

Abstract
In vascular endothelium, adherens junctions between endothelial cells are composed of VE-cadherin, an adhesive receptor crucial for the proper assembly of vascular structures and the maintenance of vascular integrity. As classical cadherins, VE-cadherin links endothelial cells together by homophilic interactions mediated by its extracellular part and associates intracellularly with actin cytoskeleton via catenins. Although, from structural crystallographic data, a dimeric structure arranged in a trans orientation emerge as a potential mechanism of cell-cell adhesion, the cadherin organization within adherens junction remains under controversy. Concerning VE cadherin, its extracellular part possesses the capacity to self-associate in solution as hexamers consisting of three antiparallel cadherin dimers. VE-cadherin based adherens junctions were reconstituted in vitro by assembly of VE-EC1-4 hexamer at the surfaces of liposomes. The artificial adherens junctions revealed by Cryo-Electron Microscopy appear as two dimensional self-assembly of hexameric structures. This cadherin organization is reminiscent of that found in native desmosomal junctions. Further structural studies performed on native VE-cadherin junctions would provide a better understanding of the cadherin organization within adherens junction.
Homophilic interactions between cadherins are intracellularly strengthened by connection to the actin cytoskeleton. Recently, we have discovered that annexin 2, an actin binding protein connects the VE-cadherin-catenin complex to the actin cytoskeleton. This novel link is labile and promotes the endothelial cell switch between a quiescent to an angiogenic state.
Keywords: Actins; metabolism; Adherens Junctions; metabolism; ultrastructure; Animals; Cadherins; chemistry; physiology; Cell Adhesion; Cryoelectron Microscopy; methods; Endothelium, Vascular; metabolism; ultrastructure; Humans; Membranes, Artificial; Models, Molecular
Keywords: Cryoelectron microscopy, VE cadherin, 3D reconstruction, endothelial junction, annexin 2
Introduction
Blood vessels are lined with a monolayer of cells that form a barrier between blood and underlying tissues. This monolayer, designated as vascular endothelium (VE), plays a central role in regulating the recruitment of leukocytes at sites of injury or inflammation, and in the growth and repair of blood vessels. All these processes are tightly controlled by adherens junctions formed between endothelial cells. In VE, adherens junctions are mainly composed of VE–cadherin, an adhesive receptor able to self-associate at endothelial cell-cell contacts as illustrated in Figure 1 [1].
Figure 1.
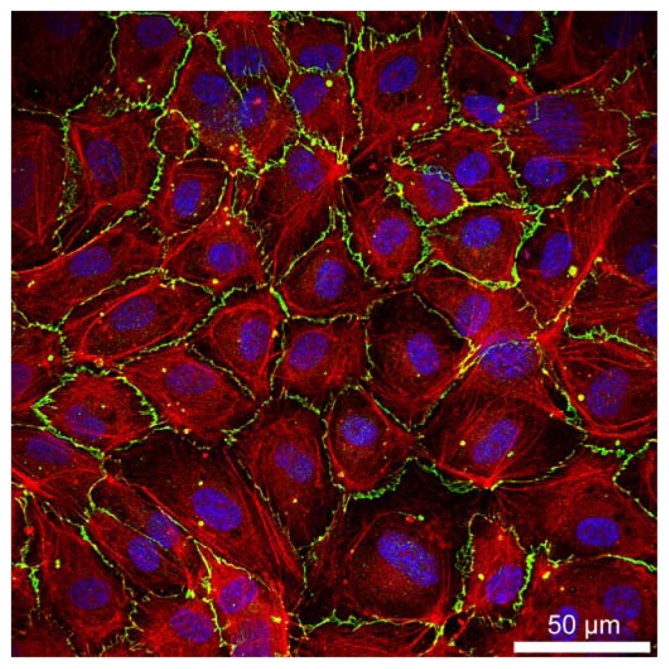
Confocal analysis of VE-cadherin and actin in confluent endothelial cell monolayer (HUVEC) Cells were plated on glass coverslips, cultured for three days, and then stained with a polyclonal anti-VE cadherin, Cy5-labelled goat-anti-rabbit (green), FITC-labelled phalloidin (red), and DAPI (blue)
VE cadherin is a member of a superfamily of Ca++- dependent adhesive receptors known to bind cells together via homotypic interactions. By promoting clustering of cells with identical phenotypes, cadherins play a crucial role in the elaboration of various tissues during embryogenesis [2] and in the maintenance of adult tissue architecture. VE cadherin is crucial for the proper - assembly of vascular structures and maintenance of vascular integrity [3]. Inactivation of VE–cadherin gene or truncation of the cytoplasmic domain of VE cadherin were demonstrated to be embryonically lethal due to vascular insufficiency [4,5]. In adult mice, antibodies directed against VE cadherin extracellular region lead to an increase of microvessel permeability or impair the assembly of new vascular structures caused by disruption of VE–cadherin–mediated cell-cell adhesion or by inhibition of adherens junction formation [6,7].
Basic molecular structure of cadherins
Classical cadherins possess a general modular structure consisting of five ExtraCellular repeats of about 110 amino acids, designated as EC1 to EC5 from the N- to the C-terminus, a transmembrane domain and a well conserved cytoplasmic tail [8,9]. Whereas the extracellular part of cadherin mediates homophilic interactions, their cytoplasmic regions bind intracellular partners including catenins (α,β,γ) and p120, which in turn bind to the actin cytoskeleton [10].
Among the cadherin superfamily, classical cadherins comprise two major subfamilies termed type I and type II cadherins. Both these types have a conserved tryptophan (Trp2) at the N-terminus while the type II cadherins, such as VE cadherin, have an additional conserved tryptophan (Trp4). Despite this difference, type I and type II EC domains contain seven stranded barrels (Greek-key sandwiches) revealing an unexpected structural similarity with the immunoglobulin fold [11–14]. The 3D structure of the entire extracellular domain of C cadherin (from Xenopus) has revealed an elongated structure containing calcium binding sites located between two consecutive EC domains [13]. Fixation of three Ca++ per site rigidifies the cadherin molecules thus reducing their sensitivity to proteolytic degradation.
Homophilic interactions mediated by the extracellular domain of cadherins
A variety of experiments demonstrate that the adhesive binding site of cadherin is primarily localized within the EC1 domain. From crystallographic studies, pairs of molecules are formed through the exchange of a N terminal β strand between the partner EC1 domains, named the swapping strand [11,13,14]. For type I cadherins, the EC1-EC1 interaction results from the insertion of the conserved Trp2 side chain from the swapping strand of the partner1 into the hydrophobic core of the partner2 molecule and vice versa. Within the E –cadherin pocket, the formation of salt bridge between Asp1 and Glu89 and hydrogen bond between Trp2 and Asp90 stabilizes the swapping interaction. The Pro5-Pro6 motif could contribute to the Trp2 exchange [15]. For type II cadherins, the side chains of the critical conserved residues Trp2 and Trp4 of the swapped strand insert into a large hydrophobic core of the partner molecule (Patel, 2006). This swapping interaction involving EC1 domain leads to the formation of antiparallel dimers that could mediate binding between cadherins exposed from opposing cells [13]. In addition to these structural data, EC1 domain emerges as the determinant area involved in the specific recognition of classical cadherins [14,16].
VE cadherin hexamer and artificial VE-cadherin based junctions
Junctional assembly is a complex process which requires various events such as the local concentration of cadherin molecules at the cell surface, the recruitment of both cytoskeleton binding proteins and regulating proteins and the reorganization of cytoskeleton network. All these processes are intimately interrelated and their individual contribution to the modulation of cell-cell junction strength remains difficult to define. Consequently, to explore the role of the VE cadherin ectodomain clustering independently of the cellular events mediated by the cytoplasmic VE cadherin-catenin complex, we have expressed in E. coli a fragment overlapping the four N-terminal extracellular domains of VE cadherin (EC1-4).
We have demonstrated that this fragment possesses the capacity to self-associate as hexamers in solution. Cryo-electron microscopy allowed us to visualize the EC1-4 hexamer as a hollow cylindrical structure of 23 nm long and 7 nm wide [17,18]. This finding suggests that the hexameric units observed in solution may participate to the elaboration of endothelial adherens junctions. Therefore we performed cryo-electron microscopy studies to provide novel insights in the way endothelial cells adhere to each other at adherens junctions [19]. Thus, a soluble VE-cadherin molecule made of the four N-terminal EC domains (EC1-4-His) was produced with a polyhistidine extension at its C-terminal end in order to allow its binding to membranes containing lipids functionalized with Ni-NTA groups and to mimic the natural orientation of cadherin molecules at cell membranes. It was shown that artificial adherens junctions can be reconstituted in vitro, by direct 2D self-assembly of the EC1-4-His fragment sandwiched between two membranes (Fig. 2A–D). The molecular architecture of these artificial junctions exhibiting a 23 nm intermembrane distance consists in the juxtaposition of the hexameric units found in solution. It appears to be similar to native desmosomal cadherin-based junctions observed in vivo [20]. This study demonstrated that the minimal information required to build up an artificial adherens junction is contained within the extracellular moiety of cadherin molecules.
Figure 2.
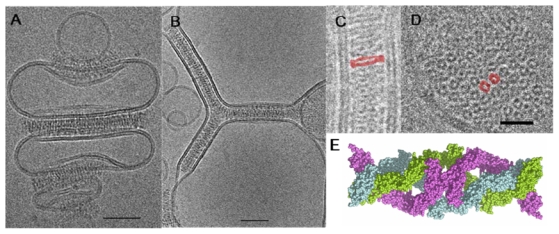
Architecture of artificial adherens junctions. (A–B) Self assembly of VE EC1-4 fragments on liposomes membranes observed by cryo-EM (scale bar 50 nm). (C, D) Detailed side and top views of junctions (scale bar 20 nm). (E) A model of VE EC1-4 hexamer [22].
Furthermore this finding underlines the propensity of the extracellular moiety of cadherin to form spontaneous clusters that may influence the adhesion. It was shown previously that the lateral clustering of cadherin mutants lacking the cytoplasmic tail increased the cadherin-mediated adhesive strength [21]. Although clustering mediated by the cadherin cytoplasmic part should contribute to a strong cadherin-based adhesion, self-assembly of the cadherin extracellular part can provide alternative modulations in the adhesive strength.
To determine the molecular architecture of the molecule, we calculated a 3D reconstruction from negatively stained electron microscopy images and produced a homology model based on the known structure of C-cadherin EC1-5 (Fig. 2E) [21]. The hexamer model consists of a trimer of dimers with each N-terminal EC1 module making an anti-parallel dimeric contact, and the EC4 modules forming extensive trimeric interactions. Each EC1-4 molecule makes a helical curve that gives some torsional flexibility to the edifice [19,22]. While there is no direct evidence for the existence of VE-cadherin hexamers at adherens junctions, this model is particularly of interest since it is supported by some of disparate results for adherens junctions. In particular, it is in agreement with the X-ray and electron microscopy results which demonstrate that the EC1 dimer is central to homotypic cadherin interaction. It provides an explanation for the force measurements of the interaction between opposing cadherin layers, which have previously been interpreted as resulting from three different interdigitating interactions [23]. It is in accord with observations of native junctions. The fact that this hexameric model of VE-cadherin can be used to explain more of the existing data on adherens junctions than any other model alone argues in favour of the existence of the hexamer at the adherens junction. In the context of the cell-cell junction, a mechanism involving cis-trimers instead of single monomer for bridging opposite cell membranes would increase the avidity and the strength of adhesive binding.
Organization of native cadherin junction
Adherens junctions and desmosomes that densely associated with actin and intermediate filaments respectively are ones of the most abundant types of intercellular adhesive junctions. They consist of classical and desmosomal cadherins. Due to the sequence and organization similarity between the extracellular domains of classical and desmosomal cadherins, both junctions adopt similar architecture. As attested by electron microscopy analyses, cadherin molecules within these adhesive junctions form a lattice that maintains the intermembrane space to a constant thickness of about 30 nm for desmosomes [20,24,25].
While there is a consensual view about the involvement of EC1 in the primary formation of trans dimer, the organization of the cadherin extracellular part within the intermembrane space remains a matter of debate. In the 3D structure of C-cadherin [13], the trans dimer at the basis of cell-cell adhesion exhibits a 38.5 nm length too long to fit within the measured intermembrane space. To take into account this geometrical constraint, Boggon and co-workers proposed that the adhesive trans dimer adopts a curved structure over the length of the extracellular domain of C-cadherin. This results in a end-to-end binding distance of 24.5 nm consistent with EM images of adherens junctions. An other model issued from electron tomography of mouse skin plastic sections proposed that adherens junction are composed of discrete groups of 10–20 cadherin molecules interacting via their tips, leading to the formation of trans dimers [25].
However, the recent observations of desmosome junctions performed on 50 nm thick human epidermal sections preserved in hydrated state using CEMOVIS (Cryo-Electron Microscopy Of VItreous Sections) technique did not support those models of cadherin assembly [20]. Desmosome junctions exhibited within the intercellular space, a regular organization of the cadherin molecules made of elongated densities perpendicular to the membrane. This indicates that cadherins are densely packed in the desmosome junction and not sparsely arranged. Our results are in agreement with the latter model of cadherin clustering in which cadherin molecules are well ordered and arranged perpendicularly to the cell membrane.
VE-cadherin Native Junction between HUVEC cells
In endothelial cell monolayers, treatment with the actin monomer sequestering drug latrunculin B induces a depolymerisation of actin cytoskeleton accompanied by a drastic destabilization of cell-cell junctions. The removal of the drug promotes the reformation of the junctions. Altogether, these results indicate that the homophilic interactions mediated by the extracellular part of cadherins must be strengthened by intracellular interactions involving actin cytoskeleton to maintain cell-cell junctions. Intracellularly, cadherins associates with β- or γ-catenin which in turn interacts with α-catenin. Until recently, it was commonly admitted that α-catenin binding to β- or γ-catenins promotes connections between cadherin-based complex and actin cytoskeleton. But this concept was challenged since it was recently demonstrated that α-catenin cannot simultaneously bind to β-catenin and the actin cytoskeleton [26]. Recently, using proteomic tools, we have discovered new partners within the VE-cadherin- based junctional complex. Among them, the membrane binding protein annexin 2 is the most abundant. We have observed that the ablation of the expression of annexin 2 in HUVEC by siRNA induces a drastic modification of the localization of VE-cad at cell-cell junctions. This protein, located in cholesterol rafts, binds both to the actin cytoskeleton and the VE-cadherin-based complex so the complex is docked to cholesterol rafts. These multiple connections prevent the lateral diffusion of the VE-cadherin-based complex thus strengthening adherens junctions in the ultimate steps of maturation. In fact, the connection of annexin 2 to the junctional VE-cadherin-based complex is very labile and governs, at least in part, the switch between immature and mature states of endothelial cell-cell junctions [27].
Conclusion
Our results support an alternative model of cadherin clustering. In this model, the extracellular part of VE-cadherin is able to elaborate hexameric self-assemblies composed of three trans-dimers, each of them exhibiting a supertwisted structure. Once attached to membrane, these hexamers pack in close arrays to build large 2D edifices reminiscent of the organization of desmosomal cadherins in native tissues. Furthermore as evidenced by the involvement of the membrane binding protein annexin 2, a more extended inventory of proteins associated to VE cadherin based junctions will allow a better a comprehensive understanding of the cadherin-mediated cell-cell interaction processes.
References
- 1.Lampugnani MG, Resnati M, Raiteri M, Pigott R, Pisacane A, Houen G, Ruco LP, Dejana E. A novel endothelial-specific membrane protein is a marker of cell-cell contacts. J Cell Biol. 1992;118:1511–1522. doi: 10.1083/jcb.118.6.1511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dawes-Hoang RE, Parmar KM, Christiansen AE, Phelps CB, Brand AH, Wieschaus EF. Folded gastrulation, cell shape change and the control of myosin localization. Development. 2005;132:4165–4178. doi: 10.1242/dev.01938. [DOI] [PubMed] [Google Scholar]
- 3.Gulino D, Delachanal E, Concord E, Genoux Y, Morand B, Valiron MO, Sulpice E, Scaife R, Alemany M, Vernet T. Alteration of endothelial cell monolayer integrity triggers resynthesis of vascular endothelium cadherin. J Biol Chem. 1998;273:29786–29793. doi: 10.1074/jbc.273.45.29786. [DOI] [PubMed] [Google Scholar]
- 4.Gory-Faure S, Prandini MH, Pointu H, Roullot V, Pignot-Paintrand I, Vernet M, Huber P. Role of vascular endothelial-cadherin in vascular morphogenesis. Development. 1999;126:2093–2102. doi: 10.1242/dev.126.10.2093. [DOI] [PubMed] [Google Scholar]
- 5.Carmeliet P, Lampugnani MG, Moons L, Breviario F, Compernolle V, Bono F, Balconi G, Spagnuolo R, Oostuyse B, Dewerchin M, Zanetti A, Angellilo A, Mattot V, Nuyens D, Lutgens E, Clotman F, de Ruiter MC, Gittenberger-de Groot A, Poelmann R, Lupu F, Herbert JM, Collen D, Dejana E. Targeted deficiency or cytosolic truncation of the VE-cadherin gene in mice impairs VEGF-mediated endothelial survival and angiogenesis. Cell. 1999;98:147–57. doi: 10.1016/s0092-8674(00)81010-7. [DOI] [PubMed] [Google Scholar]
- 6.Corada M, Zanetta L, Orsenigo F, Breviario F, Lampugnani MG, Bernasconi S, Liao F, Hicklin DJ, Bohlen P, Dejana E. A monoclonal antibody to vascular endothelial-cadherin inhibits tumor angiogenesis without side effects on endothelial permeability. Blood. 2002;100:905–911. doi: 10.1182/blood.v100.3.905. [DOI] [PubMed] [Google Scholar]
- 7.May C, Doody JF, Abdullah R, Balderes P, Xu X, Chen CP, Zhu Z, Shapiro L, Kussie P, Hicklin DJ, Liao F, Bohlen P. Identification of a transiently exposed VE-cadherin epitope that allows for specific targeting of an antibody to the tumor neovasculature. Blood. 2005;105:4337–4344. doi: 10.1182/blood-2005-01-0010. [DOI] [PubMed] [Google Scholar]
- 8.Hatta K, Nose A, Nagafuchi A, Takeichi M. Cloning and expression of cDNA encoding a neural calcium-dependent cell adhesion molecule: its identity in the cadherin gene family. J Cell Biol. 1988;106:873–881. doi: 10.1083/jcb.106.3.873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Takeichi M. Cadherins: a molecular family important in selective cell-cell adhesion. Annu Rev Biochem. 1990;59:237–252. doi: 10.1146/annurev.bi.59.070190.001321. [DOI] [PubMed] [Google Scholar]
- 10.Gumbiner BM. Cell adhesion: the molecular basis of tissue architecture and morphogenesis. Cell. 1996;84:345–57. doi: 10.1016/s0092-8674(00)81279-9. [DOI] [PubMed] [Google Scholar]
- 11.Shapiro L, Fannon AM, Kwong PD, Thompson A, Lehmann MS, Grubel G, Legrand JF, Als-Nielsen J, Colman DR, Hendrickson WA. Structural basis of cell-cell adhesion by cadherins. Nature. 1995;374:327–337. doi: 10.1038/374327a0. [DOI] [PubMed] [Google Scholar]
- 12.Overduin M, Harvey TS, Bagby S, Tong KI, Yau P, Takeichi M, Ikura M. Solution structure of the epithelial cadherin domain responsible for selective cell adhesion. Science. 1995;267:386–389. doi: 10.1126/science.7824937. [DOI] [PubMed] [Google Scholar]
- 13.Boggon TJ, Murray J, Chappuis-Flament S, Wong E, Gumbiner BM, Shapiro L. C-cadherin ectodomain structure and implications for cell adhesion mechanisms. Science. 2002;296:1308–1313. doi: 10.1126/science.1071559. [DOI] [PubMed] [Google Scholar]
- 14.Patel SD, Ciatto C, Chen CP, Bahna F, Rajebhosale M, Arkus N, Schieren I, Jessell TM, Honig B, Price SR, Shapiro L. Type II cadherin ectodomain structures: implications for classical cadherin specificity. Cell. 2006;124:1255–1268. doi: 10.1016/j.cell.2005.12.046. [DOI] [PubMed] [Google Scholar]
- 15.Parisini E, Higgins JM, Liu JH, Brenner MB, Wang JH. The Crystal Structure of Human E-cadherin Domains 1 and 2, and Comparison with other Cadherins in the Context of Adhesion Mechanism. J Mol Biol. 2007;373:401–411. doi: 10.1016/j.jmb.2007.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shapiro L, Love J, Colman DR. Adhesion molecules in the nervous system: structural insights into function and diversity. Annu Rev Neurosci. 2007;30:451–474. doi: 10.1146/annurev.neuro.29.051605.113034. [DOI] [PubMed] [Google Scholar]
- 17.Legrand P, Bibert S, Jaquinod M, Ebel C, Hewat E, Vincent F, Vanbelle C, Concord E, Vernet T, Gulino D. Self-assembly of the vascular endothelial cadherin ectodomain in a Ca2+-dependent hexameric structure. J Biol Chem. 2001;276:3581–3588. doi: 10.1074/jbc.M002667200. [DOI] [PubMed] [Google Scholar]
- 18.Bibert S, Jaquinod M, Concord E, Ebel C, Hewat E, Vanbelle C, Legrand P, Weidenhaupt M, Vernet T, Gulino-Debrac D. Synergy between extracellular modules of vascular endothelial cadherin promotes homotypic hexameric interactions. J Biol Chem. 2002;277:12790–12801. doi: 10.1074/jbc.M111597200. [DOI] [PubMed] [Google Scholar]
- 19.Lambert O, Taveau JC, Lai Kee Him J, Al Kurdi R, Gulino-Debrac D, Brisson A. The basic framework of VE-cadherin junctions revealed by cryo-EM. J Mol Biol. 2005;346:1193–1196. doi: 10.1016/j.jmb.2004.12.053. [DOI] [PubMed] [Google Scholar]
- 20.Al-Amoudi A, Norlen LP, Dubochet J. Cryo-electron microscopy of vitreous sections of native biological cells and tissues. J Struct Biol. 2004;148:131–135. doi: 10.1016/j.jsb.2004.03.010. [DOI] [PubMed] [Google Scholar]
- 21.Yap AS, Brieher WM, Pruschy M, Gumbiner BM. Lateral clustering of the adhesive ectodomain: a fundamental determinant of cadherin function. Curr Biol. 1997;7:308–15. doi: 10.1016/s0960-9822(06)00154-0. [DOI] [PubMed] [Google Scholar]
- 22.Hewat EA, Durmort C, Jacquamet L, Concord E, Gulino-Debrac D. Architecture of the VE-cadherin hexamer. J Mol Biol. 2007;365:744–751. doi: 10.1016/j.jmb.2006.10.052. [DOI] [PubMed] [Google Scholar]
- 23.Sivasankar S, Brieher W, Lavrik N, Gumbiner B, Leckband D. Direct molecular force measurements of multiple adhesive interactions between cadherin ectodomains. Proc Natl Acad Sci U S A. 1999;96:11820–11824. doi: 10.1073/pnas.96.21.11820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Drochmans P, Freudenstein C, Wanson JC, Laurent L, Keenan TW, Stadler J, Leloup R, Franke WW. Structure and biochemical composition of desmosomes and tonofilaments isolated from calf muzzle epidermis. J Cell Biol. 1978;79:427–443. doi: 10.1083/jcb.79.2.427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.He W, Cowin P, Stokes DL. Untangling desmosomal knots with electron tomography. Science. 2003;302:109–113. doi: 10.1126/science.1086957. [DOI] [PubMed] [Google Scholar]
- 26.Yamada S, Pokutta S, Drees F, Weis WI, Nelson WJ. Deconstructing the cadherin-catenin-actin complex. Cell. 2005;123:889–901. doi: 10.1016/j.cell.2005.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Heyraud S, Jaquinod M, Durmort C, Concord E, Garin J, Gulino-Debrac D. Proteomic analysis of VE cadherin-based adherens junctions. Mol Cell Biol. in press. [Google Scholar]