for in standard cages under clean conditions in separate quarters in a 12–12 h light–dark cycle with free access to water and food pellets.
Patch clamp on freshly isolated SMCs from rat corpus cavernosum
Strips of corpus cavernosum (about 1–2 mm long; see below for preparation) were stored overnight at 4°C in the solution for enzymatic digestion of the following composition (in mM): NaCl 110, KCl 5, MgCl2 2, KH2PO4 0.5, NaH2PO4 0.5, NaHCO3 10, CaCl2 0.16, EDTA 0.49, Na-HEPES 10, glucose 10, taurine 10, as well as 1.5 mg·mL−1 papain, 1.6 mg·mL−1 albumin, and 0.4 mg·mL−1 dl–dithiothreitol; pH was adjusted to 7.0. The next day, the tissue was incubated in the enzymatic solution for 5–10 min at 37°C, and the single SMCs were released by intense pipetting and after attachment to a dish, they were patched at room temperature (22–24°C). Patch pipettes (resistance 2–8 MΩ) were prepared from borosilicate glass (PG15OT- 7.5; Harvard Apparatus) pulled on a P-97 puller (Sutter Instrument Co.). Membrane currents were recorded in the whole-cell configuration with an Axopatch 200B amplifier (Axon Instruments, Inc.) and the filtered (1 kHz) data were sampled in Microcal Origin software v. 5.0 for Windows (Microcal Software, Inc.). Series resistance and capacitive current were compensated before each experiment. The current amplitudes (recorded in pA) were normalized to the cell capacitance (recorded in pF) and expressed as current density (A·F−1). The cells were held at a holding potential of −50 mV. Current–voltage characteristics were obtained from a ramp protocol (Figure 1A). Voltage ramps were performed by stepping initially from −50 mV to −100 mV for 100 ms and, then, depolarizing the whole cell from −100 mV to +80 mV with a duration of 700 ms.
Figure 1.
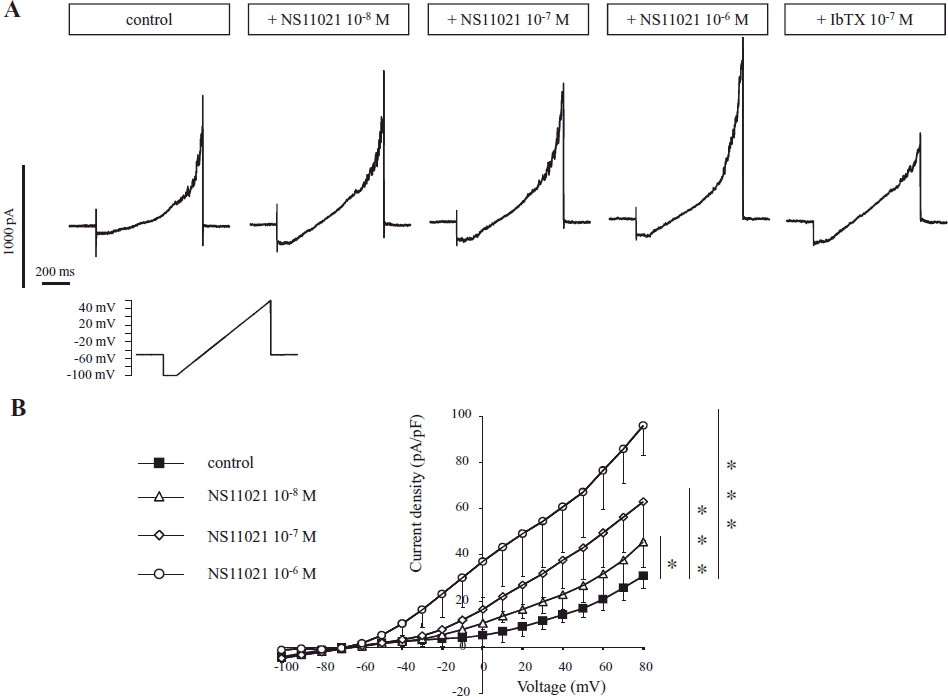
Patch clamp studies on smooth muscle cells from rat corpus cavernosum. Increasing concentrations of NS11021 (10−8, 10−7, 10−6 M) stimulate membrane conductances which were then significantly inhibited by IbTX. (A) Typical experimental recordings from smooth muscle cell (membrane capacitance 17 pF) consequently stimulated with increasing concentrations of NS11021 and, then IbTX (0.1 µM). Lower insert in (A) shows voltage ramp protocol used in these experiments. (B) shows average (n= 5, Cm = 19 ± 1 pF) current–voltage recordings of IbTX-sensitive currents under control conditions and after cumulative increase of NS11021. The current–voltage curves achieved by subtraction of resting IbTX-insensitive current obtained at the end of each experiment. *, ***P < 0.05, 0.001 versus control IbTX-sensitive current by two-way anova.
Calcium-activated potassium current was stimulated by dialysis of the cell with a pipette solution containing calcium buffered at ∼500 nM, a concentration corresponding to submaximal activation of BKCa channels (Sadoshima et al., 1988). Free [Ca2+]i in the pipette solution was estimated using WEBMAXC v. 2.22 (Chris Patton, Stanford University, CA, USA). The patch pipette solution contained (in mM): NaCl 10, KCl 122, K-HEPES 10, MgCl2 1, CaCl2 0.072, BAPTA 0.1 at pH 7.35. The bath solution contained (in mM): NaCl 135, KCl 6, Na-HEPES 10, MgCl2 1, CaCl2 0.1 at pH 7.4.
Patch clamp on human umbilical vein endothelial cells (HUVECs)
This part of the investigation conformed to the principles outlined in the Declaration of Helsinki and was approved by the local Ethics Committee of Region Midt (permission 20040154). Umbilical cords were obtained immediately after delivery. HUVECs were isolated as previously described (Ostergaard et al., 2007). In all experiments, cells were used at passages 2 or 3, where they did not express BKCa channels in qPCR studies (Krøigaard and Simonsen, unpublished observation). The endothelial cell phenotype was confirmed using phase-contrast microscopy (i.e. cuboidal, cobblestone-appearance monolayer of cells) and positive immuno-fluorescence staining with antibodies against von Willebrand factor (Ostergaard et al., 2007).
In HUVECs, membrane currents were measured using conventional whole-cell patch clamp techniques. HUVECs were dispersed using a non-enzymatic technique and plated on coverslips in a recording chamber containing physiological extracellular solution (PES) containing (in mM): NaCl 105, NaHCO3 25, KCl 4.7, KH2PO4 1.18, MgSO4 1.17, Na-HEPES 10, EGTA 0.026, glucose 5.5, pH 7.4 (with NaOH) to which 1.6 mM CaCl2 was added for Ca2+-containing PES. Recording electrodes (resistance 2–5 MΩ) were fabricated from borosilicate glass and were filled with physiological intracellular solution containing (in mM): potassium-L-aspartate 100, KCl 30, EGTA 10, MgCl2 1, Na2ATP 3, HEPES-potassium salt 5, pH 7.2 (with KOH). Membrane currents were recorded at room temperature using an EPC-10 amplifier (HEKA) filtered at 1 kHz, digitized at 10 kHz and stored on a PC for subsequent analysis. Current–voltage (IV) relationships were generated in voltage-clamped cells led at a membrane potential of −60 mV and stepped from −60 mV to +180 mV in 40 mV increments.
Functional in vitro experiments
Human erectile tissue
All patients provided informed consent and this study was in accordance with Ethical Committee, Country of Region Midtjylland, Denmark. Human corpus cavernosum tissues were obtained from 50- to 70-year-old patients with ED of various origins (diabetes, prostatic cancer or neurogenic origin). Cavernous tissue strips were retrieved at the time of penile prosthesis insertion. Immediately after removal, tissues were placed in physiological saline solution (PSS; see composition below) and kept at 4°C from the time of surgery until they were used in experiments, within 24 h.
Rat erectile tissue
The animals were killed humanely by cervical dislocation and exsanguinated after decapitation. After cutting the crura corpora cavernosa at the point of adhesion to the lower pubic bone, the penis was removed and submerged in ice-cold (4°C) PSS, and the corpus cavernosum was microsurgically dissected free.
Preparation of intracavernous artery
Ring segments (ca. 2 mm long) of intracavernous artery were dissected and mounted on two 40 µm wires of isometric double myograph (410A, Danish Myotechnology, Aarhus, Denmark) as previously described (Kun et al., 2003). The preparations were allowed to equilibrate in PSS solution, at 37°C, bubbled with a mixture of 5% CO2 in air O2 to maintain pH at 7.4. The relation between resting wall tension and internal circumference of the vessels was determined, and the internal circumference, L100, corresponding to a transmural pressure of 100 mm Hg in a relaxed vessel was calculated. Subsequently, the internal circumference of the vessels were set to L1, where L1= 0.9 × L100. The effective internal lumen diameter was determined as l1= L1/π.
Preparation of corpus cavernosum strips
Change in isometric tension of human (1 × 1 × 4 mm) and rat (0.5 × 0.5 × 3 mm) corpus cavernosum strips were investigated in a tissue organ bath system (750TOBS, Danish Myotechnology, Aarhus, Denmark). Silk ligatures were applied at both ends of the strip preparations, which were then suspended between two L-formed metal prongs in thermostatically controlled organ baths (5 mL, 37°C) containing PSS aerated with a mixture of 5% CO2 in air (21% O2, pH 7.4). During an equilibration period of 60 min, tension was adjusted until a mean stable tension of, respectively, 1.4 mN and 1.2 mN was obtained for human and rat corpus cavernosum strips, as previously described (Hedlund et al., 1999; Matsumoto et al., 2005).
Simultaneous measurements of [Ca2+]i and tension
Simultaneous measurements of smooth muscle [Ca2+]i and tension were performed in intact arteries as previously described (Villalba et al., 2007). The arteries were mounted in a myograph as described above and loaded in the dark in PSS containing 8 µM fura 2-acetoxymehyl ester (fura 2-AM) and 0.1% cremophor EL for 3 h at 37°C. The bath solution was replaced with fresh PSS and FURA-2-AM after 90 min. Fluorescence measurements were performed using an inverted microscope equipped for dual-excitation wavelength fluorimetry (Deltascan, Photon Technology International, NJ, USA). The arteries were excited with alternating wavelengths of 340 and 380 nm, and the intensity of the emitted fluorescence was collected at a wavelength of 510 nm using a photomultiplier connected to a computer running software (Felix, Photon Technology International, USA). The 340/380 fluorescence ratio (F340/F380) was taken as a measure of [Ca2+]i. Ca2+-insensitive signals were determined after quenching with Mn2+ (MnCl2) and subtracted.
Experimental procedure
To test contractility of the preparations, rat erectile tissues were exposed twice to 10 µM phenylephrine or 1 µM phenylephrine for human tissues. Arteries and corpus cavernosum strips were contracted with phenylephrine (1 µM), except human corpus cavernosum strips which were contracted with 0.5 µM phenylephrine, and cumulative concentration–response curves were constructed for NS11021 (0.1–100 µM), sildenafil (0.1 nM–30 µM) and vehicle for the drugs, or constructed in the presence of iberiotoxin (IbTX 0.1 µM), which was applied 30 min before obtaining the concentration-response curves and in a concentration causing full inhibition of BKCa channels (Galvez et al., 1990). Moreover, concentration–response curves for NS11021 were constructed in preparations contracted by 80 mM potassium-rich physiological salt solution, where NaCl was exchanged for KCl on an equivalent basis.
Electrical field stimulation (EFS)
EFS was performed with platinum electrodes (J.P. Trading, Aarhus, Denmark), which were secured in plastic heads on both sides of the mounted segment, placed approximately 2 mm from the strips. The electrodes were connected to an electrical stimulator (CS200, Danish Myotechnology, Aarhus, Denmark) with submaximal constant current output. To examine the influence of drugs on neurogenic contractions, the preparations were incubated with asymmetric dimethylarginine (ADMA, 300 µM) and propranolol (1 µM) to inhibit concomitant NO release and β-adrenoceptors respectively (Kun et al., 2003). The cumulative relaxant effect of drugs was investigated in strips contracted with 16 Hz EFS (supra-maximal current) applied with 4 min intervals.
In vivo experiments
Measurements of intracavernous and mean arterial pressure (MAP)
The rats were anaesthetized with pentobarbital sodium (Sygehus Apoteket, Aarhus, Denmark; 50 mg·kg−1; i.p.). During the experiment, the rats breathed spontaneously; the body temperature was monitored continuously and was maintained at 37°C. A heparinized (100 IE·mL−1) polyethylene catheter (PE 50) was introduced into the carotid artery to measure MAP. A 25 gauge needle attached to a heparinized polyethylene catheter was inserted into the crus corpus cavernosum to measure intracavernous pressure (ICP). Continuous direct measurements of MAP and ICP were performed with transducers (Disposable BP Transducer, ADInstruments, UK), and registered and analysed on a computerized data acquisition system (PowerLab, ADInstruments). A stabilizing period of 20–30 min was allowed before registration of basal ICP and MAP.
The cavernous nerve was isolated through a lower abdominal incision, and electrical stimulation was performed with a slender bipolar platinum electrode, which was connected to a S48 stimulator (Grass Instrument Co., Boston, MA, USA). NS11021, sildenafil and vehicle were administered by injection in the jugular vein.
Experimental procedure
A first stimulation of the cavernous nerve at parameters eliciting the maximum amplitude of the erectile response (square wave pulses of 6 Volts, 10 Hz, 1 ms for 30 s) was performed, as described earlier (Giuliano et al., 2003). Electrical parameters eliciting a submaximal erectile response were then sought between 0.6 and 1.55 Volts, other parameters remaining unchanged. Both NS11021 and sildenafil were injected at the doses of 0.1 and 1 mg·kg−1 intravenously in volumes of maximum 200 µL. One group was injected with vehicle alone. Submaximal electrical stimulation was applied 3, 13 and 23 min after the injection. At the end of each experiment, the maximal erectile response was repeated to ensure the cavernous nerve was intact.
To investigate whether the BKCa channels are involved in the effect of NS11021 in vivo, a first series of submaximal erectile responses was obtained, 6 mg·kg−1 tetraethylammonium (TEA) was injected intravenously, and after 30 min, 1 mg·kg−1 NS11021 was injected intravenously and the responses to submaximal electrical stimulation were repeated.
For each electrically induced erectile response, the ratio peak intracavernosal pressure (PICP) (mm Hg)/MAP (mm Hg) × 100 was measured, with PICP being the peak value reached by ICP during the erectile response and the MAP during the erectile response.
Analysis of data
The mechanical responses of the vessels were measured as force and expressed as active wall tension, ΔT, which is the increase in measured force, ΔF, divided by twice the segment length. The magnitude of relaxant responses is given as percentage of the contraction level just prior to the addition of the drug. The effects of NS11021 and sildenafil on EFS evoked contractions are expressed as percentages of previous control contractions
The results are expressed as means ± SEM and the concentration–response curves are presented on a semi-logarithmic scale. Differences between mean pD2 (calculated as −log EC50) and magnitude of responses were analysed using paired or unpaired t-test as indicated. n denotes the number of preparations. Differences in concentration–response relationships between treatments were analysed using two-way analysis of variance. To analyse the in vivo results, post hoc analysis was performed using Dunnett's test versus the control response. Differences at the P < 0.05 level were considered statistically significant.
Materials
The composition of PSS was as follows (in mM): NaCl 119, NaHCO3 25, KCl 4.7, KH2PO4 1.18, MgSO4 1.17, CaCl2 2.5, EDTA 0.026 and glucose 5.5 (pH 7.4). The 80 mM K+ solution had the same composition as PSS, but with NaCl replaced by KCl on an equimolar basis to attain the final 80 mM K+ concentration.
The following drugs were used: NS11021 (NeuroSearch A/S, Ballerup, Denmark), sildenafil (gift from Pfizer Ltd.), ADMA, IbTX, phenylephrine hydrochloride, propranolol hydrochloride and tetraethylammonium chloride (Sigma, St. Louis, MO, USA). NS11021 was dissolved in DMSO, and then diluted in water. The maximal DMSO concentration (0.1%) applied in vitro did not modulate smooth muscle tone in control experiments. All other drugs were dissolved in distilled water. For in vivo experiments, NS11021 stock solution was diluted in sterile 0.9% NaCl; sildenafil was dissolved in sterile 0.9% NaCl.
Results
Patch clamp electrophysiology
The effect of NS11021 on the membrane currents of freshly isolated SMCs from corpus cavernosum was tested using conventional whole-cell patch clamp technique (Figure 1). Initially, an outward rectifying membrane current with reversal potential at −21.5 ± 4.5 mV was observed. Cumulative addition of NS11021 increased whole-cell membrane current, primarily in the outward direction (Figure 1A). The current reversal potential in the presence of 10−8, 10−7, 10−6 M NS11021 was shifted to the left (−28.0 ± 5.8 mV, −36.9 ± 3.7 mV and −48.0 ± 4.6 mV respectively). Addition of IbTX (0.1 µM) inhibited membrane conductance in the outward direction. As the K+ ion in the current solution composition (see Methods) was the only ion with negative equilibrium potential (−79 mV), we suggested that NS11021 stimulated primarily BKCa conductance which was inhibited by IbTX. When the resting IbTX-insensitive current was subtracted from the whole-cell conductance, a significant potentiation of IbTX-sensitive BKCa current with NS11021 was observed (Figure 1B). The reversal potentials of IbTX-sensitive currents were −62 ± 14 mV, −63 ± 8 mV, −65 ± 7 mV and −71 ± 6 mV under control conditions and in the presence of 10−8, 10−7, 10−6 M of NS1102 respectively.
To test the hypothesis that NS11021 specifically activates BKCa current, SMCs were pretreated with IbTX (Figure 2) which significantly inhibited outward current (Figure 2A). The subsequent application of 10−8 and 10−7 M NS11021 had no effect on the membrane conductance but 10−6 M NS11021 potentiated current in the outward direction (Figure 2B), an effect inhibited by tetraethylammonium (TEA, 1 mM).
Figure 2.
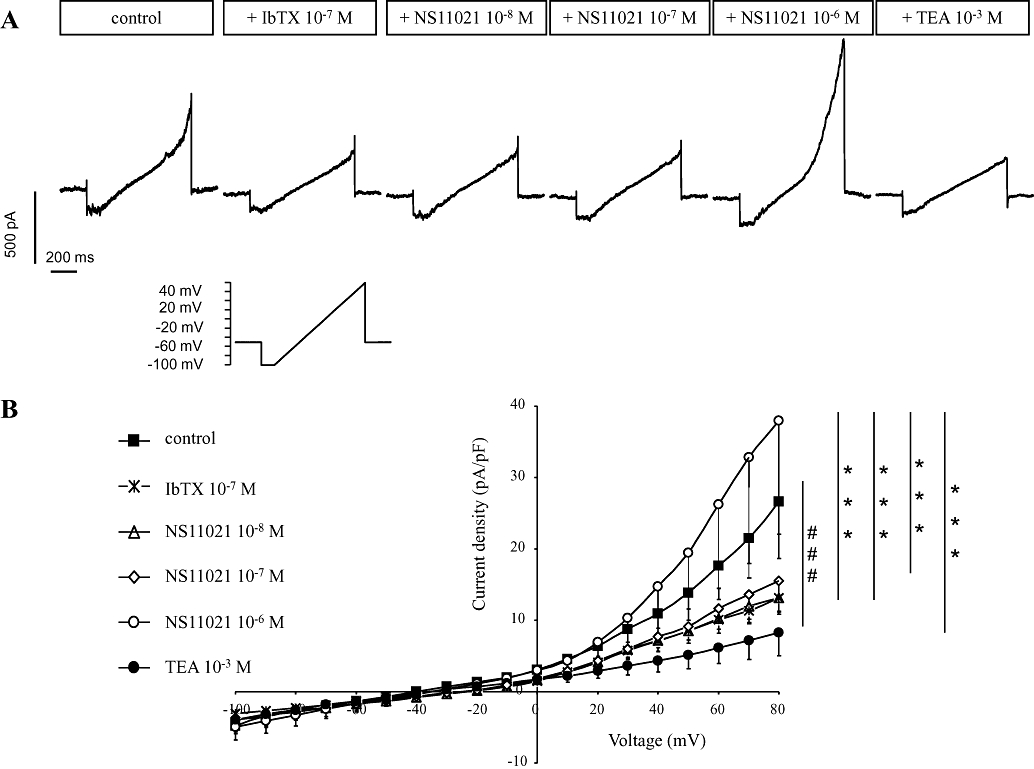
Patch clamp studies on smooth muscle cells from rat corpus cavernosum. Low concentrations (10−8 and 10−7 M) of NS11021 stimulate only IbTX-sensitive potassium current (see also Figure 1). Pretreatment with 0.1 µM IbTX inhibits initial of membrane conductances with low concentrations (10−8 and 10−7 M) of NS11021. A high concentration NS11021 (10−6 M) stimulates, however, another outward rectifying conductance which is sensitive to tetraethylammonium (TEA, 1 mM). (A) Representative experimental recordings (membrane capacitance 25 pF) and (B) averaged results (n= 5, Cm = 16 ± 3 pF) shown as current–voltage curves. ###P < 0.001 versus control current by two-way anova. ***P < 0.001 versus current in the presence of 10−6 M NS11021 by two-way anova.
To investigate the effect of NS11021 on membrane potential in HUVECs, the current–voltage relationships were determined in voltage-clamped endothelial cells. NS11021 (1 µM) did not modulate the membrane current in HUVEC (Figure 3, n= 5).
Figure 3.
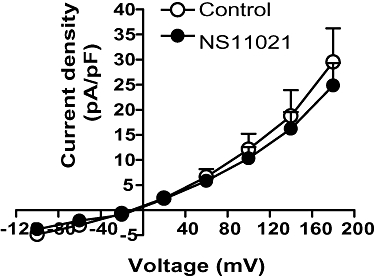
Average effect of NS11021 (10−6 M) on the current–voltage relationships of voltage-clamped human umbilical vein endothelial cells (HUVECs, n= 5).
Effect of NS11021 on [Ca2+]i
In phenylephrine(1 µM)-contracted arteries, NS11021 evoked concentration-dependent relaxations and lowered [Ca2+]i (Figure 4A). In arteries contracted by 125 mM K+, higher concentrations of NS11021 were required to lower calcium and relax the preparations (Figure 4B), and the concentration–response curves for NS11021 were markedly shifted to the right in preparations contracted by 125 mM K+ compared with those contracted by phenylephrine (Figure 4C).
Figure 4.
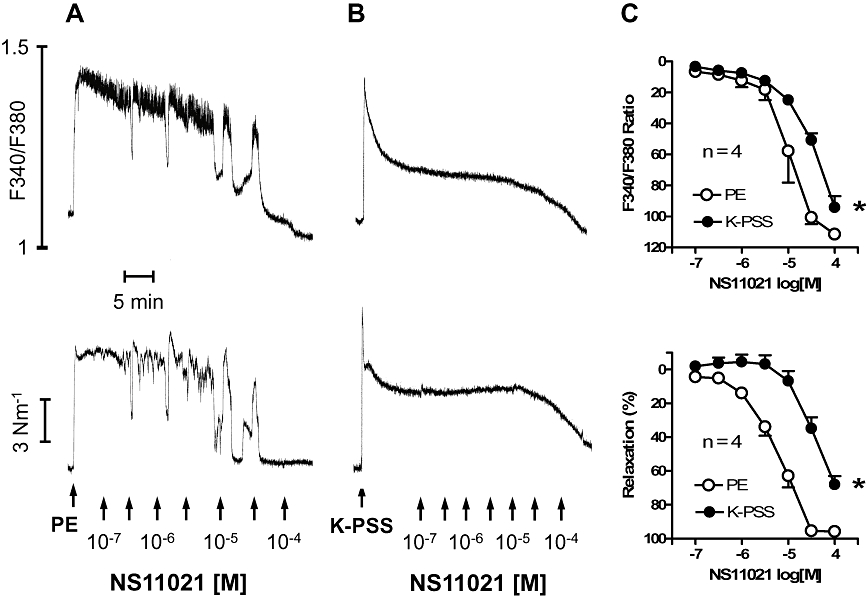
Effects of NS11021 on [Ca2+]i and tension in rat intracavernous artery. Effect of NS11021 in (A) phenylephrine (PE, 1 µM)- and (B) 125 mM K+-contracted arteries. (C) Average data showing the decrease in [Ca2+]i and tension in PE-and K-PSS-contracted preparations in response to cumulative administration of NS11021 (n= 4). *P < 0.05 by two-way anova.
In vitro investigations on rat tissues
In penile arteries with internal diameters of 195 ± 5 µM (n= 24) and corpus cavernosum strips isolated from rat penile tissue, phenylephrine (10 µM) induced contractions of, respectively, 2.1 ± 0.1 N·m−1 (n= 24) and 4.5 ± 0.2 mN (n= 87).
NS11021 (0.1–100 µM) relaxed phenylephrine-contracted (1 µM) penile arteries and corpus cavernosum strip preparations in a concentration-dependent manner (Figure 5A and B respectively) (n= 6). The pD2 values for NS11021 in phenylephrine-activated penile arteries and corpus cavernosum preparations were 5.1 ± 0.1 and 4.8 ± 0.1 respectively (n= 6). In arteries and corpus cavernosum strips contracted with 80 mM K+ or 80 mM K+ plus 1 µM phenylephrine, NS11021 relaxation was reduced, compared with preparations contracted with 1 µM phenylephrine (Figure 5A and B) (n= 4). The pD2 values in intracavernous artery and corpus cavernosum preparations contracted with 80 mM K+ were 4.4 ± 0.1 and 4.0 ± 0.1 respectively (n= 4). Vehicle had no effect (Figure 5A and B) (n= 6).
Figure 5.
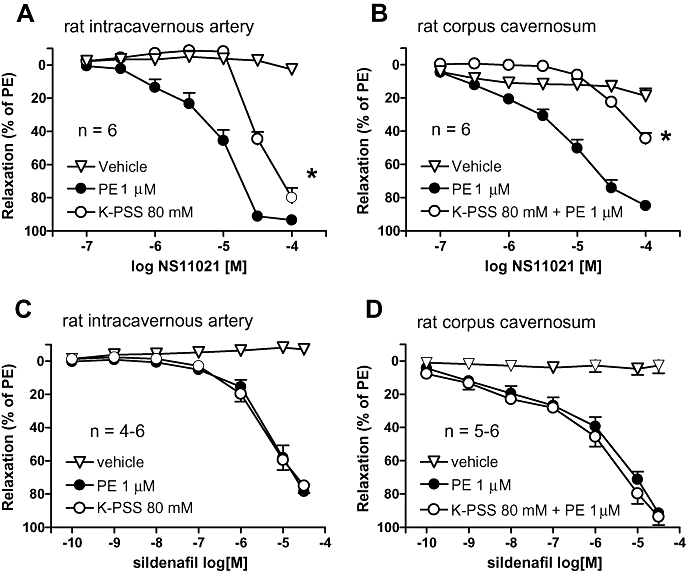
Average NS11021 (A, B) and sildenafil (C, D) induced relaxation in phenylephrine (PE)- contracted and high potassium (125 mM K+)-contracted intracavernous artery (A, C, n= 4–6) and corpus cavernosum strips (B, D, n= 5–6) isolated from rat penis. *P < 0.05 by two-way anova.
Sildenafil (0.1 nM–30 µM), an inhibitor of PDE5, induced relaxations in both rat penile arteries and corpus cavernosum strips contracted with 1 µM phenylephrine as well as in 80 mM K+-activated preparations (Figure 5C and D). The pD2 values for sildenafil in penile arteries contracted with 1 µM phenylephrine and 80 mM K+ were, respectively, 5.2 ± 0.1 and 5.5 ± 0.1 (n= 4) and in corpus cavernosum strips, respectively, 5.7 ± 0.1 and 6.0 ± 0.1 (n= 5). The vehicle of sildenafil had no effect in these preparations (Figure 5C and D).
In corpus cavernosum preparations, incubated with ADMA (300 µM) and propranolol (1 µM), EFS (16 Hz) produced reproducible, frequency-dependent contractions producing an average force of 3.4 ± 0.2 mN (n= 15). Prazosin (1 µM) inhibited the EFS-evoked contractions by 99.9 ± 0.1% (n= 3, data not shown). NS11021 induced concentration-dependent (0.1–100 µM) inhibition of EFS-induced contractions. The maximal inhibitory effect of 89 ± 5% was obtained at 100 µM NS11021 (n= 3) (Figure 6A). The pD2 value for NS11021 on contractions produced by EFS was 4.7 ± 0.3 (n= 3). Sildenafil (0.1 nM–30 µM) induced almost full inhibition, similar to NS11021, on EFS-induced contractions with a maximum of 84 ± 3% (n= 4) (Figure 6B). The pD2 value for sildenafil was 5.1 ± 0.1 (n= 4). Vehicle did not affect the EFS-evoked contractions.
Figure 6.
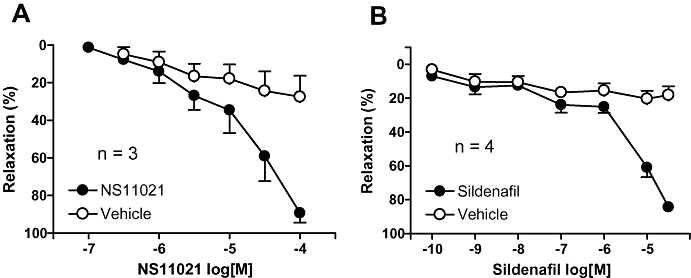
Average effect of (A) NS11021, (B) sildenafil, and vehicle on electrical field stimulation-evoked neurogenic contractions in rat corpus cavernosum strips (n= 3–4).
In vitro investigations on human tissues
In intracavernous arteries (with internal diameter of 202 ± 21 µM, n= 2), phenylephrine (10 µM) increased active tension by 4.1 ± 1.0 Nm−1 (n= 2). In two representative experiments, NS11021 and sildenafil induced concentration-dependent relaxations in penile small intracavernous arteries isolated from human tissue, while vehicle failed to alter phenylephrine contraction (data not shown).
In corpus cavernosum strips, phenylephrine (1 µM) increased force by 20.5 ± 1.8 mN (n= 16). NS11021 (0.1–100 µM) induced IbTX-sensitive concentration-dependent relaxations in phenylephrine (0.5 µM)-contracted preparations (Figure 7A). The pD2 values for NS11021 in the absence and presence of IbTX were 4.5 ± 0.1 and 4.3 ± 0.1 respectively (n= 4). Sildenafil-induced relaxations were also inhibited by IbTX (Figure 7B, n= 3). pD2 values for sildenafil in the absence and presence of IbTX were 7.0 ± 0.1 and 5.2 ± 0.2.
Figure 7.
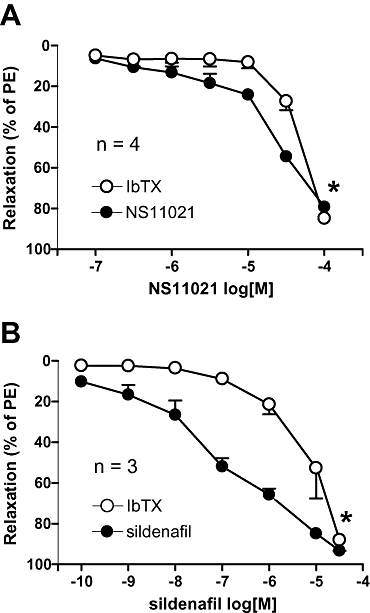
Average NS11021 (A, n= 4) and sildenafil (B, n= 3) induced relaxations in phenylephrine (PE, 0.5 µM)-contracted human corpus cavernosum strips in the presence and absence of iberiotoxin (IbTX, 0.1 µM). *P < 0.05 by two-way anova.
In vivo experiments
Original traces show the experimental procedure (Figure 8A) and the magnification of an erectile response to electrical stimulation (Figure 8B). Administration of vehicle did not influence the erectile response or blood pressure (Figure 9A, n= 5), but sildenafil (0.1 mg·kg−1) slightly increased, while a higher dose of sildenafil (1 mg·kg−1) resulted in a significant increase of the erectile response compared with the control response (Figure 9B, n= 5). Erectile responses in anaesthetized rats were significantly increased after injection of both 0.1 and 1 mg·kg−1 NS11021 (Figure 9C, n= 5). A non-selective blocker of KCa channels, TEA was applied as intravenous injection (6 mg·kg−1), and caused an initial decrease in erectile responses as well as in MAP, but at the third electrical stimulation, the erectile response and MAP had recovered to control values (Figure 9D, n= 4). However, the facilitating effect of NS11021 (1 mg·kg−1) on erectile responses was abolished in the presence of TEA (Figure 9D).
Figure 8.
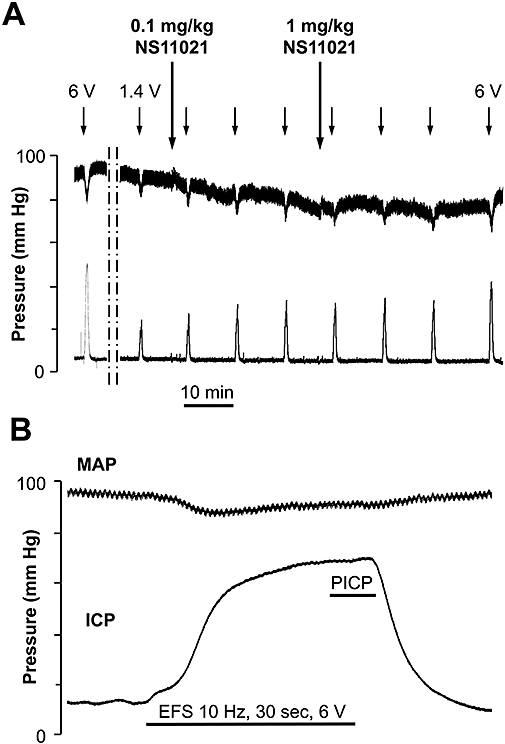
(A) Original trace of mean arterial pressure and intra-cavernous pressure recordings obtained in male Wistar rat anaesthetized with pentobarbital. NS11021 (0.1 and 1 mg·kg−1) was administered (↓) after establishing a maximal (10 Hz, 1 ms, 6 V) and submaximal response (10 Hz, 1 ms, 0.6 to 1.55 V) to cavernous nerve stimulation for 30 seconds. Submaximal stimulation was repeated 3, 13 and 23 min after drug administration. Upper trace shows the mean arterial pressure (MAP), lower trace represents the intra-cavernous pressure (ICP). (B) Represent a magnification of an erectile response induced by electrical field stimulation (EFS) of the cavernous nerve. The plateau response considered for calculation of the peak intracavernous pressure (PICP) during the erectile response. Erectile response was calculated by PICP(mm Hg)/MAP(mm Hg) × 100.
Figure 9.
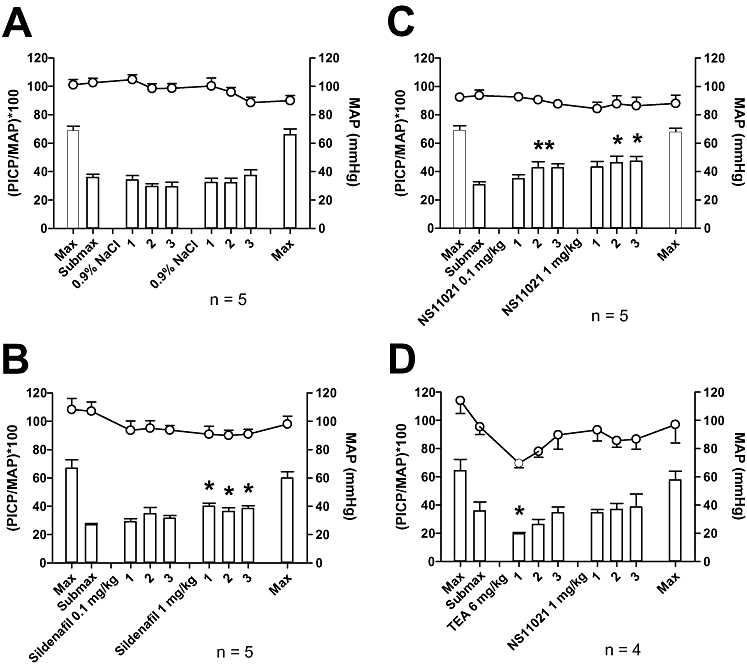
Average effect of vehicle (A), sildenafil (B) and NS11021 (C) on mean arterial pressure and erectile response obtained in male Wistar rats anaesthetized with pentobarbital. Drugs or vehicle were administered after establishing the maximal (10 Hz, 1 ms, 6 V) and submaximal responses (10 Hz, 1 ms, 0.6 to 1.55 V) to cavernous nerve stimulation for 30 s. Submaximal stimulation was repeated 3, 13 and 23 min after drug or vehicle administration. (D) Effect of intravenous tetraethylammonium (TEA, 6 mg·kg−1) pre-treatment on mean arterial pressure, erectile response and on effect of 1 mg·kg−1 NS11021. Upper traces show the mean arterial pressure (MAP), bars represent the erectile response ((PICP/MAP)*100), where PICP is the peak intracavernous pressure. *P < 0.05 compared with submaximal control response using one-way anova followed by Dunnett's test.
NS11021 (0.1 mg·kg−1) did not change MAP, but a higher dose (1 mg·kg−1) induced a transient drop in MAP (maximum fall of 9.0 ± 2.5 mmHg, n= 5), which recovered to pre-injection values after 41.8 ± 15.2 s (data not shown). TEA prevented the transient drop in blood pressure after administration of 1 mg·kg−1 NS11021 (data not shown). Sildenafil (0.1 mg·kg−1) induced a sustained drop in MAP of 24.9 ± 1.4 mm Hg, while a higher dose (1 mg·kg−1) caused a transient decrease in blood pressure of 27.3 ± 4.7 mm Hg (n= 5).
Discussion
The main findings of the present study are that NS11021, a novel selective opener of BKCa channels, increased IbTX-sensitive potassium currents in vascular smooth muscle, induced potent relaxations of penile arteries and corpus cavernosum strips, and facilitated erectile responses in anaesthetized rats. The magnitude of the effect of NS11021 was comparable with that of sildenafil, a potent inhibitor of PDE5.
NS11021 was recently found to activate BKCa channels in single-channel recordings and two-electrode voltage clamp electrophysiological recordings of monoclonal HEK293 cells (Bentzen et al., 2007). In our experiments, NS11021 increased the K+ currents in a concentration-dependent manner (10 nM–1 µM) in rat freshly isolated corpus cavernosum SMCs, and IbTX inhibited the effect of low NS11021 concentrations (≤1 µM), suggesting NS11021 increases BKCa current. NS11021 up to 1 µM did not change membrane current in HUVECs, suggesting that the effect on BKCa channels is specific. However, higher concentrations of NS11021 (≥1 µM) activated an additional outward rectifying potassium current which was sensitive to TEA (1 mM). Low and intermediate TEA concentrations (0.1–3 mM) can also block voltage-gated K+ channels of the KV7.2 and KV7.4 sub-type (Hadley et al., 2000). Moreover, in oocyte studies, NS11021 (30 µM) was found to increase voltage-gated KV7.4 current, an effect which was blocked by TEA (Bentzen et al., 2007). Therefore, the present patch clamp experiments suggest that at submicromolar concentrations, NS11021 increases BKCa current while at higher concentrations, activation of voltage-gated K+ current may also contribute to the effect of NS11021 in rat penile smooth muscle.
In the present study, NS11021-evoked maximal relaxation was inhibited but not abolished contraction induced by high extracellular potassium concentation in intracavernous arteries and corpus cavernosum strips. Moreover, the reduction in [Ca2+]i and tension was inhibited in the presence of high potassium solution only at low concentrations of NS11021 (<10 µM). Although in guinea pig ventricular cardiomyocytes, NS11021 had no effect on L- and T-type calcium currents (Bentzen et al., 2007), we can not exclude the possibility of modulation of Ca2+ currents in cavernosal SMCs. Therefore, in addition to the effect on BKCa channels at submicromolar concentrations, on KV channels at supramicromolar concentrations, blocking of calcium entry appear to contribute to the relaxant effect of high concentrations (>10 µM) of NS11021.
As mentioned in the Introduction, several lines of evidence support an important role for BKCa channels in erection (Prieto, 2008) and putative openers of BKCa channels, NS1619, 4-aryl-3-(hydroxyalkyl)quinolin-2-ones and 3-thio-quinolinones applied in drug concentrations of 10 µM have previously been found to relax corpus cavernosum strips by 20–50% (Spektor et al., 2002; Hewawasam et al., 2003; Boy et al., 2004). In the present study, NS11021 (1–10 µM) induced significant relaxations both in intracavernous arteries and corpus cavernosum strips. Moreover, IbTX induced a rightward shift in the concentration–response curves for NS11021 in human corpus cavernosum strips. These observations suggest that NS11021 is a potent and selective opener of BKCa channels in erectile smooth muscle.
Sildenafil, by inhibition of PDE5, increases cyclic GMP leading to activation of cyclic GMP-dependent protein kinase which, through phosphorylation, leads to increased Ca2+-ATPase activity and sarcoplasmatic calcium uptake, activation of myosin light chain phosphatase and opening of BKCa channels (Simonsen et al., 2002; Prieto, 2008). Thus, the relaxant effect of sildenafil in horse penile resistance arteries is due in part to cyclic GMP accumulation and to the increased effects of basally released endothelial NO (Prieto et al., 2006b), and sildenafil relaxes rabbit corpus cavernosum smooth muscle by amplifying the effects of the endogenous cyclic GMP-dependent relaxation mechanisms (Chuang et al., 1998). In previous studies, high K+ solutions and IbTX inhibited the relaxation evoked by sildenafil in horse penile small arteries and rabbit corpus cavernosum strips (Prieto et al., 2006a; El-Metwally et al., 2007), and sildenafil enhanced the NO-induced activation of BKCa channels in SMCs of human corpus cavernosum (Lee and Kang, 2001). The present results also suggest that the relaxant responses elicited by sildenafil involve the activation of BKCa channels in both intra-cavernous artery and human corpus cavernosum strips, as sildenafil relaxations were inhibited in the presence of IbTX. However, in the present study, high K+ solutions inhibited NS11021 but not sildenafil-induced relaxation in rat penile arteries and corpus cavernosum strips, and IbTX did not modify sildenafil relaxation in rat erectile tissue (data not shown). These findings suggest that there are species differences with regard to involvement of BKCa channels in sildenafil relaxation in erectile tissue.
In diabetes, smooth muscle relaxation mechanisms are often impaired and vasoconstriction in erectile tissue increased (Sáenz de Tejada et al., 2005; Wingard et al., 2007). α-Adrenoceptor-mediated contraction maintains the penis in the flaccid state (Simonsen et al., 2002). In the present study, blockade of EFS-evoked contractions by prazosin also supported the proposal that the neurogenic contraction of corpus cavernosum was elicited by activation of post-junctional α1-adrenoceptors. Both sildenafil and NS11021 caused inhibition of neurogenic contractions in erectile tissue in the present study. Therefore, in addition to the augmenting effect of NS11021 in basally released NO-mediated activation of the BKCa channels (Taniguchi et al., 1993), inhibition of α-adrenoceptor-mediated contraction due to direct activation of BKCa channels may also play a role in the relaxant effect of NS11021.
In anaesthetized rats, electrical stimulation of the cavernous nerve induces erection and increases the intracavernosal pressure. However, the PICP during the erectile response and MAP are correlated; therefore, increases in intracavernosal pressure are expressed as the ratio of the PICP over the corresponding MAP during the response (Matsumoto et al., 2005). This allows analysis of the effect of the drug treatment on the erectile response independently of their effect on blood pressure. In our experiments, NS11021, administered intravenously in 0.1 mg·kg−1 and 1 mg·kg−1 doses, significantly increased the erectile response in rats without changing MAP. Moreover, TEA abolished the effect of NS11021 1 mg·kg−1 on the erectile response. Although TEA can block several KCa channels subtypes (Sheng and Braun, 2007), the role of BKCa channels in the regulation of the circulation in vivo, investigated by TEA, is described in different animal (Eduardo da Silva-Santos and Assreuy, 1999), as well as in human studies (Bellien et al., 2005). Taken together with the findings that IbTX blocks NS11021 in vitro, our results suggest that NS11021 enhances erectile function in intact animals primarily through activation of BKCa channels.
In the present study, sildenafil was able to facilitate significantly the erectile response only at the 1 mg·kg−1 dose. These findings agree with previous studies in rats where erectile responses were also unchanged after administration of 0.1 mg·kg−1 dose of sildenafil (Giuliano et al., 2003). The in vitro results suggest that NS11021 and sildenafil are equipotent, and taking the difference in molar weight of NS11021 (367 g·mol−1) and sildenafil (474.6 g·mol−1) into account, the in vivo effect on erection of the two compounds is also of comparable magnitude.
Augmented contractility and/or impaired relaxation of arterial and corporal smooth muscle are a primary cause of ED. The fact that NS11021, an activator of BKCa channels, enhanced the erectile response in rats in vivo is consistent with previous findings showing increased erectile responses in old and diabetic rats after intracavernosal injection of the hSlo gene, which encodes the BKCa channel (Christ et al., 2004). Moreover, altered BKCa channels function may contribute to the development of other vascular diseases, such as hypertension, diabetes and atherosclerosis (Sobey, 2001). Therefore, activators of this channel may have a therapeutic potential in treatment of ED and cardiovascular disease.
In summary, the present study provides evidence suggesting that NS11021, a novel opener of BKCa channels, causes potent relaxations of erectile smooth muscle in vitro and facilitates erectile response in vivo, at doses devoid of cardiovascular side effects. These responses are comparable to those elicited by sildenafil, suggesting a potential for use of BKCa openers in the treatment of ED.
Acknowledgments
The authors are grateful to Dr. Petter Hedlund, University of Lund for his help with the in vivo experiments. The authors thank Kristina Litvin for technical assistance. U. Simonsen was supported by a grant from the Danish Medical Research Council.
Glossary
Abbreviations:
- [Ca2+]i
intracellular Ca2+ concentration
- ADMA
asymmetric dimethylarginine
- BKCa channel
big-conductance calcium-activated potassium channel
- DMSO
dimethylsulfoxide
- HUVEC
human umbilical endothelial cells
- IbTX
iberiotoxin
- KATP
ATP-sensitive potassium channel
- MAP
mean arterial blood pressure
- NS11021
1-(3,5-bis-trifluoromethyl-phenyl)-3-[4-bromo-2-(1H-tetrazol-5-yl)-phenyl]-thiourea
- PDE5
phosphodiesterase type 5
- PICP
peak intracavernosal pressure
- TEA
tetraethylammonium
Conflict of interest
U. Simonsen was supported by a grant from Neurosearch A/S, Copenhagen, Denmark.
References
- Alexander SP, Mathie A, Peters JA. Guide to receptors and channels (GRAC), 3rd edition. Br J Pharmacol. 2008;153(Suppl. 2):S1–S209. doi: 10.1038/sj.bjp.0707746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azadzoi KM, Schulman RN, Aviram M, Siroky MB. Oxidative stress in arteriogenic erectile dysfunction: prophylactic role of antioxidants. J Urol. 2005;174:386–393. doi: 10.1097/01.ju.0000161209.39959.67. [DOI] [PubMed] [Google Scholar]
- Bellien J, Joannides R, Iacob M, Arnaud P, Thuillez C. Calcium-activated potassium channels and NO regulate human peripheral conduit artery mechanics. Hypertension. 2005;46:210–216. doi: 10.1161/01.HYP.0000165685.83620.31. [DOI] [PubMed] [Google Scholar]
- Bentzen BH, Nardi A, Calloe K, Madsen LS, Olesen SP, Grunnet M. The small molecule NS11021 is a potent and specific activator of Ca2+-activated big-conductance K+ channels. Mol Pharmacol. 2007;72:1033–1044. doi: 10.1124/mol.107.038331. [DOI] [PubMed] [Google Scholar]
- Boy KM, Guernon JM, Sit SY, Xie K, Hewawasam P, Boissard CG, et al. 3-Thio-quinolinone maxi-K openers for the treatment of erectile dysfunction. Bioorg Med Chem Lett. 2004;14:5089–5093. doi: 10.1016/j.bmcl.2004.07.080. [DOI] [PubMed] [Google Scholar]
- Christ GJ, Day N, Santizo C, Sato Y, Zhao W, Sclafani T, et al. Intracorporal injection of hSlo cDNA restores erectile capacity in STZ-diabetic F-344 rats in vivo. Am J Physiol Heart Circ Physiol. 2004;287:H1544–H1553. doi: 10.1152/ajpheart.00792.2003. [DOI] [PubMed] [Google Scholar]
- Chuang AT, Strauss JD, Murphy RA, Steers WD. Sildenafil, a type-5 CGMP phosphodiesterase inhibitor, specifically amplifies endogenous cGMP-dependent relaxation in rabbit corpus cavernosum smooth muscle in vitro. J Urol. 1998;160:257–261. [PubMed] [Google Scholar]
- Davies KP, Stanevsky Y, Tar MT, Chang JS, Chance MR, Melman A. Ageing causes cytoplasmic retention of MaxiK channels in rat corporal smooth muscle cells. Int J Impot Res. 2007;19:371–377. doi: 10.1038/sj.ijir.3901541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eduardo da Silva-Santos J, Assreuy J. Long-lasting changes of rat blood pressure to vasoconstrictors and vasodilators induced by nitric oxide donor infusion: involvement of potassium channels. J Pharmacol Exp Ther. 1999;290:380–387. [PubMed] [Google Scholar]
- Edwards G, Niederste-Hollenberg A, Schneider J, Noack T, Weston AH. Ion channel modulation by NS 1619, the putative BKCa channel opener, in vascular smooth muscle. Br J Pharmacol. 1994;113:1538–1547. doi: 10.1111/j.1476-5381.1994.tb17171.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Metwally MA, Sharabi FM, Daabees TT, Senbel AM, Mostafa T. Involvement of alpha-receptors and potassium channels in the mechanism of action of sildenafil citrate. Int J Impot Res. 2007;19:551–557. doi: 10.1038/sj.ijir.3901590. [DOI] [PubMed] [Google Scholar]
- Fan SF, Brink PR, Melman A, Christ GJ. An analysis of the Maxi-K+ (KCa) channel in cultured human corporal smooth muscle cells. J Urol. 1995;153:818–825. [PubMed] [Google Scholar]
- Galvez A, Gimenez-Gallego G, Reuben JP, Roy-Contancin L, Feigenbaum P, Kaczorowski GJ, et al. Purification and characterization of a unique, potent, peptidyl probe for the high conductance calcium-activated potassium channel from venom of the scorpion Buthus tamulus. J Biol Chem. 1990;265:11083–11090. [PubMed] [Google Scholar]
- Giuliano F, Bernabe J, Alexandre L, Niewoehner U, Haning H, Bischoff E. Pro-erectile effect of vardenafil: in vitro experiments in rabbits and in vivo comparison with sildenafil in rats. Eur Urol. 2003;44:731–736. doi: 10.1016/s0302-2838(03)00377-4. [DOI] [PubMed] [Google Scholar]
- Hadley JK, Noda M, Selyanko AA, Wood IC, Abogadie FC, Brown DA. Differential tetraethylammonium sensitivity of KCNQ1-4 potassium channels. Br J Pharmacol. 2000;129:413–415. doi: 10.1038/sj.bjp.0703086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedlund P, Alm P, Andersson KE. NO synthase in cholinergic nerves and NO-induced relaxation in the rat isolated corpus cavernosum. Br J Pharmacol. 1999;127:349–360. doi: 10.1038/sj.bjp.0702556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hewawasam P, Fan W, Ding M, Flint K, Cook D, Goggins GD, et al. 4-Aryl-3-(hydroxyalkyl)quinolin-2-ones: novel maxi-K channel opening relaxants of corporal smooth muscle targeted for erectile dysfunction. J Med Chem. 2003;46:2819–2822. doi: 10.1021/jm030005h. [DOI] [PubMed] [Google Scholar]
- Hewawasam P, Fan W, Cook DA, Newberry KS, Boissard CG, Gribkoff VK, et al. 4-Aryl-3-(mercapto)quinolin-2-ones: novel maxi-K channel opening relaxants of corporal smooth muscle. Bioorg Med Chem Lett. 2004;14:4479–4482. doi: 10.1016/j.bmcl.2004.06.051. [DOI] [PubMed] [Google Scholar]
- Holland M, Langton PD, Standen NB, Boyle JP. Effects of the BKCa channel activator, NS1619, on rat cerebral artery smooth muscle. Br J Pharmacol. 1996;117:119–129. doi: 10.1111/j.1476-5381.1996.tb15163.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmquist F, Andersson KE, Hedlund H. Effects of pinacidil on isolated human corpus cavernosum penis. Acta Physiol Scand. 1990;138:463–469. doi: 10.1111/j.1748-1716.1990.tb08873.x. [DOI] [PubMed] [Google Scholar]
- Jackson G, Rosen RC, Kloner RA, Kostis JB. The second Princeton consensus on sexual dysfunction and cardiac risk: new guidelines for sexual medicine. J Sex Med. 2006;3:28–36. doi: 10.1111/j.1743-6109.2005.00196.x. [DOI] [PubMed] [Google Scholar]
- Kun A, Martinez AC, Tankó LB, Pataricza J, Papp JG, Simonsen U. Ca2+-activated K+ channels in the endothelial cell layer involved in modulation of neurogenic contractions in rat penile arteries. Eur J Pharmacol. 2003;474:103–115. doi: 10.1016/s0014-2999(03)02004-1. [DOI] [PubMed] [Google Scholar]
- Kun A, Kiraly I, Pataricza J, Marton Z, Krassoi I, Varro A, et al. C-type natriuretic peptide hyperpolarizes and relaxes human penile resistance arteries. J Sex Med. 2008;5:1114–1125. doi: 10.1111/j.1743-6109.2008.00775.x. [DOI] [PubMed] [Google Scholar]
- Lee SW, Kang TM. Effects of nitric oxide on the Ca2+-activated potassium channels in smooth muscle cells of the human corpus cavernosum. Urol Res. 2001;29:359–365. doi: 10.1007/s002400100211. [DOI] [PubMed] [Google Scholar]
- Matsumoto K, Yoshida M, Andersson KE, Hedlund P. Effects in vitro and in vivo by apomorphine in the rat corpus cavernosum. Br J Pharmacol. 2005;146:259–267. doi: 10.1038/sj.bjp.0706317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montorsi P, Ravagnani PM, Galli S, Rotatori F, Briganti A, Salonia A, et al. Common grounds for erectile dysfunction and coronary artery disease. Curr Opin Urol. 2004;14:361–365. doi: 10.1097/00042307-200411000-00012. [DOI] [PubMed] [Google Scholar]
- Moon DG, Byun HS, Kim JJ. A KATP-channel opener as a potential treatment modality for erectile dysfunction. BJU Int. 1999;83:837–841. doi: 10.1046/j.1464-410x.1999.00029.x. [DOI] [PubMed] [Google Scholar]
- Moore CR, Wang R. Pathophysiology and treatment of diabetic erectile dysfunction. Asian J Androl. 2006;8:675–684. doi: 10.1111/j.1745-7262.2006.00223.x. [DOI] [PubMed] [Google Scholar]
- Ostergaard L, Stankevicius E, Andersen MR, Eskildsen-Helmond Y, Ledet T, Mulvany MJ, et al. Diminished NO release in chronic hypoxic human endothelial cells. Am J Physiol Heart Circ Physiol. 2007;293:H2894–H2903. doi: 10.1152/ajpheart.01230.2006. [DOI] [PubMed] [Google Scholar]
- Park WS, Kang SH, Son YK, Kim N, Ko JH, Kim HK, et al. The mitochondrial Ca2+ -activated K+ channel activator, NS 1619 inhibits L-type Ca2+ channels in rat ventricular myocytes. Biochem Biophys Res Commun. 2007;362:31–36. doi: 10.1016/j.bbrc.2007.07.057. [DOI] [PubMed] [Google Scholar]
- Prieto D. Physiological regulation of penile arteries and veins. Int J Impot Res. 2008;20:17–29. doi: 10.1038/sj.ijir.3901581. [DOI] [PubMed] [Google Scholar]
- Prieto D, Rivera L, Benedito S, Recio P, Villalba N, Hernández M, et al. Ca2+-activated K+ (KCa) channels are involved in the relaxations elicited by sildenafil in penile resistance arteries. Eur J Pharmacol. 2006a;531:232–237. doi: 10.1016/j.ejphar.2005.12.033. [DOI] [PubMed] [Google Scholar]
- Prieto D, Rivera L, Recio P, Rubio JL, Hernández M, García-Sacristán A. Role of nitric oxide in the relaxation elicited by sildenafil in penile resistance arteries. J Urol. 2006b;175:1164–1170. doi: 10.1016/S0022-5347(05)00320-4. [DOI] [PubMed] [Google Scholar]
- Sadoshima J, Akaike N, Tomoike H, Kanaide H, Nakamura M. Ca-activated K channel in cultured smooth muscle cells of rat aortic media. Am J Physiol. 1988;255:H410–H418. doi: 10.1152/ajpheart.1988.255.3.H410. [DOI] [PubMed] [Google Scholar]
- Sáenz de Tejada I, Angulo J, Cellek S, González-Cadavid N, Heaton J, Pickard R, et al. Pathophysiology of erectile dysfunction. J Sex Med. 2005;2:26–39. doi: 10.1111/j.1743-6109.2005.20103.x. [DOI] [PubMed] [Google Scholar]
- Saleh SN, Angermann JE, Sones WR, Leblanc N, Greenwood IA. Stimulation of Ca2+-gated Cl- currents by the calcium-dependent K+ channel modulators NS1619 [1,3-dihydro-1-[2-hydroxy-5-(trifluoromethyl)phenyl]-5-(trifluoromethyl)-2 H-benzimidazol-2-one] and isopimaric acid. J Pharmacol Exp Ther. 2007;321:1075–1084. doi: 10.1124/jpet.106.118786. [DOI] [PubMed] [Google Scholar]
- Sheng JZ, Braun AP. Small- and intermediate-conductance Ca2+-activated K+ channels directly control agonist-evoked nitric oxide synthesis in human vascular endothelial cells. Am J Physiol Cell Physiol. 2007;293:C458–C467. doi: 10.1152/ajpcell.00036.2007. [DOI] [PubMed] [Google Scholar]
- Shiri R, Koskimäki J, Hakama M, Häkkinen J, Tammela TL, Huhtala H, et al. Effect of chronic diseases on incidence of erectile dysfunction. Urology. 2003;62:1097–1102. doi: 10.1016/j.urology.2003.07.001. [DOI] [PubMed] [Google Scholar]
- Simonsen U, Prieto D, Sáenz de Tejada I, García-Sacristán A. Involvement of nitric oxide in the non-adrenergic non-cholinergic neurotransmission of horse deep penile arteries: role of charybdotoxin-sensitive K+-channels. Br J Pharmacol. 1995;116:2582–2590. doi: 10.1111/j.1476-5381.1995.tb17211.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simonsen U, García-Sacristán A, Prieto D. Penile arteries and erection. J Vasc Res. 2002;39:283–303. doi: 10.1159/000065541. [DOI] [PubMed] [Google Scholar]
- Sobey CG. Potassium channel function in vascular disease. Arterioscler Thromb Vasc Biol. 2001;21:28–38. doi: 10.1161/01.atv.21.1.28. [DOI] [PubMed] [Google Scholar]
- Spektor M, Rodriguez R, Rosenbaum RS, Wang HZ, Melman A, Christ GJ. Potassium channels and human corporeal smooth muscle cell tone: further evidence of the physiological relevance of the Maxi-K channel subtype to the regulation of human corporeal smooth muscle tone in vitro. J Urol. 2002;167:2628–2635. [PubMed] [Google Scholar]
- Taniguchi J, Furukawa KI, Shigekawa M. Maxi K+ channels are stimulated by cyclic guanosine monophosphate-dependent protein kinase in canine coronary artery smooth muscle cells. Pflugers Arch. 1993;423:167–172. doi: 10.1007/BF00374390. [DOI] [PubMed] [Google Scholar]
- Trigo-Rocha F, Donatucci CF, Hsu GL, Nunes L, Lue TF, Tanagho EA. The effect of intracavernous injection of potassium channel openers in monkeys and dogs. Int J Impot Res. 1995;7:41–48. [PubMed] [Google Scholar]
- Venkateswarlu K, Giraldi A, Zhao W, Wang HZ, Melman A, Spektor M, et al. Potassium channels and human corporeal smooth muscle cell tone: diabetes and relaxation of human corpus cavernosum smooth muscle by adenosine triphosphate sensitive potassium channel openers. J Urol. 2002;168:355–361. [PubMed] [Google Scholar]
- Villalba N, Stankevicius E, Garcia-Sacristan A, Simonsen U, Prieto D. Contribution of both Ca2+ entry and Ca2+ sensitization to the alpha1-adrenergic vasoconstriction of rat penile small arteries. Am J Physiol Heart Circ Physiol. 2007;292:H1157–H1169. doi: 10.1152/ajpheart.01034.2006. [DOI] [PubMed] [Google Scholar]
- Vrentzos GE, Paraskevas KI, Mikhailidis DP. Dyslipidemia as a risk factor for erectile dysfunction. Curr Med Chem. 2007;14:1765–1770. doi: 10.2174/092986707781058931. [DOI] [PubMed] [Google Scholar]
- Werner ME, Zvara P, Meredith AL, Aldrich RW, Nelson MT. Erectile dysfunction in mice lacking the large-conductance calcium-activated potassium (BK) channel. J Physiol. 2005;567:545–556. doi: 10.1113/jphysiol.2005.093823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wingard C, Fulton D, Husain S. Altered penile vascular reactivity and erection in the Zucker obese-diabetic rat. J Sex Med. 2007;4:348–362. doi: 10.1111/j.1743-6109.2007.00439.x. [DOI] [PubMed] [Google Scholar]