

Abstract
Mineral surfaces were important during the emergence of life on Earth because the assembly of the necessary complex biomolecules by random collisions in dilute aqueous solutions is implausible. Most silicate mineral surfaces are hydrophilic and organophobic and unsuitable for catalytic reactions, but some silica-rich surfaces of partly dealuminated feldspars and zeolites are organophilic and potentially catalytic. Weathered alkali feldspar crystals from granitic rocks at Shap, north west England, contain abundant tubular etch pits, typically 0.4–0.6 μm wide, forming an orthogonal honeycomb network in a surface zone 50 μm thick, with 2–3 × 106 intersections per mm2 of crystal surface. Surviving metamorphic rocks demonstrate that granites and acidic surface water were present on the Earth’s surface by ∼3.8 Ga. By analogy with Shap granite, honeycombed feldspar has considerable potential as a natural catalytic surface for the start of biochemical evolution. Biomolecules should have become available by catalysis of amino acids, etc. The honeycomb would have provided access to various mineral inclusions in the feldspar, particularly apatite and oxides, which contain phosphorus and transition metals necessary for energetic life. The organized environment would have protected complex molecules from dispersion into dilute solutions, from hydrolysis, and from UV radiation. Sub-micrometer tubes in the honeycomb might have acted as rudimentary cell walls for proto-organisms, which ultimately evolved a lipid lid giving further shelter from the hostile outside environment. A lid would finally have become a complete cell wall permitting detachment and flotation in primordial “soup.” Etch features on weathered alkali feldspar from Shap match the shape of overlying soil bacteria.
Keywords: biological evolution, mineral surfaces
Mineral surfaces must have been important during the emergence of life on Earth, based on observations of minerals in modern rocks and soils. Whereas the assembly of the essential complex bio-organic molecules by random collisions in dilute aqueous soups is implausible, mineral surfaces could have played a catalytic role and acted as templates for the assembly of the first replicating organic molecules (1, 2). The most suitable sites might have been the silica-rich surfaces of partly dealuminated feldspars and zeolites. Many studies of industrial processes involving synthetic zeolites have demonstrated preferential adsorption of organic species over water and presence of aluminum-containing catalytic centers (3). Replication might have followed an organic “takeover” of the reproduction of crystals of fine-grained mineral particles (4). The evolution of a lipid cell wall might have involved organic replacement of iron sulfide membranes (5). Minerals have therefore been invoked in both catalytic and self-organizational roles. We have described periodic, self-organized tubular microstructures, in exactly the size range of modern soil bacteria, on the surfaces of weathered alkali feldspar crystals (6, 7). The microstructures also occur on feldspars in sediments undergoing diagenesis (8). They should have been commonplace in the early Earth, combining both catalytic and organizational features on the same surface. We describe their mode of origin and aspects of their surface chemistry, discuss their potential as abundant, linked catalytic micro-reactors for biosynthesis, and examine the possibility that they acted as cell walls for proto-organisms during the earliest stages of biological evolution.§
Problems recur with hypotheses for the assembly of the earliest molecules with the properties commonly associated with “life.” These include the unlikelihood that complex self-replicating molecules such as RNA could form by chance encounters even over geological time; the difficulty of protecting such molecules, once formed, from dilution and destruction by high temperatures, hydrolysis and ultra-violet radiation; and finally the difficulty of imagining how self-organization alone could lead to the encapsulation of a complex hierarchy of biochemical reactions in a membrane to form the simplest unicellular organism. Mineral-catalyzed reactions, followed by a series of fractionations, offer the most plausible route to RNA (9), but how could an extended concatenation of biochemical reactions develop without a container of suitable size and mechanical and chemical stability? Such a container, if it did not itself reproduce, need not be totally isolated, but would have to be able to transfer reactants to adjacent containers so that the essential replicating polymer could spread. We describe here abundant microstructures on the surface of alkali feldspar, a common aluminosilicate mineral, which have several features suitable for the catalysis and encapsulation of prebiotic reactants and products.
Potassium-rich alkali feldspar (K-feldspar) makes up 20–40% by vol of granites and is a major constituent of the Earth’s crust. Metamorphism has obscured the early geological record, but radioactive dating coupled with field geology and petrography is yielding valuable information on the earliest rocks. The oldest known rocks, the most extensive example of which is the Itsaq Gneiss Complex, West Greenland, in which the oldest units formed ∼3.8 Ga ago (10), are predominantly (70–80% by vol) tonalitic gneisses with a few percent of K-feldspar. A further 10% of the Itsaq rocks are granites and granitic gneisses with up to 40% K-feldspar. Water-lain volcaniclastic rocks occur in the sequence at ∼3.7 Ga and perhaps as early as ∼3.8 Ga, and sedimentary banded iron formations occur as early as 3.8 Ga (11). Thus K-feldspar must have been a common component of the Earth’s surface by ∼3.8 Ga by which time it was also undergoing weathering and particulate transport in water. In the absence of organic soils, large areas of K-feldspar would have been exposed to the atmosphere or shallow water. The oldest known fossils are microbial mats formed at ∼3.5 Ga and filamentous microbial fossils dated at ∼3.4 Ga (12). Carbon inclusions in apatite crystals from a 3.8 Ga Itsaq iron formation are unusually depleted in 13C, consistent with biological activity (11), although there are alternative explanations (13). We conclude, therefore, that a granite-rich crust was present at least by the end of the heavy bombardment recorded on the Moon at ∼3.8 Ga (14) and that the earliest life appeared shortly thereafter.
Weathered surfaces of K-feldspars from granitic source rocks and from feldspathic gneisses are complex (Fig. 1) and covered in regularly distributed etch-pits and grooves. The etch-pits generate extraordinary, cross-linked tubular networks (Fig. 2), which extend to ≥50 μm below the surface. The macroscopic effect of these pits is familiar; freshly cleaved surfaces of feldspars are strongly reflective whereas weathered surfaces are dull and chalky. The etch-pits form by dissolution of edge dislocations that develop during cooling of the igneous or metamorphic protolith (15). They are characteristic features of all alkali feldspars of the composition range found in sub-solvus granites and gneisses. In our example from Shap (Figs. 1 and 2), primary feldspar crystals grew between ∼970 and 1070 K (16). The crystals were not pure potassium feldspar but a solid solution with composition close to K0.7Na0.3AlSi3O8. Such solutions become unstable during cooling, and at ∼940 K, the crystals begin to unmix into lens-shaped lamellae of nearly end-member NaAlSi3O8 (albite) in somewhat less pure KAlSi3O8 (orthoclase). The intergrowths share a continuous (coherent) AlSi3O8 framework. Because the cell dimensions of albite and orthoclase are different, elastic coherency strains develop at the lamellar interfaces, which orientate themselves in a noncrystallographic plane close to 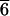 01 to minimize coherency strain energy (17). As the temperature falls, Na+ and K+ continue to diffuse through the structure, and the lamellar intergrowths (cryptoperthites) coarsen by thickening and extending in length to produce flatter lenses. The lamellae are locally regular both in thickness and separation but vary over the crystal as a whole because of differences in the local bulk composition inherited from crystal growth. The thickest lamellae are typically 1–2 μm apart and up to 400 nm thick.
01 to minimize coherency strain energy (17). As the temperature falls, Na+ and K+ continue to diffuse through the structure, and the lamellar intergrowths (cryptoperthites) coarsen by thickening and extending in length to produce flatter lenses. The lamellae are locally regular both in thickness and separation but vary over the crystal as a whole because of differences in the local bulk composition inherited from crystal growth. The thickest lamellae are typically 1–2 μm apart and up to 400 nm thick.
Figure 1.

SEM images of weathered (001) surfaces of alkali feldspar fragments originating from phenocrysts in the porphyritic granite at Shap, Cumbria, north west England. (Scale bar 5 μm.) (a) Fragment from gravel, showing the beginning of formation of etch pits by dissolution at the outcrops of edge dislocations. Lamellae of albite in orthoclase run horizontally across the image. Although appearing parallel-sided at this scale, the lamellae are in reality very flat lenses around which the dislocations form continuous loops, causing each single dislocation to form a pair of etch pits. Many of the pairs have merged to produce single, hourglass-shaped pores. (b) SEM image of a highly weathered Shap feldspar from a peat soil, inhabited by at least three varieties of rod and disc-shaped bacteria. Note the grooves where etch pits have coalesced and the paired etch pits where deeper parts of the surface have been exposed by flaking. The bacteria will have been reduced in size by the high vacuum in the microscope but nevertheless are closely similar in size to the surface features.
Figure 2.

SEM image of a resin cast of an etch-pit network near the surface of a weathered Shap alkali feldspar. (Scale bar 20 μm.) The cast was made by impregnating the feldspar with Araldite resin under vacuum, curing, and dissolving away the feldspar in concentrated HF. The surface of the feldspar is off the bottom of the micrograph, and the image is of a pile of two-dimensional networks that have fallen over to lie on top of each other. Because the resin is flexible, parts of the networks are curved. The original etch-pits were developed on edge dislocations very nearly parallel to b (horizontal) and c (vertical) in the perthite contact plane close to 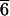 01 of the monoclinic feldspar.
01 of the monoclinic feldspar.
At 680–640 K, as the structure stiffens, the coherency stresses on the longer, thicker lamellae become unsustainable and regularly spaced edge dislocations nucleate. The dislocations have the form of very extended lens-shaped loops encircling the exsolution lamellae so that they appear in pairs when intersected by cleavage surfaces (Fig. 1). The separation between individual dislocation loops depends on the local thickness of the exsolution lamellae, in response to the magnitude of the local coherency stresses (15). The dislocations form in two orientations at right angles producing paired two-dimensional nets (Fig. 2). Thus, when the crystal reaches surface temperatures, it contains a self-organized network of edge dislocations with their associated core and peripheral strain energies.
When dissolution begins in a weathering, or diagenetic (8), environment, it proceeds more rapidly at dislocation outcrops than at a normal surface (6). The differential is most marked when surface and solution are close to equilibrium, as in the confined spaces in soils or sedimentary rocks. Under these circumstances the contribution of the dislocations to the free energy change on dissolution is proportionately greater than when dissolution is occurring far from equilibrium. The overall dissolution rate of feldspars is mainly a function of pH and temperature (18), but how these factors specifically affect dissolution at dislocations has not been investigated. The surface illustrated (Figs. 1 and 2) has been exposed to weathering for some thousands of years, since the end of the last glaciation at Shap. Thus under postglacial conditions with temperature rarely above 283 K and soil water pH ∼3.4 (7), the etch pits developed at a minimum average rate of 5 nm y−1. Dissolution might have been more rapid on the early Earth, depending on rainfall, pH, and temperature. However, inward advance of the etch tubes, which taper toward their base (7), is ultimately limited by diffusion of solutes in the very confined “internal zone” of the crystal (19). Organic solvents are not required to produce the etch-tubes, which are easily imaged by using scanning electron microscopy (SEM) after 50 s exposure to HF vapor (20), and which are detectable, using atomic force microscopy, after ∼140 days in pH 2 HCl at 298 K (7). Whatever the exact temperature and solvent chemistry in the early Archaean, it seems likely that honeycombed surfaces would develop rapidly on alkali feldspars and be as commonplace as they are now.
The potential of the etch-pit networks to act as catalytic surfaces is obvious. The (001) and (010) cleavage surfaces contain intersections with ∼2–3 ×106 pits per mm2. Individual tubes orientated normal to the grain surface are typically 0.4–0.6 μm wide, whereas those orientated parallel to the surface are narrower, typically 0.2 μm wide. A geometrical calculation assuming a 50-μm thick layer shows that a nominal 1 mm2 of weathered alkali feldspar has an actual surface area of ∼130 mm2. Although many other types of grain-scale to atomic-scale defects occur in and between minerals in coarse-grained rocks, we are not aware of any other feature which approaches alkali feldspar honeycombs in its contribution to surface area, or has similar regularity.
Crucial to the hypothesis that feldspar can provide a substrate for the catalytic assembly of polymers is the nature of the surface at the atomic level. In common with most silicates, most unmodified feldspar surfaces are hydrophilic and organophobic. However some silica-rich zeolites such as synthetic silicalite/ZSM-5 (mutinaite is the Al-bearing mineral equivalent) have electrically neutral Si-O surfaces, which strongly adsorb organic species over water (3). It is well established experimentally that acid leaching of feldspars leads to silica-rich surfaces (21, 22), although such surfaces have not yet been shown to exist on naturally weathered feldspars in the absence of biochemical weathering. However, a silica-rich layer has been detected at interfaces between lichen and feldspars on weathered quarry-faces of Shap granite (23). It is plausible that amorphous silica or zeolite-like surfaces form discontinuous coatings on feldspars and on the walls of etch-tubes (24–25). For geometrical reasons, angular surfaces, for example where tubes intersect (Fig. 2), might have topological linkages resembling the internal channel walls of silicalite. These sites would automatically have a periodic character on the scale of a few tens of nanomolars to micromolars. Feldspars commonly contain a wide range of mineral inclusions at scales from tens of nanometers to a few micrometers, at concentrations ≪1% by vol, which may be relevant to catalysis and biogenesis, many occurring in primary micropores (26, 27). Inclusions reported are clay minerals, the phosphate mineral apatite, halite, fluorite, carbonates, a Ba compound, and oxides and sulfides of various metals including Pb, Sn, Fe, Ag, Ti, and Mn. Some micropores contain ancient halogen-bearing fluids (28). Because of the large surface area of the honeycombs, and the abundance of feldspar on the early Earth, only a small proportion of feldspar surfaces need have the appropriate catalytic and chemical properties to become highly significant for prebiotic reactions.
Finally, we consider the organizational role of these honeycombs. Somewhere on the prebiotic Earth, areas of partially dissolved alkali feldspar should have been in contact with aqueous organic soups containing simple organic molecules. The feldspars might be at the surface in contact with the atmosphere or within porous, water-bearing sedimentary rocks. Either might have been near hot springs, which have been postulated in several recent treatments of the earliest life (5, 29). The surface temperatures must have been highly variable, as on the present Earth, because of many causes related to latitude, altitude, time of day, weather systems, etc. Volcanic ash containing feldspar crystals would have fallen into lakes and oceans, and lunar tides would have been higher than now causing intense sloshing. On every mm2 of weathered feldspar surface, there would have been 106 catalytic micro-reactors, open by diffusion to the dynamic reservoir of organic molecules in the primordial soups but protected from the dispersive effects of flow and convection in a fully open system and from ultraviolet radiation. Periodic desiccation would have led to strong concentrations of organic molecules, minimizing the possibility of hydrolysis. The reactors normal to the surface would have communicated laterally through the narrower connecting tubes (Fig. 2) so that ever more complex polymeric molecules, catalytically assembled on the silica-rich areas on the tube walls, could have spread through the honeycomb zone. Perhaps, in the protected, self-organized environment of the honeycomb, in a few cross-connected reactors out of the 1018 or so which would occur on a 2.5-km2 granite outcrop, the complex molecules necessary for the first self-replicating polymer would have assembled. Feeding on concentrated soups in adjacent reactors, the polymer would have colonized the feldspar surface, ultimately making the difficult step to adjacent feldspars by surface or grain-boundary diffusion.
The organized, cellular character of the honeycomb offers a possible solution to the problem of the development of a cell wall (5). It is conceivable that once biological evolution had started, progressively more complex self-organized biochemical systems developed in the etch tubes, each behaving as a single “cell”, communicating its genetic material to adjacent cells through the narrow connecting tubes. Possibly a protective lid of lipids evolved across the mouth of the tubes, to stabilize the chemistry within or protect it from dessication. Fed by nutrients, the lids would have extended into the overlying soup, leading ultimately to detachment from the mineral substrate and the emergence of the first truly independent, unicellular life. Is it pure coincidence that the dimensions of the vertical etch-tubes, particularly near their mouths, and the widths of the grooves on highly weathered feldspar surfaces, match exactly the dimensions of most modern soil bacteria (Fig. 1b)? Or are the dimensions of these exceedingly abundant organisms a distant echo of the reactors in which the first life evolved?
Acknowledgments
I.P. and M.R.L. are supported by a Natural Environment Research Council (London) Research Grant.
ABBREVIATIONS
- K-feldspar
potassium-rich alkali feldspar
- SEM
scanning electron microscopy
Footnotes
This is part II of a series. Part I (Biochemical Evolution I: Polymerization on internal, organphilic silica surfaces of dealuminated zeolites and feldspars) was published in the Proceedings (3).
References
- 1.Bernal J D. The Origin of Life. London: Weidenfeld & Nicolson; 1967. [Google Scholar]
- 2.Ferris J P, Hill A R, Jr, Liu R, Orgel L E. Nature (London) 1996;381:59–61. doi: 10.1038/381059a0. [DOI] [PubMed] [Google Scholar]
- 3.Smith J V. Proc Natl Acad Sci USA. 1998;95:3370–3375. doi: 10.1073/pnas.95.7.3370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Cairns-Smith A G. Genetic Takeover and the Mineral Origins of Life. Cambridge, U.K.: Cambridge Univ. Press; 1982. [Google Scholar]
- 5.Russell M J, Hall A J. J Geol Soc (London) 1997;154:377–402. doi: 10.1144/gsjgs.154.3.0377. [DOI] [PubMed] [Google Scholar]
- 6.Lee M R, Parsons I. Geochim Cosmochim Acta. 1995;59:4465–4488. [Google Scholar]
- 7.Lee, M. R., Hodson, M. E. & Parsons, I. (1998) Geochim. Cosmochim. Acta 62, in press.
- 8.Lee M R, Parsons I. J Sediment Res. 1998;68:191–211. [Google Scholar]
- 9.Joyce G E, Orgel L E. In: The RNA World. Gesteland R F, Atkins J F, editors. Plainview, NY: Cold Spring Harbor Lab. Press; 1993. pp. 1–25. [Google Scholar]
- 10.Nutman A P, McGregor V R, Friend C R L, Bennett V C, Kinny P D. Precambrian Res. 1996;78:1–39. [Google Scholar]
- 11.Mojzsis S J, Arrhenius G, McKeegan K D, Harrison T M, Nutman A P, Friend C R L. Nature (London) 1996;384:55–59. doi: 10.1038/384055a0. [DOI] [PubMed] [Google Scholar]
- 12.Schopf J W. In: Early Life on Earth. Bengston S, editor. New York: Columbia Univ. Press; 1994. pp. 193–207. [Google Scholar]
- 13.Eiler J M, Mojzsis S J, Arrhenius G. Nature (London) 1997;386:665. doi: 10.1038/386665a0. [DOI] [PubMed] [Google Scholar]
- 14.Taylor S R. Solar System Evolution. Cambridge, U.K.: Cambridge Univ. Press; 1992. [Google Scholar]
- 15.Lee M R, Waldron K A, Parsons I. Mineral Mag. 1995;59:63–78. [Google Scholar]
- 16.Lee M R, Parsons I. Am Mineral. 1997;82:557–570. [Google Scholar]
- 17.Willaime C, Brown W L. Acta Crystallogr A. 1974;30:316–331. [Google Scholar]
- 18.Blum A E, Stillings L L. In: Chemical Weathering Rates of Silicate Minerals. White A F, Brantley J F, editors. Vol. 31. Washington, DC: Mineral. Soc. Am.; 1995. pp. 291–351. [Google Scholar]
- 19.Hochella M F, Jr, Banfield J F. In: Chemical Weathering Rates of Silicate Minerals. White A F, Brantley J F, editors. Vol. 31. Washington, DC: Mineral. Soc. Am.; 1995. pp. 353–406. [Google Scholar]
- 20.Waldron K, Lee M R, Parsons I. Contrib Mineral Petrol. 1994;116:360–364. [Google Scholar]
- 21.Hellmann R, Eggleston C M, Hochella M F, Jr, Crerar D A. Geochim Cosmochim Acta. 1990;54:1267–1281. [Google Scholar]
- 22.Gout R, Oelkers E H, Schott J, Zwick A. Geochim Cosmochim Acta. 1997;61:3013–3018. [Google Scholar]
- 23.Lee, M. R. & Parsons, I. (1998) Chem. Geol., in press.
- 24.Smith J V. Proc Natl Acad Sci USA. 1998;95:3366–3369. doi: 10.1073/pnas.95.7.3366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Smith J V. In: Feldspars and Their Reactions. Parsons I, editor. Dordrecht, The Netherlands: Kluwer; 1994. pp. 541–595. [Google Scholar]
- 26.Worden R H, Walker F D L, Parsons I, Brown W L. Contrib Mineral Petrol. 1990;104:507–515. [Google Scholar]
- 27.Walker F D L, Lee M R, Parsons I. Mineral Mag. 1995;59:505–534. [Google Scholar]
- 28.Burgess R, Kelley S P, Parsons I, Walker F D L, Worden R H. Earth Planet Sci Lett. 1992;109:147–167. [Google Scholar]
- 29.Vergas M, Kashefi K, Blunt-Harris E L, Lovley D R. Nature (London) 1998;395:65–67. doi: 10.1038/25720. [DOI] [PubMed] [Google Scholar]