

Abstract
It has long been suspected that the relative abundance of specific nutrients can affect cognitive processes and emotions. Newly described influences of dietary factors on neuronal function and synaptic plasticity have revealed some of the vital mechanisms that are responsible for the action of diet on brain health and mental function. Several gut hormones that can enter the brain, or that are produced in the brain itself, influence cognitive ability. In addition, well-established regulators of synaptic plasticity, such as brain-derived neurotrophic factor, can function as metabolic modulators, responding to peripheral signals such as food intake. Understanding the molecular basis of the effects of food on cognition will help us to determine how best to manipulate diet in order to increase the resistance of neurons to insults and promote mental fitness.
Although food has classically been perceived as a means to provide energy and building material to the body, its ability to prevent and protect against diseases is starting to be recognized. In particular, research over the past 5 years has provided exciting evidence for the influence of dietary factors on specific molecular systems and mechanisms that maintain mental function. For instance, a diet that is rich in omega-3 fatty acids is garnering appreciation for supporting cognitive processes in humans1 and upregulating genes that are important for maintaining synaptic function and plasticity in rodents2. In turn, diets that are high in saturated fat are becoming notorious for reducing molecular substrates that support cognitive processing and increasing the risk of neurological dysfunction in both humans3 and animals4. Although these studies emphasize an important effect of food on the brain, further work is necessary to determine the mechanisms of action and the conditions for therapeutic applications in humans.
Over thousands of years, diet, in conjunction with other aspects of daily living, such as exercise, has had a crucial role in shaping cognitive capacity and brain evolution (BOX 1). Advances in molecular biology have revealed the ability of food-derived signals to influence energy metabolism and synaptic plasticity and, thus, mediate the effects of food on cognitive function, which is likely to have been crucial for the evolution of the modern brain. Feeding habits have been intrinsically associated with the development of human civilization, as people’s choice of what to eat is influenced by culture, religion and society. The newly discovered effects of food on cognition are intriguing for the general public, as they might challenge preconceptions, and they attract substantial interest from the media. The fact that feeding is an intrinsic human routine emphasizes the power of dietary factors to modulate mental health not only at the individual level, but also at the collective, population-wide level. Here I discuss the effects of both internal signals that are associated with feeding and dietary factors on cell metabolism, synaptic plasticity and mental function (FIG. 1). Throughout I use the term cognition from a neurobiological perspective, to refer to the mental processes that are involved in acquiring knowledge and to the integration of these processes into the conscious aspect of emotions, which influences mood and has psychiatric manifestations5.
Box 1. Feeding as an adaptive mechanism for the development of cognitive skills.
Adaptations that facilitated food acquisition and energy efficiency exerted strong evolutionary pressures on the formation of the modern brain and the energy-demanding development of cognitive skills. For example, the wildebeest annually travels hundreds of miles to find feeding grounds in the savannah, a behaviour that requires fully operational and complex navigational, defensive and cognitive conducts for survival. The function of brain centres that control eating behaviour is integrated with those of centres that control cognition (FIG. 1). For instance, animals that eat a potentially poisonous meal develop a perpetual aversion to its flavour through complex mechanisms of learning and memory that involve the hypothalamus, the hippocampus and the amygdala133. In turn, pleasant memories of foods have been related to brain pathways that are associated with reward134.
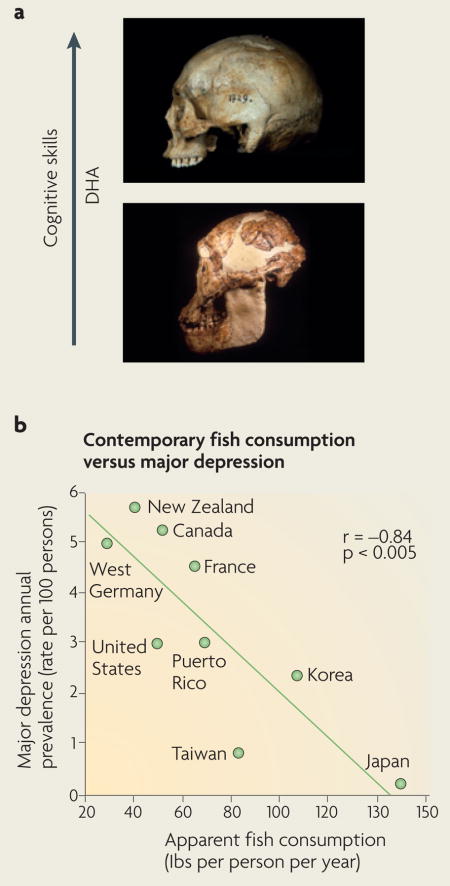
Abundant paleontological evidence suggests that there is a direct relationship between access to food and brain size, and that even small differences in diet can have large effects on survival and reproductive success135. Larger brains in humanoids are associated with the development of cooking skills, access to food, energy savings and upright walking and running136; all of these features require coordination with cognitive strategies that are centred in successful feeding. Dietary consumption of omega-3 fatty acids is one of the best-studied interactions between food and brain evolution. Docosahexaenoic acid (DHA) is the most abundant omega-3 fatty acid in cell membranes in the brain137; however, the human body is not efficient at synthesizing DHA, so we are largely dependent on dietary DHA138. It has been proposed that access to DHA during hominid evolution had a key role in increasing the brain/body-mass ratio (also known as encephalization)138 (see figure, part a). The fact that DHA is an important brain constituent supports the hypothesis that a shore-based diet high in DHA was indispensable for hominid encephalization. Indeed, archeological evidence shows that early hominids adapted to consuming fish and thus gained access to DHA before extensive encephalization occurred. The interplay between brain and environment is ongoing. Over the past 100 years, the intake of saturated fatty acids, linoleic acid and trans fatty acids has increased dramatically in Western civilizations, whereas the consumption of omega-3 fatty acids has decreased. This might explain the elevated incidence of major depression in countries such as the United States and Germany (see figure, part b)78. Both photographs in part a ©Jeffrey H. Schwartz. Part b of the figure reproduced, with permission, from REF. 78 © (1998) Lancet Publishing Group.
Figure 1. Effects of feeding on cognition.

Neural circuits that are involved in feeding behaviour show precise coordination with brain centres that modulate energy homeostasis and cognitive function. The effects of food on cognition and emotions can start before the act of feeding itself, as the recollection of foods through olfactory and visual sensory inputs alters the emotional status of the brain. The ingestion of foods triggers the release of hormones or peptides, such as insulin and glucagon-like peptide 1 (GLP1)31, into the circulation (see REF. 31 for a review); these substances can then reach centres such as the hypothalamus and the hippocampus and activate signal-transduction pathways that promote synaptic activity and contribute to learning and memory. In turn, the lack of food that is signalled by an empty stomach can elicit the release of ghrelin, which can also support synaptic plasticity and cognitive function. Chemical messages derived from adipose tissue through leptin can activate specific receptors in the hippocampus and the hypothalamus, and influence learning and memory. The positive actions of leptin on hippocampus-dependent synaptic plasticity — that is, its actions on NMDA (N-methyl-_-aspartate) receptor function and long-term potentiation facilitation — are well characterized (see REF. 13 for a review). Insulin-like growth factor 1 (IGF1) is produced by the liver and by skeletal muscle in response to signals derived from metabolism and exercise. IGF1 can signal to neurons in the hypothalamus and the hippocampus, with resulting effects on learning and memory performance. In addition to regulating appetite, the hypothalamus coordinates activity in the gut and integrates visceral function with limbic-system structures such as the hippocampus, the amyg-dala and the cerebral cortex. Visceral signals can also modulate cognition and body physiology through the hypothalamic–pituitary axis (HPA). The effects of the hypothalamus can also involve the immune system, as it heavily innervates the thymus and several immune-system molecules can affect synaptic plasticity and cognition. The parasympathetic innervation of the gut by the vagus nerve provides sensory information to the brain, enabling gut activity to influence emotions. In turn, emotions can also influence the viscera through parasympathetic efferents in the vagus nerve. Vagal nerve stimulation is being used therapeutically to treat chronic depression.
Internal signals and cognition
The influence of visceral signals on mental function has been appreciated since ancient times, and to this day lifestyle factors, such as diet and exercise, are used as part of therapies to reduce depression, schizophrenia and bipolar disorders. In this section I discuss the influence of vagal nerve stimulation (VNS) and gut hormones on cognition and emotion (FIG. 1).
Effects of vagal nerve stimulation on cognition
Vagal afferents from the gastrointestinal tract are critical for monitoring various aspects of digestion, such as the release of enzymes and food absorption. The use of VNS has become a routinely approved procedure for the treatment of refractory partial-onset seizures. Based on observations that the application of VNS to patients with epilepsy was associated with improved mood, VNS was perceived as a potential treatment for depression. In humans, VNS failed to produce improvements in depression patients who participated in a short-term open trial (lasting 10 weeks)6; however, in a longer-term study (lasting 12 months), VNS produced beneficial effects that were sustained after 2 years7. Specifically, patients treated with VNS doubled their improvement per month in the Inventory of Depressive Symptoms self report relative to patients receiving treatment as usual (TAU) by itself. TAU consisted of managing treatment-resistant depression with medication or with another therapy that was deemed appropriate by the treating physician. Based on the results of the long-term studies, the US Food and Drug Administration recently approved the use of VNS for the treatment of chronic (not acute) resistant depression (see REF. 8 for a review). Although the mechanisms that underlie the effects of VNS on depression are not well-understood, a recent study demonstrated that VNS increases the levels of the mRNAs for brain-derived neurotrophic factor (BDNF) and fibroblast growth factor 2 (FGF2) in the rat hippocampus and cerebral cortex, as well as the level of noradrenaline in the prefrontal cortex9. As elevations of BDNF10 and noradrenaline have been associated with the effects of antidepressant treatments, these findings provide insights into how signals derived from the gut can affect mood. Furthermore, on the basis that neurons of the dorsal motor nucleus of the vagus nerve retrogradely transport BDNF and other neurotrophins11, it is likely that neurotrophins are involved in sensory and motor signalling from the viscera. Interestingly, a separate line of investigations indicated that the application of VNS to humans12 or rodents12 enhanced memory performance, suggesting that the information that is signalled to the brain by the vagus nerve might serve to influence higher-order cognitive processing.
Gut hormones associated with cognition
In addition to the capacity of the gut to directly stimulate molecular systems that are associated with synaptic plasticity and learning, several gut hormones or peptides, such as leptin, ghrelin, glucagon-like peptide 1 (GLP1) and insulin have been found to influence emotions and cognitive processes (FIG. 1).
Leptin is synthesized in adipose tissue and sends signals to the brain to reduce appetite (see REF. 13 for a review). Leptin receptors have been identified in several brain areas, including the hypothalamus, the cerebral cortex and the hippocampus. The fact that leptin elevates BDNF expression in the hypothalamus suggests that BDNF might mediate the effects of leptin on food intake and energy homeostasis14. Like BDNF, leptin facilitates synaptic plasticity in the hippocampus15. Genetically obese rodents with dysfunctional leptin receptors show impairments in long-term potentiation (LTP) and long-term depression and difficulties in spatial learning16. These effects were rescued by administrating leptin into the hippocampus15,17. New studies showing that leptin promotes rapid changes in hippocampal dendritic morphology suggest that leptin exerts a direct action on hippocampal plasticity18.
Ghrelin is an adipogenic hormone that is secreted by an empty stomach (see REF. 19 for a review); it acts as an appetite stimulant in mice20 and humans21. Ghrelin is the endogenous ligand of the growth hormone secretagogue receptor, which is expressed in the arcuate nucleus in the hypothalamus22 and in the hippocampus23. Peripheral administration of ghrelin increases food intake in normal rodents24,25 and humans26,27, whereas chronic administration can lead to adiposity24,25. Ghrelin also promotes rapid reorganization of synaptic terminals in the hypothalamus28, and in the hippocampus it promotes synapse formation in dendritic spines and LTP, which are paralleled by enhanced spatial learning and memory formation29.
GLP1, which is synthesized by intestinal cells, regulates energy metabolism by stimulating pancreatic insulin secretion and subsequent glucose uptake by muscle cells, and by suppressing food intake through actions on the hypothalamus. GLP1 receptors are expressed in neurons, and infusion of GLP1 into the brain has been shown to improve associative and spatial memory in rats30. Owing to their multiple actions on somatic and neural targets, ghrelin, leptin and GLP1 can integrate processes that influence cognition and emotion.
Finally, insulin, which has classically been regarded as a gut hormone that is produced in the pancreas, has also been found to alter synaptic activity and cognitive processing (see REF. 31 for a review). Insulin secretion is normally stimulated by the mental anticipation to meals and continues during digestion and the absorption of foods into the bloodstream. Insulin can enter the brain and interact with specific signal-transduction receptors located in discrete brain regions, such as the hippocampus. Overall, the evidence seems to indicate that the act of feeding can itself modulate cognitive processes on two levels, through neural circuits that connect the gut and the brain and through the release of gut peptides into the bloodstream (FIG. 1).
Thus, as predicted from an evolutionary perspective, the gut does influence the molecular mechanisms that determine the capacity for acquiring new memories and that control emotions, as well as overall mental function. It is not surprising that visceral signals are now recognized as essential factors for the treatment of psychiatric disorders. The challenge now is to better our understanding of the molecular mechanisms by which peripheral signals can modulate mental processes.
From energy metabolism to cognition
The brain consumes an immense amount of energy relative to the rest of the body. Thus, the mechanisms that are involved in the transfer of energy from foods to neurons are likely to be fundamental to the control of brain function. Processes that are associated with the management of energy in neurons can affect synaptic plasticity32 (FIG. 2), which could explain how metabolic disorders can affect cognitive processes. Interestingly, synaptic function can, in turn, alter metabolic energy, allowing mental processes to influence somatic function at the molecular level. BDNF is an excellent example of a signalling molecule that is intimately related to both energy metabolism and synaptic plasticity: it can engage metabolic signals to affect cognitive function32. BDNF is most abundant in brain areas that are associated with cognitive and metabolic regulation: the hippocampus and the hypothalamus, respectively33. Learning to carry out a task increases BDNF-mediated synaptic plasticity in the hippocampus34,35, and genetic deletion of the BDNF gene impairs memory formation36,37. The Met variant of the Val66Met BDNF polymorphism, a common genotype in humans that is related to abnormal trafficking and secretion of BDNF in neuronal cells38, is associated with abnormal hippocampal functioning and memory processing39. In turn, BDNF has also been shown to influence multiple parameters of energy metabolism, such as appetite suppression40,41, insulin sensitivity 42,43 and glucose44 and lipid metabolism45. In addition, the hypothalamic melanocortin 4 receptor, which is crucial for the control of energy balance, regulates the expression of BDNF in the ventral medial hypothalamus46, supporting an association between energy metabolism and synaptic plasticity. In rodents, a reduction in energy metabolism caused by infusing a high dose of vitamin D3 into the brain has been shown to abolish the effects of exercise on downstream effectors of BDNF-mediated synaptic plasticity, such as calcium/calmod-ulin-dependent protein kinase II (CaMKII), synapsin I and cyclic AMP-responsive element (CRE)-binding protein (CREB)32. In humans, a de novo mutation in TrkB, a BDNF receptor, has been linked with hyperphagic obesity, as well as impairments in learning and memory 47. Although energy metabolism and BDNF-mediated synaptic plasticity seem to be interconnected, further studies are crucial to determine the confines of this relationship for the modulation of cognitive function.
Figure 2. Energy homeostasis and cognition.

Diet and exercise can affect mitochondrial energy production, which is important for maintaining neuronal excitability and synaptic function. The combination of certain diets and exercise can have additive effects on synaptic plasticity and cognitive function. ATP produced by mitochondria might activate brain-derived neurotrophic factor (BDNF) and insulin-like growth factor 1 (IGF1), which support synaptic plasticity and cognitive function. Energy-balancing molecules, such as ubiquitous mitochondrial creatine kinase (uMtCK), AMP-activated protein kinase (AMPK) and uncoupling protein 2 (UCP2)141,146, interact with BDNF to modulate synaptic plasticity and cognition. Excess energy production caused by high caloric intake or strenuous exercise results in the formation of reactive oxygen species (ROS). When ROS levels exceed the buffering capacity of the cell, synaptic plasticity and cognitive function are compromised, probably owing to a reduction in the actions of signal-transduction modulators such as BDNF. Energy metabolism can also affect molecules such as silent information regulator 1 (SIRT1), a histone deacetylase that contributes to the reduction of ROS and promotes chromatin modifications that underlie epigenetic alterations that might affect cognition146. On the basis of its demonstrated susceptibility for epigenetic modification73, another potential target for the effects of diet on epigenetics is the BDNF gene. Two main findings support a mechanism whereby exercise, similar to diet, enhances cognitive processes through effects on energy metabolism and synaptic plasticity. First, the combination of exercise and certain diets elevates the expression of uMtCK, AMPK and UCP2, which might affect energy homeostasis and brain plasticity. Second, disruption of energy homeostasis during voluntary wheel-running abolished the effects of exercise on the actions of BDNF and BDNF end products that are important for learning and memory, suggesting that energy metabolism influences BDNF function147.
The mechanism whereby BDNF affects metabolism and synaptic plasticity seems to involve insulin-like growth factor 1 (IGF1)48. IGF1 is synthesized in the liver, in skeletal muscle and throughout the brain, whereas brain IGF1 receptors are expressed mainly in the hippocampus49. A reduction of IGF1 signalling in rodents results in hyperglycaemia and insulin resistance, and infusion of IGF1 into the brain decreases plasma insulin levels and increases insulin sensitivity50. IGF1 also supports nerve growth and differentiation, neurotransmitter synthesis and release51 and synaptic plasticity52, and might contribute to sustaining cognitive function after brain insults53,54, diabetes55 and aging56. IGF1 has been shown in rodents to entrain similar downstream pathways to BDNF, such as the Akt signalling system57. Interestingly, the omega-3 fatty acid docosahexaenoic acid (DHA) stimulates neuronal plasticity through the Akt pathway58, suggesting that Akt activation might be crucial for integrating the effects of food-derived signals on brain plasticity. The phosphatidyl-inositol 3-kinase (PI3K)/Akt/mammalian target of rapamycin (mTOR) signalling pathway seems to integrate the effects of BDNF and IGF1 on energy metabolism, synaptic plasticity, and learning and memory (FIG. 3).
Figure 3. Dietary omega-3 fatty acids can affect synaptic plasticity and cognition.

The omega-3 fatty acid docosahexaenoic acid (DHA), which humans mostly attain from dietary fish, can affect synaptic function and cognitive abilities by providing plasma membrane fluidity at synaptic regions. DHA constitutes more than 30% of the total phospholipid composition of plasma membranes in the brain, and thus it is crucial for maintaining membrane integrity and, consequently, neuronal excitability and synaptic function. Dietary DHA is indispensable for maintaining membrane ionic permeability and the function of transmembrane receptors that support synaptic transmission and cognitive abilities. Omega-3 fatty acids also activate energy-generating metabolic pathways that subsequently affect molecules such as brain-derived neurotrophic factor (BDNF) and insulin-like growth factor 1 (IGF1). IGF1 can be produced in the liver and in skeletal muscle, as well as in the brain, and so it can convey peripheral messages to the brain in the context of diet and exercise. BDNF and IGF1 acting at presynaptic and postsynaptic receptors can activate signalling systems, such as the mitogen-activated protein kinase (MAPK) and calcium/calmodulin-dependent protein kinase II (CaMKII) systems, which facilitate synaptic transmission and support long-term potentiation that is associated with learning and memory. BDNF has also been shown to be involved in modulating synaptic plasticity and cognitive function through the phosphatidylinositol 3-kinase (PI3K)/Akt/mammalian target of rapamycin (mTOR) signalling pathway. The activities of the mTOR and Akt signalling pathways are also modulated by metabolic signals such as insulin and leptin (not shown). 4EBP, eukaryotic translation-initiation factor 4E binding protein; CREB, cyclic AMP-responsive element (CRE)-binding protein; IGFR, insulin-like growth factor receptor; IRS1, insulin receptor substrate 1; p70S6K, p70 S6 kinase.
Disturbances in energy homeostasis have been linked to the pathobiology of several mental diseases, and so dietary management is becoming a realistic strategy to treat psychiatric disorders. Numerous studies have found that there might be an association between abnormal metabolism (diabetes type II, obesity and metabolic syndrome) and psychiatric disorders59. In a large study of patients with manic depression60 or schizophrenia61,62, the rate of diabetes was found to be higher than in the general population (1.2% of people aged 18–44 years and 6.3% of people aged 45–64 years163). The overall prevalence of diabetes in a group of 95 patients with schizophrenia was 15.8%, and this increased to 18.9% with age61, whereas diabetes in 203 patients with manic depression ranged from 2.9% in patients of approximately 30 years of age to 25% in patients of 75–79 years of age60. However, it is difficult to ascertain a cause–effect relationship between diabetes and psychiatric disorders in these studies given that schizophrenia, manic depression and other psychiatric disorders are associated with poor quality of life and the side effects of anti-psychotic medication. On the basis of its effects on synaptic plasticity and energy metabolism, BDNF has been the focus of research into current hypotheses of schizophrenia and depression63–66. Low levels of BDNF in the plasma are associated with impaired glucose metabolism and type II diabetes67, and BDNF is reduced in the hippocampus, in various cortical areas68 and in the serum69 of patients with schizophrenia. In mice, genetic deletion of the TrkB receptor in the forebrain produces schizophrenic-like behaviour70. Furthermore, BDNF levels are reduced in the plasma of patients with major depression71, and chronic administration of antidepressants elevates hippocampal BDNF levels72. A recent study in rodents demonstrated that defeat stress, an animal model of depression, induced a lasting repression of BDNF transcripts, whereas antidepressant treatment reversed this repression by inducing histone acetylation73. Although the evidence is not conclusive to argue that BDNF has a role in mediating depression or schizophrenia, it is becoming clear that most treatments for depression or schizophrenia — that is, exercise and drugs — involve the action of BDNF.
Effects of nutrients on cognition
Several dietary components have been identified as having effects on cognitive abilities (TABLE 1). Dietary factors can affect multiple brain processes by regulating neurotransmitter pathways, synaptic transmission, membrane fluidity and signal-transduction pathways. This section focuses on recent evidence that shows the capacity of nutrients to affect neural pathways that are associated with synaptic plasticity.
Table 1.
Select nutrients that affect cognitive function
| nutrient | effects on cognition and emotion | food sources |
|---|---|---|
| Omega-3 fatty acids (for example, docosahexaenoic acid) | Amelioration of cognitive decline in the elderly148; basis for treatment in patients with mood disorders80; improvement of cognition in traumatic brain injury in rodents81; amelioration of cognitive decay in mouse model of Alzheimer’s disease149,150 | Fish (salmon), flax seeds, krill, chia, kiwi fruit, butternuts, walnuts |
| Curcumin | Amelioration of cognitive decay in mouse model of Alzheimer’s disease123; amelioration of cognitive decay in traumatic brain injury in rodents89 | Turmeric (curry spice) |
| Flavonoids | Cognitive enhancement in combination with exercise in rodents92; improvement of cognitive function in the elderly151 | Cocoa, green tea, Ginkgo tree, citrus fruits, wine (higher in red wine), dark chocolate |
| Saturated fat | Promotion of cognitive decline in adult rodents4; aggravation of cognitive impairment after brain trauma in rodents88; exacerbation of cognitive decline in aging humans3 | Butter, ghee, suet, lard, coconut oil, cottonseed oil, palm kernel oil, dairy products (cream, cheese), meat |
| B vitamins | Supplementation with vitamin B6, vitamin B12 or folate has positive effects on memory performance in women of various ages152; vitamin B12 improves cognitive impairment in rats fed a choline-deficient diet153 | Various natural sources. Vitamin B12 is not available from plant products |
| Vitamin D | Important for preserving cognition in the elderly154 | Fish liver, fatty fish, mushrooms, fortified products, milk, soy milk, cereal grains |
| Vitamin E | Amelioration of cognitive impairment after brain trauma in rodents102; reduces cognitive decay in the elderly119 | Asparagus, avocado, nuts, peanuts, olives, red palm oil, seeds, spinach, vegetable oils, wheatgerm |
| Choline | Reduction of seizure-induced memory impairment in rodents155; a review of the literature reveals evidence for a causal relationship between dietary choline and cognition in humans and rats156 | Egg yolks, soy beef, chicken, veal, turkey liver, lettuce |
| Combination of vitamins (C, E, carotene) | Antioxidant vitamin intake delays cognitive decline in the elderly157 | Vitamin C: citrus fruits, several plants and vegetables, calf and beef liver. Vitamin E: see above |
| Calcium, zinc, selenium | High serum calcium is associated with faster cognitive decline in the elderly158; reduction of zinc in diet helps to reduce cognitive decay in the elderly159; lifelong low selenium level associated with lower cognitive function in humans160 | Calcium: milk, coral. Zinc: oysters, a small amount in beans, nuts, almonds, whole grains, sunflower seeds. Selenium: nuts, cereals, meat, fish, eggs |
| Copper | Cognitive decline in patients with Alzheimer’s disease correlates with low plasma concentrations of copper161 | Oysters, beef/lamb liver, Brazil nuts, blackstrap molasses, cocoa, black pepper |
| Iron | Iron treatment normalizes cognitive function in young women162 | Red meat, fish, poultry, lentils, beans |
Dietary lipids, which were originally thought to affect the brain through their effects on cardiovascular physiology, are garnering recognition for their direct actions on the brain. Omega-3 polyunsaturated fatty acids are normal constituents of cell membranes and are essential for normal brain function (FIG. 3). In spite of the large variability in the design of experiments to evaluate the action of different dietary elements on cognitive abilities, there is a general consensus that a deficiency of omega-3 fatty acids in rodents results in impaired learning and memory74,75. Dietary deficiency of omega-3 fatty acids in humans has been associated with increased risk of several mental disorders, including attention-deficit disorder, dyslexia, dementia, depression, bipolar disorder and schizophrenia76–80. As the omega-3 fatty acid DHA is a prominent component of neuronal membranes, and as the human body is inefficient in synthesizing DHA, we are reliant on dietary DHA. Some of the mechanisms by which DHA affects brain plasticity and cognition are starting to be elucidated. For example, DHA dietary supplementation has been found to elevate levels of hippocampal BDNF and enhance cognitive function in rodent models of brain trauma81. DHA might enhance cognitive abilities by facilitating synaptic plasticity and/or enhancing synaptic membrane fluidity; it might also act through its effects on metabolism, as DHA stimulates glucose utilization82 and mitochondrial function83, reducing oxidative stress (OS)81.
Most of the studies in humans have been directed at evaluating the effects of omega-3 fatty acids on reducing the cognitive deficit that is associated with psychiatric disorders. Several other, widely publicized, attempts to determine the effects of omega-3 fatty acid supplementation on the performance of school children have been carried out. A randomized double-blind controlled trial in which half of the children received omega-3 fatty acids and the other half received placebos is being conducted across several schools in Durham, UK84. Previous studies from the same investigators showed that omega-3 fatty acid supplementation was associated with reduced cognitive deficits (in reading and spelling, and teaching-rated behaviour) in children affected with developmental coordination disorder — that is, in children with specific impairments of motor function that are independent of their motor ability 85. In the new studies, children were selected on the basis that “they were not fulfilling their potential at school” but their general ability was deemed “normal”, and they were subjected to regular tests to measure their coordination, concentration and academic ability. According to preliminary results84, some level of improvement in school performance was observed in the group receiving omega-3 fatty acids, unleashing a flurry of speculations from the media. Although the results of the Durham study require scientific scrutiny for validation, they seem to agree with those of another study86 in which omega-3 fatty acids (DHA 88 mg per day and eicosapentaenoic acid (EPA) 22 mg per day) and micro-nutrients (iron, zinc, folate and vitamins A, B6, B12 and C) were provided in a drink mix to 396 children (6–12 years of age) in Australia and 394 children in Indonesia. The results showed higher scores on tests that measured verbal intelligence and learning and memory after 6 and 12 months in both boys and girls in Australia, but in only girls in Indonesia. Although these results are consistent with described roles of omega-3 fatty acids during brain development and cognition87, it is plausible that the other dietary supplements that were present in the cocktail could have contributed to the behavioural effects. This would suggest that select dietary components might act in an additive fashion.
In contrast to the healthy effects of diets that are rich in omega-3 fatty acids, epidemiological studies indicate that diets with high contents of trans and saturated fats adversely affect cognition3. Rodent studies that evaluated the effects of “junk food”, characterized by high contents of saturated fat and sucrose, have shown a decline in cognitive performance and reduced hippocampal levels of BDNF-related synaptic plasticity after only 3 weeks of dietary treatment4. These findings suggest that the diet had a direct effect on neurons that was independent of insulin resistance or obesity. More alarming is the fact that this diet elevated the neurological burden that was associated with experimental brain injury, as evidenced by worse performance in learning tasks and a reduction of BDNF-mediated synaptic plasticity 88. Evidence that the antioxidants curcumin and vitamin E counteracted the effects of the diet suggests that increased OS might mediate the effects of the diet on plasticity 89,90.
Flavonols are part of the flavonoid family that is found in various fruits, cocoa, beans and the Ginkgo biloba tree. Although the antioxidant effects of flavonols are well established in vitro, there is general agreement that flavonols have more complex actions in vivo that require further investigation. The flavonol quercetin, a major component of G. biloba extracts, has been shown to reduce learning and memory impairment in cerebral ischaemic rodents91. Dietary supplementation with the plant-derived flavanol (−) epicathechin, which has been shown to cross the blood–brain barrier, elevated indices of synaptic spine density and angiogenesis and increased hippocampus-dependent memory in mice92. More interestingly, the positive effects of (−) epicathechin dietary supplementation on memory formation in this study were found to be further enhanced by concomitant exercise (see BOX 2).
Box 2. Additive effects of diet and exercise on synaptic plasticity and cognition.
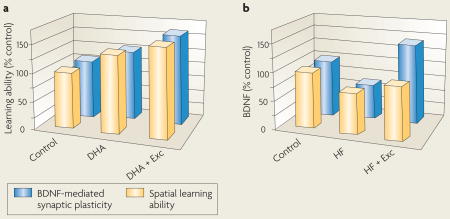
Recent studies have shown a cooperative action of diet and exercise at the molecular level, which could influence cognitive abilities. In addition to its capacity to benefit overall health, numerous studies have shown that exercise enhances learning and memory under a variety of conditions (for reviews see REFS 139, 140). In humans, exercise has been shown to counteract the mental decline that is associated with aging141, enhance the mental capacity of young adults142 and facilitate functional recovery after brain injury or disease131. Studies that showed that exercise promotes neurogenesis in the brain of adult rodents143 and humans144 have introduced the possibility that new proliferating neurons might contribute to the effects of exercise on enhancing learning and memory. In rodents, exercise (Exc) and docosahexaenoic acid (DHA) dietary supplementation combined (DHA+Exc) had a greater effect on brain-derived neurotrophic factor (BDNF)-mediated synaptic plasticity (see figure, part a, blue bars) and cognition (spatial learning ability, yellow bars) than either factor alone132, highlighting the potential of this approach for treating brain injuries. Similarly, the combination of a flavonoid-enriched diet and exercise increased the expression of genes that have a positive effect on neuronal plasticity and decreased the expression of genes that are involved in deleterious processes, such as inflammation and cell death92. Exercise has also been proven to be effective at reducing the deleterious effects of unhealthy diets, such as those that are high in saturated fat and sucrose (HF) (see figure, part b)4. Molecules that could explain the synergistic effects of diet and exercise include BDNF, which has emerged as an important factor for translating the effects of exercise on synaptic plasticity and cognitive function132,145, and several molecules that are associated with the action of BDNF on synaptic function, such as synapsin I, calcium/calmodulin-dependent protein kinase II (CaMKII) and cyclic AMP-responsive element (CRE)-binding protein (CREB). A comprehensive evaluation of how diet interacts with other lifestyle factors is important for determining the best way to enhance brain function and mental health.
Folate or folic acid is found in various foods, including spinach, orange juice and yeast. The liver generates several forms of folate after the intestine has absorbed vitamin B. Folate deficiency, which is mostly caused by low dietary intake, has been associated with a number of physiological abnormalities during development and adulthood93. Adequate levels of folate are essential for brain function, and folate deficiency can lead to neurological disorders, such as depression94 and cognitive impairment. Folate supplementation either by itself 95,96 or in conjunction with other B vitamins97,98 has been shown to be effective at preventing cognitive decline and dementia during aging, and at potentiating the effects of antidepressants99. The results of a recent randomized clinical trial indicated that a 3-year folic acid supplementation can help to reduce the age-related decline in cognitive function100. These studies, however, have sparked further debate in the scientific community that age, vitamin B12 status, genetic makeup, the presence of existing medical conditions and the current drug programme of patients receiving folic acid are important factors to be taken into consideration to reduce undesirable secondary effects, such as anaemia, low immune function and cancer101. The effects of other nutrients on cognition are summarized in TABLE 1.
Caloric intake and cognition
Caloric restriction
Altering the caloric content of the diet is a potential means by which to affect cognitive capacity. New research indicates that metabolic processes that are initiated by the burning of fuels in mitochondria can modulate select aspects of synaptic plasticity and hence have the potential to affect cognitive function (FIG. 2). Certain mechanisms that regulate cell metabolism are integrated with mechanisms that modulate synaptic function. For example, excess calories can reduce synaptic plasticity32,102 and increase the vulnerability of cells to damage103 by causing free-radical formation that surpasses the buffering capacity of the cell. Moderate caloric restriction could thus protect the brain by reducing oxidative damage to cellular proteins, lipids and nucleic acids104. Studies in rodents indicate that elevated OS decreases BDNF-mediated synaptic plasticity and cognitive function32,102. Caloric restriction also elevates levels of BDNF105,106, suggesting that BDNF might mediate the effects of low caloric intake on synaptic plasticity. Reducing caloric intake to approximately 40% of control nominal values in mice from weaning to 35 months of age decreases the deficits in motor and cognitive function that are associated with aging107. Alternate-day feeding ameliorates age-related deficits in cognitive function in a mouse model of Alzheimer’s disease when the feeding programme is maintained between 3 and 17 months of age108.
According to the ‘thrifty-gene’ hypothesis, our genome has adapted through thousands of years of evolution to profit from nominal amounts of calories in order to cope with limited food resources109. A standing concern in the field has been how caloric intake or meal frequency affects energy metabolism and health in humans. Recent studies in middle-aged men and women have established that alterations in meal frequency, without a reduction in energy intake, result in unchanged levels of several metabolic parameters, such as glucose, insulin, leptin and BDNF110. However, another study in which subjects were maintained on an alternate-day caloric-restriction diet over a 2-month period resulted in weight loss and improved cardiovascular-disease and diabetes-risk profiles111. The apparent discrepancy between these two studies suggests that the number of calories seems to be a crucial factor for the physiological effects, such that controlled meal skipping or intermittent caloric restriction might have health benefits in humans. However, further preclinical information is required for the design of therapeutic applications, so caution should be exerted in the interpretation of these studies to avoid misconceptions such as the belief that a low-calorie diet might be sufficient to promote health. This view disregards the fact that the nutritional balance of the diet is a vital requirement for the potential health benefits of low-calorie diets. It will be of considerable interest to determine how these dietary manipulations can affect other physiological parameters, such as hormonal profiles and immune-system status, which are crucial for assessing the benefits of restricted caloric intake for therapeutic purposes.
Antioxidant foods
The brain is highly susceptible to oxidative damage because of its high metabolic load and its abundance of oxidizable material, such as the poly-unsaturated fatty acids that form the plasma membranes of neural cells. Several ‘anti-oxidant diets’ have become popular for their publicized positive effects on neural function. Berries, for example, have been shown to have strong antioxidant capacity, but only a limited number of their many components have been evaluated separately: two tannins (procyanidin and prodelphinidin), anthocyanins and phenolics (see REF. 112 for a review). In rats, polyphenols have been shown to increase hippocampal plasticity (as measured by increases in HSP70 (REF. 113) and IGF1 (REF. 114)), to provide protection against kainate-induced damage115 and to benefit learning and memory performance114. It is not clear how berry extracts can benefit plasticity and cognition, but their effects are probably associated with their ability to maintain metabolic homeostasis, as this would protect membranes from lipid peroxidation and affect synaptic plasticity.
Various micronutrients with an anti-oxidant capacity that has been associated with mitochodrial activity have been shown to influence cognitive function. Alpha lipoic acid, which is found in meats such as kidney, heart and liver, and vegetables such as spinach, broccoli and potatoes, is a coenzyme that is important for maintaining energy homeostasis in mitochondria116. Alpha lipoic acid has been shown to improve memory deficits in animal models of Alzheimer’s disease117 and to reduce cognitive decay in a small group of patients with Alzheimer’s disease118. Vitamin E, or α-tocopherol, has also been implicated in cognitive performance, as decreasing serum levels of vitamin E were associated with poor memory performance in older individuals119. Vitamin E is abundant in vegetable oils, nuts, green leafy vegetables and fortified cereals, and has been shown to extend lifespan and improve mitochondrial function and neurological performance in aging mice120. The mechanisms by which vitamin E can affect cognition are not well-understood, but they are likely to be related to the putative capacity of antioxidants to support synaptic plasticity102 by protecting synaptic membranes from oxidation. Finally, the curry spice curcumin, a traditional food preservative and medicinal herb in India121,122, has been shown to reduce memory deficits in animal models of Alzheimer’s disease123 and brain trauma89. Curcumin is relatively non-toxic and has few side effects at doses greater than the low dose that has been tested in mice122. Given the high consumption of curcumin in India, it is possible that it might contribute to the low prevalence of Alzheimer’s disease in that country124. Curcumin is a strong antioxidant that seems to protect the brain from lipid peroxidation125 and nitric-oxide-based radicals126.
Diet and epigenetics
A number of innovative studies are pointing to the exciting possibility that the effects of diet on mental health can be transmitted across generations. The results of these studies indicate the importance of dietary components in influencing epigenetic events — that is, non-genetic events, such as DNA methylation, transcriptional activation, translational control and post-translational modifications that cause a potentially heritable phenotypic change — and, thus, their potential for disease modulation. The results of a longitudinal study that included more than 100 years of birth, death, health and genealogical records of 300 Swedish families in an isolated village showed that an individual’s risk for diabetes and early death was increased if their paternal grandparents grew up in times of food abundance rather than times of food shortage164. Although the molecular mechanisms for the influence of diet on epigenetics are unknown, it is known that the BDNF system is particularly susceptible to epigenetic modifications that influence cognitive function127. Chromatin modifications at specific BDNF promoters determine the differential expression of discrete BDNF splice variants. Such modifications have been observed in Alzheimer’s disease128 and can also be elicited by particular antidepressant drugs73. Accordingly, it is likely that the various BDNF splice variants have differential effects on neuronal plasticity and cognition (see REF. 65 for a review). Neural activity dissociates methyl-CpG-binding protein 2 (MECP2) from its latent location at BDNF promoter III, enabling transcription of BDNF129. A recent study in a rodent model of depression demonstrated that depressive manifestations and subsequent antidepressant treatment are associated with sustained changes in histone acetylation and methylation at BDNF promoter III73.
These studies represent a starting point for understanding how intracellular signalling that is triggered by lifestyle factors can promote lasting changes in DNA function in the brain and in cognitive capacity. Silent information regulator 2 (SIRT2), a member of the sirtuin protein family, has emerged as an important modulator of genomic stability and cellular homeostasis that seems to act by silencing the function of specific genes. A diet that is high in saturated fat reduces the expression of SIRT2 in the rat hippocampus90, whereas a diet that is high in omega-3 fatty acids has the opposite effect2. Although the mechanisms that are involved in the regulation of SIRT2 by dietary factors require further investigation, the fact that energy metabolism is involved in the modulation of SIRT2 (as with BDNF) can provide a link for the influence of dietary factors on long-term genomic stability. Interestingly, a recent study in humans examined the association between SIRT1 (homologous to the rat Sirt2 gene) gene polymorphisms and cognition130. In this study, 1,245 inhabitants of Leiden (in the Netherlands) who were at least 85 years old were genotyped for 5 SIRT1 polymorphisms during a period of 4.4 years. Those who were homozygous for one of the polymorphisms that affected the SIRT1 promoter region showed better preservation on all measurements of cognitive function than the others. Findings that SIRT2 protein is present in rodent hippocampal tissue2 and that SIRT2 function is involved in the maintenance of energy homeostasis could provide clues to how SIRT1 might relate to cognitive function.
Conclusions and future directions
Diet, exercise and other aspects of our daily interaction with the environment have the potential to alter our brain health and mental function. We now know that particular nutrients influence cognition by acting on molecular systems or cellular processes that are vital for maintaining cognitive function. This raises the exciting possibility that dietary manipulations are a viable strategy for enhancing cognitive abilities and protecting the brain from damage, promoting repair and counteracting the effects of aging. Emerging research indicates that the effects of diet on the brain are integrated with the actions of other lifestyle modalities, such as exercise (see BOX 2) and sleep131,132. The combined action of particular diets and exercise on the activation of molecular systems that are involved in synaptic plasticity has strong implications for public health and the design of therapeutic interventions. Owing to the encouraging results of clinical and preclinical studies that showed the beneficial effects of foods on the brain, the topic has attracted substantial media attention. Some of the information that has been conveyed has been hazy or exaggerated, and has contributed to people’s apprehension of taking advantage of scientific advances. As discussed, several dietary components have been found to have positive effects on cognition; however, caution is required, as a balanced diet is still the stepping-stone for any dietary supplementation. By the same token, popular dietary prescriptions that might help to reduce weight do not necessarily benefit the physiology of the body or the mind.
Brain networks that are associated with the control of feeding are intimately associated with those that are involved in processing emotions, reward and cognition. A better understanding of how these networks interact will probably produce fundamental information for the development of strategies to reduce food addiction and obesity, a major social and economic burden in Western society. It is encouraging that modern psychiatry has started to appraise the implementation of some of these concepts for the treatment of various mental disorders. For example, a consensus report from the American Psychiatric Association’s Committee on Research on Psychiatric Treatments has provided general guiding principles for the use of omega-3 fatty acids for the treatment of mood disorders80.
The fact that dietary factors and other aspects of lifestyle have an effect on a long-term timescale contributes to an under-estimation of their importance for public health. Accordingly, the slow and imperceptible cognitive decay that characterizes normal aging is within the range-of-action of brain foods, such that successful aging is an achievable goal for dietary therapies. The capacity of diet to modulate cognitive abilities might have even longer-term implications in light of recent studies that imply that nutritional effects might be transmitted over generations by influencing epigenetic events. Research indicating that an excessive intake of calories might negate the positive effects of certain diets suggests that there is an undefined line between abundance of foods and neural health. Ironically, judging by the increasing rate of obesity in Western countries, which affects individual’s health and the economy as a whole, the excessive food intake in these wealthy nations seems to be almost as harmful as the lack of it in poor countries. It is intriguing that several countries with limited resources, such as India, have a reduced prevalence of neurological disorders that have been associated with diet, such as Alzheimer’s disease. This raises the concern of whether industrialized societies are consuming a balanced diet that takes into consideration appropriate numbers of calories as well as appropriate nutrients and adequate levels of exercise. Many practical questions regarding the design of diets to specifically improve brain function, such as type, frequency and amount of nutrients that constitute healthy brain food, remain to be answered, but we are beginning to uncover the basic principles that are involved in the actions of foods on the brain. Incorporating this knowledge into the design of novel treatments could be vital to combating mental diseases and neurological weaknesses.
Acknowledgments
In the elaboration of this article, I greatly benefited from inspirational discussions with G. Cole, I. Cook, R. Edgerton, T. Jones and D. Glanzman. I would also like to thank A. She and S. Kim for valuable editorial assistance. I am thankful to the reviewers for their constructive feedback, and to the National Institute of Neurological Disorder and Stroke (NS50465) for funding support.
Footnotes
DATABASES
Entrez Gene:http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=geneAkt | BDNF | CaMKII | CREB | FGF2 | ghrelin | GLP1 | HSP70 |IGF1 | leptin | MECP2 | mTOR | Sirt 2 | SIRT1 | TrkB
FURTHER INFORMATION
Fernando Gomez-Pinilla's homepages:http://www.physci.ucla.edu/research/GomezPinilla/http://faculty.neuroscience.ucla.edu/institution/personnel?personnel%5fid=45774
All links are active in the online pdf
References
- 1.McCann JC, Ames BN. Is docosahexaenoic acid, an n-3 long-chain polyunsaturated fatty acid, required for development of normal brain function? An overview of evidence from cognitive and behavioral tests in humans and animals. Am J Clin Nutr. 2005;82:281–295. doi: 10.1093/ajcn.82.2.281. [DOI] [PubMed] [Google Scholar]
- 2.Wu A, Ying Z, Gomez-Pinilla F. Omega-3 fatty acids supplementation restores mechanisms that maintain brain homeostasis in traumatic brain injury. J Neurotrauma. 2007;24:1587–1595. doi: 10.1089/neu.2007.0313. [DOI] [PubMed] [Google Scholar]
- 3.Greenwood CE, Winocur G. High-fat diets, insulin resistance and declining cognitive function. Neurobiol Aging. 2005;26 (Suppl 1):42–45. doi: 10.1016/j.neurobiolaging.2005.08.017. [DOI] [PubMed] [Google Scholar]
- 4.Molteni R, Barnard JR, Ying Z, Roberts CK, Gomez-Pinilla F. A high-fat, refined sugar diet reduces hippocampal brain-derived neurotrophic factor, neuronal plasticity, and learning. Neuroscience. 2002;112:803–814. doi: 10.1016/s0306-4522(02)00123-9. [DOI] [PubMed] [Google Scholar]
- 5.Pessoa L. On the relationship between emotion and cognition. Nature Rev Neurosci. 2008;9:148–158. doi: 10.1038/nrn2317. [DOI] [PubMed] [Google Scholar]
- 6.Rush AJ, et al. Vagus nerve stimulation for treatment-resistant depression: a randomized, controlled acute phase trial. Biol Psychiatry. 2005;58:347–354. doi: 10.1016/j.biopsych.2005.05.025. [DOI] [PubMed] [Google Scholar]
- 7.Nahas Z, et al. Two-year outcome of vagus nerve stimulation (VNS) for treatment of major depressive episodes. J Clin Psychiatry. 2005;66:1097–1104. doi: 10.4088/jcp.v66n0902. [DOI] [PubMed] [Google Scholar]
- 8.George MS, et al. Brain stimulation for the treatment of psychiatric disorders. Curr Opin Psychiatry. 2007;20:250–254. doi: 10.1097/YCO.0b013e3280ad4698. [DOI] [PubMed] [Google Scholar]
- 9.Follesa P, et al. Vagus nerve stimulation increases norepinephrine concentration and the gene expression of BDNF and bFGF in the rat brain. Brain Res. 2007;1179:28–34. doi: 10.1016/j.brainres.2007.08.045. [DOI] [PubMed] [Google Scholar]
- 10.Nibuya M, Morinobu S, Duman RS. Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J Neurosci. 1995;15:7539–7547. doi: 10.1523/JNEUROSCI.15-11-07539.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Helke CJ, et al. Axonal transport of neurotrophins by visceral afferent and efferent neurons of the vagus nerve of the rat. J Comp Neurol. 1998;393:102–117. doi: 10.1002/(sici)1096-9861(19980330)393:1<102::aid-cne10>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- 12.Clark KB, Naritoku DK, Smith DC, Browning RA, Jensen RA. Enhanced recognition memory following vagus nerve stimulation in human subjects. Nature Neurosci. 1999;2:94–98. doi: 10.1038/4600. [DOI] [PubMed] [Google Scholar]
- 13.Harvey J. Leptin regulation of neuronal excitability and cognitive function. Curr Opin Pharmacol. 2007;7:643–647. doi: 10.1016/j.coph.2007.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Komori T, Morikawa Y, Nanjo K, Senba E. Induction of brain-derived neurotrophic factor by leptin in the ventromedial hypothalamus. Neuroscience. 2006;139:1107–1115. doi: 10.1016/j.neuroscience.2005.12.066. [DOI] [PubMed] [Google Scholar]
- 15.Shanley LJ, Irving AJ, Harvey J. Leptin enhances NMDA receptor function and modulates hippocampal synaptic plasticity. J Neurosci. 2001;21:RC186. doi: 10.1523/JNEUROSCI.21-24-j0001.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Li XL, et al. Impairment of long-term potentiation and spatial memory in leptin receptor-deficient rodents. Neuroscience. 2002;113:607–615. doi: 10.1016/s0306-4522(02)00162-8. [DOI] [PubMed] [Google Scholar]
- 17.Wayner MJ, Armstrong DL, Phelix CF, Oomura Y. Orexin-A (Hypocretin-1) and leptin enhance LTP in the dentate gyrus of rats in vivo. Peptides. 2004;25:991–996. doi: 10.1016/j.peptides.2004.03.018. [DOI] [PubMed] [Google Scholar]
- 18.O’Malley D, et al. Leptin promotes rapid dynamic changes in hippocampal dendritic morphology. Mol Cell Neurosci. 2007;35:559–572. doi: 10.1016/j.mcn.2007.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.van der Lely AJ, Tschop M, Heiman ML, Ghigo E. Biological, physiological, pathophysiological, and pharmacological aspects of ghrelin. Endocr Rev. 2004;25:426–457. doi: 10.1210/er.2002-0029. [DOI] [PubMed] [Google Scholar]
- 20.Asakawa A, et al. Antagonism of ghrelin receptor reduces food intake and body weight gain in mice. Gut. 2003;52:947–952. doi: 10.1136/gut.52.7.947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cummings DE, et al. A preprandial rise in plasma ghrelin levels suggests a role in meal initiation in humans. Diabetes. 2001;50:1714–1719. doi: 10.2337/diabetes.50.8.1714. [DOI] [PubMed] [Google Scholar]
- 22.Shuto Y, et al. Generation of polyclonal antiserum against the growth hormone secretagogue receptor (GHS-R): evidence that the GHS-R exists in the hypothalamus, pituitary and stomach of rats. Life Sci. 2001;68:991–996. doi: 10.1016/s0024-3205(00)01001-8. [DOI] [PubMed] [Google Scholar]
- 23.Guan XM, et al. Distribution of mRNA encoding the growth hormone secretagogue receptor in brain and peripheral tissues. Brain Res Mol Brain Res. 1997;48:23–29. doi: 10.1016/s0169-328x(97)00071-5. [DOI] [PubMed] [Google Scholar]
- 24.Tschop M, Smiley DL, Heiman ML. Ghrelin induces adiposity in rodents. Nature. 2000;407:908–913. doi: 10.1038/35038090. [DOI] [PubMed] [Google Scholar]
- 25.Wren AM, et al. The novel hypothalamic peptide ghrelin stimulates food intake and growth hormone secretion. Endocrinology. 2000;141:4325–4328. doi: 10.1210/endo.141.11.7873. [DOI] [PubMed] [Google Scholar]
- 26.Druce MR, et al. Subcutaneous administration of ghrelin stimulates energy intake in healthy lean human volunteers. Int J Obes (Lond) 2006;30:293–296. doi: 10.1038/sj.ijo.0803158. [DOI] [PubMed] [Google Scholar]
- 27.Wren AM, et al. Ghrelin enhances appetite and increases food intake in humans. J Clin Endocrinol Metab. 2001;86:5992. doi: 10.1210/jcem.86.12.8111. [DOI] [PubMed] [Google Scholar]
- 28.Pinto S, et al. Rapid rewiring of arcuate nucleus feeding circuits by leptin. Science. 2004;304:110–115. doi: 10.1126/science.1089459. [DOI] [PubMed] [Google Scholar]
- 29.Diano S, et al. Ghrelin controls hippocampal spine synapse density and memory performance. Nature Neurosci. 2006;9:381–388. doi: 10.1038/nn1656. [DOI] [PubMed] [Google Scholar]
- 30.During MJ, et al. Glucagon-like peptide-1 receptor is involved in learning and neuroprotection. Nature Med. 2003;9:1173–1179. doi: 10.1038/nm919. [DOI] [PubMed] [Google Scholar]
- 31.McNay EC. Insulin and ghrelin: peripheral hormones modulating memory and hippocampal function. Curr Opin Pharmacol. 2007;7:628–632. doi: 10.1016/j.coph.2007.10.009. [DOI] [PubMed] [Google Scholar]
- 32.Vaynman S, Ying Z, Wu A, Gomez-Pinilla F. Coupling energy metabolism with a mechanism to support brain-derived neurotrophic factor-mediated synaptic plasticity. Neuroscience. 2006;139:1221–1234. doi: 10.1016/j.neuroscience.2006.01.062. [DOI] [PubMed] [Google Scholar]
- 33.Nawa H, Carnahan J, Gall C. BDNF protein measured by a novel enzyme immunoassay in normal brain and after seizure: partial disagreement with mRNA levels. Eur J Neurosci. 1995;7:1527–1535. doi: 10.1111/j.1460-9568.1995.tb01148.x. [DOI] [PubMed] [Google Scholar]
- 34.Kesslak JP, So V, Choi J, Cotman CW, Gomez-Pinilla F. Learning upregulates brain-derived neurotrophic factor messenger ribonucleic acid: a mechanism to facilitate encoding and circuit maintenance? Behav Neurosci. 1998;112:1012–1019. doi: 10.1037//0735-7044.112.4.1012. [DOI] [PubMed] [Google Scholar]
- 35.Hall J, Thomas KL, Everitt BJ. Rapid and selective induction of BDNF expression in the hippocampus during contextual learning. Nature Neurosci. 2000;3:533–535. doi: 10.1038/75698. [DOI] [PubMed] [Google Scholar]
- 36.Linnarsson S, Bjorklund A, Ernfors P. Learning deficit in BDNF mutant mice. Eur J Neurosci. 1997;9:2581–2587. doi: 10.1111/j.1460-9568.1997.tb01687.x. [DOI] [PubMed] [Google Scholar]
- 37.Ma YL, Wang HL, Wu HC, Wei CL, Lee EH. Brain-derived neurotrophic factor antisense oligonucleotide impairs memory retention and inhibits long-term potentiation in rats. Neuroscience. 1998;82:957–967. doi: 10.1016/s0306-4522(97)00325-4. [DOI] [PubMed] [Google Scholar]
- 38.Chen ZY, et al. Variant brain-derived neurotrophic factor (BDNF) (Met66) alters the intracellular trafficking and activity-dependent secretion of wild-type BDNF in neurosecretory cells and cortical neurons. J Neurosci. 2004;24:4401–4411. doi: 10.1523/JNEUROSCI.0348-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Egan MF, et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell. 2003;112:257–269. doi: 10.1016/s0092-8674(03)00035-7. [DOI] [PubMed] [Google Scholar]
- 40.Kernie SG, Liebl DJ, Parada LF. BDNF regulates eating behavior and locomotor activity in mice. EMBO J. 2000;19:1290–1300. doi: 10.1093/emboj/19.6.1290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lyons WE, et al. Brain-derived neurotrophic factor-deficient mice develop aggressiveness and hyperphagia in conjunction with brain serotonergic abnormalities. Proc Natl Acad Sci USA. 1999;96:15239–15244. doi: 10.1073/pnas.96.26.15239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Pelleymounter MA, Cullen MJ, Wellman CL. Characteristics of BDNF-induced weight loss. Exp Neurol. 1995;131:229–238. doi: 10.1016/0014-4886(95)90045-4. [DOI] [PubMed] [Google Scholar]
- 43.Nakagawa T, et al. Brain-derived neurotrophic factor (BDNF) regulates glucose and energy metabolism in diabetic mice. Diabetes Metab Res Rev. 2002;18:185–191. doi: 10.1002/dmrr.290. [DOI] [PubMed] [Google Scholar]
- 44.Tonra JR, et al. Brain-derived neurotrophic factor improves blood glucose control and alleviates fasting hyperglycemia in C57BLKS-Leprdb/leprdb mice. Diabetes. 1999;48:588–594. doi: 10.2337/diabetes.48.3.588. [DOI] [PubMed] [Google Scholar]
- 45.Tsuchida A, et al. Brain-derived neurotrophic factor ameliorates lipid metabolism in diabetic mice. Diabetes Obes Metab. 2002;4:262–269. doi: 10.1046/j.1463-1326.2002.00206.x. [DOI] [PubMed] [Google Scholar]
- 46.Xu B, et al. Brain-derived neurotrophic factor regulates energy balance downstream of melanocortin-4 receptor. Nature Neurosci. 2003;6:736–742. doi: 10.1038/nn1073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Yeo GS, et al. A de novo mutation affecting human TrkB associated with severe obesity and developmental delay. Nature Neurosci. 2004;7:1187–1189. doi: 10.1038/nn1336. [DOI] [PubMed] [Google Scholar]
- 48.Ding Q, Vaynman S, Akhavan M, Ying Z, Gomez-Pinilla F. Insulin-like growth factor I interfaces with brain-derived neurotrophic factor-mediated synaptic plasticity to modulate aspects of exercise-induced cognitive function. Neuroscience. 2006;140:823–833. doi: 10.1016/j.neuroscience.2006.02.084. [DOI] [PubMed] [Google Scholar]
- 49.Islam A, et al. Changes in IGF-1 receptors in the hippocampus of adult rats after long-term adrenalectomy: receptor autoradiography and in situ hybridization histochemistry. Brain Res. 1998;797:342–346. doi: 10.1016/s0006-8993(98)00389-8. [DOI] [PubMed] [Google Scholar]
- 50.Foster LA, Ames NK, Emery RS. Food intake and serum insulin responses to intraventricular infusions of insulin and IGF-I. Physiol Behav. 1991;50:745–749. doi: 10.1016/0031-9384(91)90012-d. [DOI] [PubMed] [Google Scholar]
- 51.Anlar B, Sullivan KA, Feldman EL. Insulin-like growth factor-I and central nervous system development. Horm Metab Res. 1999;31:120–125. doi: 10.1055/s-2007-978708. [DOI] [PubMed] [Google Scholar]
- 52.Torres-Aleman I. Insulin-like growth factors as mediators of functional plasticity in the adult brain. Horm Metab Res. 1999;31:114–119. doi: 10.1055/s-2007-978707. [DOI] [PubMed] [Google Scholar]
- 53.Saatman KE, et al. Insulin-like growth factor-1 (IGF-1) improves both neurological motor and cognitive outcome following experimental brain injury. Exp Neurol. 1997;147:418–427. doi: 10.1006/exnr.1997.6629. [DOI] [PubMed] [Google Scholar]
- 54.Carro E, Trejo JL, Busiguina S, Torres-Aleman I. Circulating insulin-like growth factor I mediates the protective effects of physical exercise against brain insults of different etiology and anatomy. J Neurosci. 2001;21:5678–5684. doi: 10.1523/JNEUROSCI.21-15-05678.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lupien SB, Bluhm EJ, Ishii DN. Systemic insulin-like growth factor-I administration prevents cognitive impairment in diabetic rats, and brain IGF regulates learning/memory in normal adult rats. J Neurosci Res. 2003;74:512–523. doi: 10.1002/jnr.10791. [DOI] [PubMed] [Google Scholar]
- 56.Markowska AL, Mooney M, Sonntag WE. Insulin-like growth factor-1 ameliorates age-related behavioral deficits. Neuroscience. 1998;87:559–569. doi: 10.1016/s0306-4522(98)00143-2. [DOI] [PubMed] [Google Scholar]
- 57.Johnson-Farley NN, Patel K, Kim D, Cowen DS. Interaction of FGF-2 with IGF-1 and BDNF in stimulating Akt, ERK, and neuronal survival in hippocampal cultures. Brain Res. 2007;1154:40–49. doi: 10.1016/j.brainres.2007.04.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Akbar M, Calderon F, Wen Z, Kim HY. Docosahexaenoic acid: a positive modulator of Akt signaling in neuronal survival. Proc Natl Acad Sci USA. 2005;102:10858–10863. doi: 10.1073/pnas.0502903102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Sacks FM. Metabolic syndrome: epidemiology and consequences. J Clin Psychiatry. 2004;65 (Suppl 18):3–12. [PubMed] [Google Scholar]
- 60.Lilliker SL. Prevalence of diabetes in a manic-depressive population. Compr Psychiatry. 1980;21:270–275. doi: 10.1016/0010-440x(80)90030-9. [DOI] [PubMed] [Google Scholar]
- 61.Mukherjee S, Decina P, Bocola V, Saraceni F, Scapicchio PL. Diabetes mellitus in schizophrenic patients. Compr Psychiatry. 1996;37:68–73. doi: 10.1016/s0010-440x(96)90054-1. [DOI] [PubMed] [Google Scholar]
- 62.Dixon L, et al. Prevalence and correlates of diabetes in national schizophrenia samples. Schizophr Bull. 2000;26:903–912. doi: 10.1093/oxfordjournals.schbul.a033504. [DOI] [PubMed] [Google Scholar]
- 63.Angelucci F, Brene S, Mathe AA. BDNF in schizophrenia, depression and corresponding animal models. Mol Psychiatry. 2005;10:345–352. doi: 10.1038/sj.mp.4001637. [DOI] [PubMed] [Google Scholar]
- 64.Nestler EJ, et al. Neurobiology of depression. Neuron. 2002;34:13–25. doi: 10.1016/s0896-6273(02)00653-0. [DOI] [PubMed] [Google Scholar]
- 65.Martinowich K, Manji H, Lu B. New insights into BDNF function in depression and anxiety. Nature Neurosci. 2007;10:1089–1093. doi: 10.1038/nn1971. [DOI] [PubMed] [Google Scholar]
- 66.Duman RS. Synaptic plasticity and mood disorders. Mol Psychiatry. 2002;7 (Suppl 1):S29–S34. doi: 10.1038/sj.mp.4001016. [DOI] [PubMed] [Google Scholar]
- 67.Krabbe KS, et al. Brain-derived neurotrophic factor (BDNF) and type 2 diabetes. Diabetologia. 2007;50:431–438. doi: 10.1007/s00125-006-0537-4. [DOI] [PubMed] [Google Scholar]
- 68.Durany N, et al. Brain-derived neurotrophic factor and neurotrophin 3 in schizophrenic psychoses. Schizophr Res. 2001;52:79–86. doi: 10.1016/s0920-9964(00)00084-0. [DOI] [PubMed] [Google Scholar]
- 69.Toyooka K, et al. Decreased levels of brain-derived neurotrophic factor in serum of chronic schizophrenic patients. Psychiatry Res. 2002;110:249–257. doi: 10.1016/s0165-1781(02)00127-0. [DOI] [PubMed] [Google Scholar]
- 70.Zorner B, et al. Forebrain-specific trkB-receptor knockout mice: behaviorally more hyperactive than “depressive”. Biol Psychiatry. 2003;54:972–982. doi: 10.1016/s0006-3223(03)00418-9. [DOI] [PubMed] [Google Scholar]
- 71.Karege F, et al. Decreased serum brain-derived neurotrophic factor levels in major depressed patients. Psychiatry Res. 2002;109:143–148. doi: 10.1016/s0165-1781(02)00005-7. [DOI] [PubMed] [Google Scholar]
- 72.Chen B, Dowlatshahi D, MacQueen GM, Wang JF, Young LT. Increased hippocampal BDNF immunoreactivity in subjects treated with antidepressant medication. Biol Psychiatry. 2001;50:260–265. doi: 10.1016/s0006-3223(01)01083-6. [DOI] [PubMed] [Google Scholar]
- 73.Tsankova NM, et al. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nature Neurosci. 2006;9:519–525. doi: 10.1038/nn1659. [DOI] [PubMed] [Google Scholar]
- 74.Bourre JM, et al. The effects of dietary α-linolenic acid on the composition of nerve membranes, enzymatic activity, amplitude of electrophysiological parameters, resistance to poisons and performance of learning tasks in rats. J Nutr. 1989;119:1880–1892. doi: 10.1093/jn/119.12.1880. [DOI] [PubMed] [Google Scholar]
- 75.Moriguchi T, Greiner RS, Salem N., Jr Behavioral deficits associated with dietary induction of decreased brain docosahexaenoic acid concentration. J Neurochem. 2000;75:2563–2573. doi: 10.1046/j.1471-4159.2000.0752563.x. [DOI] [PubMed] [Google Scholar]
- 76.Adams PB, Lawson S, Sanigorski A, Sinclair AJ. Arachidonic acid to eicosapentaenoic acid ratio in blood correlates positively with clinical symptoms of depression. Lipids. 1996;31 (Suppl):157–161. doi: 10.1007/BF02637069. [DOI] [PubMed] [Google Scholar]
- 77.Peet M, Laugharne JD, Mellor J, Ramchand CN. Essential fatty acid deficiency in erythrocyte membranes from chronic schizophrenic patients, and the clinical effects of dietary supplementation. Prostaglandins Leukot Essent Fatty Acids. 1996;55:71–75. doi: 10.1016/s0952-3278(96)90148-9. [DOI] [PubMed] [Google Scholar]
- 78.Hibbeln JR. Fish consumption and major depression. Lancet. 1998;351:1213. doi: 10.1016/S0140-6736(05)79168-6. [DOI] [PubMed] [Google Scholar]
- 79.Horrobin DF. Schizophrenia: the illness that made us human. Med Hypotheses. 1998;50:269–288. doi: 10.1016/s0306-9877(98)90000-7. [DOI] [PubMed] [Google Scholar]
- 80.Freeman MP, et al. Omega-3 fatty acids: evidence basis for treatment and future research in psychiatry. J Clin Psychiatry. 2006;67:1954–1967. doi: 10.4088/jcp.v67n1217. [DOI] [PubMed] [Google Scholar]
- 81.Wu A, Ying Z, Gómez-Pinilla F. Dietary Omega-3 fatty acids normalize BDNF levels, reduce oxidative damage, and counteract learning disability after traumatic brain injury in rats. J Neurotrauma. 2004;21:1457–1467. doi: 10.1089/neu.2004.21.1457. [DOI] [PubMed] [Google Scholar]
- 82.Pifferi F, et al. (n-3) polyunsaturated fatty acid deficiency reduces the expression of both isoforms of the brain glucose transporter GLUT1 in rats. J Nutr. 2005;135:2241–2246. doi: 10.1093/jn/135.9.2241. [DOI] [PubMed] [Google Scholar]
- 83.Flachs P, et al. Polyunsaturated fatty acids of marine origin upregulate mitochondrial biogenesis and induce β-oxidation in white fat. Diabetologia. 2005;48:2365–2375. doi: 10.1007/s00125-005-1944-7. [DOI] [PubMed] [Google Scholar]
- 84.Portwood MM. The role of dietary fatty acids in children’s behaviour and learning. Nutr Health. 2006;18:233–247. doi: 10.1177/026010600601800306. [DOI] [PubMed] [Google Scholar]
- 85.Richardson AJ, Montgomery P. The Oxford-Durham study: a randomized, controlled trial of dietary supplementation with fatty acids in children with developmental coordination disorder. Pediatrics. 2005;115:1360–1366. doi: 10.1542/peds.2004-2164. [DOI] [PubMed] [Google Scholar]
- 86.Osendarp SJ, et al. Effect of a 12-mo micronutrient intervention on learning and memory in well-nourished and marginally nourished school-aged children: 2 parallel, randomized, placebo-controlled studies in Australia and Indonesia. Am J Clin Nutr. 2007;86:1082–1093. doi: 10.1093/ajcn/86.4.1082. [DOI] [PubMed] [Google Scholar]
- 87.Innis SM. Dietary (n-3) fatty acids and brain development. J Nutr. 2007;137:855–859. doi: 10.1093/jn/137.4.855. [DOI] [PubMed] [Google Scholar]
- 88.Wu A, Molteni R, Ying Z, Gomez-Pinilla F. A saturated-fat diet aggravates the outcome of traumatic brain injury on hippocampal plasticity and cognitive function by reducing brain-derived neurotrophic factor. Neuroscience. 2003;119:365–375. doi: 10.1016/s0306-4522(03)00154-4. [DOI] [PubMed] [Google Scholar]
- 89.Wu A, Ying Z, Gómez-Pinilla F. Dietary curcumin counteracts the outcome of traumatic brain injury on oxidative stress, synaptic plasticity, and cognition. Exp Neurol. 2006;197:309–317. doi: 10.1016/j.expneurol.2005.09.004. [DOI] [PubMed] [Google Scholar]
- 90.Wu A, Ying Z, Gómez-Pinilla F. Oxidative stress modulates Sir2α in rat hippocampus and cerebral cortex. Eur J Neurosci. 2006;23:2573–2580. doi: 10.1111/j.1460-9568.2006.04807.x. [DOI] [PubMed] [Google Scholar]
- 91.Pu F, et al. Neuroprotective effects of quercetin and rutin on spatial memory impairment in an 8-arm radial maze task and neuronal death induced by repeated cerebral ischemia in rats. J Pharmacol Sci. 2007;104:329–334. doi: 10.1254/jphs.fp0070247. [DOI] [PubMed] [Google Scholar]
- 92.van Praag H, et al. Plant-derived flavanol (−) epicatechin enhances angiogenesis and retention of spatial memory in mice. J Neurosci. 2007;27:5869–5878. doi: 10.1523/JNEUROSCI.0914-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Mischoulon D, Raab MF. The role of folate in depression and dementia. J Clin Psychiatry. 2007;68 (Suppl 10):28–33. [PubMed] [Google Scholar]
- 94.Herbert V. Experimental nutritional folate deficiency in man. Trans Assoc Am Physicians. 1962;75:307–320. [PubMed] [Google Scholar]
- 95.Corrada M, Kawas C, Hallfrisch J, Muller D, Brookmeyer R. Reduced risk of Alzheimer’s disease with high folate intake: The Baltimore Longitudinal Study of Aging. Alzheimers Dement. 2005;1:A4. doi: 10.1016/j.jalz.2005.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Fioravanti M, et al. Low folate levels in the cognitive decline of elderly patients and efficacy of folate as a treatment for improving memory deficits. Arch Gerontol Geriatr. 1997;26:1–13. doi: 10.1016/s0167-4943(97)00028-9. [DOI] [PubMed] [Google Scholar]
- 97.Nilsson K, Gustafson L, Hultberg B. Improvement of cognitive functions after cobalamin/folate supplementation in elderly patients with dementia and elevated plasma homocysteine. Int J Geriatr Psychiatry. 2001;16:609–614. doi: 10.1002/gps.388. [DOI] [PubMed] [Google Scholar]
- 98.Ramos MI, et al. Low folate status is associated with impaired cognitive function and dementia in the Sacramento Area Latino Study on Aging. Am J Clin Nutr. 2005;82:1346–1352. doi: 10.1093/ajcn/82.6.1346. [DOI] [PubMed] [Google Scholar]
- 99.Fava M, et al. Folate, vitamin B12, and homocysteine in major depressive disorder. Am J Psychiatry. 1997;154:426–428. doi: 10.1176/ajp.154.3.426. [DOI] [PubMed] [Google Scholar]
- 100.Durga J, et al. Effect of 3-year folic acid supplementation on cognitive function in older adults in the FACIT trial: a randomised, double blind, controlled trial. Lancet. 2007;369:208–216. doi: 10.1016/S0140-6736(07)60109-3. [DOI] [PubMed] [Google Scholar]
- 101.Folic acid: a supplementary question. Lancet Neurol. 2007;6:199. doi: 10.1016/S1474-4422(07)70038-8. [No authors listed] [DOI] [PubMed] [Google Scholar]
- 102.Wu A, Ying Z, Gomez-Pinilla F. The interplay between oxidative stress and brain-derived neurotrophic factor modulates the outcome of a saturated fat diet on synaptic plasticity and cognition. Eur J Neurosci. 2004;19:1699–1707. doi: 10.1111/j.1460-9568.2004.03246.x. [DOI] [PubMed] [Google Scholar]
- 103.Mattson MP. Energy intake, meal frequency, and health: a neurobiological perspective. Annu Rev Nutr. 2005;25:237–260. doi: 10.1146/annurev.nutr.25.050304.092526. [DOI] [PubMed] [Google Scholar]
- 104.Mattson MP. Dietary factors, hormesis and health. Ageing Res Rev. 2007;7:43–48. doi: 10.1016/j.arr.2007.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Duan W, et al. Dietary restriction normalizes glucose metabolism and BDNF levels, slows disease progression, and increases survival in huntingtin mutant mice. Proc Natl Acad Sci USA. 2003;100:2911–2916. doi: 10.1073/pnas.0536856100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Lee J, Seroogy KB, Mattson MP. Dietary restriction enhances neurotrophin expression and neurogenesis in the hippocampus of adult mice. J Neurochem. 2002;80:539–547. doi: 10.1046/j.0022-3042.2001.00747.x. [DOI] [PubMed] [Google Scholar]
- 107.Ingram DK, Weindruch R, Spangler EL, Freeman JR, Walford RL. Dietary restriction benefits learning and motor performance of aged mice. J Gerontol. 1987;42:78–81. doi: 10.1093/geronj/42.1.78. [DOI] [PubMed] [Google Scholar]
- 108.Halagappa VK, et al. Intermittent fasting and caloric restriction ameliorate age-related behavioral deficits in the triple-transgenic mouse model of Alzheimer’s disease. Neurobiol Dis. 2007;26:212–220. doi: 10.1016/j.nbd.2006.12.019. [DOI] [PubMed] [Google Scholar]
- 109.Booth FW, Chakravarthy MV, Gordon SE, Spangenburg EE. Waging war on physical inactivity: using modern molecular ammunition against an ancient enemy. J Appl Physiol. 2002;93:3–30. doi: 10.1152/japplphysiol.00073.2002. [DOI] [PubMed] [Google Scholar]
- 110.Carlson O, et al. Impact of reduced meal frequency without caloric restriction on glucose regulation in healthy, normal-weight middle-aged men and women. Metabolism. 2007;56:1729–1734. doi: 10.1016/j.metabol.2007.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Johnson JB, et al. Alternate day calorie restriction improves clinical findings and reduces markers of oxidative stress and inflammation in overweight adults with moderate asthma. Free Radic Biol Med. 2007;42:665–674. doi: 10.1016/j.freeradbiomed.2006.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Joseph JA, Shukitt-Hale B, Lau FC. Fruit polyphenols and their effects on neuronal signaling and behavior in senescence. Ann NY Acad Sci. 2007;1100:470–485. doi: 10.1196/annals.1395.052. [DOI] [PubMed] [Google Scholar]
- 113.Galli RL, Bielinski DF, Szprengiel A, Shukitt-Hale B, Joseph JA. Blueberry supplemented diet reverses age-related decline in hippocampal HSP70 neuroprotection. Neurobiol Aging. 2006;27:344–350. doi: 10.1016/j.neurobiolaging.2005.01.017. [DOI] [PubMed] [Google Scholar]
- 114.Casadesus G, et al. Modulation of hippocampal plasticity and cognitive behavior by short-term blueberry supplementation in aged rats. Nutr Neurosci. 2004;7:309–316. doi: 10.1080/10284150400020482. [DOI] [PubMed] [Google Scholar]
- 115.Duffy KB, et al. A blueberry-enriched diet provides cellular protection against oxidative stress and reduces a kainate-induced learning impairment in rats. Neurobiol Aging. 2007 May 22; doi: 10.1016/j.neurobiolaging.2007.04.002. [epub ahead of print] [DOI] [PubMed] [Google Scholar]
- 116.Liu J. The effects and mechanisms of mitochondrial nutrient α-lipoic acid on improving age-associated mitochondrial and cognitive dysfunction: an overview. Neurochem Res. 2008;33:194–203. doi: 10.1007/s11064-007-9403-0. [DOI] [PubMed] [Google Scholar]
- 117.Quinn JF, et al. Chronic dietary α-lipoic acid reduces deficits in hippocampal memory of aged Tg2576 mice. Neurobiol Aging. 2007;28:213–225. doi: 10.1016/j.neurobiolaging.2005.12.014. [DOI] [PubMed] [Google Scholar]
- 118.Holmquist L, et al. Lipoic acid as a novel treatment for Alzheimer’s disease and related dementias. Pharmacol Ther. 2007;113:154–164. doi: 10.1016/j.pharmthera.2006.07.001. [DOI] [PubMed] [Google Scholar]
- 119.Perkins AJ, et al. Association of antioxidants with memory in a multiethnic elderly sample using the Third National Health and Nutrition Examination Survey. Am J Epidemiol. 1999;150:37–44. doi: 10.1093/oxfordjournals.aje.a009915. [DOI] [PubMed] [Google Scholar]
- 120.Navarro A, et al. Vitamin E at high doses improves survival, neurological performance, and brain mitochondrial function in aging male mice. Am J Physiol Regul Integr Comp Physiol. 2005;289:R1392–R1399. doi: 10.1152/ajpregu.00834.2004. [DOI] [PubMed] [Google Scholar]
- 121.Ammon HP, Wahl MA. Pharmacology of Curcuma longa. Planta Med. 1991;57:1–7. doi: 10.1055/s-2006-960004. [DOI] [PubMed] [Google Scholar]
- 122.Kelloff GJ, et al. Progress in cancer chemoprevention: development of diet-derived chemopreventive agents. J Nutr. 2000;130:467S–471S. doi: 10.1093/jn/130.2.467S. [DOI] [PubMed] [Google Scholar]
- 123.Frautschy SA, et al. Phenolic anti-inflammatory antioxidant reversal of Aβ-induced cognitive deficits and neuropathology. Neurobiol Aging. 2001;22:993–1005. doi: 10.1016/s0197-4580(01)00300-1. [DOI] [PubMed] [Google Scholar]
- 124.Ganguli M, et al. Apolipoprotein E polymorphism and Alzheimer disease: the Indo-US Cross-National Dementia Study. Arch Neurol. 2000;57:824–830. doi: 10.1001/archneur.57.6.824. [DOI] [PubMed] [Google Scholar]
- 125.Martin-Aragon S, Benedi JM, Villar AM. Modifications on antioxidant capacity and lipid peroxidation in mice under fraxetin treatment. J Pharm Pharmacol. 1997;49:49–52. doi: 10.1111/j.2042-7158.1997.tb06751.x. [DOI] [PubMed] [Google Scholar]
- 126.Sreejayan N, Rao MN. Nitric oxide scavenging by curcuminoids. J Pharm Pharmacol. 1997;49:105–107. doi: 10.1111/j.2042-7158.1997.tb06761.x. [DOI] [PubMed] [Google Scholar]
- 127.Tsankova N, Renthal W, Kumar A, Nestler EJ. Epigenetic regulation in psychiatric disorders. Nature Rev Neurosci. 2007;8:355–367. doi: 10.1038/nrn2132. [DOI] [PubMed] [Google Scholar]
- 128.Garzon D, Yu G, Fahnestock M. A new brain-derived neurotrophic factor transcript and decrease in brain-derived neurotrophic factor transcripts 1, 2 and 3 in Alzheimer’s disease parietal cortex. J Neurochem. 2002;82:1058–1064. doi: 10.1046/j.1471-4159.2002.01030.x. [DOI] [PubMed] [Google Scholar]
- 129.Chen WG, et al. Derepression of BDNF transcription involves calcium-dependent phosphorylation of MeCP2. Science. 2003;302:885–889. doi: 10.1126/science.1086446. [DOI] [PubMed] [Google Scholar]
- 130.Kuningas M, Putters M, Westendorp RG, Slagboom PE, van Heemst D. SIRT1 gene, age-related diseases, and mortality: the Leiden 85-plus study. J Gerontol A Biol Sci Med Sci. 2007;62:960–965. doi: 10.1093/gerona/62.9.960. [DOI] [PubMed] [Google Scholar]
- 131.Vaynman S, Gomez-Pinilla F. Revenge of the “sit”: how lifestyle impacts neuronal and cognitive health through molecular systems that interface energy metabolism with neuronal plasticity. J Neurosci Res. 2006;84:699–715. doi: 10.1002/jnr.20979. [DOI] [PubMed] [Google Scholar]
- 132.Wu A, Ying Z, Gomez-Pinilla F. DHA dietary supplementation enhances the effects of exercise on synaptic plasticity and cognition. Neuroscience. doi: 10.1016/j.neuroscience.2008.05.061. (in the press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Garcia J, Ervin FR. Appetites, aversions, and addictions: a model for visceral memory. Recent Adv Biol Psychiatry. 1968;10:284–293. doi: 10.1007/978-1-4684-9072-5_24. [DOI] [PubMed] [Google Scholar]
- 134.Arana FS, et al. Dissociable contributions of the human amygdala and orbitofrontal cortex to incentive motivation and goal selection. J Neurosci. 2003;23:9632–9638. doi: 10.1523/JNEUROSCI.23-29-09632.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Thompson ME, Wrangham RW. Diet and reproductive function in wild female chimpanzees (Pan troglodytes schweinfurthii) at Kibale National Park, Uganda. Am J Phys Anthropol. 2008;135:171–181. doi: 10.1002/ajpa.20718. [DOI] [PubMed] [Google Scholar]
- 136.Gibbons A. Paleoanthropology. Food for thought Science. 2007;316:1558–1560. doi: 10.1126/science.316.5831.1558. [DOI] [PubMed] [Google Scholar]
- 137.Crawford MA, Sinclair AJ. The limitations of whole tissue analysis to define linolenic acid deficiency. J Nutr. 1972;102:1315–1321. doi: 10.1093/jn/102.10.1315. [DOI] [PubMed] [Google Scholar]
- 138.Crawford MA, et al. Evidence for the unique function of docosahexaenoic acid during the evolution of the modern hominid brain. Lipids. 1999;34 (Suppl):39–47. doi: 10.1007/BF02562227. [DOI] [PubMed] [Google Scholar]
- 139.Hillman CH, Erickson KI, Kramer AF. Be smart, exercise your heart: exercise effects on brain and cognition. Nature Rev Neurosci. 2008;9:58–65. doi: 10.1038/nrn2298. [DOI] [PubMed] [Google Scholar]
- 140.Gomez-Pinilla F. The influences of diet and exercise on mental health through hormesis. Ageing Res Rev. 2008;7:49–62. doi: 10.1016/j.arr.2007.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Kramer AF, et al. Ageing, fitness and neurocognitive function. Nature. 1999;400:418–419. doi: 10.1038/22682. [DOI] [PubMed] [Google Scholar]
- 142.Winter B, et al. High impact running improves learning. Neurobiol Learn Mem. 2007;87:597–609. doi: 10.1016/j.nlm.2006.11.003. [DOI] [PubMed] [Google Scholar]
- 143.van Praag H, Kempermann G, Gage FH. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nature Neurosci. 1999;2:266–270. doi: 10.1038/6368. [DOI] [PubMed] [Google Scholar]
- 144.Pereira AC, et al. An in vivo correlate of exercise-induced neurogenesis in the adult dentate gyrus. Proc Natl Acad Sci USA. 2007;104:5638–5643. doi: 10.1073/pnas.0611721104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Vaynman S, Ying Z, Gomez-Pinilla F. Hippocampal BDNF mediates the efficacy of exercise on synaptic plasticity and cognition. Eur J Neurosci. 2004;20:2580–2590. doi: 10.1111/j.1460-9568.2004.03720.x. [DOI] [PubMed] [Google Scholar]
- 146.Laurin D, Verreault R, Lindsay J, MacPherson K, Rockwood K. Physical activity and risk of cognitive impairment and dementia in elderly persons. Arch Neurol. 2001;58:498–504. doi: 10.1001/archneur.58.3.498. [DOI] [PubMed] [Google Scholar]
- 147.Ding Q, Vaynman S, Souda P, Whitelegge JP, Gomez-Pinilla F. Exercise affects energy metabolism and neural plasticity-related proteins in the hippocampus as revealed by proteomic analysis. Eur J Neurosci. 2006;24:1265–1276. doi: 10.1111/j.1460-9568.2006.05026.x. [DOI] [PubMed] [Google Scholar]
- 148.van Gelder BM, Tijhuis M, Kalmijn S, Kromhout D. Fish consumption, n-3 fatty acids, and subsequent 5-y cognitive decline in elderly men: the Zutphen Elderly Study. Am J Clin Nutr. 2007;85:1142–1147. doi: 10.1093/ajcn/85.4.1142. [DOI] [PubMed] [Google Scholar]
- 149.Hashimoto M, et al. Chronic administration of docosahexaenoic acid ameliorates the impairment of spatial cognition learning ability in amyloid β-infused rats. J Nutr. 2005;135:549–555. doi: 10.1093/jn/135.3.549. [DOI] [PubMed] [Google Scholar]
- 150.Calon F, et al. Docosahexaenoic acid protects from dendritic pathology in an Alzheimer’s disease mouse model. Neuron. 2004;43:633–645. doi: 10.1016/j.neuron.2004.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Letenneur L, Proust-Lima C, Le Gouge A, Dartigues JF, Barberger-Gateau P. Flavonoid intake and cognitive decline over a 10-year period. Am J Epidemiol. 2007;165:1364–1371. doi: 10.1093/aje/kwm036. [DOI] [PubMed] [Google Scholar]
- 152.Bryan J, Calvaresi E, Hughes D. Short-term folate, vitamin B-12 or vitamin B-6 supplementation slightly affects memory performance but not mood in women of various ages. J Nutr. 2002;132:1345–1356. doi: 10.1093/jn/132.6.1345. [DOI] [PubMed] [Google Scholar]
- 153.Sasaki H, et al. Vitamin B12 improves cognitive disturbance in rodents fed a choline-deficient diet. Pharmacol Biochem Behav. 1992;43:635–639. doi: 10.1016/0091-3057(92)90204-s. [DOI] [PubMed] [Google Scholar]
- 154.Przybelski RJ, Binkley NC. Is vitamin D important for preserving cognition? A positive correlation of serum 25-hydroxyvitamin D concentration with cognitive function. Arch Biochem Biophys. 2007;460:202–205. doi: 10.1016/j.abb.2006.12.018. [DOI] [PubMed] [Google Scholar]
- 155.Holmes GL, et al. Seizure-induced memory impairment is reduced by choline supplementation before or after status epilepticus. Epilepsy Res. 2002;48:3–13. doi: 10.1016/s0920-1211(01)00321-7. [DOI] [PubMed] [Google Scholar]
- 156.McCann JC, Hudes M, Ames BN. An overview of evidence for a causal relationship between dietary availability of choline during development and cognitive function in offspring. Neurosci Biobehav Rev. 2006;30:696–712. doi: 10.1016/j.neubiorev.2005.12.003. [DOI] [PubMed] [Google Scholar]
- 157.Wengreen HJ, et al. Antioxidant intake and cognitive function of elderly men and women: the Cache County Study. J Nutr Health Aging. 2007;11:230–237. [PubMed] [Google Scholar]
- 158.Schram MT, et al. Serum calcium and cognitive function in old age. J Am Geriatr Soc. 2007;55:1786–1792. doi: 10.1111/j.1532-5415.2007.01418.x. [DOI] [PubMed] [Google Scholar]
- 159.Ortega RM, et al. Dietary intake and cognitive function in a group of elderly people. Am J Clin Nutr. 1997;66:803–809. doi: 10.1093/ajcn/66.4.803. [DOI] [PubMed] [Google Scholar]
- 160.Gao S, et al. Selenium level and cognitive function in rural elderly Chinese. Am J Epidemiol. 2007;165:955–965. doi: 10.1093/aje/kwk073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Pajonk FG, et al. Cognitive decline correlates with low plasma concentrations of copper in patients with mild to moderate Alzheimer’s disease. J Alzheimers Dis. 2005;8:23–27. doi: 10.3233/jad-2005-8103. [DOI] [PubMed] [Google Scholar]
- 162.Murray-Kolb LE, Beard JL. Iron treatment normalizes cognitive functioning in young women. Am J Clin Nutr. 2007;85:778–787. doi: 10.1093/ajcn/85.3.778. [DOI] [PubMed] [Google Scholar]
- 163.Adams PF, Marano MA. Current estimates for the national health interview survey, 1994. Vital Health Stat 10. 1995;193 (Pt 2):261–520. [PubMed] [Google Scholar]
- 164.Kaati G, Bygren LO, Pembrey M, Sjöström M. Transgenerational response to nutrition, early life circumstances and longevity. Eur J Hum Genet. 2007;15:784–790. doi: 10.1038/sj.ejhg.5201832. [DOI] [PubMed] [Google Scholar]