

Abstract
Abscisic acid (ABA) mediates resistance to abiotic stress and controls developmental processes in plants. The group-A PP2Cs, of which ABI1 is the prototypical member, are protein phosphatases that play critical roles as negative regulators very early in ABA signal transduction. Because redundancy is thought to limit the genetic dissection of early ABA signalling, to identify redundant and early ABA signalling proteins, we pursued a proteomics approach. We generated YFP-tagged ABI1 Arabidopsis expression lines and identified in vivo ABI1-interacting proteins by mass-spectrometric analyses of ABI1 complexes. Known ABA signalling components were isolated including SnRK2 protein kinases. We confirm previous studies in yeast and now show that ABI1 interacts with the ABA-signalling kinases OST1, SnRK2.2 and SnRK2.3 in plants. Interestingly, the most robust in planta ABI1-interacting proteins in all LC-MS/MS experiments were nine of the 14 PYR/PYL/RCAR proteins, which were recently reported as ABA-binding signal transduction proteins, providing evidence for in vivo PYR/PYL/RCAR interactions with ABI1 in Arabidopsis. ABI1–PYR1 interaction was stimulated within 5 min of ABA treatment in Arabidopsis. Interestingly, in contrast, PYR1 and SnRK2.3 co-immunoprecipitated equally well in the presence and absence of ABA. To investigate the biological relevance of the PYR/PYLs, we analysed pyr1/pyl1/pyl2/pyl4 quadruple mutant plants and found strong insensitivities in ABA-induced stomatal closure and ABA-inhibition of stomatal opening. These findings demonstrate that ABI1 can interact with several PYR/PYL/RCAR family members in Arabidopsis, that PYR1–ABI1 interaction is rapidly stimulated by ABA in Arabidopsis and indicate new SnRK2 kinase-PYR/PYL/RCAR interactions in an emerging model for PYR/PYL/RCAR-mediated ABA signalling.
Keywords: abscisic acid, ABI1, ABI1 interacting proteins, proteome, PYR/PYL/RCAR
Introduction
The plant hormone abscisic acid (ABA) controls important abiotic stress-induced and developmental responses, including maintenance of seed dormancy, seed development, growth regulation and stomatal closure. Forward and reverse genetic analyses have led to the identification of many loci encoding ABA signalling components that function in ABA metabolism and in ABA signalling (Finkelstein et al., 2002; Nambara and Marion-Poll, 2005; Israelsson et al., 2006; Yamaguchi-Shinozaki and Shinozaki, 2006). However, genetic redundancy has hampered the identification of many important ABA signalling components. Furthermore, proteins that mediate ABA reception are a subject of present debate and insufficient evidence exists on the interactions of some of the candidate proteins with known ABA signalling components (McCourt and Creelman, 2008).
Protein phosphorylation and dephosphorylation events are important mediators of ABA signal transduction. ABA-activated SNF1-related protein kinases 2 (SnRK2s) and Ca2+ dependent protein kinases (CDPKs) are positive transducers of ABA signalling (Li et al., 2000; Mustilli et al., 2002; Yoshida et al., 2002; Mori et al., 2006; Fujii et al., 2007; Zhu et al., 2007). Furthermore calcineurin B-like proteins (CBLs) and CBL-binding protein kinases (CIPKs), encode another class of SNF1-related protein kinases that negatively affect ABA responses (Hrabak et al., 2003; Batistic and Kudla, 2009; Luan, 2009). The ABA-activated protein kinase OST1 (SnRK2.6/SnRK2E) functions as a positive regulator of ABA-induced stomatal closure (Mustilli et al., 2002; Yoshida et al., 2002). SnRK2.2 and SnRK2.3 are homologues of OST1 and are also activated by ABA and regulate ABA responses in seed germination, root growth and gene expression (Fujii et al., 2007). Recently, snrk2.2/2.3/2.6 triple mutants were shown to cause a strong ABA insensitive phenotype in seed germination, root growth and gene expression, suggesting that SnRK2.2, SnRK2.3 and OST1/SnRK2.6 have overlapping functions in ABA signalling (Fujii and Zhu, 2009; Nakashima et al., 2009).
The dominant ABA insensitive mutants abi1-1 and abi2-1 exhibit ABA insensitivity in seed germination and root growth responses (Koornneef et al., 1984). ABI1 and ABI2 encode type 2C protein phosphatases (PP2Cs) (Leung et al., 1994, 1997; Meyer et al., 1994; Rodriguez et al., 1998). The Arabidopsis genome includes more than 76 PP2C genes (Schweighofer et al., 2004; Xue et al., 2008). Signal transduction analysis in guard cells showed that abi1-1 impairs ABA signalling mechanisms including ABA activation of S-type anion channels (Pei et al., 1997), ABA-induced [Ca2+] cyt elevations (Allen et al., 1999), ABA-induced Ca2+ channel activation and ROS production (Murata et al., 2001) and ABA activation of the OST1 protein kinase in Arabidopsis (Mustilli et al., 2002). These findings led to the model that ABI1 PP2C functions very early in ABA signal transduction (Pei et al., 1997; Allen et al., 1999; Murata et al., 2001; Mustilli et al., 2002). These signal transduction studies prompted us to search for in vivo ABI1-interacting proteins via protein complex purifications in the present study to identify possible redundant early ABA signal transduction proteins.
Studies have shown that six of the nine Arabidopsis PP2Cs belonging to cluster A of the PP2Cs family (Schweighofer et al., 2004), ABI1, ABI2, HAB1, HAB2, AHG1 and PP2CA/AHG3 function as negative regulators of ABA signalling and have independent and overlapping functions (Gosti et al., 1999; Merlot et al., 2001; Leonhardt et al., 2004; Saez et al., 2004, 2006; Kuhn et al., 2006; Yoshida et al., 2006b; Nishimura et al., 2007; Rubio et al., 2009). Yeast two hybrid screens have identified substrates of ABI1, including the homeodomain transcriptional factor ATHB6 (Himmelbach et al., 2002), the protein kinases, CIPK15, CIPK20 and OST1 (Guo et al., 2002; Ohta et al., 2003; Yoshida et al., 2006a), gluthathione peroxidase3 (Miao et al., 2006) and a homologue of the yeast SWI/SNF chromatin-remodelling complex, SWI3B (Saez et al., 2008).
Recently two independent groups have reported that two Bet VI family proteins, PYR1 and RCAR1, interact with ABI1, ABI2 and HAB1 (Ma et al., 2009; Park et al., 2009). These studies have provided evidence that the recombinant protein PYR1 and the RCAR1-ABI2 protein complex, may function as ABA receptors in ABA signalling (Ma et al., 2009; Park et al., 2009). Two dimensional NMR analyses of PYR1 showed that ABA causes shifts in the conformation of PYR1 (Park et al., 2009). Furthermore, ABA stimulates the interaction of PYR1 with PP2Cs and this ABA-dependent interaction down regulates PP2C activity in vitro (Park et al., 2009). RCAR1 also negatively regulates PP2C activity after ABA treatment in vitro, though RCAR1 constitutively interacts with PP2Cs. However, whether PYR/PYL/RCAR proteins interact with ABI1 in Arabidopsis remains unknown.
Here, to identify unknown redundant early ABA signalling proteins in vivo, we pursued YFP–ABI1 complex affinity column-based purifications from Arabidopsis plants. Interestingly, we identified nine of the 14 PYR/PYL/RCAR family members as the most robustly co-purified proteins with ABI1. These data show ABI1 interaction with multiple members of PYR/PYL/RCAR family in Arabidopsis.
Here we further address the following emerging questions: (i) Does the ABI1 protein phosphatase interact with the PYR1 protein in vivo (ii) Does ABA affect this interaction in vivo and within which time frame? (iii) Does ABI1 interact with SnRK2.2, SnRK2.3 and OST1/SnRK2.6 in plants? (iv) Does PYR form complexes with these ABA signalling SnRK2 kinases and does ABA affect this interaction? and (v) Do PYR/PYL/RCAR function in ABA-induced stomatal closure and ABA inhibition of stomatal opening?
Results
Isolation of YFP–ABI1 over-expression plants
To assess further the ABA signalling cascade, we pursued experiments to identify ABI1-interacting proteins in Arabidopsis using affinity column-based protein complex purifications. We generated transgenic YFP–ABI1 and YFP Arabidopsis expression lines in an abi1-3 knockout mutant background (Saez et al., 2006). Five independent transgenic YFP–ABI1 expression lines were isolated. Western blot and fluorescence microscopy analyses showed that these Arabidopsis lines express the YFP–ABI1 and YFP proteins, respectively (Figure 1a,b). Interestingly, YFP–ABI1 expression plants showed growth and ABA insensitive phenotypes that were comparable to the dominant abi1-1 mutant. 5-week-old YFP–ABI1 plants were significantly smaller in size than control YFP expression plants (Figure 1b). Previous research has reported that ABI1–GFP over-expressing lines do not show any ABA response phenotypes compared with vector control lines (Moes et al., 2008). In our investigations, YFP–ABI1 fusion expression plants showed ABA insensitive phenotypes during seed germination, root growth and in stomatal responses compared with the control YFP expression plants confirming ABI1 function (Figure 1c–e).
Figure 1.
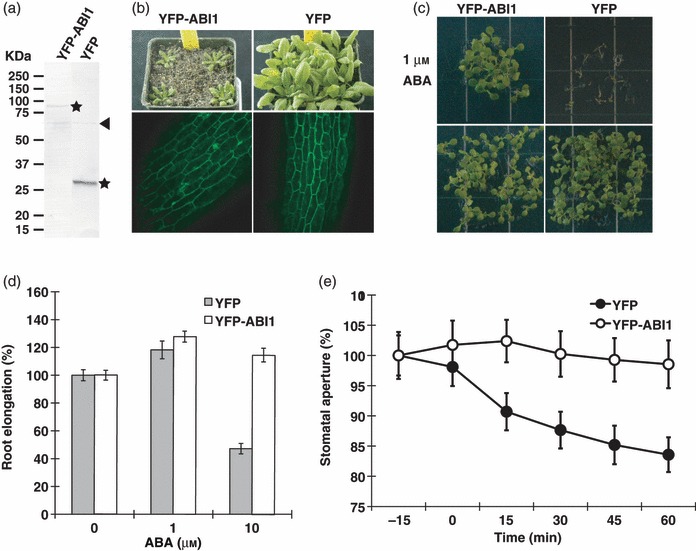
Constitutive expression of YFP–ABI1 causes ABA insensitivity.
(a) Western blot analysis of YFP-tagged ABI1 (left) and YFP control (right) proteins in transgenic Arabidopsis. Stars indicate predicted YFP–ABI1 and YFP bands. Arrow head: non-specific band.
(b) Morphology and subcellular localization of Arabidopsis plants constitutively expressing YFP–ABI1 (left) and YFP (right) at the rosette plant stage. Plants were grown for 5 weeks in soil.
(c) ABA-insensitive phenotype of constitutively expressed YFP–ABI1 (left) and ABA response in control YFP-expressing seedlings (right). Top: in the presence of 1 μm ABA. Bottom: without added ABA. Transgenic plant seeds were sown on agar MS plates with or without 1 μm exogenous ABA.
(d) ABA-dependent root growth responses of Arabidopsis seedlings constitutively expressing YFP and YFP–ABI1. Seedlings were germinated and grown on hormone-free MS plates for 5 days and then transferred to MS plates containing the indicated ABA concentrations. Root length was measured 4 days after transfer. Error bars show standard deviations.
(e) Time course experiments of ABA-induced stomatal closing in YFP control and YFP–ABI1 expressing leaves (genotype blind experiments). Stomatal apertures were individually mapped and images captured (Siegel et al., 2009) and measured before and after addition of 1 μm ABA. Average stomatal apertures at time −15 min: 3.47 ± 0.14 μm (YFP plants), 3.75 ± 0.18 μm (YFP–ABI1 plants). YFP plants: n = 23 stomata, YFP–ABI1 plants: n = 26 stomata. Error bars show SEM.
Identification of ABI1-interacting proteins
Using YFP–ABI1 and YFP expression in the abi1-3 knock-out background, we purified ABI1-interacting proteins. A GFP affinity column was loaded with whole protein extracts from YFP–ABI1 and control YFP expression plants grown on MS plates for 21 days with or without ABA treatment. Affinity purified protein complexes were identified by mass spectrometric analyses. The specificity of the proteins purified by YFP affinity purification was analysed in parallel negative control experiments using YFP expression plants in the abi1-3 mutant background (Tables S1 and S2). Upon silver staining, some visible bands overlapped with controls and specific bands associated that YFP–ABI1 samples were also consistently observed (Figure S1). Mass-spectrometrical analyses of five samples without ABA treatment (four independent samples and one duplicate) and five samples treated with ABA (three independent samples and two duplicates) allowed identification of proteins that associated with YFP–ABI1. Interestingly, the identified proteins included known ABA signalling components SnRK2.2, SnRK2.3, RPN10 and OST2/AHA1 (Tables 1, 2, S5 and S6) (Smalle et al., 2003; Fujii et al., 2007; Merlot et al., 2007). SnRK2.2 and SnRK2.3 were not identified in any of the YFP control experiments (Table S2).
Table 1.
Known ABA signalling proteins co-purified with ABI1 from Arabidopsis plants without addition of exogenous ABA
| Sequence coverage (%) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Symbol | Locus | AGI | Ex1 | Ex2 | Ex3 | Ex4 | Ex5 | Average |
| ABI1 | AT4G26080.1 | AT4G26080 | 90.6 | 87.1 | 83.2 | 87.1 | 87.6 | 87.1 ± 2.6 |
| SnRK2.2 | AT3G50500.1 | AT3G50500 | – | – | 22.4 | – | 13.5 | 18.0 ± 6.3 |
| SnRK2.3 | AT5G66880.1 | AT5G66880 | – | – | 8.9 | 12.5 | 16.6 | 12.7 ± 3.9 |
| RPN10 | AT4G38630.1 | AT4G38630 | 10.6 | – | 14.0 | 7.3 | 11.7 | 10.9 ± 2.8 |
| OST2 | AT2G18960.1 | AT2G18960 | 5.6 | – | 12.9 | 2.8 | – | 7.1 ± 5.2 |
Table 2.
Known ABA signalling proteins co-purified with ABI1 from Arabidopsis plants exposed to exogenous ABA
| Sequence coverage (%) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Symbol | Locus | AGI | Ex6 | Ex7 | Ex8 | Ex9 | Ex10 | Average |
| ABI1 | AT4G26080.1 | AT4G26080 | 63.4 | 85.5 | 89.4 | 84.6 | 82.3 | 81.0 ± 10.2 |
| SnRK2.2 | AT3G50500.1 | AT3G50500 | 13.5 | 16.6 | 26.0 | – | – | 18.7 ± 6.5 |
| SnRK2.3 | AT5G66880.1 | AT5G66880 | 13.6 | 11.4 | 19.9 | – | – | 15.0 ± 4.4 |
| RPN10 | AT4G38630.1 | AT4G38630 | 14.5 | 11.7 | 13.7 | 7.3 | – | 11.8 ± 3.2 |
| OST2 | AT2G18960.1 | AT2G18960 | – | 3.9 | 10.3 | – | – | 7.1 ± 4.5 |
Analyses of mass-spectra identified nine proteins in the absence of exogenously applied ABA that were identified in all YFP–ABI1 experiments, but not in YFP controls (Tables 3, S3, and S7). In the presence of exogenous ABA, eleven proteins were identified in all YFP–ABI1 samples (Tables 4, S4, and S8). Initially, we named these proteins candidate ABI1 interacting proteins (ABIP). The protein with the largest sequence coverage was ABI1 (Table 3). Six of the nine proteins in samples that were not treated with exogenous ABA encoded proteins from a, at the time uncharacterized, Bet VI subfamily of proteins (Tables 3 and S3). These Bet VI family proteins including PYR1 and other close homologues have recently been independently identified as candidate ABA-binding and ABA signalling proteins (Ma et al., 2009; Park et al., 2009). Interestingly the proteins co-purified with ABI1 with the highest total and unique sequence coverage and number of unique peptides and spectrum count were all members of the PYR/PYL/RCAR protein family (Figure 2a, Tables 3, S3 and S7). Two additional proteins from the 14 member PYR/PYL/RCAR protein family were identified in three or four of the five experiments (Table 3, PYL7 and PYL10). Moreover, in ABA-treated samples eight PYR/PYL/RCAR proteins had the largest total and unique sequence coverage and number of unique peptides and spectrum count of the isolated ABI1-interacting proteins and were identified in all five LC-MS/MS experiments (Tables 4, S4 and S8). As Bet VI proteins were the most significantly and consistently represented proteins from among all sequenced protein peptides in YFP–ABI1 samples, but not in YFP control samples (Table S2), these findings provide evidence that the ABI1 protein phosphatase can form complexes with several of the 14 PYR/PYL/RCAR Bet VI family members in vivo (Figure 2a, Tables 3, 4, S1, S3, S4, S7 and S8).
Table 3.
Candidate ABI1-interacting proteins with the largest mass-spectrometrical sequence coverage from five LC-MS/MS experiments analyzing ABI1 complexes isolated from Arabidopsis in the absence of exogenous ABA
| Sequence coverage (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Locus | AGI | Symbol | Ex1 | Ex2 | Ex3 | Ex4 | Ex5 | Average | |
| ABI1 | AT4G26080.1 | AT4G26080 | ABI1 | 90.6 | 87.1 | 83.2 | 87.1 | 87.6 | 87.1 ± 2.6 |
| ABIP1 | AT5G53160.2 | AT5G53160 | PYL8 | 78.2 | 63.3 | 79.8 | 61.2 | 64.9 | 69.5 ± 8.8 |
| ABIP2 | AT2G38310.1 | AT2G38310 | PYL4 | 87.9 | 72.5 | 55.1 | 40.6 | 62.8 | 63.8 ± 17.8 |
| ABIP3 | AT5G05440.1 | AT5G05440 | PYL5 | 74.4 | 63.5 | 71.9 | 35.5 | 57.1 | 60.5 ± 15.7 |
| ABIP4 | AT1G01360.1 | AT1G01360 | PYL9 | 67.9 | 50.3 | 44.9 | 62.6 | 59.9 | 57.1 ± 9.4 |
| ABIP5 | AT2G40330.1 | AT2G40330 | PYL6 | 57.7 | 22.3 | 48.4 | 28.4 | 34 | 38.2 ± 14.6 |
| ABIP6 | AT4G17870.1 | AT4G17870 | PYR1 | 39.8 | 19.4 | 36.1 | 28.8 | 48.2 | 34.5 ± 10.9 |
| ABIP11 | AT3G53430.1 | AT3G53430 | RPL12B | 28.3 | 15.1 | 31.3 | 5.4 | 5.4 | 17.1 ± 12.3 |
| ABIP12 | AT5G20280.1 | AT5G20280 | ATSPS1F | 9.2 | 4.9 | 9.4 | 5 | 4 | 6.5 ± 2.6 |
| ABIP7a | AT4G01026.1 | AT4G01026 | PYL7 | 46.9 | – | 37 | 25.1 | 26.1 | 33.8 ± 10.3 |
| ABIP8a | AT4G27920.1 | AT4G27920 | PYL10 | – | – | 15.3 | 7.7 | 7.7 | 10.2 ± 4.4 |
| ABIP9a | AT5G46790.1 | AT5G46790 | PYL1 | 22.6 | – | – | – | – | 22.6 ± 0.0 |
Three additional PYR/PYL/RCAR family members identified in 4, 3 and 1 of the experiments, respectively.
Table 4.
Candidate ABI1-interacting proteins with the largest mass-spectrometrical sequence coverage from five LC-MS/MS experiments analyzing Arabidopsis ABI1 complexes exposed to exogenous ABA
| Sequence coverage (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Locus | AGI | Symbol | Ex6 | Ex7 | Ex8 | Ex9 | Ex10 | Average | |
| ABI1 | AT4G26080.1 | AT4G26080 | ABI1 | 63.4 | 85.5 | 89.4 | 84.6 | 82.3 | 81.0 ± 10.2 |
| ABIP2 | AT2G38310.1 | AT2G38310 | PYL4 | 50.7 | 67.6 | 83.1 | 83.1 | 61.8 | 69.3 ± 14.0 |
| ABIP6 | AT4G17870.1 | AT4G17870 | PYR1 | 71.2 | 73.3 | 70.7 | 62.8 | 42.9 | 61.2 ± 12.5 |
| ABIP1 | AT5G53160.2 | AT5G53160 | PYL8 | 62.2 | 71.8 | 69.7 | 50 | 42 | 59.1 ± 12.8 |
| ABIP9 | AT5G46790.1 | AT5G46790 | PYL1 | 55.2 | 70.6 | 73.3 | 41.2 | 18.1 | 51.7 ± 22.8 |
| ABIP4 | AT1G01360.1 | AT1G01360 | PYL9 | 27.3 | 77 | 61.5 | 45.5 | 32.6 | 48.8 ± 20.6 |
| ABIP3 | AT5G05440.1 | AT5G05440 | PYL5 | 30 | 43.3 | 83.3 | 42.4 | 34.5 | 46.7 ± 21.2 |
| ABIP7 | AT4G01026.1 | AT4G01026 | PYL7 | 12.3 | 64.9 | 69.7 | 49.3 | 17.5 | 42.7 ± 26.6 |
| ABIP5 | AT2G40330.1 | AT2G40330 | PYL6 | 32.1 | 42.3 | 52.1 | 30.7 | 23.3 | 36.1 ± 11.2 |
| ABIP13 | AT3G19390.1 | AT3G19390 | MLD14.3 | 13.7 | 21.9 | 7.3 | 13.3 | 13.3 | 13.9 ± 5.2 |
| ABIP14 | AT2G20580.1 | AT2G20580 | AtRPN1a | 2.8 | 10.7 | 10.1 | 5.6 | 5.6 | 7.0 ± 3.3 |
| ABIP8a | AT4G27920.1 | AT4G27920 | PYL10 | – | 7.7 | – | – | – | 7.7 ± 0.0 |
PYL10 was identified in one experiment as shown.
Figure 2.
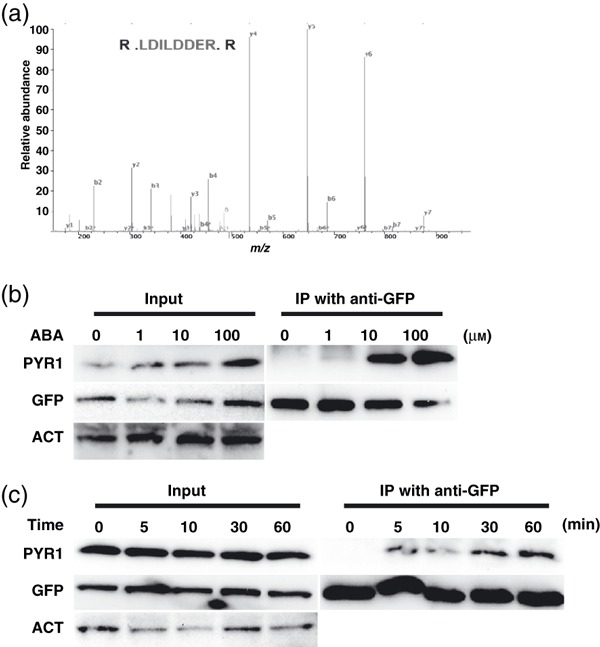
ABA causes ABI1–PYR1 interaction in Arabidopsis.
(a) Example of raw tandem mass spectrometry data identifying PYR1 peptide in YFP–ABI affinity purified samples. The predicted PYR1 peptide sequence is shown in the top inset.
(b) Increasing concentrations of exogenously applied ABA strongly enhanced co-immunoprecipitation of YFP–ABI1 with PYR1 in Arabidopsis. Transgenic Arabidopsis plants were exposed to ABA for 3 h.
(c) ABA triggers interaction of YFP–ABI1 with PYR1 within 5 min of exposure of Arabidopsis plants to exogenous ABA. Left: Total protein extracts (left, Input) from YFP–ABI1 plants. Plants were grown on MS plates for 3 weeks and were treated with or without 100 μm exogenous ABA. After co-immunoprecipitation using anti-GFP beads, input and co-immunoprecipitated samples were detected with anti-PYR1 and anti-GFP antibodies.
ABI1 interacts with PYR1 in Arabidopsis
It was recently shown that ABA enhances the interaction of YFP–ABI1 with HA–PYR1 transiently expressed in Nicotiana benthamiana leaves (Park et al., 2009). To assess whether these interactions occur in vivo in Arabidopsis, we performed co-immunoprecipitation experiments in stably transformed YFP–ABI1 and YFP control expression lines using a PYR1 antibody. YFP–ABI1 interacted with PYR1 in Arabidopsis with a strong enhancement upon application of 100 μm exogenous ABA for 48 h (Figure S2). Subsequently, co-immunoprecipitation experiments were performed three h after exogenous application of 0, 1, 10 and 100 μm ABA. ABA caused a clear enhancement in ABI1–PYR1 interaction in vivo (Figure 2b). To determine the timing of this ABA response in Arabidopsis, co-immunoprecipitation experiments were performed 0, 5, 10, 30 or 60 min after application of exogenous ABA. These experiments showed a clear ABA induction of a YFP–ABI1 and PYR1 interaction within 5 min of ABA treatment in vivo (Figure 2c).
PP2Cs, PYR1 and SnRK2s interact with each other in ABA signalling
ABI1 has been reported to interact with OST1/SnRK2E/SnRK2.6 in yeast (Yoshida et al., 2006a). However it remains unknown whether this interaction occurs in plant cells and whether ABI1 interacts with other SnRK2s. It also remains unknown whether any ABI1–SnRK2 interactions are regulated by ABA similar to the ABI1–PYR1 interaction. As SnRK2.2 and SnRK2.3 were co-purified with ABI1 in our affinity column purification experiments (Tables 1 and 2), we co-expressed YFP–ABI1 with either HA–SnRK2.2, HA–SnRK2.3 or HA–OST1/SnRK2.6 in N. benthamiana and performed co-immunoprecipitation experiments. YFP–ABI1 constitutively interacted with HA–SnRK2.2, HA–SnRK2.3 and HA–OST1/SnRK2.6 both in the absence and presence of exogenously applied ABA (Figure 3).
Figure 3.
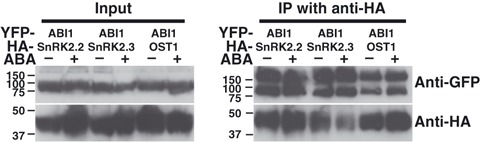
ABI1 PP2C co-immunoprecipitates with all three ABA signalling SnRK2 kinases in plants.
YFP–ABI1 co-immunoprecipitates with HA–SnRK2.2, HA–SnRK2.3 and HA– OST1/SnRK2.6 both in the presence and absence of exogenously applied ABA. Total protein extracts (left, Input) from transformed Nicotiana benthamiana leaves were harvested 3 days after inoculation and were treated with or without 100 μm ABA for 24 h before harvesting. After co-immunoprecipitation using an anti-HA matrix, input and immunoprecipitated samples were detected with anti-GFP and anti-HA antibodies.
We analysed whether PYR1 might co-purify with SnRK2s. We co-expressed YFP–PYR1 with either HA–SnRK2.3 or HA–OST1/SnRK2.6 in N. benthamiana and performed co-immunoprecipitation experiments. Interestingly, YFP–PYR1 co-immunoprecipitated with HA–SnRK2.3 both in the presence and absence of ABA, but not detectably or very weakly with HA–OST1/SnRK2.6 (Figure 4).
Figure 4.
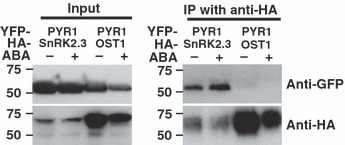
Interaction of PYR1 with SnRK2.3 and OST1.
YFP–PYR1 co-immunoprecipitates with HA–SnRK2.3. In contrast HA–OST1/SnRK2.6 did not co-immunoprecipitate with YFP–PYR1. Total protein extracts (left, Input) from transformed Nicotiana benthamiana leaves were harvested 3 days after inoculation and were treated with or without 100 μm ABA for 24 h before harvesting. After co-immunoprecipitation using an anti-HA matrix, input and immunoprecipitated samples were detected with anti-GFP and anti-HA antibodies.
pyr1/pyl1/pyl2/pyl4 quadruple mutant plants exhibit ABA insensitive phenotype in guard cells
The abi1-1 protein impairs ABA signalling upstream of known early ABA responses in Arabidopsis guard cells (Pei et al., 1997; Allen et al., 1999; Murata et al., 2001; Mustilli et al., 2002). A pyr1/pyl1/pyl2/pyl4 quadruple mutant was recently shown to cause strong ABA insensitivities in seed germination, root growth and ABA-induced gene expression (Park et al., 2009). Whether PYR/PYL/RCAR genes mediate ABA response in guard cells remains unknown. Public microarray data show that the PYR1, PYL1, PYL2, PYL4, PYL5, PYL7 and PYL8 mRNAs are highly expressed in guard cells and seedlings (Leonhardt et al., 2004; Goda et al., 2008; Yang et al., 2008) (Figure S4). Stomatal response analyses were performed with pyr1/pyl1/pyl2/pyl4 quadruple mutant plants. pyr1/pyl1/pyl2/pyl4 quadruple mutants showed a strong ABA insensitive phenotype in double-blinded ABA-induced stomatal closing and ABA inhibition of stomatal opening analyses (Figures 5a,b and S5). Studies have provided evidence that Ca2+ signalling functions downstream of the abi1-1 and abi2-1 PP2Cs in guard cells (Allen et al., 1999; Murata et al., 2001). Experiments were pursued to determine whether pyr1/pyl1/pyl2/pyl4 quadruple mutant plants impair Ca2+-induced stomatal closing. Repetitive addition and removal of Ca2+ to the extracellular solution bathing leaf epidermes while shifting the K+ equilibrium potential in guard cells, results in Ca2+-induced stomatal closure (Allen et al., 2001). Such imposed extracellular Ca2+ pulses caused similar Ca2+-reactive and Ca2+-programmed stomatal closure responses in both pyr1/pyl1/pyl2/pyl4 quadruple mutant and wild type epidermes (Figure 5c). These results suggest that these PYR/PYL/RCAR proteins are critical for ABA signalling in guard cells upstream of Ca2+ signalling.
Figure 5.
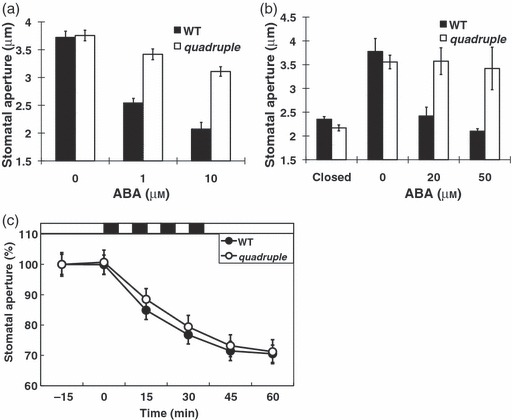
ABA-induced stomatal closure and ABA-inhibition of stomatal opening but not Ca2+ -induced stomatal closure are impaired in pyr1pyl1pyl2pyl4 quadruple mutant plants.
(a) ABA-induced stomatal closure in pyr1pyl1pyl2pyl4 quadruple mutant and wild type rosette leaves treated with the indicated ABA concentrations for 1 h (n = 4 experiments, 30 stomata per experiment and condition; genotype and [ABA] blind experiments).
(b) ABA inhibition of stomatal opening in pyr1pyl1pyl2pyl4 quadruple mutant and wild type abaxial leaf epidermes treated with the indicated ABA concentrations for 1 h (n = 3 experiments, 30 stomata per experiment and condition; genotype and [ABA] blind experiments). Closed means incubation in the dark for 3 h (before light and ABA treatment).
(c) Stomatal closure in response to four repetitive extracellular Ca2+ pulses (5 min each) is not impaired in pyr1pyl1pyl2pyl4 quadruple mutant suggesting that PYR/PYL/RCARs function upstream of Ca2+. Four 5-min extracellular applications of 1 mm CaCl2 and 1 mm KCl were sequentially applied to abaxial leaf epidermes (black bars at top) followed by 50 mm KCl and 0 added CaCl2 exposure (White bars at top) (n = 31 individually mapped stomata for both wild type and pyr1pyl1pyl2pyl4 quadruple mutant) Average stomatal apertures at time = −15 min: 5.24 ± 0.19 μm (wild type), 5.52 ± 0.14 μm (pyr1pyl1pyl2pyl4 quadruple mutant). Error bars show SEM.
Discussion
Studies have suggested that ABI1 functions upstream of all analysed events in early ABA signal transduction in guard cells (Pei et al., 1997; Allen et al., 1999; Murata et al., 2001; Mustilli et al., 2002). We hypothesized that redundant proteins may act upstream of ABI1, because forward genetic screens had not identified other upstream loci. Our early attempts to co-purify ABI1-complexed proteins from Arabidopsis, showed that additional purification steps led to loss of many ABI1-associated proteins (I. Mori & J. I. Schroeder unpublished data). A rapid purification method using GFP affinity column-affinity purification, as reported here, was more fruitful for isolating redundant ABA signalling proteins in vivo (Tables 1–4, S1–S4, S7, S8). Here we identify nine of the 14 PYR/PYL/RCAR gene family proteins as in vivo ABI1-co-purified proteins with the largest sequence coverage and number of unique peptides and spectrum count of all identified proteins (Tables 3, 4, S3, S4, S7 and S8; Figures 2 and 3). After identifying these Bet VI family proteins as the main co-purified proteins in ABI1 complexes from Arabidopsis, we learned that PYR1 had been independently isolated in a chemical genetics screen as a candidate ABA signalling protein (Park et al., 2009). In the absence of ABA, PYR1 and PYL4 proteins were co-purified as ABI1-interacting proteins (Tables 3, S3 and S7), although ABA enhanced the interaction of ABI1–PYR1 (Figure 2) and HAB1–PYL4 (Park et al., 2009). Co-purification results in the absence of exogenously applied ABA may be due to the known endogenous ABA levels in Arabidopsis plants (Nambara and Marion-Poll, 2005). On the other hand, ABI1 seems to interact constitutively with PYL5 to PYL9 in Arabidopsis, in the absence of ABA, based on yeast two hybrid data (Ma et al., 2009; Park et al., 2009). Over-expression of several other PP2Cs causes ABA insensitive phenotypes in seed germination stages (Saez et al., 2004; Kuhn et al., 2006; Yoshida et al., 2006b; Nishimura et al., 2007). YFP–ABI1 expression plants showed ABA insensitive phenotypes during seed germination, root growth and in stomatal responses compared with the control YFP expression plants (Figure 1c–e).
In addition to the identification of PYR/PYL/RCAR proteins as the major proteins co-purified with ABI1 from Arabidopsis plants, the present study reports several further relevant new findings: (i) A new PYR1 antibody was used and showed that the ABI1 protein phosphatase interacts with the PYR1 protein in vivo (ii) ABA stimulates this interaction in Arabidopsis within 5 min of ABA exposure. (iii) ABI1 co-immunoprecipitates with the SnRK2.2, SnRK2.3 and OST1/SnRK2.6 protein kinases in plants, extending previous yeast two hybrid findings (Yoshida et al., 2006a). (iv) PYR1 forms complexes with these ABA signalling SnRK2 kinases in plants both in the presence and absence of ABA. (v) PYR/PYL/RCAR genes are critical for ABA-induced stomatal closure and ABA inhibition of stomatal opening.
Protein phosphorylation and dephosphorylation events in ABA signalling
The protein kinase OST1/SnRK2.6 is phosphorylated within 5 min of ABA application (Mustilli et al., 2002; Yoshida et al., 2006a). ABA binds to PYR1 and RCAR1–ABI2 thus down-regulating PP2C activities (Ma et al., 2009; Park et al., 2009). In addition, PYR/PYL/RCAR genes are required for ABA activation of the SnRK2 kinases (Park et al., 2009). Here ABA is shown to cause PYR1–ABI1 interaction in Arabidopsis, whereas PYR1 forms a constitutive complex with the SnRK2.3 kinase both in the absence and presence of ABA (Figure 4). Whether PYR1 directly binds to SnRK2s remains to be determined. Furthermore, ABI1 constitutively interacts with the SnRK2.2, SnRK2.3 and OST1/SnRK2.6 kinases in planta (Figure 3). These co-immunoprecipitation results support the hypothesis that these major ABA signalling components, PYR/PYL/RCARs, PP2Cs and SnRK2s, may form an ABA signalosome complex, rather than forming a simple sequential PYR1→ABI1→SnRK2 signalling pathway. An initial sequential model (Park et al., 2009) would predict that SnRK2 kinases may only enter into a complex with PYR1 after ABA exposure.
One of the substrates of SnRK2s is a b-ZIP type transcription factor ABRE/ABF (Furihata et al., 2006; Fujii et al., 2007). Arabidopsis ost2 and slac1 mutants show strong ABA insensitive phenotypes in guard cells and act downstream of the OST1 kinase (Li et al., 2000; Merlot et al., 2007; Negi et al., 2008; Vahisalu et al., 2008). We isolated OST2/AHA1 as a candidate ABI1-interacting protein (Tables 1 and 2), which would be consistent with the model that the ABA signalling PP2Cs regulate multiple downstream signalling proteins (Pei et al., 1997; Allen et al., 1999; Murata et al., 2001; Furihata et al., 2006; Kuhn et al., 2006; Nishimura et al., 2007; Saez et al., 2008;).
PYR/PYL/RCAR family members regulate ABA signalling in guard cells
Several ABA receptors have been reported to impair ABA-induced stomatal closure; CHLH was reported as an ABA receptor but only one stable and available genetic allele cch (Mochizuki et al., 2001) showed a strong ABA insensitive phenotype (Shen et al., 2006). Recently, double mutants in the candidate ABA receptors, GTG1 and GTG2, were shown to cause ABA insensitivity in seed germination, root growth, stomatal movements and ABA-induced gene expression (Pandey et al., 2009). For ABA-induced stomatal closure, gtg1gtg2 double mutants exhibited a limited insensitivity to ABA compared with WT, suggesting that additional ABA receptors are needed to perceive ABA and transduce the ABA signal independently. More recently, the PYR/PYL/RCAR family proteins were identified as new ABA binding and signalling proteins (Ma et al., 2009; Park et al., 2009). Here we show that pyr1/pyl1/pyl2/pyl4 quadruple mutant plants exhibit a strong ABA insensitive phenotype in ABA-induced stomatal closing and ABA inhibition of stomatal opening but not in Ca2+-induced stomatal closing (Figure 5), suggesting that PYR1/PYL1/PYL2/PYL4 function upstream of Ca2+ in ABA signalling and likely also upstream of Ca2+-independent signalling (Allan et al., 1994; Siegel et al., 2009). Furthermore PYR1/PYL1/PYL2/PYL4 mediate ABA signalling not only in seed germination and seedling stages (Park et al., 2009) but also in guard cells (Figure 5).
In conclusion, in vivo affinity column purifications of ABI1 complex proteins provide evidence that the PYR/PYL/RCAR family of proteins are the most abundant and robust in vivo ABI1-complexed interactors in Arabidopsis. ABA rapidly stimulates PYR1–ABI1 interaction in Arabidopsis and additional findings indicate the possible assembly of an early ABA signalosome.
Experimental procedures
Plant materials and growth conditions
The Arabidopsis thaliana (L.) Heynh. ecotype Columbia (Col) was used in this study. Plant growth conditions, seed germination and root elongation assays were performed as described previously (Nishimura et al., 2004; Mori et al., 2006). ABA-induced and repetitive Ca2+ pulse-stomatal movement responses and apertures were analysed as described previously (Vahisalu et al., 2008). For ABA inhibition of stomatal opening, to pre-close stomata, intact leaf epidermes attached to a cover slip were pre-incubated in buffer (10 mm KCl, 7.5 mm iminodiacetic acid and 10 mm MES/Tris, pH 6.2) for 3 h in the dark and exposed to the indicated ABA concentrations in the light (100 μmol m−2 s−1).
Cloning and construction of transgenic YFP–ABI1 Arabidopsis lines
The full length ABI1 cDNA was cloned into pDONR vector (Invitrogen, http://www.invitrogen.com) and sequenced and then transferred to the destination vectors pH35YG and pEarleyGate 201 by Gateway LR recombination reaction (Invitrogen). Agrobacterium tumefaciens strain GV3101 was used to transform the abi1-3 T-DNA insertion disruption line and wild-type Col-0 with plants by the floral dip method (Clough and Bent, 1998). Transgenic plant lines were screened for hygromycin resistance and homozygous T3 lines were isolated.
Affinity column purification of YFP–ABI1 complexed proteins
Identification of proteins by mass spectrometry is described in the Experimental procedures S1.
Co-immunoprecipitation in Arabidopsis and Nicotiana benthamiana
For protein extractions, 3-week-old seedlings (1 g) grown on horizontal MS plates were incubated for 2 h in water before ABA treatment. Co-immunoprecipitations were performed using Dynabeads Protein G (Invitrogen) captured with anti-GFP antibody (Abcam, http://www.abcam.com) according to the manufacture’s specifications. Co-immunoprecipitations in N. benthamiana were performed as described previously without modifications (Park et al., 2009).
Acknowledgments
We thank Drs M. Böhmer and D.G. Mendoza-Cozatl for technical advice, Dr T.H. Kim for critical comments and reading of the manuscript and Drs I. Mori and M. Ghassemian (UCSD) for pursuing early abi1-1 and ABI1 complex purification attempts in the Schroeder laboratory. We are also grateful to the Arabidopsis Biological Resource Center (ABRC) and Dr T. Demura (NAIST) for the pH35YG plasmid. Requests for the pyr1pyl1pyl2pyl4 quadruple mutant seeds should be sent to Sean.cutler@ucr.edu. J.C. is an investigator of the Howard Hughes Medical Institute. This research was supported by NIH (GM060396-ES010337) and NSF (MCB0417118-MCB0918220) grants to J.I.S., NSF (IOS-0820508) to S.R.C., NIH (R01 GM052413) to J.C. and NIH (P41 RR011823) to J.R.Y.
Author contributions: N.N. performed experiments in Figures 1, 2, 3 and 4 and analysed mass spectrometry data. A.S. and J.R.Y. performed mass spectrometry and A.S., P.C., N.N. and J.I.S. analysed mass spectrometry data. K.N. and J.C. provided GFP antibody and assisted with affinity purifications. S.Y.P. and S.R.C. generated PYR1 antibody and pyr1pyl1pyl2pyl4 quadruple mutant seeds. A.W., S.L. and D.F.C. performed experiments in Figures 1(d,e) and 5. J.I.S. conceived of the project and J.I.S. and N.N. developed experimental designs. N.N. and J.I.S. wrote the paper.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online version of this article:
Figure S1. Identification of ABI1-interacting proteins in Arabidopsis.
Figure S2. ABA-induced interaction between ABI1 and PYR1 in Arabidopsis.
Figure S3. Tandem mass spectrum of peptides from proteins ABIP8 and ABIP11 listed in Tables S3 and S4 that showed one unique sequence to the corresponding proteins.
Figure S4. Transcription profiles of ABI1, PYR1/PYLs and SnRK2s in seedlings, guard cells and mesophyll cells.
Figure S5. ABA-induced stomatal closure isimpaired in pyr1pyl1-pyl2pyl4 quadruple mutant plants.
Table S1. List of candidate ABI1-interacting proteins in YFP–ABI1 plants derived from LC-MS/MS data.
Table S2. List of candidate YFP-interacting proteins in control YFP plants derived from LC-MS/MS data.
Table S3. The number of unique and total peptides and spectrum count of candidate ABI1-interacting proteins co-purified with ABI1 from Arabidopsis plants in the absence of exogenous ABA.
Table S4. The number of unique and total peptides and spectrum count of candidate ABI1-interacting proteins co-purified with ABI1 from plants exposed to exogenous ABA.
Table S5. The unique sequence coverage of known ABA signalling proteins co-purified with ABI1 from Arabidopsis plants without addition of exogenous ABA.
Table S6. The unique sequence coverage of known ABA signalling proteins co-purified with ABI1 from Arabidopsis plants exposed to exogenous ABA.
Table S7. The unique sequence coverage of candidate ABI1-interacting proteins co-purified with ABI1 from Arabidopsis plants in the absence of exogenous ABA.
Table S8. The unique sequence coverage of candidate ABI1-interacting proteins co-purified with ABI1 from plants exposed to exogenous ABA.
Experimental procedures S1.
Purification of YFP–ABI1 interacting proteins by affinity column purification methods.
Multidimensional protein identification technology (MudPIT).
Analysis of tandem mass spectra.
PYR1 expression and antiserum preparation.
Construction of the erecta+ pyr1/pyl1/pyl2/pyl4 quadruple mutant.
Please note: Wiley-Blackwell are not responsible for the content or functionality of any supporting materials supplied by the authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
References
- Allan AC, Fricker MD, Ward JL, Beale MH, Trewavas AJ. Two transduction pathways mediate rapid effects of abscisic acid in commelina guard cells. Plant Cell. 1994;6:1319–1328. doi: 10.1105/tpc.6.9.1319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen GJ, Kuchitsu K, Chu SP, Murata Y, Schroeder JI. Arabidopsis abi1-1 and abi2-1 phosphatase mutations reduce abscisic acid-induced cytoplasmic calcium rises in guard cells. Plant Cell. 1999;11:1785–1798. doi: 10.1105/tpc.11.9.1785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen GJ, Chu SP, Harrington CL, Schumacher K, Hoffmann T, Tang YY, Grill E, Schroeder JI. A defined range of guard cell calcium oscillation parameters encodes stomatal movements. Nature. 2001;411:1053–1067. doi: 10.1038/35082575. [DOI] [PubMed] [Google Scholar]
- Batistic O, Kudla J. Plant calcineurin B-like proteins and their interacting protein kinases. Biochim. Biophys. Acta. 2009;1793:985–992. doi: 10.1016/j.bbamcr.2008.10.006. [DOI] [PubMed] [Google Scholar]
- Clough SJ, Bent AF. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998;16:735–743. doi: 10.1046/j.1365-313x.1998.00343.x. [DOI] [PubMed] [Google Scholar]
- Finkelstein RR, Gampala SS, Rock CD. Abscisic acid signalling in seeds and seedlings. Plant Cell. 2002;14:S15–S45. doi: 10.1105/tpc.010441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujii H, Zhu JK. Arabidopsis mutant deficient in 3 abscisic acid-activated protein kinases reveals critical roles in growth, reproduction and stress. Proc. Natl Acad. Sci. USA. 2009;106:8380–8385. doi: 10.1073/pnas.0903144106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujii H, Verslues PE, Zhu JK. Identification of two protein kinases required for abscisic acid regulation of seed germination, root growth and gene expression in Arabidopsis. Plant Cell. 2007;19:485–494. doi: 10.1105/tpc.106.048538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furihata T, Maruyama K, Fujita Y, Umezawa T, Yoshida R, Shinozaki K, Yamaguchi-Shinozaki K. Abscisic acid-dependent multisite phosphorylation regulates the activity of a transcription activator AREB1. Proc. Natl Acad. Sci. USA. 2006;103:1988–1993. doi: 10.1073/pnas.0505667103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goda H, Sasaki E, Akiyama K, et al. The AtGenExpress hormone and chemical treatment data set: experimental design, data evaluation, model data analysis and data access. Plant J. 2008;55:526–542. doi: 10.1111/j.0960-7412.2008.03510.x. [DOI] [PubMed] [Google Scholar]
- Gosti F, Beaudoin N, Serizet C, Webb AA, Vartanian N, Giraudat J. ABI1 protein phosphatase 2C is a negative regulator of abscisic acid signalling. Plant Cell. 1999;11:1897–1910. doi: 10.1105/tpc.11.10.1897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo Y, Xiong L, Song CP, Gong D, Halfter U, Zhu JK. A calcium sensor and its interacting protein kinase are global regulators of abscisic acid signalling in Arabidopsis. Dev. Cell. 2002;3:233–244. doi: 10.1016/s1534-5807(02)00229-0. [DOI] [PubMed] [Google Scholar]
- Himmelbach A, Hoffmann T, Leube M, Hohener B, Grill E. Homeodomain protein ATHB6 is a target of the protein phosphatase ABI1 and regulates hormone responses in Arabidopsis. EMBO J. 2002;21:3029–3038. doi: 10.1093/emboj/cdf316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hrabak EM, Chan CW, Gribskov M, et al. The Arabidopsis CDPK-SnRK superfamily of protein kinases. Plant Physiol. 2003;132:666–680. doi: 10.1104/pp.102.011999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Israelsson M, Siegel RS, Young J, Hashimoto M, Iba K, Schroeder JI. Guard cell ABA and CO2 signalling network updates and Ca2+ sensor priming hypothesis. Curr. Opin. Plant Biol. 2006;9:654–663. doi: 10.1016/j.pbi.2006.09.006. [DOI] [PubMed] [Google Scholar]
- Koornneef M, Reulling G, Karssen C. The isolation and characterization of abscisic acid-insensitive mutants of Arabidopsis thaliana. Physiol. Plant. 1984;74:377–383. [Google Scholar]
- Kuhn JM, Boisson-Dernier A, Dizon MB, Maktabi MH, Schroeder JI. The protein phosphatase AtPP2CA negatively regulates abscisic acid signal transduction in Arabidopsis and effects of abh1 on AtPP2CA mRNA. Plant Physiol. 2006;140:127–139. doi: 10.1104/pp.105.070318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leonhardt N, Kwak JM, Robert N, Waner D, Leonhardt G, Schroeder JI. Microarray expression analyses of Arabidopsis guard cells and isolation of a recessive abscisic acid hypersensitive protein phosphatase 2C mutant. Plant Cell. 2004;16:596–615. doi: 10.1105/tpc.019000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leung J, Bouvier-Durand M, Morris PC, Guerrier D, Chefdor F, Giraudat J. Arabidopsis ABA response gene ABI1: features of a calcium-modulated protein phosphatase. Science. 1994;264:1448–1452. doi: 10.1126/science.7910981. [DOI] [PubMed] [Google Scholar]
- Leung J, Merlot S, Giraudat J. The Arabidopsis ABSCISIC ACID-INSENSITIVE2 (ABI2) and ABI1 genes encode homologous protein phosphatases 2C involved in abscisic acid signal transduction. Plant Cell. 1997;9:759–771. doi: 10.1105/tpc.9.5.759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J, Wang XQ, Watson MB, Assmann SM. Regulation of abscisic acid-induced stomatal closure and anion channels by guard cell AAPK kinase. Science. 2000;287:300–303. doi: 10.1126/science.287.5451.300. [DOI] [PubMed] [Google Scholar]
- Luan S. The CBL–CIPK network in plant calcium signalling. Trends Plant Sci. 2009;14:37–42. doi: 10.1016/j.tplants.2008.10.005. [DOI] [PubMed] [Google Scholar]
- Ma Y, Szostkiewicz I, Korte A, Moes D, Yang Y, Christmann A, Grill E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science. 2009;324:1064–1068. doi: 10.1126/science.1172408. [DOI] [PubMed] [Google Scholar]
- McCourt P, Creelman R. The ABA receptors—we report you decide. Curr. Opin. Plant Biol. 2008;11:474–478. doi: 10.1016/j.pbi.2008.06.014. [DOI] [PubMed] [Google Scholar]
- Merlot S, Gosti F, Guerrier D, Vavasseur A, Giraudat J. The ABI1 and ABI2 protein phosphatases 2C act in a negative feedback regulatory loop of the abscisic acid signalling pathway. Plant J. 2001;25:295–303. doi: 10.1046/j.1365-313x.2001.00965.x. [DOI] [PubMed] [Google Scholar]
- Merlot S, Leonhardt N, Fenzi F, et al. Constitutive activation of a plasma membrane H+-ATPase prevents abscisic acid-mediated stomatal closure. EMBO J. 2007;26:3216–3226. doi: 10.1038/sj.emboj.7601750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer K, Leube MP, Grill E. A protein phosphatase 2C involved in ABA signal transduction in Arabidopsis thaliana. Science. 1994;264:1452–1455. doi: 10.1126/science.8197457. [DOI] [PubMed] [Google Scholar]
- Miao Y, Lv D, Wang P, Wang XC, Chen J, Miao C, Song CP. An Arabidopsis glutathione peroxidase functions as both a redox transducer and a scavenger in abscisic acid and drought stress responses. Plant Cell. 2006;18:2749–2766. doi: 10.1105/tpc.106.044230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mochizuki N, Brusslan JA, Larkin R, Nagatani A, Chory J. Arabidopsis genomes uncoupled 5 (GUN5) mutant reveals the involvement of Mg-chelatase H subunit in plastid-to-nucleus signal transduction. Proc. Natl Acad. Sci. USA. 2001;98:2053–2058. doi: 10.1073/pnas.98.4.2053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moes D, Himmelbach A, Korte A, Haberer G, Grill E. Nuclear localization of the mutant protein phosphatase abi1 is required for insensitivity towards ABA responses in Arabidopsis. Plant J. 2008;54:806–819. doi: 10.1111/j.1365-313X.2008.03454.x. [DOI] [PubMed] [Google Scholar]
- Mori IC, Murata Y, Yang Y, et al. CDPKs CPK6 and CPK3 function in ABA regulation of guard cell S-type anion- and Ca2+ -permeable channels and stomatal closure. PLoS Biol. 2006;4:e327. doi: 10.1371/journal.pbio.0040327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murata Y, Pei ZM, Mori IC, Schroeder J. Abscisic acid activation of plasma membrane Ca2+ channels in guard cells requires cytosolic NAD(P)H and is differentially disrupted upstream and downstream of reactive oxygen species production in abi1-1 and abi2-1 protein phosphatase 2C mutants. Plant Cell. 2001;13:2513–2523. doi: 10.1105/tpc.010210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mustilli AC, Merlot S, Vavasseur A, Fenzi F, Giraudat J. Arabidopsis OST1 protein kinase mediates the regulation of stomatal aperture by abscisic acid and acts upstream of reactive oxygen species production. Plant Cell. 2002;14:3089–3099. doi: 10.1105/tpc.007906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakashima K, Fujita Y, Kanamori N, et al. Three Arabidopsis SnRK2 protein kinases, SRK2D/SnRK2.2, SRK2E/SnRK2.6/OST1 and SRK2I/SnRK2.3, involved in ABA signalling are essential for the control of seed development and dormancy. Plant Cell Physiol. 2009;50:1345–1363. doi: 10.1093/pcp/pcp083. [DOI] [PubMed] [Google Scholar]
- Nambara E, Marion-Poll A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 2005;56:165–185. doi: 10.1146/annurev.arplant.56.032604.144046. [DOI] [PubMed] [Google Scholar]
- Negi J, Matsuda O, Nagasawa T, Oba Y, Takahashi H, Kawai-Yamada M, Uchimiya H, Hashimoto M, Iba K. CO2 regulator SLAC1 and its homologs are essential for anion homeostasis in plant cells. Nature. 2008;452:483–486. doi: 10.1038/nature06720. [DOI] [PubMed] [Google Scholar]
- Nishimura N, Yoshida T, Murayama M, Asami T, Shinozaki K, Hirayama T. Isolation and characterization of novel mutants affecting the abscisic acid sensitivity of Arabidopsis germination and seedling growth. Plant Cell Physiol. 2004;45:1485–1499. doi: 10.1093/pcp/pch171. [DOI] [PubMed] [Google Scholar]
- Nishimura N, Yoshida T, Kitahata N, Asami T, Shinozaki K, Hirayama T. ABA-Hypersensitive Germination1 encodes a protein phosphatase 2C, an essential component of abscisic acid signalling in Arabidopsis seed. Plant J. 2007;50:935–949. doi: 10.1111/j.1365-313X.2007.03107.x. [DOI] [PubMed] [Google Scholar]
- Ohta M, Guo Y, Halfter U, Zhu JK. A novel domain in the protein kinase SOS2 mediates interaction with the protein phosphatase 2C ABI2. Proc. Natl Acad. Sci. USA. 2003;100:11771–11776. doi: 10.1073/pnas.2034853100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pandey S, Nelson DC, Assmann SM. Two novel GPCR-type G proteins are abscisic acid receptors in Arabidopsis. Cell. 2009;136:136–148. doi: 10.1016/j.cell.2008.12.026. [DOI] [PubMed] [Google Scholar]
- Park SY, Fung P, Nishimura N, et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science. 2009;324:1068–1071. doi: 10.1126/science.1173041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pei ZM, Kuchitsu K, Ward JM, Schwarz M, Schroeder JI. Differential abscisic acid regulation of guard cell slow anion channels in Arabidopsis wild-type and abi1 and abi2 mutants. Plant Cell. 1997;9:409–423. doi: 10.1105/tpc.9.3.409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez PL, Benning G, Grill E. ABI2, a second protein phosphatase 2C involved in abscisic acid signal transduction in Arabidopsis. FEBS Lett. 1998;421:185–190. doi: 10.1016/s0014-5793(97)01558-5. [DOI] [PubMed] [Google Scholar]
- Rubio S, Rodrigues A, Saez A, Dizon MB, Galle A, Kim TH, Santiago J, Flexas J, Schroeder JI, Rodriguez PL. Triple loss-of-function of protein phosphatases type 2C leads to partial constitutive response to endogenous ABA. Plant Physiol. 2009;150:1345–1355. doi: 10.1104/pp.109.137174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saez A, Apostolova N, Gonzalez-Guzman M, Gonzalez-Garcia MP, Nicolas C, Lorenzo O, Rodriguez PL. Gain-of-function and loss-of-function phenotypes of the protein phosphatase 2C HAB1 reveal its role as a negative regulator of abscisic acid signalling. Plant J. 2004;37:354–369. doi: 10.1046/j.1365-313x.2003.01966.x. [DOI] [PubMed] [Google Scholar]
- Saez A, Robert N, Maktabi MH, Schroeder JI, Serrano R, Rodriguez PL. Enhancement of abscisic acid sensitivity and reduction of water consumption in Arabidopsis by combined inactivation of the protein phosphatases type 2C ABI1 and HAB1. Plant Physiol. 2006;141:1389–1399. doi: 10.1104/pp.106.081018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saez A, Rodrigues A, Santiago J, Rubio S, Rodriguez PL. HAB1–SWI3B interaction reveals a link between abscisic acid signalling and putative SWI/SNF chromatin-remodelling complexes in Arabidopsis. Plant Cell. 2008;20:2972–2988. doi: 10.1105/tpc.107.056705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schweighofer A, Hirt H, Meskiene I. Plant PP2C phosphatases: emerging functions in stress signalling. Trends Plant Sci. 2004;9:236–243. doi: 10.1016/j.tplants.2004.03.007. [DOI] [PubMed] [Google Scholar]
- Shen YY, Wang XF, Wu FQ, et al. The Mg-chelatase H subunit is an abscisic acid receptor. Nature. 2006;443:823–826. doi: 10.1038/nature05176. [DOI] [PubMed] [Google Scholar]
- Siegel RS, Xue S, Murata Y, Yang Y, Nishimura N, Wang A, Schroeder JI. Calcium elevation-dependent and attenuated resting calcium-dependent abscisic acid induction of stomatal closure and abscisic acid-induced enhancement of calcium sensitivities of S-type anion and inward-rectifying K channels in Arabidopsis guard cells. Plant J. 2009;59:207–220. doi: 10.1111/j.1365-313X.2009.03872.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smalle J, Kurepa J, Yang P, Emborg TJ, Babiychuk E, Kushnir S, Vierstra RD. The pleiotropic role of the 26S proteasome subunit RPN10 in Arabidopsis growth and development supports a substrate-specific function in abscisic acid signalling. Plant Cell. 2003;15:965–980. doi: 10.1105/tpc.009217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vahisalu T, Kollist H, Wang YF, et al. SLAC1 is required for plant guard cell S-type anion channel function in stomatal signalling. Nature. 2008;452:487–491. doi: 10.1038/nature06608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue T, Wang D, Zhang S, Ehlting J, Ni F, Jakab S, Zheng C, Zhong Y. Genome-wide and expression analysis of protein phosphatase 2C in rice and Arabidopsis. BMC Genomics. 2008;9:550. doi: 10.1186/1471-2164-9-550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi-Shinozaki K, Shinozaki K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006;57:781–803. doi: 10.1146/annurev.arplant.57.032905.105444. [DOI] [PubMed] [Google Scholar]
- Yang Y, Costa A, Leonhardt N, Siegel RS, Schroeder JI. Isolation of a strong Arabidopsis guard cell promoter and its potential as a research tool. Plant Methods. 2008;4:1–15. doi: 10.1186/1746-4811-4-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida R, Hobo T, Ichimura K, Mizoguchi T, Takahashi F, Aronso J, Ecker JR, Shinozaki K. ABA-activated SnRK2 protein kinase is required for dehydration stress signalling in Arabidopsis. Plant Cell Physiol. 2002;43:1473–1483. doi: 10.1093/pcp/pcf188. [DOI] [PubMed] [Google Scholar]
- Yoshida R, Umezawa T, Mizoguchi T, Takahashi S, Takahashi F, Shinozaki K. The regulatory domain of SRK2E/OST1/SnRK2.6 interacts with ABI1 and integrates abscisic acid (ABA) and osmotic stress signals controlling stomatal closure in Arabidopsis. J. Biol. Chem. 2006a;281:5310–5318. doi: 10.1074/jbc.M509820200. [DOI] [PubMed] [Google Scholar]
- Yoshida T, Nishimura N, Kitahata N, Kuromori T, Ito T, Asami T, Shinozaki K, Hirayama T. ABA-hypersensitive germination3 encodes a protein phosphatase 2C (AtPP2CA) that strongly regulates abscisic acid signalling during germination among Arabidopsis protein phosphatase 2Cs. Plant Physiol. 2006b;140:115–126. doi: 10.1104/pp.105.070128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu SY, Yu XC, Wang XJ, et al. Two calcium-dependent protein kinases, CPK4 and CPK11, regulate abscisic acid signal transduction in Arabidopsis. Plant Cell. 2007;19:3019–3036. doi: 10.1105/tpc.107.050666. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.