

Abstract
Background
There is growing evidence suggesting that hypothalamic galanin (GAL), which is known to stimulate intake of a fat-rich diet, has a role in promoting the consumption of ethanol. The present study further examined this possibility in GAL knockout (GALKO) mice.
Methods
Two groups of female and male GALKO mice, compared to wild-type (WT) controls, were trained to voluntarily drink increasing concentrations of ethanol, while maintained on lab chow and water. They were examined in terms of their daily ethanol intake and preference, acute consumption of a high-fat diet, preference for flavored solutions, and expression of different peptides shown to stimulate ethanol intake.
Results
In the GALKO mice compared to WT, the results revealed: 1) a 35-45% decrease in ethanol intake and preference, which was evident only at the highest (15%) ethanol concentration, was stronger in female than in male mice, and was seen with comparisons to littermate as well as non-littermate WT mice; 2) a 48% decrease in acute intake of a fat-rich diet, again stronger in female than male mice; 3) no difference in consumption of sucrose or quinine solutions in preference tests; 4) a total loss of GAL mRNA in the hypothalamic paraventricular nucleus (PVN) of female and male mice; and 5) a gender-specific change in mRNA levels of peptides in the perifornical lateral hypothalamus (PFLH), orexin and melanin-concentrating hormone, which are known to stimulate ethanol and food intake and were markedly decreased in females while increased in males
Conclusions
These results provide strong support for a physiological role of PVN GAL in stimulating the consumption of ethanol, as well as a fat-rich diet. Ablation of the GAL gene produced a behavioral phenotype, particularly in females, which may reflect the functional relationship of galanin to ovarian steroids. It also altered the peptides in the PFLH, with their reduced expression contributing to the larger behavioral effects observed in females and their increased expression attenuating these effects in males.
Keywords: Ethanol, Hypothalamus, Galanin, Orexin, Dietary fat
INTRODUCTION
The peptide, galanin (GAL), is known to have a stimulatory effect on food intake (Kyrkouli et al., 1986). This effect is strongest with GAL administration directly into the hypothalamic paraventricular nucleus (PVN), where injection of a GAL receptor antagonist suppresses food intake (Leibowitz and Kim, 1992). A variety of evidence from injection studies and measurements of endogenous peptide has linked PVN GAL to dietary fat (Akabayashi et al., 1994; Leibowitz and Kim, 1992; Nagase et al., 2002; Yun et al., 2005). Administration of this peptide induces a preference for a high-fat diet over a low-fat diet (Leibowitz, 2007) and stimulates daily food consumption with repeated injections only on diets with at least 35% fat (Yun et al., 2005). Moreover, fat consumption and preference for a fat-rich diet is reduced by PVN injection of a GAL antagonist or antisense oligonucleotide to GAL mRNA that reduces GAL peptide levels (Akabayashi et al., 1994; Leibowitz and Kim, 1992). Measurements of endogenous peptides show GAL mRNA and peptide levels in the PVN to be increased in fat-preferring rats and stimulated by acute or chronic consumption of a fat-rich diet or injection of the fat emulsion, intralipid, in association with a rise in circulating triglycerides (Akabayashi et al., 1994; Chang et al., 2004; Gaysinskaya et al., 2007; Leibowitz et al., 2004). Together, these findings suggest that GAL-induced feeding has a specific relationship to dietary fat that is bidirectional in nature, with GAL acting in a positive feedback loop to stimulate the consumption of a high-fat diet that, in turn, further stimulates its expression, leading to overeating of the diet.
In addition to stimulating food intake, GAL also increases the consumption of ethanol (Lewis et al., 2004; Rada et al., 2004). This effect occurs with GAL injection into the ventricles as well as directly into the PVN, and it is evident even in the presence of food, with GAL preferentially stimulating ethanol but not food intake in rats trained to voluntarily drink ethanol. Also, the opposite effect, a marked decrease in ethanol intake, can be observed with PVN injection of the GAL antagonist, M40 (Rada et al., 2004). These results suggest that feeding-stimulatory peptides may have an additional role in potentiating the consumption of ethanol. Clinical evidence for the involvement of endogenous GAL in ethanol intake is supported by reports demonstrating an association of GAL haplotypes with alcoholism in distinct populations (Belfer et al., 2007) and also with circulating triglycerides (Plaisier et al., 2009) that rise with consumption of ethanol as well as a fat-rich diet (Contaldo et al., 1989; Schrezenmeir, 1996).
As with dietary fat, this positive relationship of GAL to ethanol intake appears to be bidirectional, with ethanol intake stimulating the expression and production of endogenous GAL. In rats injected with 10% ethanol or trained to voluntarily drink ethanol, GAL mRNA or peptide is increased specifically in the PVN (Chang et al., 2007; Leibowitz et al., 2003). Further, levels of PVN GAL mRNA are positively correlated with the amount of ethanol consumed as well as levels of blood ethanol and circulating lipids (Chang et al., 2007), and naloxone-induced withdrawal from the opioid effects of ethanol ingestion reverses this ethanol effect on GAL, significantly reducing peptide expression below baseline levels (Leibowitz et al., 2003). These studies support the existence of a positive feedback loop between this peptide and ethanol, with GAL stimulating the drinking of ethanol that in turn enhances endogenous peptide, resulting in overconsumption of ethanol. This GAL-ethanol relationship may involve elevated triglyceride levels induced by ethanol intake, similar to that proposed for the GAL-fat feedback loop (Leibowitz, 2007). With behavioral studies revealing a close, positive relationship between ethanol and fat intake (Carrillo et al., 2004), it is possible that GAL and perhaps other peptides known to stimulate both ethanol and fat intake play a mediating role (Leibowitz, 2007).
To date, studies with GAL mutants indicate that mice with deletions of the GAL gene or GAL receptor genes or that overexpress GAL are able to defend their food intake and body weight when fed ad libitum or fasted on a standard, low-fat diet (Hohmann et al., 2003). Whereas this suggests that GAL may not have a major role in controlling feeding on a standard, laboratory diet, the lack of response in GAL mutants may be attributed to the activation of compensatory mechanisms that help the animals to maintain nutrient homeostasis, as well as an insufficiency of dietary fat which is required for GAL to exhibit its behavioral effects. The latter possibility is supported by a chronic injection study, which showed GAL to significantly increase daily food intake and body weight only in rats maintained on a fat-rich diet (Yun et al., 2005). It is also substantiated by studies in GAL knockout mice (GALKO), which exhibited a decrease in daily food intake and specifically fat intake when maintained on mixed fat-rich or pure macronutrient diets (Adams et al., 2008), and in mice lacking the GAL type 1 receptor gene, which also showed a decrease in chronic consumption of a high-fat diet (Zorrilla et al., 2007). These results support a specific role for endogenous GAL in controlling feeding behavior on a fat-rich diet.
The present study examined GALKO mice to test the possibility that GAL may have a similar function in the control of ethanol intake. These mice were trained to voluntarily drink increasing concentrations of ethanol, and their daily intake was compared to that of wild-type (WT) controls. With evidence showing GAL to function in close, positive relation to the ovarian steroids in controlling ingestive behavior (Leibowitz et al., 1998; Leibowitz et al., 2007; Leibowitz et al., 2009), we examined both females and males to determine if the behavioral changes induced by GAL deficiency are gender specific. We also measured hypothalamic expression of GAL as well as other orexigenic peptides that may be altered by a deficiency in GAL and thus contribute to the phenotype of GALKO mice. With studies of orexin (ORX) and melanin-concentrating hormone (MCH) showing these peptides to have similarities to GAL in relation to dietary fat, ethanol and ovarian steroids (Chang et al., 2008; Duncan et al., 2005; Leibowitz and Wortley, 2004; Porkka-Heiskanen et al., 2004; Schneider et al., 2007; Viale et al., 1999), changes in endogenous GAL in the mutant mice may, in turn, have impact on these two peptides expressed predominantly in the nearby, perifornical lateral hypothalamus (PFLH). The results of these tests provide support for the hypothesis that GAL acts physiologically to stimulate the ingestion of ethanol as well as a high-fat diet.
MATERIALS AND METHODS
Animals
The GALKO mice were generated on the 129Ola/Hsd background (Wynick et al., 1998). As described previously, exons one to five of the GAL gene were replaced with a PGK-Neo cassette in reverse orientation, removing the signal peptide, coding region, and the majority of the GAL-associated peptide. In homozygous GALKO mice, the expression of GAL was undetectable in all tissues examined (Wynick et al., 1998). Once generated, the chimeras were bred to co-isogenic 129Ola/Hsd background to generate mice heterozygous for the GALKO allele. These mice became the founders of a colony bred by Dr. Marina Picciotto at Yale School of Medicine, who generously provided us with 4 WT and 4 KO mice, each set with 2 males and 2 females. To generate large numbers of animals of both genders for the two groups examined in this study (n=5-8/genotype/gender), we bred WT × WT and KO × KO mating pairs to obtain non-littermate WT and GALKO offspring for Group 1 and also WT × KO offspring to get HET × HET mating pairs that, in turn, generated littermate WT and GALKO offspring for Group 2. To confirm or determine the genotype of the mice, the samples were sent to Transnetyx Inc. (see below). Only the homozygous GALKO mice were examined in this study. Experimental testing occurred when the mice were 2-5 months of age, with both females and males tested in each experiment. The mice, which were sex-and age-matched in both groups, were individually housed in plastic cages, in a fully accredited AAALAC facility (22°C, with a 12:12-h light-dark cycle with lights off at 1 pm), according to institutionally approved protocols as specified in the NIH Guide to the Use and Care of Animals and also with the approval of the Rockefeller University Animal Care Committee. The mice were maintained ad libitum on laboratory chow and water, and their food intake, water intake, and body weights were measured 2-3 times per week.
Genotyping
At 19-20 days of age, the mice from both Group 1 and Group 2 were anesthetized with isoflourene, and a sample from the tip of their tail was collected and sent to Transnetyx (Cordova, TN) for genotype analysis using the primers and PCR conditions previous described (Wynick et al., 1998).
Ad libitum ethanol consumption and preference
The female and male WT and GALKO mice in Groups 1 and 2, while maintained on lab chow, were individually housed and habituated in their home cage to drink from two bottles. These bottles were 50 ml conical centrifuge tubes purchased from Nalgene Nunc International (Rochester, NY). For the first week, both bottles contained plain water. After that, the mice were given access to one bottle of ethanol mixed with water and the other bottle of plain water. The concentration of ethanol (v/v) was increased every 4 days, from 3% to 8%, 10% and then 15%, over the course of the experiment. The position of the bottles was changed daily to control for position preference. Intake of ethanol, water and chow was measured daily, and body weight was measured twice weekly. The measure of daily ethanol intake (g/kg body weight) was obtained by averaging ethanol intake over the 4 days of measurements at each concentration. Relative ethanol preference was calculated in terms of the ratio of ethanol intake to total fluid volume consumed.
Assessment of high-fat meal consumption
While maintained ad libitum on 15% ethanol, the female and male mice of Group 1 were then tested with a high-fat diet (5.15 Kcals/g), which was given for a brief period at dark onset. This diet, described in detail elsewhere (Chang et al., 2008), consisted of 50% fat (with 80% lard and 20% vegetable oil), 25% carbohydrate (with 30% dextrin, 30% cornstarch and 40% sucrose), and 25% protein (with casein from Bioserv, Frenchtown, NJ, and 0.3% L-cystine and DL-methionine from MP Biomedicals, Solon, OH) and was supplemented with 4% minerals (Briggs N Salt Mixture, MP Biomedicals) and 3% vitamins (Vitamin Diet Fortification Mixture, MP Biomedicals). To adapt them to the diet before the test, the mice were given at dark onset a daily sample (5 Kcal) over a 5-day period. For the actual experiment, the chow and ethanol were removed 2 h before the test, and the mice were then given the high-fat diet at dark onset, which they were allowed to consume ad libitum over the next 2-h period. This test was repeated 4 times over 4 consecutive days, and the final intake measure (Kcal/2 h) was the average of these scores obtained over the 4 days. One week after completing these behavioral tests, blood was collected by retro-orbital eye bleeds for measurement of blood ethanol levels. On the day of the collection, the mice were allowed to drink for 3 h after dark onset, during which time the GALKO consumed slightly but not significantly less ethanol than the WT controls (Females: 4.9 ± 0.25 vs 5.4 ± 0.31 g/kg; Males: 3.5 ± 0.48 vs 3.9 ± 0.25 g/kg). Ocular blood was then collected, and ethanol levels were measured using the Analox GM7 Fast Enzymatic Metabolizer (Analox, Lunenburg, MA).
Ad libitum sucrose/quinine access procedure
The mice of Group 2 were taken off ethanol and examined 2 weeks later for their preference for sucrose and quinine. Over an 8-day period, the mice were given continuous access to plain water in one bottle and sucrose or quinine in the other, with the position of the bottles switched daily. They had access to these solutions over a 48-h period, with the sucrose solutions (1.70% and 4.25%) tested first, followed by the quinine solutions (0.03 mM and 0.10 M). Preference for these solutions was determined by dividing the volume of the taste solution by the total volume of taste solution and water.
Brain dissection
Levels of peptide mRNA in the littermate WT and GAL KO of Group 2 (n=4-8/genotype/gender) were examined. One week after completing the sucrose/quinine preference tests, the mice were sacrificed by rapid decapitation 2 h before dark onset. Their brains were rapidly removed and placed in a mouse matrix with the ventral surface facing up, and three 1.0 mm coronal sections were made, with the middle optic chiasma as the anterior boundary and the middle infundibulum as the posterior boundary. The sections were placed on a glass slide, and the PVN (Bregma -0.58 to -1.06 mm) and PFLH (Bregma -1.70 to -2.06 mm) were rapidly microdissected under a microscope, with the assistance of a mouse brain atlas (Paxinos and Franklin, 2001) and using the fornix and third ventricle as landmarks. The PVN was dissected as a reversed isosceles triangle, 1.0 mm bilateral to the ventricle and between the fornix structures. For the PFLH, the dissection was taken from the area surrounding the fornix, within a range of 0.2 mm medial and ventral to the fornix, 0.3 mm dorsal and 0.1 mm lateral. These dissections were immediately frozen in liquid nitrogen and stored at -80° C until processed.
Real-time quantitative PCR analysis of mRNA
Real-time quantitative PCR was used to measure mRNA levels of GAL in the PVN and ORX and MCH in the PFLH, as previously described (Chang et al., 2008). Briefly, total RNA was extracted with TRIzol reagent, and template cDNA was synthesized using an oligo-dT primer and SuperScript II reverse transcriptase. Primers were designed with Applied Biosystems Primer Express V1.5a software. The sequences of the primers were: GAL (5’-TTCCCACCACTGCTCAAGATG-3’ and 5’-TGGCTGACAGGGTTGCAA-3’) (GenBank #NM033237) generate a 78 bp amplicon corresponding to the nucleotide 127-205 of the sequence, ORX (5’-AGATACCATCTCTCCGGATTGC-3’ and 5’CCAGGGAACCTTTGTAGAAGGA-3’) (GenBank #AF019565) generate a 73 bp amplicon corresponding to the nucleotide 48-121 of the sequence, MCH (5’-ATCGGTTGTTGCTCCTTCTCTG-3’ and 5’-TCTGCTTGGAGCCTGTGTTCTT-3’) (GenBank #NM012625) generate a 101 bp amplicon corresponding to the nucleotide 224-324 of the sequence, β-actin (5’-GGCCAACCGTGAAAAGATGA-3’ and 5’-CACAGCCTGGATGGCTACGT-3’ (GenBank #NM031144) generate a 79 amplicon corresponding to the nucleotide 420-498 of the sequence. The SYBR Green PCR core reagents kit (Applied Biosystems, Foster City, CA) was used for PCR, which was performed in MicroAmp Optic 96-well Reaction Plates (Applied Biosystems, Foster City, CA) on an ABI PRISM 7900 Sequence Detection system (Applied Biosystems, Foster City, CA), with the condition of 2 min at 50°C, 10 min at 95°C, then 40 cycles of 15 s at 95°C and 1 min at 60°C. Each study consisted of 4 independent runs of PCR in triplicate, and each run included a standard curve, non-template control, and negative RT control. The concentrations of primers were 100 to 200 nM, and all reagents, unless indicated, were from Invitrogen (Carlsbad, CA). The levels of target gene expression were quantified relative to the level of β-actin, using standard curve method. The specificities of RT-PCR products were confirmed by both a single dissociation curve of the product and a single band with corresponding molecular weight revealed by agarose gel electrophoresis. They were also verified by an anatomical negative control by using the corpus callosum in the same brain
Data Analysis
All values are expressed as mean ± SEM. With a standard statistical package (SPSS), statistical analyses comparing the different behavioral or peptide measures for the subgroups were performed using a 2-way ANOVA, with repeated measures where appropriate, or an unpaired, two-tailed Student’s t-test. The criterion for use of the term “significant” in the text is that the probability value for a given test is p<0.05.
RESULTS
Experiment 1: Ethanol consumption and taste preference
In this experiment, female and male mice lacking the GAL gene were given ad libitum access to increasing concentrations of ethanol, from 3% to 15%, and they were compared to their respective female and male WT control mice. In Group 1 with GALKO mice compared to non-littermate WT, a 2-way ANOVA relating genotype to ethanol intake on the different concentrations revealed a main behavioral effect, reduced intake of ethanol in GALKO mice, which was statistically significant in female mice [F(3,67) = 5.74, p < 0.001] but did not reach significance in males [F(3,47) = 1.86, p = 0.161]. This effect was observed only on the 15% ethanol concentration, when the female GALKO mice exhibited a significant, 43% decrease in intake (p<0.001) (Fig. 1A) and the male GALKO mice showed only a 29% decrease (Table 1). Preference for ethanol relative to water was also significantly decreased in females [F(3,63) = 12.13, p < 0.001] but not males [F(3,43) = 2.40, p = 0.09], with the 15% ethanol concentration revealing a 34% reduction in females (Fig. 1B) and a 21% reduction in males (Table 1). Both the female and male GALKO mice throughout the experiment were similar to their respective WT controls (Table 2) in the measures of daily chow intake, water intake, total fluid intake, and body weight, except for total fluid intake on 15% ethanol which was significantly reduced (p<0.05) in female GALKO mice due to their decrease in ethanol intake. The GALKO and WT were also similar in their blood ethanol levels when consuming 15% ethanol, as indicated by the measures in both females (69 ± 0.7 vs 73 ± 3.6 mg/dl) and males (67 ± 4.4 vs 68 ± 3.5 mg/dl). Gender comparisons showed a tendency for female mice, both WT and GALKO, to drink more ethanol than the males across all concentrations, although this effect did not reach statistical significance [F(3,109) = 3.12, p = 0.28].
Figure 1.
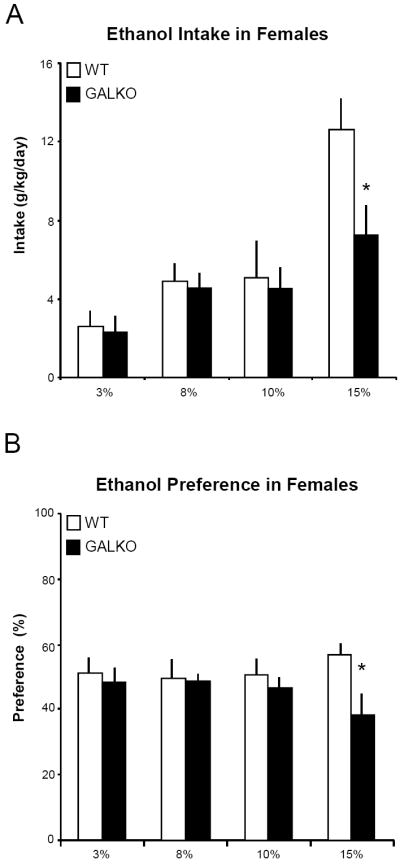
Ethanol intake and preference in female galanin knockout (GALKO) and wild-type (WT) mice voluntarily drinking increasing concentrations of ethanol (3%-15%): (A) Ethanol intake (g/kg/day), plotted as a function of ethanol concentration (4-day average) in a two-bottle, continuous-access paradigm; and (B) Ethanol preference (%), calculated as volume of ethanol consumed divided by total volume of solution consumed (x 100). The GALKO mice compared to WT showed a significant decrease in ethanol intake and preference on 15% ethanol (*, p<0.001).
Table 1.
Ethanol Intake and Ethanol Preference in Male Mice
| WT | GALKO | |
|---|---|---|
| Ethanol Intake (g/kg/day) | ||
| 3% | 1.5 ± 0.5 | 1.9 ± 0.2 |
| 8% | 2.9 ± 0.9 | 2.0 ± 0.6 |
| 10% | 4.0 ± 1.9 | 1.6 ± 1.5 |
| 15% | 9.0 ± 2.2 | 6.2 ± 1.8 |
| Ethanol Preference (%) | ||
| 3% | 37 ± 3.0 | 43 ± 5.2 |
| 8% | 34 ± 4.3 | 27 ± 3.5 |
| 10% | 36 ± 4.2 | 15 ± 5.8 |
| 15% | 38 ± 3.1 | 30 ± 4.2 |
Table 2.
Behavioral Measures in Female and Male Mice Drinking Ethanol Ad Libitum at Different Concentrations
| Female | Male | |||
|---|---|---|---|---|
| WT | GALKO | WT | GALKO | |
| Chow Intake (kcal/day) | ||||
| 3% | 13 ± 0.53 | 14 ± 1.50 | 18 ± 0.72 | 16 ± 0.32 |
| 8% | 18 ± 1.40 | 17 ± 0.42 | 16 ± 0.41 | 14 ± 0.64 |
| 10% | 17 ± 0.73 | 18 ± 0.64 | 16 ± 0.52 | 13 ± 2.40 |
| 15% | 17 ± 1.20 | 16 ± 0.92 | 15 ± 0.62 | 14 ± 2.10 |
| Water Intake (g/kg/day) | ||||
| 3% | 84 ± 5.0 | 89 ± 6.2 | 85 ± 7.2 | 85 ± 5.7 |
| 8% | 65 ± 2.3 | 68 ± 2.1 | 70 ± 8.1 | 68 ± 4.9 |
| 10% | 51 ± 6.0 | 66 ± 1.4 | 80 ± 6.5 | 88 ± 6.1 |
| 15% | 65 ± 7.2 | 73 ± 0.8 | 99 ± 5.8 | 92 ± 8.7 |
| Total Fluid Intake (g/kg/day) | ||||
| 3% | 171 ± 7.3 | 166 ± 9.5 | 138 ± 7.2 | 148 ± 5.7 |
| 8% | 126 ± 13.5 | 162 ± 15.0 | 106 ± 7.2 | 93 ± 3.2 |
| 10% | 111 ± 12.3 | 122 ± 11.6 | 125 ± 8.4 | 104 ± 3.1 |
| 15% | 149 ± 9.4 | 129 ± 9.1* | 159 ± 14.4 | 133 ± 14.6 |
| Body Weight (g) | ||||
| 3% | 22 ± 0.7 | 23 ± 0.7 | 26 ± 0.8 | 27 ± 1.3 |
| 8% | 23 ± 0.7 | 22 ± 0.4 | 27 ± 0.9 | 28 ± 1.1 |
| 10% | 23 ± 1.0 | 22 ± 0.4 | 27 ± 0.5 | 30 ± 1.0 |
| 15% | 24 ± 1.3 | 24 ± 1.0 | 29 ± 0.8 | 30 ± 0.9 |
| Sucrose Preference (%) | ||||
| 1.70% | 82 ± 7.2 | 89 ± 6.4 | 92 ± 3.0 | 91 ± 3.0 |
| 4.30% | 81 ± 7.8 | 87 ± 9.1 | 96 ± 9.5 | 87 ± 4.0 |
| Quinine Preference (%) | ||||
| 0.03 mM | 18 ± 2.3 | 17 ± 1.6 | 15 ± 2.1 | 15 ± 3.2 |
| 0.10 mM | 19 ± 2.6 | 15 ± 1.0 | 13 ± 1.0 | 15 ± 1.3 |
p<0.05 comparing GALKO to WT mice
This effect of genotype on 15% ethanol intake was not litter dependent, as it was similarly seen in Group 2 with GALKO and littermate WT controls. As shown in Group 1, reduced intake of 15% ethanol was seen in the female GALKO vs WT (7.92 ± 0.94 vs 12.9 ± 1.0 g/kg/day, p < 0.03), with only a trend observed in the male GALKO vs WT (7.22 ± 0.96 vs 9.18 ± 0.88 g/kg/day, p = 0.21). This decrease in intake was not due to any obvious change in taste preference. Tests conducted in Group 2 one week after withdrawing ethanol revealed no difference between genotypes in their preference for the sucrose solution in females [F(1,15) = 0.02, p = 0.93] or males [F(1,13) = 0.03, p = 0.96] or for the quinine solution in females [F(1,15) = 1.42, p = 0.27] or males [F(1,13) = 0.45, p = 0.53] relative to water (Table 2). Together, these results demonstrate that a deficiency of GAL reduces ad libitum consumption of ethanol in females more than males, that this behavioral effect can be seen when comparing GALKO mice to littermate as well as non-littermate WT controls, and that the reduced ethanol intake does not reflect any clear difference in preference for other flavored solutions or in blood levels of ethanol.
Experiment 2: Consumption of a high-fat diet
In a recent study that examined GALKO mice of mixed sex on a chronic high-fat diet (Adams et al., 2008), daily consumption of dietary fat, in experiments using a mixed high-fat diet or jars of pure macronutrients, was found to be reduced by 40-50%, similar to the decrease in daily ethanol intake shown in Experiment 1. To further examine the changes in eating behavior and fat preference induced by GAL deficiency, the WT and GALKO female and male mice of Group 1, while being maintained on lab chow, were tested for their ingestion of a high-fat diet provided during a brief period at dark onset. Measurements of caloric intake during the 2-h test revealed a marked change in the GAL deficient mice. While showing normal daily intake of lab chow (Table 2), the female GALKO mice compared to WT demonstrated a 55% reduction in the amount of high-fat diet ingested during the acute test (Fig. 2). The male mice, which consumed less high-fat diet compared to females (p < 0.01) during the 2-h test period, exhibited a smaller, statistically insignificant genotype effect, with the GALKO mice showing only a 35% decrease in intake compared to WT (Fig. 2). Thus, in addition to confirming the findings obtained with chronic consumption of a high-fat diet (Adams et al., 2008), these measures of a brief period of diet consumption show this reduction in high-fat diet intake to be stronger in females, similar to that observed with the measures of ethanol intake.
Figure 2.
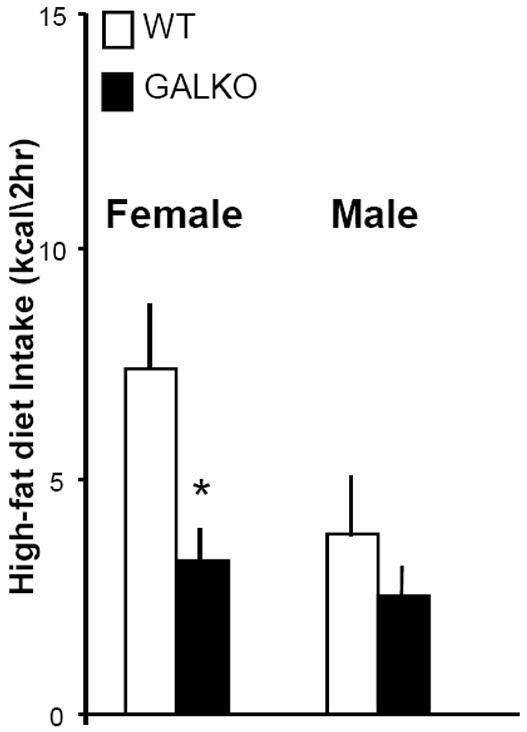
High-fat diet intake (kcal/2h) in female and male GALKO and WT mice during a 2-h test period at dark onset (average of 4 tests), with chow and ethanol removed. Compared to WT, the GALKO females but not males exhibited a significant decrease in high-fat diet intake (*, p<0.01).
Experiment 3: Expression of orexigenic peptides in the hypothalamus
This experiment was designed to confirm the deficiency of GAL in the PVN of GALKO mice and also measure other hypothalamic peptides involved in ethanol intake that may be affected by this GAL deficiency. One week after completing the sucrose/quinine preference tests, the female and male WT and GALKO mice of Group 2, while maintained on only lab chow and water, were examined for peptide gene expression using real-time quantitative PCR. The orexigenic peptides studied were GAL, ORX and MCH, each of which is found to stimulate ethanol intake (Duncan et al., 2005; Lewis et al., 2004; Rada et al., 2004; Schneider et al., 2007). In agreement with the reduction in GAL protein levels described in whole hypothalamus (Wynick et al., 1998), the GALKO mice compared to WT of this study exhibited a reduction in GAL mRNA in the PVN to background levels, in both females and males (Fig. 3). Additional measurements showed that this loss of GAL was accompanied by gender-specific changes in the expression of ORX and MCH in the PFLH (Fig. 3). In the GALKO compared to WT mice, the expression of these two peptides was greatly reduced (-75% and -39%, respectively) in females but markedly increased (+213% and +243%, respectively) in males. These results demonstrate that ablation of the GAL gene has effects that extend beyond GAL and involve other peptides that affect ethanol as well as fat intake and thus possibly contribute to gender-specific alterations in the behavioral phenotype.
Figure 3.
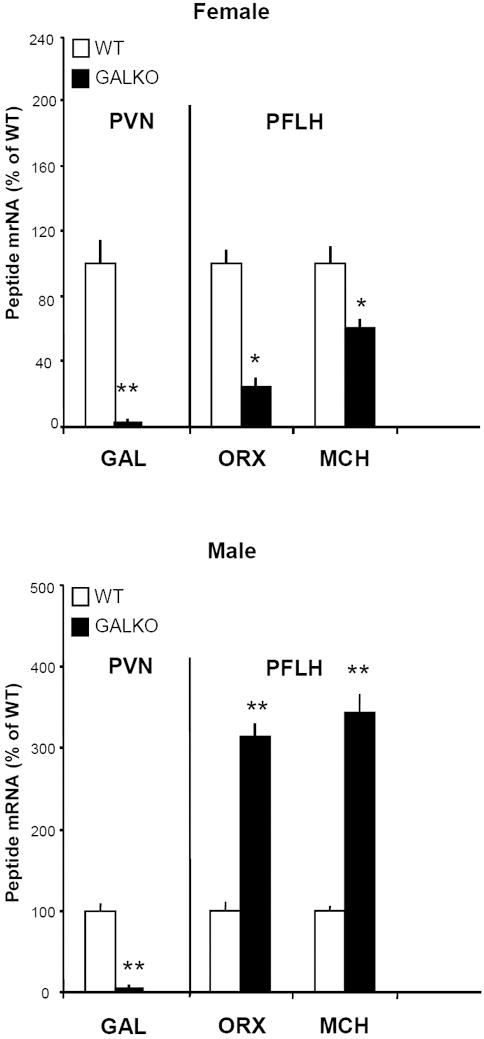
Gene expression of galanin (GAL) in the paraventricular nucleus (PVN) and orexin (ORX) and melanin-concentrating hormone (MCH) in the perifornical lateral hypothalamus (PFLH) of GALKO and WT mice, as measured by real-time quantitative PCR. Both female and male GALKO mice displayed a reduction in GAL mRNA to background levels in the PVN. Expression of ORX and MCH in the PFLH was markedly reduced in female GALKO mice (*, p < 0.01), while increased in male GALKO mice (**, p<0.001).
DISCUSSION
There is considerable evidence, from studies with injections and measurements of GAL, suggesting a positive relationship between this peptide and ethanol intake that is bidirectional in nature and impacts on voluntary ethanol intake in Sprague-Dawley rats. The present results in GAL mutant mice provide further support for this relationship.
Effect of GAL deficiency on ethanol intake as a function of gender
In accordance with the known stimulatory effect of GAL injection on the consumption of ethanol, the female GALKO mice compared to WT controls exhibited a significant reduction in their daily intake of ethanol and a parallel decrease in preference for ethanol relative to water. This change in female mice became evident only at the 15% concentration of ethanol, when the switch from a 10% to 15% ethanol solution increased ethanol intake (in g/kg) by more than 150% in the WT mice but less than 60% in the GALKO mice. This difference between genotypes suggests that endogenous GAL in stimulating ethanol intake becomes active only at the highest, 15% concentration of ethanol, when the mice are most vulnerable to the effects of GAL gene ablation. This decrease in ethanol intake was not litter dependent, since it was observed when the GALKO mice were compared to littermate as well as non-littermate WT controls. The results of GAL ablation on ethanol intake was apparently not attributed to a change in taste for flavored solutions, as preference tests with sweet (sucrose) and bitter (quinine) solutions in both females and males revealed no differences between the WT and GALKO mice. Also, it did not reflect a change in ethanol metabolism, as blood levels of ethanol were not altered in the GALKO mice on the 15% concentration. While the male GALKO mice showed a tendency toward a decrease in ethanol intake and preference, this did not reach statistical significance, suggesting that mechanisms involving reproductive hormones (see below) may contribute to GAL’s role in the control of ethanol intake.
Effect of GAL deficiency on food intake and body weight
There are numerous studies with injection and measurements of GAL suggesting a role for this peptide in stimulating feeding behavior and weight gain in rats and mice (Hohmann et al., 2003; Leibowitz, 2007). In the present study of mice drinking ethanol, however, measurements of daily food intake and body weight showed no difference between genotype in either females or males. While consistent with published studies in ethanol-naive, GALKO mice (Adams et al., 2008; Hohmann et al., 2003; Wynick et al., 1998), this lack of change in food intake in these mutant mice may reflect other confounding factors rather than a lack of function for GAL. In addition to the diet which is important for revealing the effects of GAL (see below), recent studies in rats trained to drink ethanol have shown that PVN injection of GAL stimulates only ethanol intake and not food intake as one normally sees in the absence of ethanol (Lewis et al., 2004; Rada et al., 2004). This finding has led to the proposal that chronic availability of ethanol can usurp the role of endogenous GAL in feeding and cause a shift in its stimulatory effect on ingestive behavior to the consumption of ethanol (Leibowitz, 2007).
Possible mechanisms underlying the gender-specificity of reduced ethanol intake
The stronger effect of GAL deficiency on ethanol intake observed in females compared to males suggests that reproductive hormones may be involved in the stimulatory effect of GAL on consumption of ethanol. This is supported by evidence showing that GAL, in the stimulation of food intake, acts in close association with the ovarian steroids, estrogen (E2) and progesterone (P4), both in pubertal and adult female rats (Leibowitz et al., 1998; Leibowitz et al., 2007; Leibowitz et al., 2009). These steroids have a strong stimulatory effect on endogenous GAL in the PVN and medial preoptic nucleus (MPN), and they are essential for the stimulatory effect of dietary fat on GAL expression in these nuclei (Leibowitz et al., 2007; Leibowitz et al., 2009). Their circulating levels rise in close association with the increase in GAL mRNA in the PVN and MPN, during proestrous in adult rats (Leibowitz et al., 1998; Leibowitz et al., 2007; Rossmanith et al., 1996) and at puberty onset (Leibowitz et al., 2009), and they themselves are found to stimulate the consumption of ethanol, in both ovariectomized and intact female rats (Ford et al., 2004; Marinelli et al., 2003; Rajasingh et al., 2007; Reid et al., 2002). While GAL is known to modulate the release of gonadotropins (López et al., 1993), pituitary content of these hormones is unaltered in ethanol-naïve, GALKO mice (Wynick et al., 1998), leading us to consider the involvement of other reproductive hormones in GAL’s effect on ethanol intake. One such hormone may be prolactin. Pituitary levels of prolactin mRNA and protein are markedly reduced in female GALKO mice (Wynick et al., 1998). Moreover, in addition to stimulating ethanol intake, GAL has a strong, stimulatory effect on the secretion of prolactin (Koshiyama et al., 1987; Ottlecz et al., 1988), which in turn can increase food intake (Cai et al., 1998; O’Halloran et al., 1991), and prolactin in E2-primed rats stimulates GAL (Hammond et al., 1997). Whereas there are no studies in female rats showing a causal effect of prolactin on ethanol intake, there is evidence that chronic consumption of ethanol increases circulating prolactin levels in females, while having no effect in males (Emanuele et al., 2001; Salonen and Huhtaniemi, 1990; Sanchis et al., 1985). This evidence supports the possibility that prolactin contributes to the positive feedback loop between GAL and ethanol and performs this function in a gender-specific manner.
Relationship between GAL, dietary fat and ethanol intake
In addition to the steroids and prolactin, GAL is known to have a close, positive relationship to dietary fat, which in turn is related to the consumption of ethanol (Leibowitz, 2007). Two recent studies in mutant mice provide further support for these associations with dietary fat. When chronically maintained on a mixed high-fat diet or allowed to choose a diet from pure macronutrients, GAL deficient mice compared to WT (gender unspecified) consume less of the mixed diet and choose less of the pure fat, and this effect is partially reversed by central administration of GAL (Adams et al., 2008). Further, mice lacking the GAL type 1 receptor also show a decrease in chronic consumption of high-fat diet (Zorrilla et al., 2007). The results of the present investigation confirm and extend these findings. They show that GALKO mice compared to WT, while maintained on chronic lab chow, exhibit the expected decline in the consumption of a high-fat diet, consuming less calories of the diet when made available only during a brief period at the start of the feeding cycle. In addition, they demonstrate that this reduced intake of dietary fat is considerably stronger in females than males. This gender difference, consistent with the stronger reduction in ethanol intake observed in female GALKO mice, further links these two nutrients and their underlying mechanisms. The tendency of females to exhibit greater fat intake and show a stronger preference for this macronutrient than males, as shown here in mice and also in rats (Leibowitz et al., 1991; Leibowitz et al., 2009), has been attributed to GAL in the PVN and MPN, where it exhibits greater expression in females compared to males, and also to progesterone, which in E2-primed rats stimulates GAL in these nuclei and increases the consumption of fat (Leibowitz et al., 1998; Leibowitz et al., 2007). There are reports suggesting that females may consume more ethanol compared to males (Almeida et al., 1998; Cailhol and Mormede, 2001), and this gender difference may also be attributed, in part, to GAL. Thus, the reduced consumption of a fat-rich diet as well as ethanol specifically in female GALKO mice is an expected consequence of GAL deficiency. It substantiates the existence of a positive feedback loop between dietary fat, ethanol, steroids and GAL, in a circuit that involves the PVN and MPN and possibly the anterior pituitary where GAL is heavily expressed and similarly responsive to ovarian steroids and dietary fat (Leibowitz et al., 2007).
Effect of GAL gene ablation on GAL expression in the PVN
The original characterization of the GALKO mice used radioimmunoassay to measure GAL peptide levels in whole hypothalamus as well as other tissues and observed a 50% decline in peptide levels in heterozygotes and undetectable levels of GAL in homozygote mutants (Wynick et al., 1998). To confirm that endogenous GAL is deficient in the present GALKO mice, the expression of this peptide was measured, using real-time quantitative PCR, specifically in the PVN where GAL is believed to act in the control of consummatory behavior (Leibowitz, 2007; Yun et al., 2005). In the GALKO compared to WT controls, the results demonstrated only background levels of PVN GAL mRNA, validating the genotype of the animals and their deficiency of GAL. While this result and the behavioral changes support a role for GAL in controlling both ethanol and fat intake, it fails to provide any explanation for the gender differences, with female GALKO mice showing much stronger effects than males. While it may be the combination of disturbances in GAL and prolactin that contribute to the behavioral changes in females, it is important to consider additional, possibly neurochemical changes in males that might mitigate the effects of GAL gene ablation and rescue their normal consummatory behavior.
Effect of GAL gene ablation on expression of other orexigenic peptides
To provide information on the possibility that other orexigenic peptides may be involved in these behavioral changes seen in GALKO mice, we measured the expression of two additional peptides, ORX and MCH, which have been related to dietary fat and ethanol. Like GAL, these two peptides are found to stimulate the ingestion of food (Clegg et al., 2002; Sweet et al., 1999) and ethanol (Duncan et al., 2005; Schneider et al., 2007), and they, in turn, are shown to be stimulated by the consumption of a high-fat diet as well as ethanol (Chang et al., 2008; Chang et al., 2004; Kennedy et al., 2007; Wortley et al., 2003). These peptides may also rise in relation to the ovarian steroids (Porkka-Heiskanen et al., 2004; Viale et al., 1999). To our knowledge, there is only one report that has investigated the effects of GAL gene ablation on other peptides, reporting no difference between GALKO and WT mice in their hypothalamic content of neuropeptide Y, in both females and males (Wynick et al., 1998). The present study revealed marked changes in the expression of ORX and MCH in the PFLH and showed these changes in the two peptides to be remarkably similar, as well as gender specific. The expression of both ORX and MCH in GALKO mice, compared to WT, was significantly reduced in the females but greatly increased, by more than 3-fold, in the males. Whereas there is other evidence showing gene ablation of a feeding-related peptide to affect the expression of another such peptide (Marsh et al., 1999; Qian et al., 2002), the present results are particularly distinctive in revealing such dramatic changes in ORX and MCH in GALKO mice, ones that are diametrically opposite in females and males. While the deficiency in GAL may explain the reduction in both ethanol and fat intake observed in female GALKO mice, the differences between females and males in their patterns of ORX and MCH expression may also be important in explaining the gender specificity of their behavioral phenotype. In line with the evidence that both ORX and MCH have a stimulatory effect on ethanol intake when injected into the brain (Duncan et al., 2005; Schneider et al., 2007), the marked reduction in ORX and MCH mRNA in female GAKKO mice may contribute, together with GAL deletion, to their decrease in ethanol consumption. On the other hand, the male GALKO mice, despite their loss of GAL, may be consuming normal ethanol and fat intake due to the increase in PFLH ORX and MCH expression. Thus, although both female and male mice have absent GAL expression, only females appear to be sensitive to the absence of the GAL gene, with the males able to compensate by stimulating other neurochemical systems.
Acknowledgments
This research is funded by USPHS Grants, AA 12882 and DA 21518. We thank Dr. Marina Picciotto at Yale School of Medicine for providing breeding pairs of GALKO and WT mice and Ms. Helen Martirosova and Olga Berman for their technical assistance.
References
- Adams AC, Clapham JC, Wynick D, Speakman JR. Feeding behaviour in galanin knockout mice supports a role of galanin in fat intake and preference. J Neuroendocrinol. 2008;20(2):199–206. doi: 10.1111/j.1365-2826.2007.01638.x. [DOI] [PubMed] [Google Scholar]
- Akabayashi A, Koenig J, Watanabe Y, Alexander J, Leibowitz S. Galanin-containing neurons in the paraventricular nucleus: a neurochemical marker for fat ingestion and body weight gain. Proc Natl Acad Sci U S A. 1994;91(22):10375–9. doi: 10.1073/pnas.91.22.10375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almeida O, Shoaib M, Deicke J, Fischer D, Darwish M, Patchev V. Gender differences in ethanol preference and ingestion in rats. The role of the gonadal steroid environment. J Clin Invest. 1998;101(12):2677–85. doi: 10.1172/JCI1198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belfer I, Hipp H, Bollettino A, McKnight C, Evans C, Virkkunen M, Albaugh B, Max M, Goldman D, Enoch M. Alcoholism is associated with GALR3 but not two other galanin receptor genes. Genes Brain Behav. 2007;6(5):473–81. doi: 10.1111/j.1601-183X.2006.00275.x. [DOI] [PubMed] [Google Scholar]
- Cai A, Bowers R, Moore JJ, Hyde J. Function of galanin in the anterior pituitary of estrogen-treated Fischer 344 rats: autocrine and paracrine regulation of prolactin secretion. Endocrinology. 1998;139(5):2452–8. doi: 10.1210/endo.139.5.6025. [DOI] [PubMed] [Google Scholar]
- Cailhol S, Mormède P. Sex and strain differences in ethanol drinking: effects of gonadectomy. Alcohol Clin Exp Res. 2001;25(4):594–9. [PubMed] [Google Scholar]
- Carrillo CA, Leibowitz SF, Karatayev O, Hoebel BG. A high-fat meal or injection of lipids stimulates ethanol intake. Alcohol. 2004;34(23):197–202. doi: 10.1016/j.alcohol.2004.08.009. [DOI] [PubMed] [Google Scholar]
- Chang G-Q, Gaysinskaya V, Karatayev O, Leibowitz SF. Maternal high-fat diet and fetal programming: increased proliferation of hypothalamic peptide-producing neurons that increase risk for overeating and obesity. J Neurosci. 2008;28(46):12107–19. doi: 10.1523/JNEUROSCI.2642-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang G-Q, Karatayev O, Ahsan R, Avena NM, Lee C, Lewis MJ, Hoebel BG, Leibowitz SF. Effect of ethanol on hypothalamic opioid peptides, enkephalin, and dynorphin: relationship with circulating triglycerides. Alcohol Clin Exp Res. 2007;31(2):249–59. doi: 10.1111/j.1530-0277.2006.00312.x. [DOI] [PubMed] [Google Scholar]
- Chang G-Q, Karatayev O, Davydova Z, Leibowitz SF. Circulating triglycerides impact on orexigenic peptides and neuronal activity in hypothalamus. Endocrinology. 2004;145(8):3904–12. doi: 10.1210/en.2003-1582. [DOI] [PubMed] [Google Scholar]
- Clegg D, Air E, Woods S, Seeley R. Eating elicited by orexin-a, but not melanin-concentrating hormone, is opioid mediated. Endocrinology. 2002;143(8):2995–3000. doi: 10.1210/endo.143.8.8977. [DOI] [PubMed] [Google Scholar]
- Contaldo F, D’Arrigo E, Carandente V, Cortese C, Coltorti A, Mancini M, Taskinen MR, Nikkila EA. Short-term effects of moderate alcohol consumption on lipid metabolism and energy balance in normal men. Metabolism. 1989;38(2):166–171. doi: 10.1016/0026-0495(89)90257-6. [DOI] [PubMed] [Google Scholar]
- Duncan E, Proulx K, Woods S. Central administration of melanin-concentrating hormone increases alcohol and sucrose/quinine intake in rats. Alcohol Clin Exp Res. 2005;29(6):958–64. doi: 10.1097/01.alc.0000167741.42353.10. [DOI] [PubMed] [Google Scholar]
- Emanuele N, Lapaglia N, Emanuele M. Impact of acute and chronic ethanol exposure on prolactin in both male and female rats. Endocrine. 2001;16(1):29–37. doi: 10.1385/ENDO:16:1:29. [DOI] [PubMed] [Google Scholar]
- Ford M, Eldridge J, Samson H. Determination of an estradiol dose-response relationship in the modulation of ethanol intake. Alcohol Clin Exp Res. 2004;28(1):20–8. doi: 10.1097/01.ALC.0000108647.62718.5A. [DOI] [PubMed] [Google Scholar]
- Gaysinskaya V, Karatayev O, Chang G, Leibowitz S. Increased caloric intake after a high-fat preload: relation to circulating triglycerides and orexigenic peptides. Physiol Behav. 2007;91(1):142–53. doi: 10.1016/j.physbeh.2007.02.002. [DOI] [PubMed] [Google Scholar]
- Hammond P, Khandan-Nia N, Withers D, Jones P, Ghatei M, Bloom S. Regulation of anterior pituitary galanin and vasoactive intestinal peptide by oestrogen and prolactin status. J Endocrinol. 1997;152(2):211–9. doi: 10.1677/joe.0.1520211. [DOI] [PubMed] [Google Scholar]
- Hohmann J, Krasnow S, Teklemichael D, Clifton D, Wynick D, Steiner R. Neuroendocrine profiles in galanin-overexpressing and knockout mice. Neuroendocrinology. 2003;77(6):354–66. doi: 10.1159/000071308. [DOI] [PubMed] [Google Scholar]
- Kennedy AR, Pissios P, Otu H, Xue B, Asakura K, Furukawa N, Marino FE, Liu FF, Kahn BB, Libermann TA, Maratos-Flier E, Roberson R. A high-fat, ketogenic diet induces a unique metabolic state in mice. Am J Physiol Endocrinol Metab. 2007;292(6):E1724–39. doi: 10.1152/ajpendo.00717.2006. [DOI] [PubMed] [Google Scholar]
- Koshiyama H, Kato Y, Inoue T, Murakami Y, Ishikawa Y, Yanaihara N, Imura H. Central galanin stimulates pituitary prolactin secretion in rats: possible involvement of hypothalamic vasoactive intestinal polypeptide. Neurosci Lett. 1987;75(1):49–54. doi: 10.1016/0304-3940(87)90073-5. [DOI] [PubMed] [Google Scholar]
- Kyrkouli S, Stanley B, Leibowitz S. Galanin: stimulation of feeding induced by medial hypothalamic injection of this novel peptide. Eur J Pharmacol. 1986;122(1):159–60. doi: 10.1016/0014-2999(86)90175-5. [DOI] [PubMed] [Google Scholar]
- Leibowitz S. Overconsumption of dietary fat and alcohol: mechanisms involving lipids and hypothalamic peptides. Physiol Behav. 2007;91(5):513–21. doi: 10.1016/j.physbeh.2007.03.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leibowitz S, Akabayashi A, Alexander J, Wang J. Gonadal steroids and hypothalamic galanin and neuropeptide Y: role in eating behavior and body weight control in female rats. Endocrinology. 1998;139(4):1771–80. doi: 10.1210/endo.139.4.5867. [DOI] [PubMed] [Google Scholar]
- Leibowitz S, Akabayashi A, Wang J, Alexander J, Dourmashkin J, Chang G. Increased caloric intake on a fat-rich diet: role of ovarian steroids and galanin in the medial preoptic and paraventricular nuclei and anterior pituitary of female rats. J Neuroendocrinol. 2007;19(10):753–66. doi: 10.1111/j.1365-2826.2007.01584.x. [DOI] [PubMed] [Google Scholar]
- Leibowitz S, Avena N, Chang G, Karatayev O, Chau D, Hoebel B. Ethanol intake increases galanin mRNA in the hypothalamus and withdrawal decreases it. Physiol Behav. 2003;79(1):103–11. doi: 10.1016/s0031-9384(03)00110-0. [DOI] [PubMed] [Google Scholar]
- Leibowitz S, Dourmashkin J, Chang G, Hill J, Gayles E, Fried S, Wang J. Acute high-fat diet paradigms link galanin to triglycerides and their transport and metabolism in muscle. Brain Res. 2004;1008(2):168–78. doi: 10.1016/j.brainres.2004.02.030. [DOI] [PubMed] [Google Scholar]
- Leibowitz S, Kim T. Impact of a galanin antagonist on exogenous galanin and natural patterns of fat ingestion. Brain Res. 1992;599(1):148–52. doi: 10.1016/0006-8993(92)90863-5. [DOI] [PubMed] [Google Scholar]
- Leibowitz S, Lucas D, Leibowitz K, Jhanwar Y. Developmental patterns of macronutrient intake in female and male rats from weaning to maturity. Physiol Behav. 1991;50(6):1167–74. doi: 10.1016/0031-9384(91)90578-c. [DOI] [PubMed] [Google Scholar]
- Leibowitz SF, Akabayashi A, Alexander J, Karatayev O, Chang GQ. Puberty Onset in Female Rats: Relationship With Fat Intake, Ovarian Steroids and the Peptides, Galanin and Enkephalin, in the Paraventricular and Medial Preoptic Nuclei. Journal of Neuroendocrinology. 2009;21(6):538–549. doi: 10.1111/j.1365-2826.2009.01870.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leibowitz SF, Wortley KE. Hypothalamic control of energy balance: different peptides, different functions. Peptides. 2004;25(3):473–504. doi: 10.1016/j.peptides.2004.02.006. [DOI] [PubMed] [Google Scholar]
- Lewis M, Johnson D, Waldman D, Leibowitz S, Hoebel B. Galanin microinjection in the third ventricle increases voluntary ethanol intake. Alcohol Clin Exp Res. 2004;28(12):1822–8. doi: 10.1097/01.alc.0000148099.12344.c8. [DOI] [PubMed] [Google Scholar]
- López F, Meade EJ, Negro-Vilar A. Endogenous galanin modulates the gonadotropin and prolactin proestrous surges in the rat. Endocrinology. 1993;132(2):795–800. doi: 10.1210/endo.132.2.7678800. [DOI] [PubMed] [Google Scholar]
- Marinelli PW, Quirion R, Gianoulakis C. Estradiol valerate and alcohol intake: a comparison between Wistar and Lewis rats and the putative role of endorphins. Behav Brain Res. 2003;139(12):59–67. doi: 10.1016/s0166-4328(02)00057-8. [DOI] [PubMed] [Google Scholar]
- Marsh D, Miura G, Yagaloff K, Schwartz M, Barsh G, Palmiter R. Effects of neuropeptide Y deficiency on hypothalamic agouti-related protein expression and responsiveness to melanocortin analogues. Brain Res. 1999;848(12):66–77. doi: 10.1016/s0006-8993(99)01962-9. [DOI] [PubMed] [Google Scholar]
- Nagase H, Nakajima A, Sekihara H, York D, Bray G. Regulation of feeding behavior, gastric emptying, and sympathetic nerve activity to interscapular brown adipose tissue by galanin and enterostatin: the involvement of vagal-central nervous system interactions. J Gastroenterol. 2002;37(Suppl 14):118–27. doi: 10.1007/BF03326430. [DOI] [PubMed] [Google Scholar]
- O’Halloran D, Jones P, Ghatei M, Bloom S. Rat anterior pituitary neuropeptides following chronic prolactin manipulation: a combined radioimmunoassay and mRNA study. J Endocrinol. 1991;131(3):411–9. doi: 10.1677/joe.0.1310411. [DOI] [PubMed] [Google Scholar]
- Ottlecz A, Snyder GD, McCann SM. Regulatory role of galanin in control of hypothalamic-anterior pituitary function. Proc Natl Acad Sci U S A. 1988;85(24):9861–9865. doi: 10.1073/pnas.85.24.9861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paxinos G, Franklin KBJ. The mouse brain in stereotaxic coordinates. Academic Press; San Diego, Calif: 2001. pp. 92101–0095. [Google Scholar]
- Plaisier C, Kyttälä M, Weissglas-Volkov D, Sinsheimer J, Huertas-Vazquez A, Riba L, Ramírez-Jiménez S, de Bruin T, Tusié-Luna T, Aouizerat B, Pullinger C, Malloy M, Kane J, Cruz-Bautista I, Herrera M, Aguilar-Salinas C, Kuusisto J, Laakso M, Taskinen M, van der Kallen C, Pajukanta P. Galanin preproprotein is associated with elevated plasma triglycerides. Arterioscler Thromb Vasc Biol. 2009;29(1):147–52. doi: 10.1161/ATVBAHA.108.178533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porkka-Heiskanen T, Kalinchuk A, Alanko L, Huhtaniemi I, Stenberg D. Orexin A and B levels in the hypothalamus of female rats: the effects of the estrous cycle and age. Eur J Endocrinol. 2004;150(5):737–42. doi: 10.1530/eje.0.1500737. [DOI] [PubMed] [Google Scholar]
- Qian S, Chen H, Weingarth D, Trumbauer M, Novi D, Guan X, Yu H, Shen Z, Feng Y, Frazier E, Chen A, Camacho R, Shearman L, Gopal-Truter S, MacNeil D, Van der Ploeg L, Marsh D. Neither agouti-related protein nor neuropeptide Y is critically required for the regulation of energy homeostasis in mice. Mol Cell Biol. 2002;22(14):5027–35. doi: 10.1128/MCB.22.14.5027-5035.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rada P, Avena N, Leibowitz S, Hoebel B. Ethanol intake is increased by injection of galanin in the paraventricular nucleus and reduced by a galanin antagonist. Alcohol. 2004;33(2):91–7. doi: 10.1016/j.alcohol.2004.05.002. [DOI] [PubMed] [Google Scholar]
- Rajasingh J, Bord E, Qin G, Ii M, Silver M, Hamada H, Ahluwalia D, Goukassian D, Zhu Y, Losordo D, Kishore R. Enhanced voluntary alcohol consumption after estrogen supplementation negates estrogen-mediated vascular repair in ovariectomized mice. Endocrinology. 2007;148(8):3618–24. doi: 10.1210/en.2006-1357. [DOI] [PubMed] [Google Scholar]
- Reid LD, Marinelli PW, Bennett SM, Fiscale LT, Narciso SP, Oparowski CJ, Reid ML, Merrigan BA, Moricone J, Hubbell CL, Gianoulakis C. One injection of estradiol valerate induces dramatic changes in rats’ intake of alcoholic beverages. Pharmacol Biochem Behav. 2002;72(3):601–16. doi: 10.1016/s0091-3057(02)00732-3. [DOI] [PubMed] [Google Scholar]
- Rossmanith W, Marks D, Clifton D, Steiner R. Induction of galanin mRNA in GnRH neurons by estradiol and its facilitation by progesterone. J Neuroendocrinol. 1996;8(3):185–91. doi: 10.1046/j.1365-2826.1996.04473.x. [DOI] [PubMed] [Google Scholar]
- Salonen I, Huhtaniemi I. Effects of chronic ethanol diet on pituitary-testicular function of the rat. Biol Reprod. 1990;42(1):55–62. doi: 10.1095/biolreprod42.1.55. [DOI] [PubMed] [Google Scholar]
- Sanchis R, Esquifino A, Guerri C. Chronic ethanol intake modifies estrous cyclicity and alters prolactin and LH levels. Pharmacol Biochem Behav. 1985;23(2):221–4. doi: 10.1016/0091-3057(85)90560-x. [DOI] [PubMed] [Google Scholar]
- Schneider ER, Rada P, Darby RD, Leibowitz SF, Hoebel BG. Orexigenic peptides and alcohol intake: Differential effects of orexin, galanin, and ghrelin. Alcoholism-Clinical and Experimental Research. 2007;31(11):1858–1865. doi: 10.1111/j.1530-0277.2007.00510.x. [DOI] [PubMed] [Google Scholar]
- Schrezenmeir J. Hyperinsulinemia, hyperproinsulinemia and insulin resistance in the metabolic syndrome. Experientia. 1996;52(5):426–432. doi: 10.1007/BF01919311. [DOI] [PubMed] [Google Scholar]
- Sweet DC, Levine AS, Billington CJ, Kotz CM. Feeding response to central orexins. Brain Res. 1999;821(2):535–538. doi: 10.1016/s0006-8993(99)01136-1. [DOI] [PubMed] [Google Scholar]
- Viale A, Kerdelhué B, Nahon J. 17beta-estradiol regulation of melanin-concentrating hormone and neuropeptide-E-I contents in cynomolgus monkeys: a preliminary study. Peptides. 1999;20(5):553–9. doi: 10.1016/s0196-9781(99)00007-8. [DOI] [PubMed] [Google Scholar]
- Wortley K, Chang G, Davydova Z, Leibowitz S. Peptides that regulate food intake: orexin gene expression is increased during states of hypertriglyceridemia. Am J Physiol Regul Integr Comp Physiol. 2003;284(6):R1454–65. doi: 10.1152/ajpregu.00286.2002. [DOI] [PubMed] [Google Scholar]
- Wynick D, Small C, Bacon A, Holmes F, Norman M, Ormandy C, Kilic E, Kerr N, Ghatei M, Talamantes F, Bloom S, Pachnis V. Galanin regulates prolactin release and lactotroph proliferation. Proc Natl Acad Sci U S A. 1998;95(21):12671–6. doi: 10.1073/pnas.95.21.12671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yun R, Dourmashkin JT, Hill JO, Gayles EC, Fried SK, Leibowitz SF. PVN galanin increases fat storage and promotes obesity by causing muscle to utilize carbohydrate more than fat. Peptides. 2005;26(11):2265–2273. doi: 10.1016/j.peptides.2005.04.005. [DOI] [PubMed] [Google Scholar]
- Zorrilla E, Brennan M, Sabino V, Lu X, Bartfai T. Galanin type 1 receptor knockout mice show altered responses to high-fat diet and glucose challenge. Physiol Behav. 2007;91(5):479–85. doi: 10.1016/j.physbeh.2006.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]