

Abstract
The review focuses on the central neuronal circuits involved in energy homeostasis and the opportunities these offer for pharmacological intervention to decrease feeding behaviour and reduce weight. This article is based on the presentation ‘New central targets for the treatment of obesity’ (Sargent, British Pharmacological society, Clinical Section Symposium, December 2008).
Central neuronal substrates controlling weight offer numerous opportunities for pharmacological intervention. These opportunities range from non-specific enhancement of monoamine signalling (triple reuptake inhibitors) to targeting specific monoamine receptor subtypes (5-HT2c and 5-HT6). The data reviewed suggest that these approaches will lead to weight loss; whether this is sufficient to produce clinically meaningful effect remains to be determined. Combination therapy targeting more than one mechanism may be a means of increasing the magnitude of the response. Preclinical studies also suggest that novel approaches targeting specific neuronal pathways within the hypothalamus, e.g. MCH1 receptor antagonism, offer an opportunity for weight reduction. However, these approaches are at an early stage and clinical studies will be needed to determine if these novel approaches lead to clinically meaningful weight loss and improvements in co-morbid conditions such as diabetes and cardiovascular disorders.
Keywords: 5-HT2c, MCH1, neuropeptides, obesity, sibutramine, weight loss agents
The obesity problem
Obesity is a major global health problem and rates of obesity [body mass index (BMI) >30 kg m−2] have risen steadily year on year. Recent figures show an incidence of obesity of more than 1 in 4 in many parts of the USA [1]. These high rates are similar in other developed countries and the incidence of weight gain is increasing in less developed regions. Obesity is not just a cosmetic or lifestyle illness but leads to many life-threatening health complications. As the BMI increases the incidence of metabolic disorders (Type 2 diabetes), hypertension, cancer and musculoskeletal problems increases [1–3]. In 1999, Calle et al. reported mortality rates for all causes doubled when the BMI increased from 25 to >35 [4]. The annual direct healthcare costs associated with obesity in the USA are in excess of $93 billion [5]. Particularly worrying is the fact that the incidence of obesity in children and young adults is on the increase, with some reports suggesting that up to 25% of children may be obese. The long-term health implications of these findings are far reaching and suggest that for the first time intergenerational increase in life expectancy will not occur and these children will have a shorter life span than their parents because of health complications associated with weight gain [6] (Figure 1).
Figure 1.
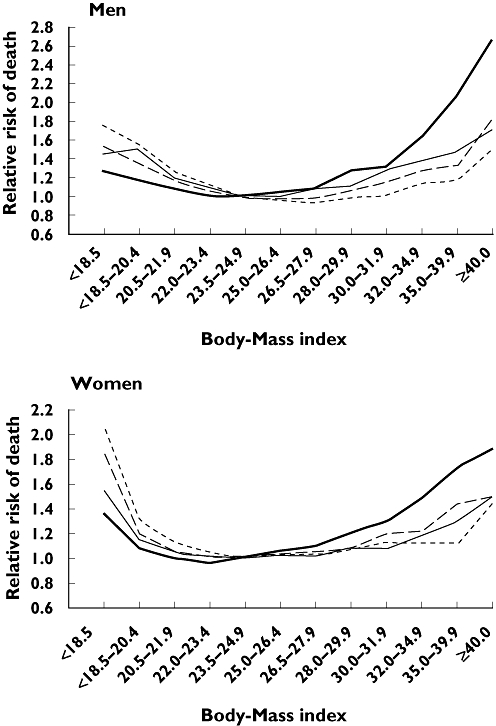
Multivariate relative risk of death from all causes among men and women according to body mass index, smoking status, and disease status. The four subgroups are mutually exclusive. Nonsmokers had never smoked. The reference category was made up of subjects with a body mass index of 23.5–24.9. Calle et al. N Engl J Med 1999; 341: 1097–105. Current or former smokers with a history of disease (----); Current or former smokers with on history of disease (– –); Nonsmokers with a history of disease (—); Nonsmokers with no history of disease ( )
)
Obesity occurs when the balance between energy input (food intake) and energy expenditure (exercise, activity) is disrupted, i.e. more food is consumed than utilized, leading to excess fat stores being laid down. Although for some people a calorie-restricted diet and exercise can lead to a reduction in weight gain, for many people this is not enough and weight loss cannot be maintained. There are many environmental factors that predispose individuals to gain weight, e.g. freely available high-calorie food and sedentary life style [7]. Genetic factors also contribute to this imbalance; for example, the ‘Thrifty-gene’ hypothesis proposes that humans particularly prone to famine have evolved to favour the storage of excess calories in the form of fat [7]. In the severely obese, surgical intervention may be necessary (see review in this supplement). An alternative approach is to develop therapeutic agents that can either reduce food consumption or increase energy utilization.
This review is based on the presentation ‘New central targets for the treatment of obesity’ (Sargent, British Pharmacological society, Clinical Section Symposium December 2008) and focuses on central neuronal circuits involved in energy homeostasis and the opportunities these offer for pharmacological intervention to reduce feeding behaviour and reduce weight gain. The review is not intended as an in-depth analysis of each individual mechanism, but will highlight the breadth of approaches and direct readers to other recent reviews and articles that address these approaches in much more detail.
Hypothalamus and the central control of feeding and energy homeostasis
The hypothalamus is the key processing area within the brain for the integration of numerous signals related to energy homeostasis. Peripheral signals of satiety are integrated and passed on to other areas of the brain via nuclei in the hypothalamus. All agents that act to reduce food intake and weight gain will directly or indirectly influence activity within the hypothalamus. Peripheral signals from adipose stores, the gastrointestinal tract and endocrine system influence the activity of neurons within the arcuate nucleus of the hypothalamus. When fat stores are reduced and energy levels are low, hunger signals mediated via an increase in the gut hormone, ghrelin, and reductions in insulin, glucose, leptin and cholecystokinin (CCK) cause increases in the activity of both neuropeptide Y (NPY) and agouti-related protein (AgRP) neurons, which in turn leads to decreased activity of the melanocortin system, leading to disinhibition of melanin-concentrating hormone (MCH) and orexin (ORX) signalling producing a marked orexigenic effect. Following a meal the reverse occurs, with high levels of glucose, insulin, CCK and reduced ghrelin levels leading to increases in pro-opiomelanocortin (POMC) and cocaine-amphetamine regulated transcript (CART) neuronal activity, in turn increasing α-melanocyte-stimulating hormone (α-MSH) release and decreasing MCH/ORX activity, leading to satiation and a termination of feeding (Figure 2) [8–21].
Figure 2.
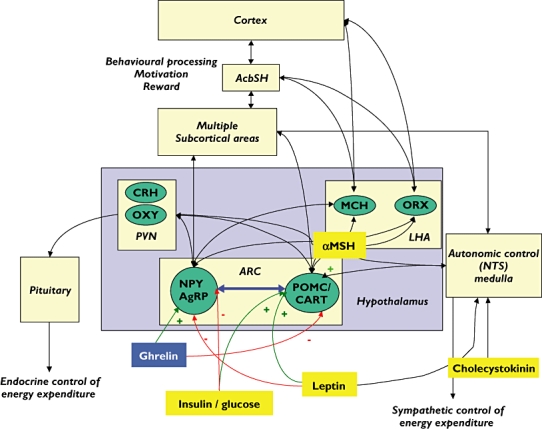
Circuits implicated in the central control of feeding. AcbSH, nucleus accumbens shell; AgRP, agouti gene-related peptide; ARC, arcuate nucleus; CART, cocaine-amphetamine regulated transcript; CCK, cholecystokinin; CRH, corticotrophin releasing hormone; LHA, lateral hypothalamus; α-MSH, alpha-melanocyte stimulating hormone; MCH, melanin-concentrating hormone; MC4R, melanocortin 4 receptor; NPY, neuropeptide Y; NTS, tractus solitari; ORX-orexin (hypocretin); OXY, oxytocin; POMC, pro-opiomelanocortin; PVN, periventricular nucleus
Centrally acting pharmacological treatments of obesity
A number of central agents have been developed for the treatment of obesity [21, 22]. These agents produced their actions through the modification of biogenic amine function [17, 23]. The first agents used were amphetamine-like compounds, and although they produced significant weight loss, the stimulant properties and abuse liability led to their demise. Analogues that did not have the dopamine reuptake/releasing properties were subsequently developed, culminating in the development of the serotonin releaser, d-fenfluramine and the sympathomimetic, phentermine. These compounds, particularly when used in combination (fen/phen) produced significant weight loss. d-Fenfluramine produces its action via the release of 5-HT plus direct stimulation of 5-HT2C receptors located in the hypothalamus. The increase in serotonin within the hypothalamus activates POMC neurons, via the 5-HT2C receptor. The POMC neurons play a key role in the regulation of feeding by sending anorectic signals to the periventricular nucleus (PVN) and other brain areas associated with energy homeostasis. Fenfluramine and the fen/phen combination were withdrawn following a number of cases of pulmonary hypertension and cardiac valvulopathy [17, 21].
One compound, acting via monoamine reuptake inhibition, currently marketed is sibutramine (Table 1). Sibutramine is a serotonin/noradrenaline reuptake inhibitor that produces robust weight loss in rodent models of obesity. Sibutramine acts by increasing noradrenaline and serotonin levels in the hypothalamus, nucleus accumbens and the brainstem, all regions associated with energy homeostasis [23]. The desmethyl metabolites of sibutramine have significant dopamine reuptake inhibition, and this may contribute to the overall profile of the compound in vivo[24, 25]
Table 1.
Centrally acting marketed or compounds in clinical development for the treatment of obesity
| Company | Product name/code | Product type | Phase |
|---|---|---|---|
| Abbott | Sibutramine | NA/5HT reuptake inhibitor | Authorized |
| Arena Pharma | Lorcaserin | 5HT2C agonist | Phase III |
| Athersys | ATHX-105 | 5HT2C agonist | Phase I |
| Biovitrum | BVT.74316 | 5HT6 antagonist | Phase I |
| Dov Pharmaceuticals | Dov 21947 | NA/DA/5HT reuptake inhibitor | Phase II |
| EPIX Pharma | PRX-07034 | 5HT6 antagonist | Phase I |
| NeuroSearch | Tesofensine | NA/DA/5HT reuptake inhibitor | Phase II |
| Orexigen | Contrave | Bupropion + naltrexone | Phase III |
| Orexigen | Empatic | Bupropion + zonisamide | Phase II |
| Shionogi | S-2367 | Neuropeptide Y5 inhibitor | Phase II |
| Vivus | Qnexa | Phentermine + topiramate | Phase II |
In the clinic, sibutramine causes weight loss, of about 4% compared with placebo, over the course of 12–18 months. Sibutramine also increased the percentage of patients achieving at least a 5 or 10% weight reduction. Sibutramine-induced weight loss was also accompanied by a significant reduction in high-density lipoprotein-cholesterol and triglycerides, but raised heart rate and blood pressure [26].
A recent report has questioned the utility of currently available antiobesity agents on both an efficacy and cost basis. In the survey, payers stated a threshold of about 18% weight loss from placebo would influence formulary coverage and reimbursement [27].
It is clear that additional compounds with superior efficacy and improved safety are required.
New pharmacological approaches to the regulation of food intake and weight loss
Monoamine reuptake inhibitors
Monoamine regulation of food intake remains an area of great interest. Astrup et al. recently reported Phase II clinical data on tesofensine. Tesofensine is a triple reuptake inhibitor that blocks the reuptake of noradrenaline, serotonin and dopamine [28]. Tesofensine, combined with diet, produced approximately a 10% (12.8 kg) reduction in weight at the highest dose tested (1 mg) compared with a 2% (2.2 kg) weight loss in the placebo group. However, this dose was also associated with a significant increase in heart rate and blood pressure. Lower doses (0.25–0.5 mg) produced a 4.5–9% decrease in body weight with no associated change in blood pressure. A slight tachycardia was noted with the 0.5-mg dose. Dov Pharmaceuticals are also developing a compound that acts by inhibiting the reuptake of dopamine, serotonin and noradrenaline. Dov 21947 at doses up to 150 mg has been reported to produce a significant decrease in body weight of 2.1 kg compared with the placebo group in an 8-week study [29].
5-HT2C agonists as potential obesity treatments
Studies using serotonin reuptake inhibitors and selective serotonin agonists and antagonists have clearly implicated serotonin in the regulation of feeding and energy homeostasis. Studies in animals using both transgenic knock-out animals and selective agonists show that stimulation of the 5-HT2C receptor decreases food intake and body weight in both lean and obese rodents. Stimulation of 5-HT2C receptors has been implicated in the hypophagic action of fenfluramine. Many 5-HT2C agonists have been developed over the years; however, selectivity over other 5-HT2 receptors has remained a major challenge. Stimulation of 5-HT2C receptors produces a hypophagic effect and blockade of the 5-HT2C receptor has been implicated in drug-induced weight gain [30]. Stimulation of 5-HT2A receptors produces unwanted hallucinogenic-like effects and activity at the 5-HT2B receptor implicated in valvulopathy and pulmonary hypertension limiting the use of relatively nonselective agents [21].
5-HT2C agonists are thought to exert their hypophagic actions via stimulation of receptors located on POMC containing neurons within the arcuate (ARC) nucleus. Recent transgenic data add support to the proposal that 5-HT2C receptors on POMC neurons play a crucial role in the regulation of feeding. Mice lacking the 5-HT2C receptor developed hyperphagia and weight gain and showed an attenuated response to the anorexigenic 5-HT2C agonist, 1-(m-chlorophenyl)piperazine (mCPP). Selective re-introduction of 5-HT2C receptors on POMC containing neurons reversed the KO-induced deficits and normalized both feeding and weight gain [30–33].
Lorcaserin (ADP359) is a selective 5-HT2C agonist currently in clinical development for obesity. In a Phase II study subjects with a BMI = 30–45 received lorcaserin (10 mg q.d., 15 mg q.d. or 10 mg b.i.d.) for 12 weeks. Lorcaserin produced a dose-related increase in the number of patients displaying a >5% weight loss. For example, only 2.3% of patients on placebo demonstrated >5% weight loss, whereas 31% of patients receiving 10 mg b.i.d. showed >5% weight loss. Fasting blood sugar, cholesterol and waist circumference were also reduced. Adverse events were modest and no increased incidence of valvulopathy or pulmonary hypertension was noted [34].
5-HT6 antagonists as potential obesity targets
The 5-HT6 receptor has recently attracted attention as a potential target for the treatment of obesity [35]. 5-HT6 KO animals demonstrated a resistance to weight gain when placed on a high-fat diet. In another series of experiments, it was found that antisense oligonucleotides directed at the 5-HT6 receptor decreased body weight and food intake in rats. Woolley et al. [36] also observed that a high dose of the 5-HT6 antagonist Ro 04-6790 significantly attenuated body weight gain in growing rats when administered for 3 days. Since these original studies, a number of others have reported reductions in food intake following 5-HT6 antagonist administration [37]. The exact mechanism of 5-HT6-mediated reductions in food intake is unclear. However, 5-HT6 receptor mRNA is found in the ARC and it has been proposed that the blockade of 5-HT6 receptors reduces GABAergic inhibitory control of POMC neurons and the subsequent increase in the release of α-MSH and stimulation of MC-4 receptors leading to a reduction in food intake [35].
It should be noted that in most cases relatively high doses of 5-HT6 antagonists are required to influence food intake compared with doses efficacious in cognition models. This may suggest that the overall effect on food intake is mediated via 5-HT6 antagonism plus additional actions.
Cannabinoid antagonists as antiobesity agents
There is substantial evidence that endocannabinoids are involved in the control of appetite and body weight, and these effects appear to be mediated via the CB1 receptor [38, 39]. Tetrahydrocannabinol and cannabis both increase high fat and highly palatable diet intake. The endogenous cannabinoid anandamide has also been shown to increase food intake when injected into the brain, while CB1 KO mice show reduced food intake and weight gain. The highly selective CB1 antagonist rimonobant inhibits intake of palatable foods by marmosets and reduces intake of high-fat diet in rodents. However, rimonobant has no effect on normal laboratory chow intake. Pair feeding studies also show that the weight loss produced by CB1 antagonists is greater than that predicted based on reduction in calories consumed alone, suggesting that CB1 antagonists increase energy expenditure in addition to reducing food intake [21, 40].
The central actions of endocannabinoids on food intake are mediated by direct action within the hypothalamus. Endocannabinoids are under negative control by leptin, the adipocyte hormone that inhibits orexigenic signalling in the hypothalamus. In genetically obese animals that have disrupted leptin function, the endocannabinoids system is found to be overactive. Activation of CB1 receptors within the hypothalamus results in the disinhibition of MCH release, producing an orexigenic action [21].
Rimonobant and a number of other CB1 antagonists (taranabant, otenabant, surinabant and ibipinabant) have been taken into clinical development and shown to produce significant decreases in body weight. In a recent meta-analysis, rimonobant was shown to produce a 4.7-kg reduction in body weight after 1 year, an effect size similar to that observed for sibutramine. Rimonobant also significantly reduced placebo subtracted waist circumference, blood pressure and triglyceride concentrations [41]. The CB1 antagonist significantly increased the incidence of psychiatric adverse events (notably, depression, anxiety, irritability and aggression). These adverse events led to the withdrawal of rimonobant and termination of the development of several other CB1 antagonists for obesity [42].
Combination pharmacotherapy as an alternative approach
Clinical experience with existing treatments suggests that weight loss produced by any one agent is limited and rarely exceeds 10% of the starting weight. This initial drop also plateaus. The ‘ceiling effect’ observed is probably due to adaptive changes as the body continues to maintain a balance in favour of energy conservation. One approach that some have adopted to overcome this issue is to develop combination therapies that tackle the problem through more than one mechanism, thus minimizing the impact of adaptive changes.
The noradrenaline/dopamine reuptake inhibitor, bupropion, when combined with the opiate antagonist naltrexone, has been reported to produce a significant loss of 9.3–11.5% of baseline body weight in a recent the Phase III trial. Twice as many subjects lost ≥10% weight compared with the placebo group. The mechanism of action of the bupropion/naltrexone combinations is thought to be through the dual action of monoamine elevation enhancing POMC neuronal activation accompanied by a blockade of the natural endorphin negative feedback that inhibits α-MSH release [43]. Bupropion has also been combined with zonisamide, the anticonvulsant compound, to augment the bupropion-induced weight loss [43].
Another combination therapy in development is phentermine and the anticonvulsant, topiramate (Qnexa). In a Phase II trial, the combination treatment produced significantly more weight loss than either compound alone. In a recently reported Phase III study, the percentage of patients achieving >10% loss was 66% for the combination treatment compared with 7% in the placebo group. Additional analysis showed a significant control of blood sugar in these nondiabetic subjects compared with the placebo group. Subjects receiving active treatment had a significant improvement in glycaemic control as measured by a reduction in haemoglobin A1c compared with placebo [44].
Direct modulation on neuropeptide transmitters within the hypothalamus
Within the hypothalamus, a number of peptide transmitters are involved in the integration of peripheral satiation and hunger signals and have either orexigenic or anorexigenic effects. Pharmacological manipulation of these targets offers another approach to regulating food intake [11].
Neuropeptide Y antagonists
NPY is a 36 amino acid neurotransmitter widely distributed within the brain. The arcuate nucleus has a large concentration of NPY-containing neurons that then innervate the PVN and lateral hypothalamus. NPY actions are mediated via multiple receptors (NPY1–6). Studies using the peptide have shown that administration of NPY produces marked stimulation of feeding and energy expenditure by activating NPY1 and 5 receptors within the hypothalamus [45, 46]. NPY levels are regulated by leptin and other peripheral modulators (e.g. gut hormones); following a meal when leptin levels are low, NPY activity is reduced [47]. Conversely, NPY levels are elevated during hunger and energy depletion [21]. Selective NPY antagonists, particularly targeting NPY5 receptors, inhibit feeding and reduce body weight in models of obesity [46]. The Y5 antagonist MK0557 was recently shown to induce only modest weight loss (3.4 kg) compared with placebo (1.8 kg) in a 52-week study. These results led the authors to conclude that Y5 receptor activation is involved in energy homeostasis in man; however, selective blockade of this receptor alone was insufficient to produce clinically meaningful weight loss [47].
Shionogi recently reported positive effects of their NPY5 antagonist, S-2367, in a Phase IIa proof of concept study. The safety and efficacy of S-2367 was assessed following once-daily treatment in a population of 342 obese subjects. Over the 12-week treatment period, subjects in the high-dose group of S-2367 lost an average of 5.3 kg over 16 weeks, or 5.6% of baseline weight, vs. 2.5 kg or 2.7% of baseline weight for placebo. In addition, there was also a clear dose–response between the 400 mg day−1 and 1600 mg day−1 dosing groups. S-2367 was well tolerated, and no significant or serious safety adverse events occurred [48]. Further long-term studies will be necessary to establish if this weight reduction is clinically relevant.
Melanocortin MC-4 receptor agonists
Decreased activity of the melanocortin system is associated with hunger and the initiation of feeding. α-MSH released by POMC neurons acts on MC4 receptors to exert an inhibitory effect on the secondary ‘leptin-sensitive’ MCH and ORX pathways. Injections of MC4 agonists decrease food intake, whereas inhibition of MC4 receptor activity by infusion of an MC receptor antagonist or with the inverse agonist AgRP results in increased food intake. MC4 activity affects meal size and meal choice, but not meal frequency. The type of diet also affects the efficacy of MC4 agonists to reduce food intake in rats [15, 49, 50].
Mutations in the human melanocortin MC4 receptor have been associated with obesity, which underscores the relevance of this receptor as a drug target to treat obesity. A number of groups have looked at the development of selective agonists for MC4 as potential treatments for obesity. However, the lack of suitable small-molecular-weight nonpeptide agonist ligands or adverse events have limited clinical development of this target [15, 49, 50].
Melanin concentrating hormone antagonists
ARC NPY-AgRP and POMC neurons project to brain regions involved in energy homeostasis, including the PVN, LHA and nucleus accumbens shell. These so-called ‘leptin-sensitive’ neurons express significant amounts of MCH. MCH has been identified as an orexigenic peptide that exerts an effect on food intake and body weight regulation [51, 52]. MCH is a cyclic 19 amino acid neuropeptide expressed in the lateral hypothalamus in response to both energy restriction and leptin deficiency. MCH is known to stimulate feeding when injected into the lateral ventricle of rats, and the mRNA for MCH is upregulated in the hypothalamus of genetically obese mice (ob/ob) and in fasted control animals. In addition, animals treated with MCH show increases in glucose, insulin and leptin levels, mimicking human metabolic syndrome. Mice lacking MCH receptors are hypophagic and lean with increased metabolic rate, whereas animals overexpressing MCH gain excess weight on both standard and high-fat diets. MCH exerts an orexigenic effects via activation of MCH1 receptors. Indeed, a variety of small-molecule, MCH1 antagonists have been shown to produce significant weight loss in mouse and rat models of obesity. These compounds have been shown to produce weight loss by reducing meal size [53, 54].
Despite the discovery and preclinical development of many selective MCH1 receptor antagonists, very few have entered clinical development. One of the main reasons for the lack of clinical development has been the propensity for selective agents to have significant cardiovascular liabilities. Many of the early compounds displayed significant human ether-a-go-go-related gene-binding activity and drug-induced QTc prolongation [52].
Despite the significant challenges encountered by many groups, MCH1 receptor antagonism remains a very attractive target for obesity. Recently a number of compounds have been reported that possess significant MCH1 antagonist activity and cause marked weight loss in animals, without producing changes in cardiovascular function associated with other, older MCH1 antagonists. For example, AMR-MCH-1 was found to bind to the human MCH1 receptor with a Ki value of 2.6 nM and demonstrated significant and sustained reductions in food intake and body weight in a chronic, 28-day feeding study in male dietary-induced obese C57BL/6J mice. At 30 mg kg−1 and 60 mg kg−1 (po b.i.d.), AMR-MCH-1 produced weight losses of 11.1% and 13.9%, respectively, compared with 5.8% for positive control sibutramine (20 mg kg−1 po q.d.). Fat pad analysis indicated that the weight loss caused by AMR-MCH-1 was associated with reductions in fat mass of 27.5% and 44.6% compared with vehicle. These data indicate that AMR-MCH-1 is a high-affinity MCH1 receptor antagonist that causes sustained weight loss [55].
Another MCH1 antagonist, NGD-4715, entered Phase I clinical testing; the initial phase of the multiple ascending dose (MAD) study utilized three times per day dosing for 14 days in healthy obese subjects exposed to a high caloric diet. During the initial phase of this MAD study, induction of the liver enzyme CYP 3A4 occurred and a lipid-lowering effect was also observed. A follow-up study designed to test twice per day dosing in healthy obese subjects on a restricted caloric diet for 14 days was also conducted. Using b.i.d. dosing, CYP3A4 induction was substantially reduced. With less frequent dosing and caloric restriction, no effect on lipids was observed. As in the first Phase I study, no serious adverse events were observed. However, vivid dreams and awakenings were reported by half the drug-treated subjects during the first week of dosing. Development of this compound has subsequently been terminated [56].
Conclusions
Central neuronal substrates of feeding and energy balance offer numerous opportunities for pharmacological intervention. These opportunities range from nonspecific enhancement of monoamine signalling (triple reuptake inhibitors) to targeting specific monoamine receptor subtypes (5-HT2c and 5-HT6). To date, the evidence shows that these approaches may lead to statistically significant weight loss, but whether this is sufficient to produce clinically meaningful weight reduction remains to be determined. Combination therapy targeting more than one pathway/mechanism may be a means of increasing the magnitude of the response.
Based on animal data, novel approaches targeting specific neuronal pathways within the hypothalamus, e.g. MCH1 receptor antagonism, offer an opportunity for weight reduction. However, these approaches are at an early stage and clinical studies will be needed to determine if these novel approaches lead to clinically meaningful weight loss and improvements in comorbid conditions such as diabetes and cardiovascular disorders.
Competing interests
None to declare.
We would like to thank P. Guzzo, M. Surman, A. Henderson and M. Luche for their helpful comments and suggestions.
REFERENCES
- 1.Center for Disease Control and Prevention. Available at http://www.cdc.gov/nccdphp/dnpa/obesity/trend/maps/ (last accessed.
- 2.Poirier P, Eckel RH. Cardiovascular consequences of obesity. Drug Discov Today Ther Strat. 2008;5:45–51. doi: 10.1016/j.ddstr.2008.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mittendorfer B, Peterson LR. Cardiovascular consequences of obesity and targets for treatment. Drug Discov Today Ther Strat. 2008;5:53–61. doi: 10.1016/j.ddstr.2008.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Calle EE, Thun MJ, Petrelli JM, Rogriguez C, Heath CW. Body-mass index and mortality in a prospective cohort of U.S. adults. N Engl J Med. 1999;341:1097–105. doi: 10.1056/NEJM199910073411501. [DOI] [PubMed] [Google Scholar]
- 5.Finkelstein EA, Fiebelkorn IC, Wang G. National medical spending attributable to overweight and obesity: how much, and who's paying? Health Affairs. 2003;W3:219–26. doi: 10.1377/hlthaff.w3.219. [DOI] [PubMed] [Google Scholar]
- 6.Daniels SR. The consequences of childhood overweight and obesity. Future Child. 2006;16:47–67. doi: 10.1353/foc.2006.0004. [DOI] [PubMed] [Google Scholar]
- 7.Bloom SR, Kuhajda FP, Laher I, Pi-Sunyer X, Ronnett GV, Tan TMM, Weigle DS. The obesity epidemic: pharmacological challenges. Mol Interv. 2008;8:82–98. doi: 10.1124/mi.8.2.6. [DOI] [PubMed] [Google Scholar]
- 8.Blundell JE, Levin F, King NA, Barkeling B, Gustafson T, Hellstrom PM, Holst JJ, Naslund E. Overconsumption and obesity: peptides and susceptibility to weight gain. Regul Pept. 2008;149:32–8. doi: 10.1016/j.regpep.2007.10.009. [DOI] [PubMed] [Google Scholar]
- 9.Schwartz MW, Woods SC, Porte D, Seeley RJ, Baskin DG. Central nervous system control of food intake. Nature. 2000;404:661–71. doi: 10.1038/35007534. [DOI] [PubMed] [Google Scholar]
- 10.Woods SC, D'Alessio DA. Central control of body weight and appetite. J Clin Endocrinol Metab. 2008;93:S37–50. doi: 10.1210/jc.2008-1630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Abizaid A, Horvath TL. Brain circuits regulating energy homeostasis. Regul Pept. 2008;149:3–10. doi: 10.1016/j.regpep.2007.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hameed S, Dhillo WS, Bloom SR. Gut hormones and appetite control. Oral Dis. 2009;15:18–26. doi: 10.1111/j.1601-0825.2008.01492.x. [DOI] [PubMed] [Google Scholar]
- 13.Knecht S, Ellger T, Levine JA. Obesity in neurobiology. Prog Neurobiol. 2008;84:85–103. doi: 10.1016/j.pneurobio.2007.09.003. [DOI] [PubMed] [Google Scholar]
- 14.Valassi E, Scacchi M, Cavagnini F. Neuroendocrine control of food intake. Nutr Metab Cardiovasc Dis. 2008;18:158–68. doi: 10.1016/j.numecd.2007.06.004. [DOI] [PubMed] [Google Scholar]
- 15.Wikberg JES, Mutulis F. Targeting melanocortin receptors: an approach to treat weight disorders and sexual dysfunction. Nat Rev Drug Discov. 2008;7:307–23. doi: 10.1038/nrd2331. [DOI] [PubMed] [Google Scholar]
- 16.Adamantidis A, de Lecea L. Sleep and metabolism: shared circuits, new connections. Trends Endocrinol Metab. 2008;19:362–70. doi: 10.1016/j.tem.2008.08.007. [DOI] [PubMed] [Google Scholar]
- 17.Adan RAH, Vanderschuren LJMJ, la Fleur SE. Anti-obesity drugs and neural circuits of feeding. Trends Pharmacol Sci. 2008;29:208–17. doi: 10.1016/j.tips.2008.01.008. [DOI] [PubMed] [Google Scholar]
- 18.Moran TH, Aja S, Ladenheim EE. Leptin modulation of peripheral controls of meal size. Physiol Behav. 2006;89:511–6. doi: 10.1016/j.physbeh.2006.04.020. [DOI] [PubMed] [Google Scholar]
- 19.Coll AP, Farooqi IS, O'Rahilly S. The hormonal control of food intake. Cell. 2007;129:251–62. doi: 10.1016/j.cell.2007.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Benoit SC, Tracy AL, Davis JF, Choi D, Clegg DJ. Novel functions of orexigenic hypothalamic peptides: from genes to behavior. Nutrition. 2008;24:843–7. doi: 10.1016/j.nut.2008.06.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bray GA, Greenway FL. Pharmacological treatment of the overweight patient. Pharmacol Rev. 2007;59:151–84. doi: 10.1124/pr.59.2.2. [DOI] [PubMed] [Google Scholar]
- 22.Clapham JC, Arch JRS, Tadayyon M. Anti-obesity drugs: a critical review of current therapies and future opportunities. Pharmacol Ther. 2001;89:81–121. doi: 10.1016/s0163-7258(00)00105-4. [DOI] [PubMed] [Google Scholar]
- 23.Clifton PG, Kennett GA. Monoamine receptors in the regulation of feeding behaviour and energy balance. CNS Neurol Disord Drug Targets. 2006;5:293–312. doi: 10.2174/187152706777452254. [DOI] [PubMed] [Google Scholar]
- 24.Billes SK, Cowley MA. Inhibition of dopamine and norepinephrine reuptake produces additive effects on energy balance in lean and obese mice. Neuropsychopharmacology. 2007;32:822–34. doi: 10.1038/sj.npp.1301155. [DOI] [PubMed] [Google Scholar]
- 25.Glick SD, Haskew RE, Maisonneuve IM, Carlson JN, Jerussi TP. Enantioselective behavioral effects of sibutramine metabolites. Eur J Pharmacol. 2000;397:93–102. doi: 10.1016/s0014-2999(00)00216-8. [DOI] [PubMed] [Google Scholar]
- 26.Rucker D, Padwal R, Li SK, Curioni C, Lau DCW. Long term pharmacotherapy for obesity and overweight: updated meta-analysis. BMJ. 2007;335:1194–9. doi: 10.1136/bmj.39385.413113.25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Greenapple R. Available at http://www.fiercepharma.com/special-reports/can-obesity-treatment-be-cost-effective (last accessed 15 October 2009)
- 28.Astrup A, Madsbad S, Breum L, Jensen TJ, Kroustrup JP, Larsen TM. Effect of tesofensine on bodyweight loss, body composition, and quality of life in obese patients: a randomised, double-blind, placebo-controlled trial. Lancet. 2008;372:1859–60. doi: 10.1016/S0140-6736(08)61525-1. [DOI] [PubMed] [Google Scholar]
- 29. Company information. Available at http://www.medicalnewstoday.com/articles/83621.php (last accessed 15 October 2009.
- 30.Xu Y, Jones JE, Kohno D, Williams KW, Lee CE, Choi MJ, Anderson JG, Heisler LK, Zigman JM, Lowell BB, Elmquist JK. 5-HT2CRs expressed by pro-opiomelanocortin neurons regulate energy homeostasis. Neuron. 2008;60:582–9. doi: 10.1016/j.neuron.2008.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Qiu J, Xue C, Bosch MA, Murphy JG, Fan W, Rønnekleiv OK, Kelly MA. Serotonin 5-hydroxytryptamine2c receptor signaling in hypothalamic proopiomelanocortin neurons: role in energy homeostasis in females. Mol Pharmacol. 2007;72:885–96. doi: 10.1124/mol.107.038083. [DOI] [PubMed] [Google Scholar]
- 32.Heisler LK, Jobst EE, Sutton GM, Zhou L, Borok E, Thornton-Jones Z, Liu HY, Zigman JM, Balthasar N, Kishi T, Lee CE, Aschkenasi CJ, Zhang C-Y, Yu J, Boss O, Mountjoy KG, Clifton PG, Lowell BB, Friedman JM, Horvath T, Butler AA, Elmquist JK, Cowley MA. Serotonin reciprocally regulates melanocortin neurons to modulate food intake. Neuron. 2006;51:239–49. doi: 10.1016/j.neuron.2006.06.004. [DOI] [PubMed] [Google Scholar]
- 33.Zhou L, Sutton GM, Rochford JJ, Semple RK, Lam DD, Oksanen LJ, Thornton-Jones ZD, Clifton PG, Yueh C-Y, Evans ML, McCrimmon RJ, Elmquist JK, Butler AA, Heisler LK. Serotonin 2C receptor agonists improve type 2 diabetes via melanocortin-4 receptor signaling pathways. Cell Metab. 2007;6:398–405. doi: 10.1016/j.cmet.2007.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Company information. Available at http://www.arenapharm.com (last accessed.
- 35.Heal DJ, Smith SL, Fisas A, Codony X, Buschmann H. Selective 5-HT6 receptor ligands: progress in the development of a novel pharmacological approach to the treatment of obesity and related metabolic disorders. Pharmacol Ther. 2008;117:207–31. doi: 10.1016/j.pharmthera.2007.08.006. [DOI] [PubMed] [Google Scholar]
- 36.Woolley ML, Bentley JC, Sleight AJ, Marsden CA, Fone KC. A role for 5-HT6 receptors in retention of spatial learning in the Morris water maze. Neuropharmacology. 2001;41:210–9. doi: 10.1016/s0028-3908(01)00056-9. [DOI] [PubMed] [Google Scholar]
- 37.Fisas A, Codony X, Romero G, Dordal A, Giraldo J, Merce R, Holenz J, Heal D, Buschmann H, Pauwels PJ. Chronic 5-HT6 receptor modulation by E-6837 induces hypophagia and sustained weight loss in diet-induced obese rats. Br J Pharmacol. 2006;148:973–83. doi: 10.1038/sj.bjp.0706807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Akbas F, Gasteyger C, Sjödin A, Astrup A, Larsen TM. A critical review of the cannabinoid receptor as a drug target for obesity management. Obes Rev. 2008;10:58–67. doi: 10.1111/j.1467-789X.2008.00520.x. [DOI] [PubMed] [Google Scholar]
- 39.Di Marzo V. CB1 receptor antagonism: biological basis for metabolic effects. Drug Discov Today. 2008;13:1026–41. doi: 10.1016/j.drudis.2008.09.001. [DOI] [PubMed] [Google Scholar]
- 40.Trillou CR, Arnone M, Delgorge C, Gonalons N, Keane P, Maffrand J-P, Soubrie P. Anti-obesity effect of SR141716, a CB1 receptor antagonist, in diet-induced obese mice. Am J Physiol Regul Integr Comp Physiol. 2003;284:R345–53. doi: 10.1152/ajpregu.00545.2002. [DOI] [PubMed] [Google Scholar]
- 41.Pi-Sunyer FX, Aronne LJ, Heshmati HM, Devin J, Rosenstock J. Effect of rimonabant, a cannabinoid-1 receptor blocker, on weight and cardiometabolic risk factors in overweight or obese patients: RIO-North America: a randomized controlled trial. J Am Med Assoc. 2006;295:761–75. doi: 10.1001/jama.295.7.761. [DOI] [PubMed] [Google Scholar]
- 42.Plieth J. Obesity: what next after the CB1 antagonists' failure? Scrip. 2008. pp. 44–7. 7 November 2008.
- 43. Company information. Available at http://www.orexigen.com (last accessed.
- 44. Company information. Available at http://www.vivus.com (last accessed.
- 45.Mashiko S, Ishihara A, Iwaasa H, Sano H, Ito J, Gomori A, Oda Z, Moriya R, Matsushita H, Jitsuoka M, Okamoto O, MacNeil DJ, Van der Ploeg LHT, Fukami T, Kanatani A. A pair-feeding study reveals that a Y5 antagonist causes weight loss in diet-induced obese mice by modulating food intake and energy expenditure. Mol Pharmacol. 2007;71:602–8. doi: 10.1124/mol.106.029991. [DOI] [PubMed] [Google Scholar]
- 46.Levens NR, Della-Zuana O. Neuropeptide Y Y5 receptor antagonists as anti-obesity drugs. Curr Opin Investig Drugs. 2003;4:1198–204. [PubMed] [Google Scholar]
- 47.Erondu N, Gantz I, Musser B, Suryawanshi S, Mallick M, Addy C, Cote J, Bray G, Fujioka K, Bays H, Hollander P, Sanabria-Bohorquez SM, Eng W, Langstrom B, Hargreaves RJ, Burns HD, Kanatani A, Fukami T, MacNeil DJ, Gottesdiener KM, Amatruda JM, Kaufman KD, Heymsfield SB. Neuropeptide Y5 receptor antagonism does not induce clinically meaningful weight loss in overweight and obese adults. Cell Metab. 2006;4:275–82. doi: 10.1016/j.cmet.2006.08.002. [DOI] [PubMed] [Google Scholar]
- 48. Company information. Available at http://www.shionogiusa.com/media.html (last accessed.
- 49.Bertolini A, Tacchi R, Vergoni AV. Brain effects of melanocortins. Pharmacological Res. 2008;59:13–47. doi: 10.1016/j.phrs.2008.10.005. [DOI] [PubMed] [Google Scholar]
- 50.Nogueiras R, Wiedmer P, Perez-Tilve D, Veyrat-Durebex C, Keogh JM, Sutton GM, Pfluger PT, Castaneda TR, Neschen S, Hofmann SM, Howles PN, Morgan DA, Benoit SC, Szanto I, Schrott B, Schürmann A, Joost H-G, Hammond C, Hui DY, Woods SC, Rahmouni K, Butler AA, Farooqi IS, O'Rahilly S, Rohner-Jeanrenaud F, Tschöp MH. The central melanocortin system directly controls peripheral lipid metabolism. J Clin Invest. 2007;117:3475–88. doi: 10.1172/JCI31743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Luthin DR. Anti-obesity effects of small molecule melanin-concentrating hormone receptor1 (MCHR1) antagonists. Life Sci. 2007;81:423–40. doi: 10.1016/j.lfs.2007.05.029. [DOI] [PubMed] [Google Scholar]
- 52.Mendez-Andino J, Wos JA. MCH-R1 antagonists: what is keeping most research programs away from the clinic? Drug Discov Today. 2007;12:972–9. doi: 10.1016/j.drudis.2007.08.010. [DOI] [PubMed] [Google Scholar]
- 53.Kowalski TJ, Farley C, Cohen-Williams ME, Varty G, Spar BD. Melanin concentrating hormone-1 receptor antagonism decreases feeding by reducing meal size. Eur J Pharmacol. 2004;497:41–7. doi: 10.1016/j.ejphar.2004.06.027. [DOI] [PubMed] [Google Scholar]
- 54.Shearman LP, Camacho RE, Sloan Stribling D, Zhou D, Bednarek MA, Hreniuk DL, Feighner SD, Tan CP, Howard AD, Van der Ploeg LH, MacIntyre DE, Hickey GJ, Alison M, Strack AM. Chronic MCH-1 receptor modulation alters appetite, body weight and adiposity in rats. Eur J Pharmacol. 2003;475:37–47. doi: 10.1016/s0014-2999(03)02146-0. [DOI] [PubMed] [Google Scholar]
- 55.Sargent B, Surman M, Freeman E, Grabowski J, Jiang X-W, Jiang G, Hadden M, Luche M, Khmelnitsky Y, Lindsay J, Mozhaeva L, Zaremba J, Viggers J, Vickers S, Goddard S, Cheetham S, Guzzo P. Comparative pharmacology of AMR-MCH-1 and AMR-MCH-2, MCH1 receptor antagonists for the treatment of obesity. Proceedings of the British Pharmacological Society, Available at http://www.pA2online.org/abstracts/Vol6Issue4abst114P.pdf (last accessed.
- 56. Company information. Available at http://www.neurogen.com/index.php?option=com_content&view=article&id=63&Itemid=78&p=irol-news (last accessed.