

Abstract
• Background and Aims Rhinanthus minor is a root hemiparasitic plant that attacks a wide range of host species which are severely damaged by the parasite. Rhinanthus minor also attempts unsuccessfully to form connections to a range of non-hosts which in contrast are not damaged by the parasite; however, the underlying physiological basis of these differences is not fully understood.
• Methods Biomass of host–parasite combinations was studied, and histology, electron microscopy and FT-IR microspectroscopy were used to determine the cellular-level interactions between Rhinanthus haustoria (the parasite's connective structure) and the roots of a range of potential host species.
• Results Two distinct defence responses were observed in the non-host forbs Plantago lanceolata and Leucanthemum vulgare. Firstly, L. vulgare was able to encapsulate the parasite's invading structures preventing it from gaining access to the stele. This was supported by FT-IR microspectroscopy, used to monitor lignification in response to Rhinanthus haustoria. Secondly, host cell fragmentation was observed at the interface between the parasite and P. lanceolata. Growth data confirmed the non-host status of the two forbs whilst, in contrast, grasses and a legume which were good hosts showed no evidence of defence at the host/parasite interface.
• Conclusions Variable resistance to Rhinanthus is shown for the first time to be controlled by cellular-level resistance to haustoria by either cell fragmentation or lignification at the host/parasite interface.
Keywords: Parasitic plant, Rhinanthus minor (yellow rattle), resistance, host defence, FITR spectroscopy, histology, electron microscopy
INTRODUCTION
Parasitic plants are a group of exploitative organisms that have evolved to utilize another plant's resources (for reviews, see Phoenix and Press, 2005; Press and Phoenix, 2005). They tap into the xylem or phloem stream of their host and withdraw water, mineral nutrients and carbohydrates through the haustorium, a structure which provides vascular continuity from parasite to host (Riopel and Timko, 1995; Dorr, 1997; Gurney et al., 2003). Following successful attachment, hosts show reduced biomass accumulation (Seel et al., 1993; Davies and Graves, 2000) and reproductive output (Gurney et al., 1995). Parasitic plants may also attach to ‘non-host’ plants but, in these instances, the non-host remains unaffected. Damage to the host can be dramatic; for example, the root hemiparasite Striga hermonthica has become an agriculturally significant weed leading to major losses of crop productivity (Parker and Riches, 1993).
The potential host may be able to respond to infection by a parasite through a number of mechanisms. First, the host may innately possess or actively induce a physical barrier against the invading parasite. Previous studies have shown that some potential host species can form an encapsulation layer that the parasite is unable to penetrate at the interface between the two species. This mechanism was observed in a sunflower/Orobanche association (Labrousse et al., 2001) and in the Striga/sorghum (variety ‘Framida’) association (Arnaud et al., 1999). Electron-dense compounds were found to accumulate at the host/parasite interface of Striga and sorghum (Olivier et al., 1991); these may be defence-related compounds such as phytoalexins and lignin. Indeed lignification and cell wall thickening were seen at the point of parasite penetration in the sunflower–Orobanche and cowpea–Striga associations (Jorrin et al., 1996; Lane et al., 1996). It is, however, unclear whether these physical barriers are structurally and chemically distinct from each other. Secondly, the potential host may show localized die-back at the point of infection. Lane et al. (1993) observed physical separation of host and parasite in the form of localized host cell death at the host/parasite interface in a Striga/cowpea association. Goldwasser et al. (2000) also reported cell death, observing necrotic lesions at the interface between the haustorium of Orobanche and vetch roots. Thirdly, the potential host may mount a chemical defence against the parasite, as seen in the phytoalexin accumulation at the interface between Orobanche cernua and sunflower (Jorrin et al., 1996). In contrast to these three defence responses, the potential host may put up no apparent defence at all and be unable to prevent free access to the parasite resulting in the associated negative effects on the host (Cechin and Press, 1993; Gurney et al., 1995; Hibberd et al., 1996; Dorr, 1997).
Most studies of parasitic plant–host interfaces investigate the interaction between a parasite and a single species or group of closely related hosts. This is because many of the parasitic plants of greatest socio-economic importance are host-specific (Press, 1989), and only germinate in the presence of potential hosts (Parker and Riches, 1993). In this case, a host can be defined as a species to which a parasitic plant will attach and which will confer benefits to the parasite in terms of its growth and reproduction. Conversely, a non-host is a species that will not confer any benefits to the parasite. Importantly though, a greater spectrum of responses (from true host to non-host and all stages in between) are displayed by potential hosts as a result of attack by some of the non-host-specific parasitic plants. These catholic parasites and their potential hosts thus represent an excellent opportunity to study a range of host and non-host responses.
Rhinanthus minor (subsequently referred to as Rhinanthus) is a facultative root hemi-parasite, which is a common species in natural and semi-natural grasslands and which is widely distributed throughout northern temperate regions (Westbury, 2004). It has a wide host range extending to at least 20 species (Gibson and Watkinson, 1989) and, in common with other parasitic plants, it significantly affects the biomass of the hosts to which it successfully attaches (Davies and Graves, 2000), although these species do not all represent equally good hosts. Broadly, graminoid and legume species support the greatest parasite biomass (i.e. are better hosts) (Seel and Press, 1993; Cameron, 2004), while the forbs (non-leguminous perennial dicots) support the least (i.e. are poorer hosts) (Seel and Press, 1993; Cameron, 2004). However, reasons for the variable success of Rhinanthus on different species have never been fully elucidated (Cameron et al., 2005). The aim of this study, therefore, is to investigate whether host and non-host resistance is implicated in the variable success of Rhinanthus and the subsequent variation in host damage. To evaluate these interactions, a novel combination of growth studies, histological analysis of haustorial morphology, and FT-IR microspectroscopy of the parasite–host interface, has been employed to analyse the nature of any resistance to Rhinanthus.
MATERIALS AND METHODS
Plant material
Ten 2-week-old seedlings of each potential host [the grasses Cynosurus cristatus L. and Phleum bertolonii DC. (both Poaceae), a legume Vicia cracca (Leguminosae) and two forbs Leucanthemum vulgare Lam. (Asteraceae) and Plantago lanceolata L. (Plantaginaceae)] were grown for 4 weeks in 15-cm-diameter pots (one plant per pot) containing 50 : 50 sand : John Innes No. 3 compost. Rhinanthus seeds were surface sterilized for 5 min in 3 % sodium hypochlorite solution, washed in distilled water and preconditioned on moist filter paper at 4 °C until germination (approx. 8 weeks) (Keith et al., 2004). Four seedlings of Rhinanthus were transplanted into five of the pots for each host species. The parasites were subsequently reduced to one per host when the first parasite showed morphological changes associated with attachment (Klaren and Janssen, 1978). Water was supplied to the soil daily and the pots were arranged in a randomized block design with replicate number (1–5) as the blocking factor. The hosts, together with the parasites, were then grown for a further 14 weeks (after parasite attachment) in a glasshouse (temperature 28 °C max, 16 °C min). The total above-ground biomass of both host and parasite was harvested after 14 weeks, dried in an oven at 80 °C for 2 d, and the biomass recorded along with the number of parasite flowers. The below-ground biomass was not measured as the roots of the host and parasite were not readily distinguishable.
Primary haustoria were removed from the fresh roots and preserved in 2 % glutaraldehyde and 2·5 % paraformaldehyde in 0·1 m phosphate buffer (pH 7·2), placed under vacuum for 10 min and then stored at 4 °C for a further 7 d to allow the fixative to penetrate the tissue. Roots from uninfected hosts were also collected and fixed in the same way. After fixatation the haustoria were washed in 0·1 m phosphate buffer (pH 7·2) and dehydrated through a graduated ethanol series.
Embedding procedure for light microscopy (LM)
Tissue was embedded in LR-White resin (TAAB, UK) and ten haustoria per replicate plant (five plants in total) from each of the five host species were serially sectioned at 2-μm intervals (between 80 and 100 sections were cut per haustorium). Sections were floated onto polysine microscope slides (TAAB), placed on an 80 °C hot plate for 5 min to dry and were stained on the hotplate with 0·5 % toluidine blue O in boric acid for 30 s. Sections were permanently mounted using histomount (TAAB) and a glass cover slip and examined using a Leitz Laborlux 5 microscope (Leica, Milton Keynes, UK) in bright field. Images were captured using a Polaroid DMC Ie CCD digital camera (Datacell, Berkshire, UK).
Haustoria preparation for scanning electron microscopy (SEM)
Ten haustoria per replicate plant (five plants in total) from the five host species were fixed and dehydrated as detailed above. Haustoria were cut in half longitudinally and dried by being introduced into liquid carbon dioxide. The liquid carbon dioxide was brought to its critical point; this is critical as it avoids the problem of surface tension, during drying, resulting in disruption to tissue and causing visible distortions. After drying, the samples were attached to aluminium pegs and the surface of the tissue coated in a vacuum with an electrically conductive layer of gold. Haustoria were examined using a Cambridge 590B scanning electron microscope (LEO Electron Microscopy, Cambridge, UK). Images were captured digitally.
Haustoria preparation and sectioning for transmission electron microscopy (TEM)
Ten haustoria per replicate plant from the five host species were fixed and dehydrated as detailed above with the exception of tissue being infiltrated with 1 % osmium tetraoxide for 24 h prior to the ethanol series and washed clean with phosphate buffer before de-hydration. Tissue was embedded in epoxy resin (EPON; TAAB) and gold thickness (80–100 nm) ultra thin serial sections were cut and floated onto 300-mesh copper grids (TABB) and examined using a Philips 301 transmission electron microscope (FEWE Electron Optics, Cambridge, UK). Images were captured using 35-mm film photography and digitized using Adobe Photoshop Elements v2.
Procedure for Fourier-transform infrared (FT-IR) micro-spectroscopy
FT-IR micro-spectroscopy can probe the chemical structure of a given material from sample areas down to approx. 50 × 50 μm (Coats et al., 2003). The technique combines high spatial precision with high chemical specificity. In this study, FT-IR micro-spectroscopy was used to identify the chemical constituents of areas of the sections of the haustoria and host roots based on published spectra for a range of molecules, including key plant defence compounds such as lignin (Seca et al., 1998) and suberin (Cordeiro et al., 1998; Zeier and Schreiber, 1999). Samples were embedded in the same way as for LM and haustoria from each replicate host (four biological replicates, three spectra from each interface) sectioned to 2 μm and floated on to barium fluoride discs (Linkam, UK) using double-distilled water, then allowed to air dry for 2 d.
The host–parasite interface (Fig. 1) was analysed in all five host species using FT-IR micro-spectroscopy. All infrared spectra were measured in transmission mode using a Spectra Tech continuum microscope coupled to a Nicolet Nexus spectrometer. A resolution of 4 wave numbers (cm−1) was used with 64 scans. Water and carbon dioxide absorption bands were eliminated by purging the system with dry air before and during data collection. Spectra were measured from areas of 200 × 100 μm at the interface between the host and the parasite in infected roots and in the same region of uninfected roots (Fig. 1). This sample area maximized the signal to noise ratio in the spectra, but avoided signal dilution effects of sampling regions outside the haustorial interface. Spectra of individual haustoria from four separate host plants per host species were taken. The spectra from this tissue were compared with those of two known key plant defence compounds: lignin (Seca et al., 1998; Yao et al., 2003) and suberin (Cordeiro et al., 1998). The frequencies of bands associated with lignin and suberin were assigned relative to the reference spectrum with a tolerance of ±4 cm−1 (data not shown). Due to the relatively low signal to noise ratio below 900 cm−1 only those samples with absorption bands at least double the baseline intensity (and hence clearly distinct) were included for this region.
Fig. 1.

Host roots uninfected (A) and infected with the parasitic plant Rhinanthus minor (B) showing host root cortex (1), endodermis (2), stele (3), FT-IR sample collection region is represented by the dashed box and, where present, host/parasite interface (4), parasite endophyte (5) and parasite haustorium (6).
Statistical analysis
Differences between treatment means were analysed by ANOVA and Fisher's multiple comparison test using Minitab 13 (Minitab Inc., PA, USA). Where necessary to satisfy the test assumptions, data were arcsine square root transformed. Untransformed means and associated standard errors are presented. Principal component analysis (PCA) was used to elucidate changes in the lignin and suberin FT-IR profiles pre- and post-infection by R. minor using the PC-Ord multivariate analysis package (MJM Software Design, Gleneden Beach, OR, USA).
RESULTS
Growth study
The grasses (C. cristatus and P. bertolonii) and legume (V. cracca) were effective hosts for Rhinanthus in terms of the parasite's growth (Fig. 2A) and reproduction (Fig. 2B). In contrast, the two forb species (P. lanceolata and L. vulgare) were ineffective hosts, the parasite achieving significantly lower biomass than when grown without a host (Fig. 2A, B). Rhinanthus significantly reduced the growth of both grasses (hosts), but did not significantly reduce the biomass of the legume (host) or either forb species (non-hosts) (Fig. 3).
Fig. 2.
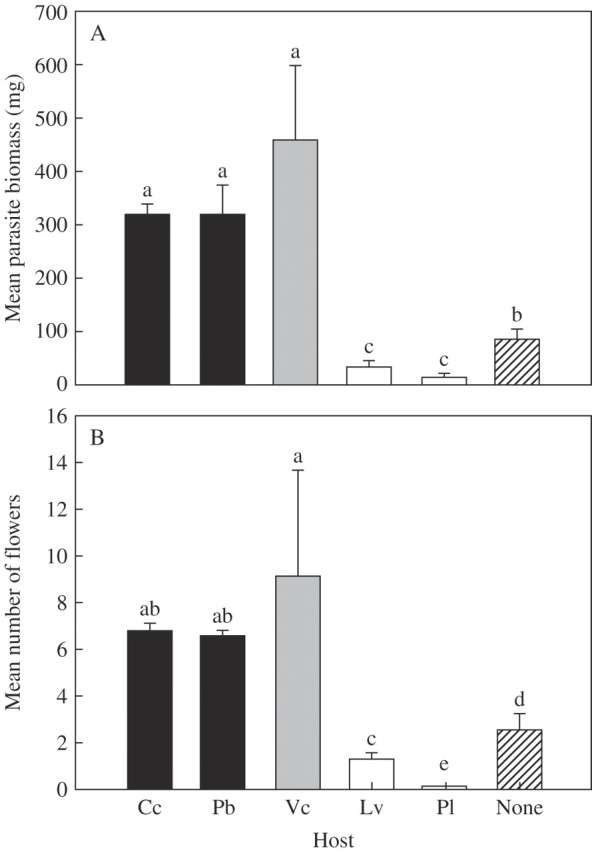
Biomass (A) and number of flowers (B) of the parasitic plant Rhinanthus minor attached to five potential hosts: grasses C. cristatus (Cc) and P. bertolonii (Pb); a legume V. cracca (Vc) and two forbs L. vulgare (Lv) and P. lanceolata (Pl). Columns sharing the same letter are not significantly different [P > 0·05; Fisher's multiple comparison test following one-way ANOVA (biomass, df = 5,26; F = 17·37; P < 0·001; number of flowers, df = 5,25; F = 60·54; P < 0·001)—biomass data were log10-transformed and flower data were arcsin square root transformed prior to ANOVA analysis].
Fig. 3.

Biomass of five potential hosts infected with Rhinanthus minor (black columns) and uninfected (white columns): grasses C. cristatus (Cc) and P. bertolonii (Pb); a legume V. cracca (Vc) and two forbs L. vulgare (Lv) and P. lanceolata (Pl). Pairs of columns marked with asterisks are significantly different (ANOVA: *, P < 0·05; ***, P < 0·001).
Histology
LM revealed extensive host penetration by the parasite in both grasses and the legume, with the formation of xylem bridges by the parasite (Fig. 4A–C). Conversely, in the two forbs the parasite did not penetrate into the vascular tissues. At the interface with the forbs, a darkly staining layer formed between the host and parasite tissue (Fig. 4D). In P. lanceolata there was localized host cell fragmentation and heavy staining at the point of attempted parasite attachment (Fig. 4E).
Fig. 4.
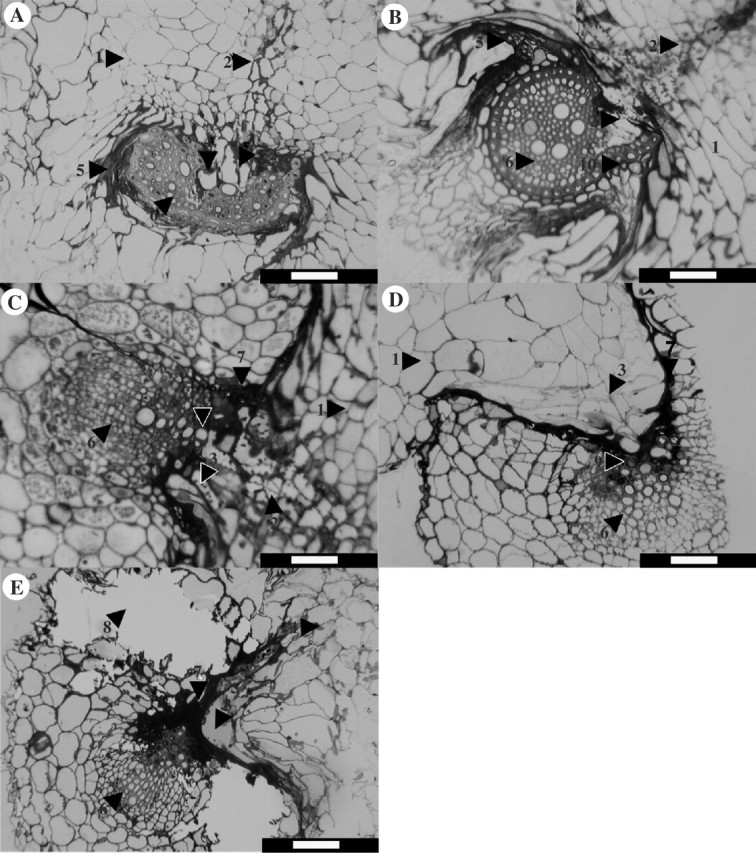
Transverse sections through the haustoria of the parasitic angiosperm Rhinanthus minor attacking the roots of the grasses C. cristatus (A) and P. bertolonii (B), a legume V. cracca (C) and forbs L. vulgare (D) and P. lanceolata (E). Sections were 2 μm thick and were stained with toluidine blue for 30 s. The numbers indicate: 1, parasite haustorium; 2, secondary parasite xylem; 3, penetration peg (endophyte); 4, parasite penetrating host xylem; 5, crushed host cortical cells; 6, host stele; 7, darkly staining material; 8, fragmented host cells; 9, occluded cells; 10, peeled endodermis. Scale bars = 55 μm.
Further investigation of haustoria using SEM confirmed parasite access to the stele of both grass and legume roots (Fig. 5A–E). In the latter case, oscula [defined by Dorr (1997) as conduits between parasite and host], were observed penetrating the host xylem. Parasite attachment to the two forbs was confirmed not to compromise host vasculature (Fig. 5F, G) and in P. lanceolata all haustoria studied had lesions of fragmenting cells (Fig. 5G).
Fig. 5.
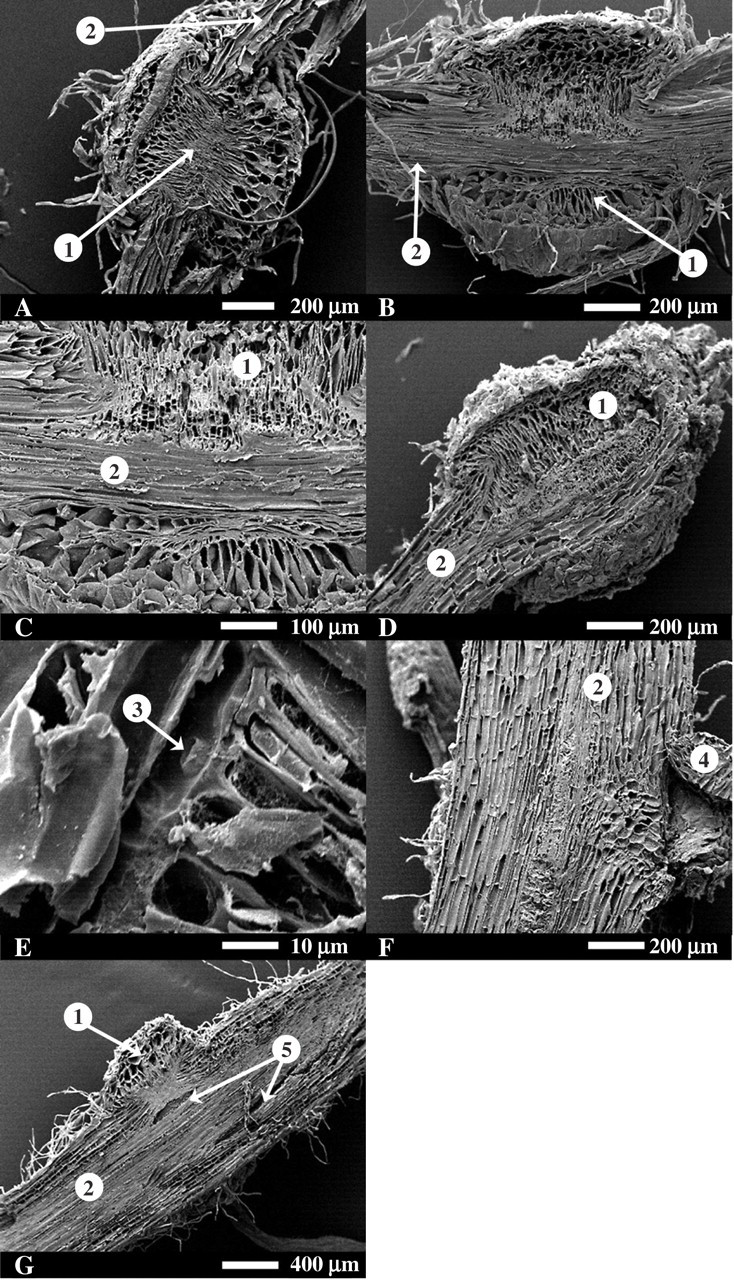
Scanning electron micrographs of longitudinal cuts through the haustoria of the parasitic angiosperm Rhinanthus attacking the roots of C. cristatus (A), P. bertolonii (B and C), V. cracca (D and E), L. vulgare (F) and P. lanceolata (G). The numbers indicate: 1, parasite haustorium; 2, host root; 3, osculum; 4, lateral root; 5, lesions in host root. Scale bars are shown with each image. The parasite haustorium in (D) is behind the host root.
Ultrastructural analysis of the host/parasite interface by TEM reconfirmed the vascular penetration of the grasses and legume by the parasite (Fig. 6A–C). In front of the invading parasite, the cell walls of C. cristatus and their middle lamellae appeared to be fragmenting (Fig. 6A). Additionally, crushed host cells at the parasite interface with P. bertolonii were observed (Fig. 6A).
Fig. 6.
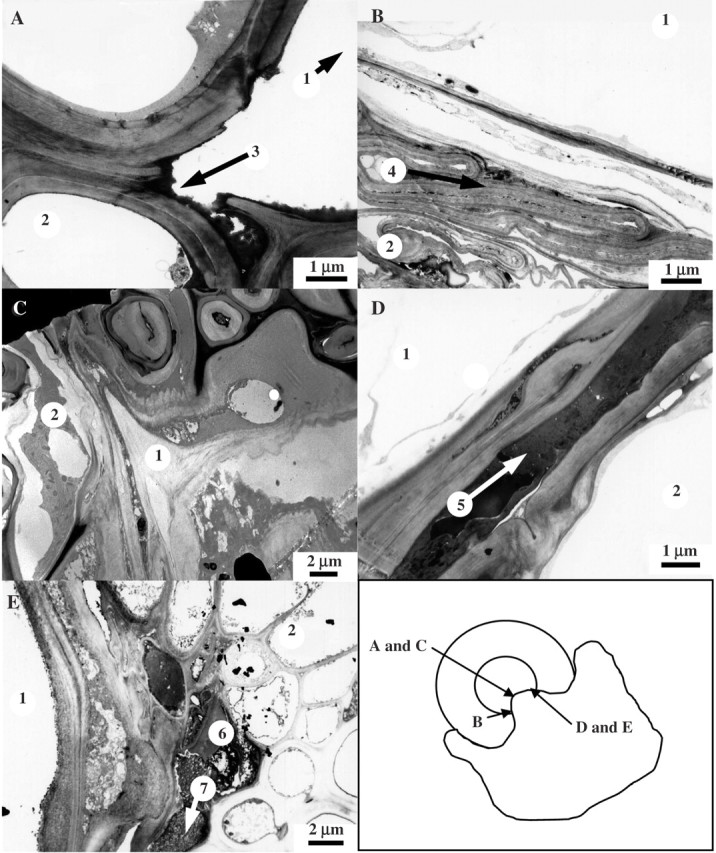
Transmission electron micrographs of transverse sections through the haustoria of the parasitic angiosperm Rhinanthus attacking the roots of C. cristatus (A), P. bertolonii (B) V. cracca (C), L. vulgare (D) and P. lanceolata (E). The numbers indicate: 1, host tissue; 2, parasite tissue; 3, host cells degraded by the parasite; 4, crushed host cortical cells; 5, layer of dark staining material; 6, dead/decaying host cells; 7, coagulated cytoplasm (indicative of hypersensitive response). Scale bars are shown with each image.
In the forbs, parasite penetration was highly restricted and failed to reach the vasculature. Electron-dense material accumulated in the contact zone between L. vulgare and Rhinanthus (Fig. 6D). A three-cell-deep layer of dying host cells containing dark coagulated cytoplasm occurred at the interface between P. lanceolata and Rhinanthus (Fig. 6E).
FT-IR micro-spectroscopy analysis
The host–parasite interface (Fig. 1) was analysed in all five host species using FT-IR. FT-IR spectra of the parasite interface of two potential host species are shown in Fig. 7. There are eight peaks associated with lignin at the interface between Rhinanthus and L. vulgare (Fig. 7A), while the parasite interface with C. cristatus shows no lignin bands (Fig. 7B). PCA was used to elucidate a pattern of the presence or absence of FT-IR absorption bands attributed to lignin and suberin in infected tissues relative to uninfected controls. Figure 8A shows a PCA plot for lignin peaks in infected and uninfected samples. There is some clustering of infected versus uninfected samples, indicating differences in the composition of lignins in infected versus uninfected tissue. The number of lignin-associated bands detected was generally increased by infection, but this effect was only significant for L. vulgare (Fig. 9A; see Table 1 for statistical analysis). The PCA plot for bands associated with suberin showed no clustering (Fig. 8B) and there were no significant differences in the numbers of suberin bands detected between infected and uninfected tissues of any species (Fig. 9B; see Table 1 for statistical analysis).
Fig. 7.

Fourier-transform infrared spectra of (A) C. cristatus and (B) L. vulgare. Dotted lines show the occurrence of bands/peaks associated with lignin and the wavelength (cm−1).
Fig. 8.
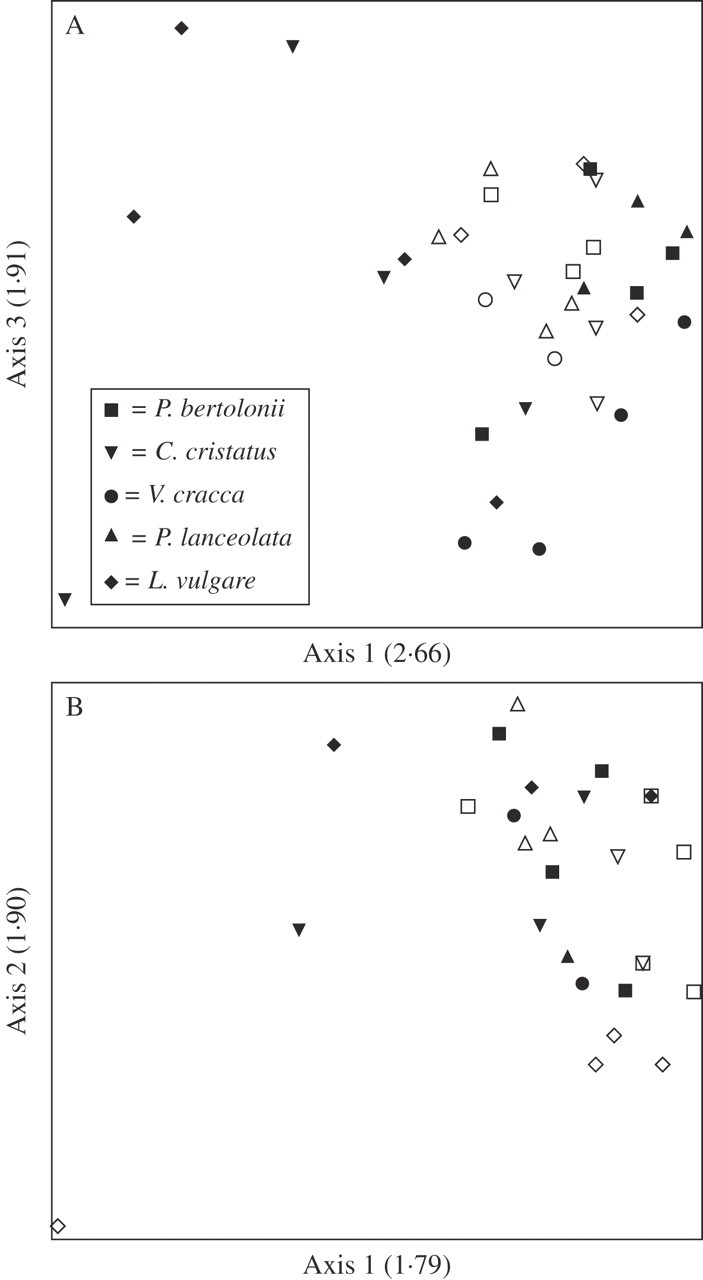
Principal component analysis plots of bands associated with (A) lignin and (B) suberin. Open symbols represent infected hosts and closed symbols represent uninfected hosts. Eigenvalues for each axis are given in parentheses.
Fig. 9.
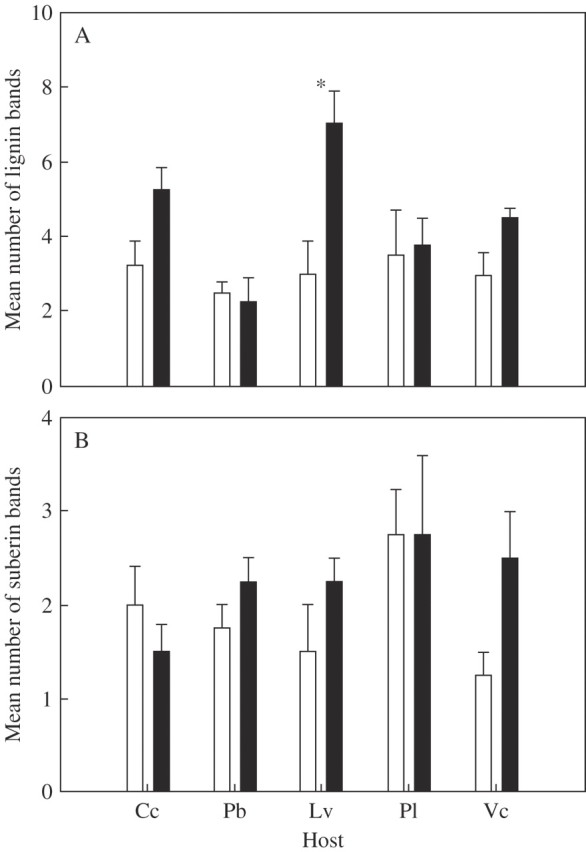
The number of detectable bands in the FT-IR spectra associated with (A) lignin and (B) suberin. Error bars represent 1 s.e. Bars sharing asterisks are significantly different (ANOVA: P < 0·05). Black columns are infected roots, white columns uninfected roots. n = 4.
Table 1.
One-way ANOVA output for number of detectable bands in the FT-IR spectrum associated with lignin and suberin in host root tissue infected with Rhinanthus compared with uninfected tissue for five potential hosts
| Lignin | Suberin | |||||
|---|---|---|---|---|---|---|
| Species | d.f. | P | df | P | ||
| Cynosurus cristatus | 7 | (as) | 0·065↑ | 7 | (as) | 0·372↓ |
| Phleum bertolonii | 7 | (as) | 0·658↓ | 7 | (as) | 0·209↑ |
| Plantago lanceolata | 7 | (as) | 0·852↑ | 7 | (as) | 0·075↑ |
| Leucanthemum vulgare | 7 | (as) | 0·025*↑ | 7 | (as) | 0·929− |
| Vicia cracca | 7 | (as) | 0·062↑ | 7 | (bc) | 0·097↑ |
The type of data transformation is given in parentheses in front of the P value: as, arc sin square root transformation; bc, box cox transformation (performed using Mintab version 13).
↑, An increase in bands in infected tissue; ↓, a decrease in bands in infected tissue; −, no change relative to uninfected tissue.
* Significant difference in number of lignin bands P < 0·05.
DISCUSSION
Using five different host species, this study has established for the first time that the success of the hemiparasitic plant, Rhinanthus minor is intrinsically linked to the ability of the host to mount successful defences against the invading haustorium. Given the clear relationship between the defence capacity (of the potential host) and Rhinanthus success in the present study, it seems reasonable to propose such defence mechanisms may play a role in mediating Rhinanthus success. The implications of this therefore, have considerable broad importance in understanding the impacts of Rhinanthus, both on host plants and on communities as a whole. Rhinanthus is considered a key tool in the restoration of meadow systems owing to its suppression of grasses (‘good hosts’) in the field, allowing increases in abundance of forb species which are less affected by Rhinanthus (Westbury and Davies, 2005). From the present study it can now be seen how such community-level impacts can be driven by physiological and morphological responses at the parasite/host interface.
Variation in the success of haustorial formation
The grasses and legume were all susceptible to the parasite. Rhinanthus was able to colonize their roots and compromise their vascular system, forming well-developed haustoria. These haustoria encompassed the host root, crushing the cortex, and developed a penetration peg containing differentiated secondary xylem. In contrast, haustorial development on the forbs was poor. The parasite, in all replicates studied, failed to penetrate forb vasculature. The resultant haustoria were poorly developed, lacking hyaline bodies and never encompassing the host root. In some examples, penetration pegs were formed, although these always lacked differentiated secondary xylem.
If the haustorium is successful, the parasite is likely to access host resources both through the apoplast and symplast by accessing the host vasculature. The parasite can directly access host vessels through structures known as oscula, defined by Dorr (1997) as tube-like structures which act as direct conduits between host and parasite. The SEM studies of haustoria revealed oscula penetrating host xylem in the Rhinanthus–V. cracca association; the first time these structures have been reported in Rhinanthus. The TEM studies revealed that in the C. cristatus–Rhinanthus association the host cells and the middle lamella were fragmenting ahead of the invading parasite endophyte in all sections viewed. This implicates extracellular enzyme activity in the penetration process. This is consistent with other indirect studies of cell-wall degrading enzymes or their products in the haustorial interface of Orobanche–Helianthus annuus (Losner-Goshen et al., 1998) and Rhinanthus–C. cristatus (D. Kraushaar, D. D. Cameron and W. E. Seel, unpubl. res.). In the latter case the products of pectinase were localized in infected grass roots, although this was not observed in any of the forbs studied.
Fragmented host cells present at the interface between P. lanceolata and Rhinanthus were seen using LM. Moreover, a three-cell-deep layer of fragmented host cells bordering the juncture with the parasite was seen in the TEM. These cells were distinct from the rest of the host cells and the cytoplasm was vacuolized, a sign of localized cell death or hypersensitive response (HR) (Greenberg and Yao, 2004). HR is a widely observed plant response to fungal attack and has been shown to serve as a successful defence against fungal pathogens (Silva et al., 2002). It is initiated by parasite/pathogen-derived compounds, elicitors, which trigger a gene-for-gene response leading to the HR. The gene-for-gene model of plant defence suggests that if a potential host is resistant to a parasite/pathogen then it will have a resistance gene (R) which corresponds to an avirulence (avr) gene in the parasite/pathogen (Takken and Joosten, 2000). However, in the absence of the potential host R gene, the avr genes do provide a selective advantage to the parasite (Bonas and Lahaye, 2002). The reasons underlying the lack of this response in the other species is unclear; however, pathogenic and mycorrhizal fungi can suppress the production of some or all of these elicitors which in turn can prevent the expression of HR (for a review, see Greenberg and Yao, 2004), thus the ability of the parasite to suppress the potential host's HR response (Heath, 1998) may account for the lack of HR in the other hosts studied, including L. vulgare.
Encapsulation and lignification at the interface between other parasitic plants and their hosts has also been reported by Labrousse et al. (2001), Arnaud et al. (1999), Olivier et al. (1991) and Jorrin et al. (1996). They hypothesized that this barrier served as a resistance mechanism preventing the parasite accessing the host's vasculature. In the present study, encapsulation of the parasite endophyte was observed in the LM sections which showed that Rhinanthus did not compromise the xylem of either of the forbs. Additionally, the parasite endophyte was surrounded by a dense, darkly stained material most notably in L. vulgare where there was also an accumulation of electron-dense material at the interface between the host and the parasite in the TEM electron micrographs.
Changes in lignification
The darkly staining material was investigated using FT-IR micro-spectroscopy by comparing infra-red spectra of infected and un-infected host roots with published spectra for two key defence compounds, lignin and suberin. Multivariate analysis of the spectra indicated that there were changes in the spectrum of lignin when plants were colonized by the parasite. This was most notable in the forb L. vulgare where there was a statistically significant increase in the detectable number of bands in the spectrum associated with lignin. In contrast, when the same analysis was undertaken for suberin, another likely defence compound, there was no change in the spectra when potential hosts were attacked. Changes in lignin chemistry in the presence of the parasite may be expected since defence and structural lignins are known to differ in their chemical composition (Hawkins and Boudet, 2003). However, it is surprising that an increase in lignin bands associated with one potential host species (L. vulgare), correlating with the darkly staining material seen at the interface, was only detected in this species. Sensitivity to the parasite and the subsequent induction of lignification in response thus appears to be a significant factor determining the expression of a successful defence response. Moreover, during colonization, pathogenic (Niks and Rubiales, 2002, Greenberg and Yao, 2004) and mycorrhizal fungi (Kapulnik et al., 1996) have the capacity to suppress the pathway which leads to the biosynthesis of lignin. The potential for parasitic plants to interrupt the defence chemistry of compatible hosts needs to be investigated further.
Growth responses
The growth and reproductive success of the parasite and the subsequent parasite-induced host damage are strongly related. It has been shown that when the parasite achieves the greatest biomass it inflicts the most damage on the host. The grasses and legume proved very good hosts for Rhinanthus; the parasite grew significantly larger and produced more flowers compared with host-less individuals. The biomass of the grasses was significantly suppressed by the parasite and these species were successfully colonized by the parasite. Neither of the grasses or the legume mounted successful resistance responses and appeared to be equally susceptible. However, it is perplexing that, in contrast, the biomass of the legume V. cracca was not suppressed, despite the parasite being able to colonize its roots successfully. The reasons for this are unclear. However, V. cracca is a vigorous species and, as a legume, is not reliant on soil nitrogen; thus is perhaps able to tolerate the drain on its resources inflicted by the parasite.
The two forbs were not significantly damaged by the parasite and the parasite achieved a small biomass and produced few flowers. Indeed when growing with the forbs, Rhinanthus performance was worse than when it had no host. Rhinanthus could not compromise the vasculature of these species and, although haustoria were formed, they were poorly differentiated. Therefore, at least one of the factors underlying the poor parasite growth on the forbs is its inability to successfully colonize their roots, and so the forbs studied do not represent true hosts. How far it is possible to generalize the occurrence of successful defences in all forbs is unclear and requires further investigation.
Wider implications
We have highlighted that such varying growth and physiological responses to Rhinanthus may have important implications at the community level. Rhinanthus minor has been shown to significantly influence the structure of its grassland habitat (Gibson and Watkinson, 1992) by suppressing grasses and thus releasing forbs from competitive exclusion and enhancing botanical diversity (Westbury, 2004; Ameloot et al., 2005). In an earlier paper, Gibson and Watkinson (1991) suggest that this effect is mediated by host selectivity, although some forbs were considered preferred hosts. The present findings suggest that an additional or alternative mechanism may be in place, supporting the idea that variable resistance to the parasite by different host groups may be implicated in determining community-level effects (Westbury, 2004). Alternatively, variable resistance could indeed be an integral part of the host selectivity suggested by Gibson and Watkinson (1991).
Supplementary Material
Acknowledgments
We would like to thank Daniel Kraushaar (Georgia, USA) for help with processing the FT-IR data, Prof. Wolfram Hartung, Dr Fan Jiang (Würzburg, Germany), Dr Arthur Berg, and Dr Sarah Dalrymple (Aberdeen, UK) for useful discussions. We also thank Drs Jonathan Leake, Barry Lomax and Gareth Phoenix (Sheffield, UK) for critical reading of the manuscript, Kevin MacKenzie (Aberdeen, UK) for help with the microscopy and Janet Woo, David Hadwen and Norma McKay (Aberdeen, UK) for technical support. This research was funded by The Natural Environment Research Council (UK) (to D.D.C. and W.E.S.) and The Arthritis Research Campaign (to A.M.C.).
LITERATURE CITED
- Ameloot E, Verheyen K, Hermy M. (2005) Meta-analysis of standing crop reduction by Rhinanthus spp. and its effect on vegetation structure. Folia Geobotanica 40289–310. [Google Scholar]
- Arnaud MC, Véronési C, Thalouan P. (1999) Physiology and histology of resistance to Striga hermonthica in Sorghum bicolor var. Framida. Australian Journal of Plant Physiology 2663–70. [Google Scholar]
- Bonas U and Lahaye T. (2002) Plant disease resistance triggered by pathogen-derived molecules: refined models of specific recognition. Current Opinion in Microbiology 544–50. [DOI] [PubMed] [Google Scholar]
- Cameron DD. (2004) A role for differential host resistance to the hemiparasitic angiosperm, Rhinanthus minor, in determining the structure of host plant communities?(University of Aberdeen, UK) PhD Thesis.
- Cameron DD, Hwangbo J-K, Keith AM, Geniez J-M, Kraushaar D, Rowntree J, et al. (2005) Interactions between the hemiparasitic angiosperm Rhinanthus minor and its hosts: from the cell to the ecosystem. Folia Geobotanica 40217–230. [Google Scholar]
- Cechin I and Press MC. (1993) Nitrogen relations of the Sorghum–Striga hermonthica host–parasite association: growth and photosynthesis. Plant, Cell and Environment 16237–247. [DOI] [PubMed] [Google Scholar]
- Coats AM, Hukins DWL, Imrie CT, Aspden RM. (2003) Polarization artefacts of an FTIR microscope and the consequences for intensity measurements on anisotropic materials. Journal of Microscopy 21163–66. [DOI] [PubMed] [Google Scholar]
- Cordeiro N, Belgacem MN, Silvestre AJD, Neto CP, Gandini A. (1998) Cork suberin as a new source of chemicals. 1. Isolation and chemical characterization of its composition. International Journal of Biological Macromolecules 2271–80. [DOI] [PubMed] [Google Scholar]
- Davies DM and Graves JD. (2000) The impact of phosphorus on interactions of the hemiparasitic angiosperm Rhinanthus minor and its host Lolium perenne. Oecologia 124100–106. [DOI] [PubMed] [Google Scholar]
- Dorr I. (1997) How Striga parasitizes its host: a TEM and SEM study. Annals of Botany 79463–472. [Google Scholar]
- Gibson CC and Watkinson AR. (1989) The host range and selectivity of a parasitic plant—Rhinanthus minor L. Oecologia 78401–406. [DOI] [PubMed] [Google Scholar]
- Gibson CC and Watkinson AR. (1991) Host selectivity and the mediation of competition by the root hemiparasite. Rhinanthus. Oecologia 8681–87. [DOI] [PubMed] [Google Scholar]
- Gibson CC and Watkinson AR. (1992) The role of the hemiparasitic annual Rhinanthus minor in determining grassland community structure. Oecologia 8962–68. [DOI] [PubMed] [Google Scholar]
- Goldwasser Y, Plakhine D, Kleifeld Y, Zamski E, Rubin B. (2000) The differential susceptibility of vetch (Vicia spp.) to Orobanche aegyptiaca: anatomical studies. Annals of Botany 85257–262. [Google Scholar]
- Greenberg JT and Yao N. (2004) The role and regulation of programmed cell death in plant–pathogen interactions. Cellular Microbiology 6201–211. [DOI] [PubMed] [Google Scholar]
- Gurney AL, Press MC, Ransom JK. (1995) The parasitic angiosperm Striga hermonthica can reduce photosynthesis of its sorghum and maize hosts in the field. Journal of Experimental Botany 461817–1823. [Google Scholar]
- Gurney AL, Grimanelli D, Kanampiu F, Hoisington D, Scholes JD, Press MC. (2003) Novel sources of resistance to Striga hermonthica in Tripsacum dactyloides, a wild relative of maize. New Phytologist 160557–568. [DOI] [PubMed] [Google Scholar]
- Hawkins S and Boudet A. (2003) ‘Defence lignin’ and hydroxycinnamyl alcohol dehydrogenase activities in wounded Eucalyptus gunnii. Forest Pathology 3391–104. [Google Scholar]
- Heath MC. (1998) Apoptosis, programmed cell death and the hypersensitive response. European Journal of Plant Pathology 104117–124. [Google Scholar]
- Hibberd JM, Quick WP, Press MC, Scholes JD. (1996) The influence of the parasitic angiosperm Striga gesnerioides on the growth and photosynthesis of its host, Vigna unguiculata. Journal of Experimental Botany 47507–512. [Google Scholar]
- Jorrin J, De Ruck E, Serghini K, Perez de Luque A, Munoz-Garcia J. Cubero JI, Berner D, Joel DM, Mussleman LJ, Parker C. (1996) Biochemical aspects of the parasitism of sunflower by Orobanche. Advances in parasitic plant research. Proceedings of the 6th international parasitic weed symposium, Cordoba, Junta, Andalusia.
- Kapulnik Y, Volpin H, Itzhaki H, Ganon D, Galili S, David R, et al. (1996) Suppression of defence responses in mycorrhizal alfalfa and tobacco roots. New Phytologist 13359–64. [Google Scholar]
- Keith AM, Cameron DD, Seel WE. (2004) Spatial interactions between the hemiparasitic angiosperm Rhinanthus minor are species specific. Functional Ecology 18435–442. [Google Scholar]
- Klaren CH and Janssen G. (1978) Physiological changes in the hemi-parasite Rhinanthus minor serotinus before and after attachment. Physiologia Plantarum 42151–155. [Google Scholar]
- Labrousse P, Arnaud MC, Serieys H, Berville A, Thalouarn P. (2001) Several mechanisms are involved in resistance of Helianthus to Orobanche cumana Wallr. Annals of Botany 88859–868. [Google Scholar]
- Lane JA, Bailey JA, Butler RC, Terry PJ. (1993) Resistance of cowpea [Vigna unguiculata (L) walp] to Striga gesnerioides (willd) vatke, a parasitic angiosperm. New Phytologist 125405–412. [DOI] [PubMed] [Google Scholar]
- Lane JA, Moore THM, Child DV, Bailey JA. (1996) Post-infection resistance mechanisms against Striga in cowpea and sorghum. In Cubero JI, Berner D, Joel DM, Mussleman LJ, Parker C (Eds.). Advances in parasitic plant research. Proceedings of the 6th international parasitic weed symposium(Junta Andalusia, Cordoba, Spain).
- Losner-Goshen D, Portnoy VH, Mayer AM, Joel DM. (1998) Pectolytic activity by the haustorium of the parasitic plant Orobanche L. (Orobanchaceae) in host roots. Annals of Botany 81319–326. [Google Scholar]
- Niks RE and Rubiales D. (2002) Potentially durable resistance mechanisms in plants to specialised fungal pathogens. Euphytica 124201–216. [Google Scholar]
- Olivier A, Benhamou N, Leroux GD. (1991) Cell-surface interactions between sorghum roots and the parasitic weed Striga hermonthica: cytochemical aspects of cellulose distribution in resistant and susceptible host tissues. Canadian Journal of Botany 691679–1690. [Google Scholar]
- Parker C and Riches CR. (1993) Parasitic weeds of the world: biology and control(CAB International, Wallingford).
- Phoenix GK and Press MC. (2005) Linking physiological traits to impacts on community structure and function: the role of root hemiparasitic Orobanchaceae (ex Scrophulariaceae). Journal of Ecology 9367–78. [Google Scholar]
- Press MC. (1989) Autotrophy and heterotrophy in root hemiparasites. Trends in Ecology and Evolution 4258–263. [DOI] [PubMed] [Google Scholar]
- Press MC and Phoenix GK. (2005) Impacts of parasitic plants on natural communities. New Phytologist 166737–751. [DOI] [PubMed] [Google Scholar]
- Riopel JL and Timko MP. (1995) Haustorial initiation and differentiation. In Press MC and Graves JD (Eds.). Parasitic plants(Chapman and Hall, London) pp. 39–73.
- Seca AML, Cavaleiro JAS, Domingues FMJ, Silvestre AJD, Evtuguin D, Neto CP. (1998) Structural characterization of the bark and core lignins from kenaf (Hibiscus cannabinus). Journal of Agricultural and Food Chemistry 463100–3108. [Google Scholar]
- Seel WE and Press MC. (1993) Influence of the host on three sub-arctic annual facultative root hemiparasites. 1. Growth, mineral accumulation and aboveground dry matter partitioning. New Phytologist 125131–138. [DOI] [PubMed] [Google Scholar]
- Seel WE, Cooper RE, Press MC. (1993) Growth, gas-exchange and water-use efficiency of the facultative hemiparasite Rhinanthus minor minor associated with hosts differing in foliar nitrogen concentration. Physiologia Plantarum 8964–70. [Google Scholar]
- Silva MC, Nicole M, Guerra-Guimaraes L, Rodrigues CJ. (2002) Hypersensitive cell death and post-haustorial defence responses arrest the orange rust (Hemileia vastatrix) growth in resistant coffee leaves. Physiological and Molecular Plant Pathology 60169–183. [Google Scholar]
- Takken FLW and Joosten MHAJ. (2000) Plant resistance genes: their structure, function and evolution. European Journal of Plant Pathology 106699–713. [Google Scholar]
- Westbury DW and Davies A. (2005) Yellow rattle—its natural history and use in grassland restoration. British Wildlife pp. 93–98.
- Westbury DW. (2004) Rhinanthus minor L. Biological flora of the British Isles. Journal of Ecology 92906–927. [Google Scholar]
- Yao J, Xu XW, Feng YY. (2003) FTIR studies on the chemical composition of wheat straw in different layers. Spectroscopy and Spectral Analysis 2358–60. [PubMed] [Google Scholar]
- Zeier J and Schreiber L. (1999) Fourier transform infrared-spectroscopic characterisation of isolated endodermal cell walls from plant roots: chemical nature in relation to anatomical development. Planta 209537–542. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}