

Abstract
Biosynthetic pathways can be hijacked to yield novel compounds by introduction of novel starting materials. Here we have altered tryptamine, which serves as the starting substrate for a variety of alkaloid biosynthetic pathways, by replacing the indole with one of four aza-indole isomers. We show that two aza-tryptamine substrates can be successfully incorporated into the products of the monoterpene indole alkaloid pathway in Catharanthus roseus. Use of unnatural heterocycles in precursor directed biosynthesis, in both microbial and plant natural product pathways, has not been widely demonstrated, and successful incorporation of starting substrate analogs containing the aza-indole functionality has not been previously reported. This work serves as a starting point to explore fermentation of aza-alkaloids from other tryptophan and tryptamine derived natural product pathways.
Aza-indoles are important pharmacophores that mimic the indole scaffold. These unique heterocycles are isosteric to the indole moiety, but display improved water solubility and unique hydrogen bonding properties (Merour and Joseph, 2001). Several natural products contain aza-indole moieties (Perry et al., 1994; Schumacher and Davidson, 1995; Trimurtulu et al., 1994), and the anti-tumor properties of a synthetic aza analog of the indole natural product rebeccamycin have also been examined (Marminon et al., 2003) (Figure 1). Moreover, a number of synthetic drug candidates contain the aza-indole scaffold (Leblanc et al., 2009; Tsai et al., 2008; Wang et al., 2003) (Figure 1). We synthesized aza-tryptamines 1–4 and demonstrated that two of these compounds, 1 and 4, could serve as surrogates for tryptamine in the monoterpene indole alkaloid biosynthetic pathway to generate aza-alkaloids (Figure 2). Notably, these two deactivated aza-indole substrates were successfully utilized in an enzymatic Pictet-Spengler reaction, a key step in this biosynthetic pathway.
Figure 1.

The aza-indole moiety (highlighted in red) in natural and synthetic compounds. A. Aza-indoles in natural products. B. Aza-indole moiety in an indole natural product analog. C. Synthetic pharmaceutically important compounds containing an aza-indole group.
Figure 2.

Synthesis of aza-tryptamine compounds 1–4.
A general synthetic procedure for the production of 4-, 5-, 6-, and 7- aza-tryptamine 1–4 was developed. Ortho-iodoanilines, which were either commercially available or were prepared by copper-catalyzed halogen exchange reaction using the diamine ligand trans-N,N′-dimethylcyclohexane (Klapars and Buchwald, 2002) (Figure 2), could be subjected to palladium-catalyzed heteroannulation with a trimethylsiliated alkyne, followed by the acid hydrolysis to give all four 3-substituted aza-indole products in good yields (Pullagurla et al., 2005; Ujjainwalla and Warner, 1998) (Figure 2).
We investigated the use of aza-tryptamines in a Pictet-Spengler reaction, which is widely used in synthetic organic chemistry (Royer et al., 2004), and also serves as the first committed enzymatic step in a number of alkaloid biosynthetic pathways (O’Connor and Maresh, 2006) (Figure 3). In the Pictet-Spengler reaction, tryptamine 5 reacts with an aldehyde to form an iminium ion. The nucleophilic indole then attacks the iminium ion, resulting in the formation of a tetrahydro-β-carboline product (Figure 3) (Maresh et al., 2008). Since aza-tryptamines are more electron deficient than tryptamine 5, the efficiency of the Pictet-Spengler reaction is expected to be greatly decreased with substrates 1–4. However, when incubated with both the aldehyde secologanin 6 and the “Pictet-Spenglerase” strictosidine synthase from the monoterpene indole alkaloid biosynthetic pathway in Catharanthus roseus, 4- and 7-aza-tryptamines 1 and 4 were converted into the corresponding aza-tetrahydro-β-carboline products 7 and 10. No product formation was observed with 5- and 6-aza-tryptamines 2 and 3. Overall, the catalytic efficiency of 7-aza-tryptamine 4 (kcat: 11.8 ± 0.4 min−1 and KM: 65 μM ± 9.1) was approximately 100 fold lower compared to tryptamine 5 (kcat: 96 ± 4.7 min−1 and KM: 5 μM ± 1.4), and 4-aza-tryptamine 1 (kcat: 10.2 ± 0.3 min−1 and a KM: 151 μM ± 9.3) was 280 fold reduced relative to tryptamine 5.
Figure 3.

Figure 3A. Reaction of aza-tryptamine compounds 1–4 in a Pictet-Spengler reaction. B. Formation of downstream alkaloid and aza-alkaloid products.
Strictosidine synthase, in general, has been shown to display a greater tolerance for substituents at the 4 and 7 positions of the indole ring compared to substituents at the 5 and 6 positions (Figure 3) (Bernhardt et al., 2007; Loris et al., 2007; McCoy et al., 2006), a pattern consistent with the reactivity observed with aza-tryptamines 1–4. Although steric factors did not seem likely to play a major role in the recognition of the aza-tryptamines–the size of nitrogen (van der waals radius 1.55 Å) is similar to carbon (1.70 Å)–hydrogen bonding properties of the aza-indoles could affect whether the substrate interacts productively with the enzyme. Given that 7-aza-tryptamine 4 is the best substrate of the series, it is notable that the 7-position of the indole ring is pointed toward the solvent, away from any protein contact, while 4, 5 and 6 positions are located in the interior of the protein (Ma et al., 2006).
We asked whether the differing electronic properties of 1–4 also played a role in the relative rates of enzymatic reaction, since the Pictet-Spengler reaction depends upon the electron density of the aromatic amine substrate. After formation of the iminium ion, the Pictet-Spengler reaction can proceed by one of two mechanisms. In one mechanism, the C2 carbon can attack the iminium moiety to form the cyclized intermediate shown in Figure 3. Alternatively, the C3 carbon can attack the iminium moiety to form a spiro intermediate, which then undergoes a 1,2 shift to rearrange to the cyclized intermediate shown in Figure 3. Computational evidence suggests that C2, rather than C3, specifically acts as the nucleophile that attacks the iminium moiety, and that the spiro intermediate is not part of the productive reaction mechanism (Figure 3) (Kowalski et al., 1995; Maresh et al., 2008). Therefore, it is the electron density of the C2 position that should have the most impact upon the relative reactivity of 1–4. Since electron density can be inferred from 13C chemical shifts (Lauterbur, 1961), 1H-13C-HSQC spectroscopy was used to measure the chemical shifts of the C2 atom in 1–4. These measurements indicated that the C2 position of 7-aza-tryptamine 4 (115.9 ppm) is the most electron rich, followed by 5-aza-tryptamine 2 (131.0 ppm), then 4-aza-tryptamine 1 (135.2 ppm) and lastly 6-aza-tryptamine 3 (137.5 ppm). While these data are consistent with previous reports that 4- and 6-aza-indoles are more deactivated than 5- and 7-aza-indoles (Zhang et al., 2002), the electronic density of these aza-indole substrates do not correspond to the order of enzyme reactivity, where 7-aza-tryptamine 4 > 4-aza-tryptamine 1 ≫> 5-aza-tryptamine 2 ~ 6-aza-tryptamine 3. Therefore, we conclude that the electron density of the C2 atom of the aza-tryptamines is not the dominating factor in enzymatic reactivity.
It is also known that the pyridine moieties of 5- and 6-aza-indoles are more basic than 4- and 7-aza-indoles (7-aza-indole pKa: 4.6, 4-aza-indole pKa: 6.9, 6-aza-indole pKa; 7.9, 5-aza-indole pKa: 8.3 (Adler and Albert, 1963)). Therefore, 5- and 6-aza-tryptamines 2 and 3 are more likely to be protonated at neutral pH. Protonation of the pyridine would reduce the electron density of the aza-indole, and the rate of the Picet-Spengler reaction would be further diminished. However, although strictosidine synthase is active at pH 8 (Maresh et al., 2008), both 2 and 3 still failed to form product when incubated with strictosidine synthase and 6 at this more basic pH value. Additionally, the tryptamine-binding pocket of strictosidine synthase consists primarily of hydrophobic residues (Ma et al., 2006), suggesting that an amino acid of the enzyme does not protonate the pyridine ring of any of the aza-tryptamines. Therefore, we conclude that neither the higher pKa values of 5- and 6-aza-tryptamines, nor the corresponding protonation state of the pyridine moiety, are responsible for the lack of enzymatic reactivity with 2 and 3. Instead, the reactivity is consistent with the previously reported substrate specificity of strictosidine synthase, in which tryptamine analogs with a variety of substituents at the 4 and 7 positions are accepted, while most substrates substituted at the 5 and 6 positions are not (Bernhardt et al., 2007; Loris et al., 2007; McCoy et al., 2006). Importantly, the aza group has different hydrogen bonding properties and is much more hydrophilic than the corresponding carbocycle. In addition to discriminating against 5- and 6 substituted tryptamines for steric reasons, strictosidine synthase also discriminates against these isosteric changes.
It is notable that strictosidine synthase allows turnover of chemically deactivated substrates. We speculate that for successful turnover of 1 and 4, the enzyme uses the active site glutamic acid residue to keep the iminium ion nitrogen protonated (Maresh et al., 2008) (Figure 3), and therefore electrophilic, while at the same time ensuring that the pyridine moiety of the aza-indole remains neutral. This highlights the superb capacity of enzymes to catalyze reactions by selectively protonating spatially separated functional groups.
The enzymatically active substrates, 4- and 7-aza-tryptamine 1 and 4, were chosen for further analysis in this biosynthetic pathway, in the steps beyond the Pictet-Spengler reaction catalyzed by strictosidine synthase (Figure 3). To examine whether the aza-strictosidine analogs 7 and 10 formed by strictosidine synthase could be subjected to further enzymatic transformations, these compounds were incubated with Catharanthus roseus cell culture that is known to produce a variety of monoterpene indole alkaloids derived from strictosidine 11 (O’Connor and Maresh, 2006). Co-culture of C. roseus with varying concentrations of 1 and 4, or the corresponding strictosidine synthase products, aza-strictosidines 7 and 10, did not result in the formation of any new alkaloid compounds as evidenced by analysis of the cell culture extracts by LC-MS. However, when 4- and 7-aza-strictosidine 7 and 10 were deglycosylated in vitro by the strictosidine glucosidase enzyme that acts directly downstream from strictosidine synthase, and then co-cultured with C. roseus, we observed a variety of new compounds that were not present in control cultures lacking aza-indole starting substrates (Figure 3 and Figure 4). Monoterpene indole alkaloid biosynthesis is a highly branched pathway in C. roseus, with over 100 compounds arising from the deglycosylated strictosidine intermediate. The molecular weight of the new compounds formed appeared to correlate with aza analogs of some these known alkaloids.
Figure 4.
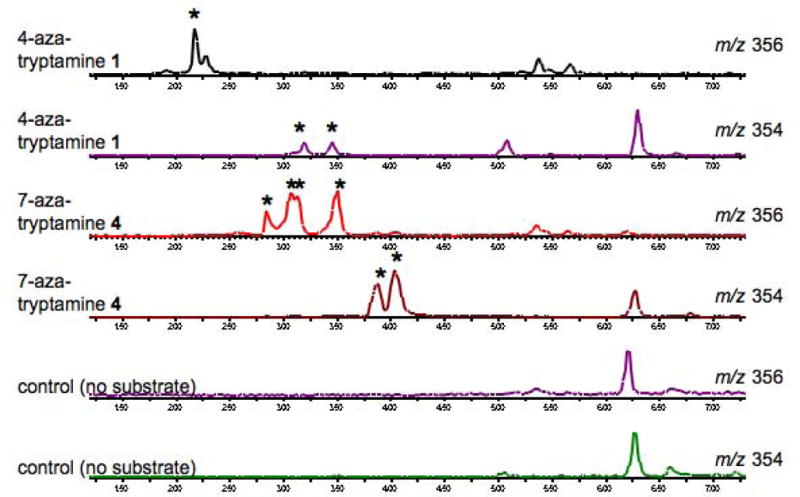
LC-MS spectra showing production of new compounds by Catharanthus roseus when supplemented with deglycosylated strictosidine derived from 1 and 4. Starred peaks indicate compounds found only in cultures supplemented with deglycosylated strictosidine derived from 1 or 4. LC-MS spectra showing the results of feeding deglycosylated strictosidine 11 derived from natural tryptamine 5 to cell cultures are shown in the Supporting Information.
Several compounds with m/z 356 and m/z 354 derived from 4-aza-tryptamine 1 and 7-aza-tryptamine 4 were observed (Figure 4). Natural tryptamine derived compounds with m/z 353 (corresponding to m/z 354 for the aza-tryptamine analog) in these cell lines are heteroyohimbine alkaloids, with ajmalicine 12 as the major compound. Additionally, there are several known sitsirikine type alkaloids 13 with m/z 355 (corresponding to m/z 356 for the aza-indole analog) that have been previously isolated from C. roseus. One of the most abundant unnatural products (m/z 356, HPLC retention time at 3.6 min), which was derived from 7-aza-tryptamine 4, was isolated and characterized by 1H-NMR and g-COSY. The 1H-NMR spectrum of the isolated compound most closely matched that of isositsirikine. Notably, isositsirikine isomers have been shown to exhibit anti-neoplastic activity (Mukhopadhyay et al., 1983). NMR analysis of all isositsirikine stereoisomers have been previously reported (Lounasmaa et al., 1994). The NMR data for the isolated aza compound 14 was most consistent with the NMR data reported for (3α, 16R, 19E)- isositsirikine (Lounasmaa et al., 1994) (Figure 3). (3α, 16R, 19E)- isositsirikine has been previously isolated from C. roseus cell culture, though typically in small amounts. Therefore, while the natural isositsirikine alkaloid is produced in low yields compared to other monoterpene indole alkaloid classes, the aza-tryptamine substrate 4 preferentially partitions into this pathway branch.
No compounds with molecular weights consistent with aza-analogs of catharanthine 15 or tabersonine 16 (m/z 337 for parent, m/z 338 for aza analog) or akuammicine 17 (m/z 323 for parent, m/z 324 for aza analog), the other major alkaloids normally observed in C. roseus culture, were observed (Figure 3). While heteroyohimbine type alkaloids such as ajmalicine 12 and isositisirikine 13 result from enzymatic reduction of deglycosylated strictosidine (Bernhardt et al., 2009; Hemscheidt and Zenk, 1985; Stockigt et al., 1983), formation of both catharanthine 15, tabersonine 16 and akuammicine 17 require a complex rearrangement of the deglycosylated strictosidine intermediate (Qureshi and Scott, 1968; Scott et al., 1971). Although the mechanism of this rearrangement remains poorly understood, it clearly involves the C2 and C3 positions of the indole moiety. The diminished electron density of the aza-tryptamine substrates 1 and 4 do not appear to be compatible with this rearrangement. In contrast, formation of alkaloid products such as 12 and 13 entail a reduction reaction that does not involve participation of the indole group, enabling the biosynthesis of milligram quantities of aza-heteroyohimbine products.
Significance
By introducing unnatural starting materials to a producing organism, biosynthetic pathways can be hijacked to yield novel compounds. Here we have altered tryptamine 5, which serves as the starting substrate for a variety of alkaloid biosynthetic pathways, by replacing the indole with one of four aza-indole isomers. We show that two aza-tryptamine substrates can be successfully incorporated into the products of the monoterpene indole alkaloid pathway in Catharanthus roseus. A key step of this alkaloid biosynthetic pathway is an enzymatic Pictet-Spengler reaction that converts tryptamine 5 and secologanin 6 into strictosidine 11, a tetrahydro-β-carboline product. Two of the electron deficient aza-tryptamines, 1 and 4, were able to undergo this enzymatic reaction, resulting in the formation of pyrrolodipyridines 7 and 10, the aza analogs of tetrahydro-β-carbolines. These pyrrolodipyridines could then be transformed by C. roseus into heteroyohimbine type alkaloids. Use of unnatural heterocycles in precursor directed biosynthesis, in both microbial and plant natural product pathways, has not been widely demonstrated, and successful incorporation of starting substrate analogs containing the aza-indole functionality has not, to the best of our knowledge, been previously reported. The most abundant of the aza-alkaloids produced in C. roseus culture was isolated, characterized and shown to be an analog of isositsirikine, a natural product that has demonstrated anti-neoplastic activity. Normally, the level of isositsirikine in C. roseus culture is low; however, aza-tryptamine substrate 4 appeared to form this particular product preferentially compared to other classes of monoterpene alkaloids. Future efforts will focus on evaluation of the bioactivity of isolated aza-analogs. Additionally, this work serves as a starting point to explore fermentation of aza-alkaloids from other tryptophan and tryptamine derived natural product pathways.
Experimental
Characterization of compounds 1–4
Compound 1
1H NMR (500 MHz, D2O) δ 8.40 (d, J=9.0 Hz, 1H), 8.37 (d, J=6.0 Hz, 1H),7.93 (s, 1H), 7.56 (d, J=7.5 Hz, 1H), 7.63 (d, J=8.0 Hz, 1H), 3.33 (t, J=7.0 Hz, 2H), 3.19 (d, J=7.2 Hz, 2H. 13C NMR (125 MHz, D2O) δ 135.2, 134.5, 133.6, 132.7, 128.4, 116.7, 105.0. Exact mass (ESI-TOF): m/z calc (M+H): 162.1026, found: 162.1032.
Compound 2
1H NMR (500 MHz, D2O) δ 8.94 (s, 1H), 8.16 (d, J=6.5 Hz, 1H), 7.74 (dd, J=6.5 Hz, 1H), 7.60 (s, 1H), 3.30 (t, J=7.2 Hz, 2H), 3.16 (t, J=7.2 Hz, 2H). 13C NMR (125 MHz, D2O) δ 143.1, 134.4, 131.0, 130.9, 123.9, 113.6, 109.8, 40.0, 22.7. Exact mass (ESI-TOF): m/z calc (M+H): 162.1026, found: 162.1033.
Compound 3
1H NMR (500 MHz, D2O) δ 8.79 (s, 1H), 8.05 (d, J=6.5 Hz, 1H), 7.96 (s, 1H), 7.92 (d, J=6.5 Hz, 1H), 3.30 (t, J=7.2 Hz, 2H), 3.14 (d, J=7.2 Hz, 2H). 13C NMR (125MHz, D2O) δ 137.5, 136.2, 131.1, 127.5, 126.7, 115.3, 112.0, 39.7, 21.9. Exact mass (ESI-TOF): m/z calc (M+H): 162.1026, found: 162.1030.
Compound 4
1H NMR (500 MHz, D2O) δ 8.53 (d, J=8.0 Hz, 1H), 8.23 (d, J=5.5 Hz, 1H), 7.49 (s, 1H), 7.44 (t, J=6,8 Hz, 1H), 3.26 (t, J=6.8 Hz, 2H), 3.09 (t, J=7.0 Hz, 2H), 13C NMR (125MHz, D2O) δ 138.8, 137.1, 133.1, 128.2, 125.9, 115.9, 111.9. Exact mass (ESI-TOF): m/z calc (M+H): 162.1026, found: 162.1024.
Enzyme assays
Strictosidine synthase (C. roseus) was expressed in E. coli and purified as previously described (Bernhardt et al., 2009; Maresh et al., 2008; McCoy et al., 2006). Activity was measured using an HPLC assay following previously reported procedures (Maresh et al., 2008). Secologanin (2.5 mM), 1-naphthalene acetic acid as an internal standard (60 μM) and tryptamine analogs (1 μM – 500 μM) in PBS buffer (50 mM, pH 7.0) were pre-incubated at 30°C. Reactions were initiated by addition of strictosidine synthase (1 nM – 200 nM). The concentration of 1 and 4 was calculated from the extinction coefficients (1: ε = 6,006 M−1cm−1, 4: ε = 7,900 M−1cm−1, 290 nm, PBS buffer (50 mM) pH 7.0). Enzymatic reactions were quenched by addition of 2.0 M aqueous sodium hydroxide. Quenched aliquots of the reaction were directly injected onto an analytical HPLC using a solvent gradient of 10% to 70% acetonitrile in 0.1% aqueous trifluoroacetic acid. The absorbance of tryptamine analogs, strictosidine analogs and NAA were measured at 280 and 290nm. Peak areas were integrated and normalized to the internal standard. Initial rates were determined from the slope by linear least-squares fit of four normalized data points for each concentration of varied substrate. Kinetic parameters Vmax and Km were estimated from the initial rates directly fit to appropriate forms of the Michaelis-Menten equation by nonlinear least squares fit using Origin 6.0 software (OriginLab Corp.)
Feeding studies with 1 and 4
To a solution of strictosidine synthase and secologanin (10 mg, 25.8 mmol) in 50 mM PBS buffer (3 ml, pH 7.0) was added 4- or 7-aza-tryptamine 1 or 4 (4.6 mg, 20 mmol) in 50 mM PBS buffer (1 ml, pH 7.0). The reaction mixture was incubated at 30°C for 48 hours. Stricto sidine glucosidase was then added and the mixture was allowed to incubate for an additional 48 hours. The solution was evaporated, dissolved in 150 μl ethanol and added to C. roseus cell suspension culture (150 ml) that had been subcultured for 10 days. The cultures were allowed to incubate for an additional 7 days, at which point the cells were filtered and ground in methanol. The methanol extract was dried, redissolved in ethyl acetate and washed with water. The organic layer was concentrated, dissolved in methanol and subjected to LC-MS analysis. UPLC analysis was performed on an Acquity Ultra Performance BEH C18 column with a 1.7 μm particle size, 2.1 × 100 mm dimension, and a flow rate of 0.6 mL min−1 in tandem with a Micromass LCT Premier TOF Mass Spectrometer with an ESI source (Waters Corporation). The capillary and sample cone voltages were 3000 V and 30 V, respectively. The desolvation and source temperatures were 300 and 100 °C, respectively. The cone and desolvation gas flow rates were 60 and 800 L h−1. Analysis was performed with MassLynx 4.1. Compound 14 was purified from reverse phase preparative HPLC using a gradient of 0–30% acetonitrile in water with 0.1% trifluoroacetic acid over 40 minutes.
7-Aza-isositsirikine (3α, 16R, 19E) 14
1H NMR (600 MHz, CD3OD) δ 8.25 (d, J=3.7 Hz, 1H), 8.00 (d, J=7.7 Hz, 1H), 7.17 (dd, J=7.7 Hz, 4.9 Hz, 1H), 6.04 (q, J=6.7 Hz, 1H), 3.88 (m, 3H), 3.78 (s, 3H), 3.70 (m, 3H), 3.56 (m, 2H), 3.19 (m, 1H), 3.06 (m, 1H), 2.62 (m, 2H), 2.46 (m, 2H). Exact mass (ESI-TOF): m/z calc (M+H): 356.1969, found: 356.1974.
Supplementary Material
Acknowledgments
We acknowledge support from GM074820.
Footnotes
Supporting Information: Additional synthetic procedures, enzyme assay conditions, and additional spectroscopic data are included in the Supporting Information.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Adler TK, Albert A. The biological and physical properties of the azaindoles. J Med Chem. 1963;6:480–483. doi: 10.1021/jm00341a003. [DOI] [PubMed] [Google Scholar]
- Bernhardt P, McCoy E, O’Connor SE. Rapid identification of enzyme variants for reengineered alkaloid biosynthesis in periwinkle. Chem Biol. 2007;14:888–897. doi: 10.1016/j.chembiol.2007.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernhardt P, Yerkes N, O’Connor SE. Bypassing stereoselectivity in the early steps of alkaloid biosynthesis. Org Biomol Chem. 2009;7:4166–4168. doi: 10.1039/b916027m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hemscheidt T, Zenk MH. Partial purification and characterization of a NADPH dependent tetrahydroalstonine synthase from Catharanthus roseus cell suspension cultures. Plant Cell Rep. 1985;4:216–219. doi: 10.1007/BF00269293. [DOI] [PubMed] [Google Scholar]
- Klapars A, Buchwald SL. Copper-catalyzed halogen exchange in aryl halides: An aromatic Finkelstein reaction. J Am Chem Soc. 2002;124:14844–14845. doi: 10.1021/ja028865v. [DOI] [PubMed] [Google Scholar]
- Kowalski P, Bojarski AJ, Mokrosz JL. Structure and spectral properties of β-carbolines. 8 Mechanism of the Pictet-Spengler cyclization: an MNDO approach. Tetrahedron. 1995;51:2737–2742. [Google Scholar]
- Lauterbur PC. C13 nuclear magnetic resonance spectroscopy (NMR). I Aromatic hydrocarbons. J Am Chem Soc. 1961;83:1838–1846. [Google Scholar]
- Leblanc Y, Roy P, Dufresne C, Lachance N, Wang ZY, O’Neill G, Greig G, Denis D, Mathieu MC, Slipetz D, et al. Discovery of potent and selective DP1 receptor antagonists in the azaindole series. Bioorg Med Chem Lett. 2009;19:2125–2128. doi: 10.1016/j.bmcl.2009.03.010. [DOI] [PubMed] [Google Scholar]
- Loris EA, Panjikar S, Ruppert M, Barleben L, Unger M, Schubel H, Stockigt J. Structure-based engineering of strictosidine synthase: Auxiliary for alkaloid libraries. Chem Biol. 2007;14:979–985. doi: 10.1016/j.chembiol.2007.08.009. [DOI] [PubMed] [Google Scholar]
- Lounasmaa M, Jokela R, Hanhinen P, Miettinen J, Salo J. Preparation and conformational study of Z-isositsirikine and E-isositsirikine epimers and model compounds - Determination of their C-16 configurations. Tetrahedron. 1994;50:9207–9222. [Google Scholar]
- Ma X, Panjikar S, Koepke J, Loris E, Stockigt J. The structure of Rauvolfia serpentina strictosidine synthase is a novel six-bladed beta-propeller fold in plant proteins. Plant Cell. 2006;18:907–920. doi: 10.1105/tpc.105.038018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maresh JJ, Giddings LA, Friedrich A, Loris EA, Panjikar S, Trout BL, Stöckigt J, Peters B, O’Connor SE. Strictosidine synthase: mechanism of a Pictet-Spengler catalyzing enzyme. J Am Chem Soc. 2008;130:710–723. doi: 10.1021/ja077190z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marminon C, Pierre A, Pfeiffer B, Perez V, Leonce S, Joubert A, Bailly C, Renard P, Hickman J, Prudhomme M. Syntheses and antiproliferative activities of 7-azarebeccamycin analogues bearing one 7-azaindole moiety. J Med Chem. 2003;46:609–622. doi: 10.1021/jm0210055. [DOI] [PubMed] [Google Scholar]
- McCoy E, Galan MC, O’Connor SE. Substrate specificity of strictosidine synthase. Bioorg Med Chem Lett. 2006;16:2475–2478. doi: 10.1016/j.bmcl.2006.01.098. [DOI] [PubMed] [Google Scholar]
- Merour JY, Joseph B. Synthesis and reactivity of 7-azaindoles (1H-pyrrolo[2,3-b]pyridine) Curr Org Chem. 2001;5:471–506. [Google Scholar]
- Mukhopadhyay S, El-Sayed A, Handy GA, Cordell GA. Catharanthus alkaloids XXXVII. 16-Epi-Z-isositsirikine, a monomeric indole alkaloid with antineoplastic activity from Catharanthus roseus and Rhazya stricta. J Nat Prod. 1983;46:409–413. doi: 10.1021/np50027a019. [DOI] [PubMed] [Google Scholar]
- O’Connor SE, Maresh J. Chemistry and biology of monoterpene indole alkaloid biosynthesis. Nat Prod Rep. 2006;23:532–547. doi: 10.1039/b512615k. [DOI] [PubMed] [Google Scholar]
- Perry NB, Ettouati L, Litaudon M, Blunt JW, Munro MHG. Alkaloids from the antarctic sponge kirkpatrickia-varialosa. 1 Variolin-b, a new antitumor and antiviral compound. Tetrahedron. 1994;50:3987–3992. [Google Scholar]
- Pullagurla M, Dukat M, Roth BL, Setola V, Glennon RA. 5-Azatryptamine analogs as h5-HT6 serotonin receptor ligands. Med Chem Res. 2005:1–18. [Google Scholar]
- Qureshi AA, Scott AI. Interconversion of corynanthe, aspidosperma and iboga alkaloids a model for indole alkaloid biosynthesis. Chem Comm. 1968:945–946. [Google Scholar]
- Royer J, Bonin M, Micouin L. Chiral heterocycles by iminium ion cyclization. Chem Rev. 2004;104:2311–2352. doi: 10.1021/cr020083x. [DOI] [PubMed] [Google Scholar]
- Schumacher RW, Davidson BS. Didemnolines a-d, new n9-substituted beta-carbolines from the marine ascidian Didemnum sp. Tetrahedron. 1995;51:10125–10130. [Google Scholar]
- Scott AI, Reichardt PB, Slaytor MB, Sweeny JG. Mechanisms of indole alkaloid biosynthesis. Bioorg Chem. 1971;1:157–173. [Google Scholar]
- Stockigt J, Hemscheidt T, Hofle G, Heinstein P, Formacek V. Steric course of hydrogen transfer during enzymatic formation of 3α-heteroyohimbine alkaloids. Biochemistry. 1983;22:3448–3452. [Google Scholar]
- Trimurtulu G, Faulkner DJ, Perry NB, Ettouati L, Litaudon M, Blunt JW, Munro MHG, Jameson GB. Alkaloids from the antarctic sponge kirkpatrickia-varialosa. 2 variolin-a and n(3′)-methyl tetrahydrovariolin-b. Tetrahedron. 1994;50:3993–4000. [Google Scholar]
- Tsai J, Lee JT, Wang W, Zhang J, Cho H, Mamo S, Bremer R, Gillette S, Kong J, Haass NK, et al. Discovery of a selective inhibitor of oncogenic B-Raf kinase with potent antimelanoma activity. Proc Natl Acad Sci USA. 2008;105:3041–3046. doi: 10.1073/pnas.0711741105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ujjainwalla F, Warner D. Synthesis of 5-, 6- and 7-azaindoles via palladium-catalyzed heteroannulation of internal alkynes. Tetrahedron Lett. 1998;39:5355–5358. [Google Scholar]
- Wang T, Zhang ZX, Wallace OB, Deshpande M, Fang HQ, Yang Z, Zadjura LM, Tweedie DL, Huang S, Zhao F, et al. Discovery of 4-benzoyl-1-[(4-methoxy-1H-pyrrolo[2,3-b]lpyridin-3-yl)oxoacetyl]-2-(R)- methylpiperazine (BMS-378806): A novel HIV-1 attachment inhibitor that interferes with CD4-gp120 interactions. J Med Chem. 2003;46:4236–4239. doi: 10.1021/jm034082o. [DOI] [PubMed] [Google Scholar]
- Zhang ZX, Yang Z, Wong H, Zhu J, Meanwell NA, Kadow JF, Wang T. An effective procedure for the acylation of azaindoles at C-3. J Org Chem. 2002;67:6226–6227. doi: 10.1021/jo020135i. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.