

Abstract
Background
Human immunodeficiency virus (HIV) vaccine development remains a global priority. We describe the safety and immunogenicity of a multi-clade DNA vaccine prime with a replication-defective Adenovirus type 5 (rAd5) boost.
Methods
The vaccine is a 6-plasmid mixture encoding HIV envelope (env) subtypes A, B and C and subtype B gag, pol and nef, and a rAd5 expressing identical genes, with the exception of nef. Three hundred and twenty-four participants were randomized to receive placebo (n=138), a single dose of rAd5 at 1010 (n=24) or 1011 particle units (n=24), or DNA at 0, 1 and 2 months followed by rAd5 at either 1010 (n=114) or 1011 particle units (n=24) boosting at 6 months. Participants were followed for 24 weeks after the final immunization.
Results
The vaccine was safe and well tolerated. HIV-specific T cell responses were detected in 63% of vaccinees. Pre-existing Ad5 neutralizing antibody titer did not impact the frequency and magnitude of T cell responses in prime-boost recipients, but did impact the response rates in participants receiving rAd5 alone (p=0.037).
Conclusion
The DNA/rAd5 immunization regimen was safe and induced HIV-1 multi-clade T cell responses, which were not significantly affected by pre-existing rAd5 neutralizing antibody titer.
Keywords: HIV-1 Vaccine, DNA plasmid vaccine, recombinant Adenovirus vaccine, Africa vaccine trial
INTRODUCTION
The HIV epidemic continues to grow with 33 million persons living with HIV/AIDS and an additional 7,400 new infections daily in 2007 [1]. While multiple HIV prevention strategies have been evaluated with some recent successes [2, 3], a vaccine to prevent HIV infection, or alter disease course, remains the cornerstone of a successful global HIV control program. Despite the failure of the VaxGen and Merck candidate HIV vaccines in efficacy testing [4–6], there is consensus that both basic discovery and clinical research pursuing a vaccine is justified [7]. A vaccine that induces primarily T cell immunity may only afford a reduction in viral burden among recipients based upon animal model studies, yet be a valuable contribution to HIV control programs by slowing disease progression and reducing the rate of secondary infections [8, 9].
The Vaccine Research Center (VRC), National Institutes of Allergy and Infectious Diseases candidate HIV vaccine is a multi-clade HIV DNA prime vaccine expressing HIV-1 Gag, Pol, Nef and Env proteins, boosted with recombinant Adenovirus type 5 (rAd5) expressing matching proteins with the exception of Nef. The VRC DNA/rAd5 vaccine was designed for induction of HIV specific T cell immunity, to ideally prevent HIV acquisition and, failing that, to reduce viral load in vaccine recipients who did become infected [10]. Unlike the Merck vaccine, which failed to provide benefit in efficacy testing, the VRC prime-boost candidate provided a survival benefit in non-human primate studies using a homologous, intravenous, pathogenic SIV challenge [11–14]. Both the VRC HIV DNA prime and the rAd5 vaccines were safe, well-tolerated and immunogenic when administered separately to HIV uninfected adults in the US [15, 16].
The VRC HIV DNA/rAd5 vaccine was evaluated for safety and immunogenicity in three studies among lower risk populations of HIV uninfected adults in the US, Caribbean, South Africa and East Africa to establish safety and immunogenicity at sites similar to those which might provide higher risk volunteers for efficacy testing of the vaccine. The combined data from these studies were considered sufficient to determine whether this candidate vaccine should proceed to proof of concept efficacy testing [10]. Here we describe one of these studies assessing safety and immunogenicity of a replication-defective rAd5 administered alone or following priming with a multi-clade DNA in East African populations with high pre-existing Adenovirus 5 (Ad5) neutralizing antibody titers.
METHODS
Study Design
RV 172 was a randomized, double-blinded placebo-controlled trial performed at three clinical research sites: the Makerere University-Walter Reed Project (MUWRP), Kampala, Uganda, the Walter Reed Project (WRP), Kericho, Kenya and the Mbeya Medical Research Programme (MMRP), Mbeya, Tanzania. The protocol was approved by ethical review boards in each country and the US Department of Defense. All volunteers provided written informed consent. The trial was conducted in two parts as shown in Table 1. Part A, a phase I study in 144 volunteers (Groups 1–4) evaluated rAd5 at two doses, 1010 and 1011 particle units (PU)/mL, administered as a single dose alone (Groups 1 and 2, respectively) or as a boost following three injections of the DNA prime vaccine (Groups 3 and 4, respectively). Two dose regimens of rAd5 were evaluated to provide information about the safety and immunogenicity of this vaccine in African populations where pre-existing neutralizing antibody to the vector is common [17]. Part B, a phase II study in 180 volunteers, further defined safety, tolerability and immunogenicity for the preferred prime-boost combination for efficacy testing (Group 5). In Part B, participants were randomized to either vaccine or placebo at a ratio of 1:1. Solicited adverse events (AE) commonly associated with vaccination and unsolicited AE, including a battery of blood and urine tests, were assessed at clinical encounters with volunteers 2 weeks after each vaccination and 4, 6, 12 and 24 weeks following the final immunization.
Table 1.
Study Design
| Immunization Schedule | ||||
|---|---|---|---|---|
| Group (No. of vaccinees/placebos) | Day 0 | Day 28 | Day 56 | Day 168 |
| 1 (24/12) | VRC-HIVADV014-00-VP 1010 PU | |||
| 2 (24/12) | VRC-HIVADV014-00-VP 1011PU | |||
| 3 (24/12) | VRC-HIVDNA-016-00-VP (4mg) | VRC-HIVDNA-016-00-VP (4mg) | VRC-HIVDNA-016-00-VP (4mg) | VRC-HIVADV014-00-VP 1010 PU |
| 4 (24/12) | VRC-HIVDNA-016-00-VP (4mg) | VRC-HIVDNA-016-00-VP (4mg) | VRC-HIVDNA-016-00-VP (4mg) | VRC-HIVADV014-00-VP 1011PU |
| 5 (90/90) | VRC-HIVDNA-016-00-VP (4mg) | VRC-HIVDNA-016-00-VP (4mg) | VRC-HIVDNA-016-00-VP (4mg) | VRC-ADV014-00- VP 1010PU |
Vaccine product
The VRC vaccines have been previously described [15, 16]. Briefly, the VRC DNA-HIV vaccine (VRC-HIVDNA016-00-VP) and VRC rAd5-HIV vaccine (VRC-HIVADV014-00-VP) contain largely matched HIV gene inserts, but they are not identical. VRC-HIVDNA016-00-VP (Vaccine Research Center, Bethesda, MD, USA) is composed of six closed, circular, DNA plasmids: three plasmids expressing HIV-1 Gag, Pol and Nef proteins from clade B and representing 50% by weight; and three plasmids each expressing HIV-1 Env glycoprotein from clades A, B and C, and representing the other 50% by weight (16.67% each). VRC-HIVADV014-00-VP (Vaccine Research Center, Bethesda, MD, USA), is composed of four replication-defective, recombinant serotype 5 adenoviral vector composed of four rAd5 vectors (in a 3:1:1:1 ratio) encoding HIV-1 Gag/Pol polyproteins from clade B and HIV-1 Env glycoproteins from clades A, B, and C. The envelope genes were chosen as representative primary isolates from each of the three clades. Mutations and deletions were introduced into HIV genes to preclude expression of functional proteins. Envelope sequences were truncated and the DNA plasmids and rAd5 vectors express gp145 (cytoplasmic tail deleted) and gp140 (transmembrane domain and cytoplasmic tail deleted) envelope proteins, respectively. In addition, the EnvB construct in the rAd5 had a V1/V2 deletion not present in the DNA product. The DNA vaccine was delivered intramuscularly in a dose of 4 mg using a needle free injection system (Biojector, Bioject Incorporated, Bedminster, NJ, USA). The rAd5 vaccine (1ml) was delivered intramuscularly by needle injection in a dose of either 1010 or 1011 PU/mL.
HIV Diagnosis
Monitoring for HIV-1 infection was performed using a sequential algorithm of the following U.S. FDA approved tests: Genetic Systems rLAV ELISA (BioRad Laboratories, Redmond, WA), Vironostika HIV-1 Microelisa (BioMerieux, Durham, NC), Genetic Systems HIV-1 Western Blot Kit (BioRad Laboratories) and Amplicor HIV-1 Monitor version 1.5 RT-PCR (Roche Diagnostics, Indianapolis, IN).
Adenovirus type 5 neutralizing antibody (Ad5 Nab) assay
Sera collected from participants prior to vaccination were used for determining Ad5 Nab titers. A validated assay was performed at the NIAID Vaccine Immune T-cell and Antibody Laboratory (NVITAL, Gaithersburg, MD, USA), a contract laboratory supporting NIAID funded clinical vaccine trials. Serum dilutions, ranging from 1:12 to 1:8748, were incubated with a luciferase reporter rAd5 for 30 minutes and added to A549 (human lung carcinoma) cells. Following incubation for 24 hours, luciferase activity was assessed. The 90% inhibition serum titer was determined to be the serum dilution, which can be interpolated to have 10% of the maximum luciferase activity relative to the assay run in the absence of serum.
Cell Preparation
Peripheral blood mononuclear cells (PBMC) were prepared from acid-citrate dextrose anti-coagulated tubes by Ficoll-Hypaque PLUS density gradient centrifugation (Pharmacia, Uppsala, Sweden) using Leucosep tubes (Greiner Bio-one, Essen, Germany) within 8h of collection. PBMCs were cryopreserved in RPMI medium (Sigma, St. Louis, MO) containing 20% heat-inactivated fetal-calf serum and 10% dimethylsulfoxide (DMSO;Sigma) in a Cryo 1°C freezing container (Nalgene, Rochester, NY). Cells were stored at ≤ −140°C. Immunogenicity assessments were performed on thawed specimens; average cell viability was >94% immediately after thawing.
IFN-γ ELISpot assay
PBMC were thawed and rested overnight prior to testing. The IFN-γ ELISpot assay was performed at a central laboratory as previously described [18] HIV peptides of 15-mer overlapping by 11 amino acids were prepared and divided into 5 peptide pools, representing Env (clades A and B), Pol 1 and Pol 2 (clade B) and Nef (clade B) (Vaccine Research Center, NIH, Bethesda, MD). These peptide pools matched the vaccine HIV gene products. An Env C peptide pool was kindly provided by the International AIDS Vaccine Initiative (IAVI) and HIV clade B-Gag (LAI) was from New England Peptide LLC (Gardner, MA). All peptides had a purity of >85% as measured by high performance liquid chromatography and mass spectrometry. All peptides were used at a final concentration of 1μg/ml. Since all peptides were reconstituted in DMSO, wells containing PBMC and media only were supplemented with the equivalent concentration of DMSO and served as negative controls. Phytohemagglutinin (PHA;Sigma) was used as a positive control. PBMC plus peptide antigens or PHA were tested in 3 replicate wells. Negative controls were performed in quadruplicate. Spots were counted with a C.T.L. analyzer (Shaker Heights, OH) and software (version 4.0.19, C.T.L. Analyzers). Results are expressed as spot-forming cells (SFC)/106 PBMC. A positive IFN-γ response was defined as at least 55 SFC/106 PBMC (uncorrected) and at least 4 times the DMSO treated wells [19]. The assay was scored as invalid if PHA treated PBMC failed to produce a positive response and/or the negative control was ≥ 55 SFC/106 PBMC.
Data Analysis
Comparisons of reactogenicity and adverse events were made by assigning scores of 0, 1, 2 and 3 to severities of none, mild, moderate and severe, respectively. The Wilcoxon Rank sum test was used for 2 group comparisons. Otherwise, the Kruskal-Wallis test was used. Fisher’s exact test was used for 2 by 2 tables. All comparisons were based on the maximum severity per volunteer. The proportion of volunteers with positive ELISpot responses was tested using Fisher’s exact test. The Cochran-Armitage test for trend was used to compare responses rates across increasing pre-immunization Ad5 Nab titer. The magnitude of ELISpot responses was tested by ANOVA on log10 response. All tests are 2-tailed and all analyses were performed using SAS.
RESULTS
Enrollment and vaccination of 324 volunteers commenced in May 2006 and was completed in October 2006 with 144, 120 and 60 participants at the Ugandan, Kenyan and Tanzanian sites, respectively. The average age was 28 years (range 18–49) and 103 (32%) participants were female. Three hundred and eleven (96%) participants completed the 12-month study and 13 (4%) terminated the study before the final visit. Nineteen (6%) participants did not complete the vaccination schedule (8 placebo and 11 vaccine recipients). The most common reason for discontinuation of vaccination was pregnancy (N = 8). Two individuals prematurely stopped vaccination due to adverse events (one vaccine recipient for urticaria, and one placebo recipient for generalized itching). No deaths occurred during the 12 months of the study. A single HIV infection was observed during the study period in a vaccine recipient. All traceable volunteers were offered counseling and HIV testing following public disclosure of the Merck STEP study results and 264 (81%) attended briefing and VCT sessions. A total of 4 additional HIV infections have been identified in extended follow-up (one vaccine recipient and 3 placebo recipients) yielding an overall infection rate of 0.611 per 100 person-years observation. The infection rate observed by MHRP in community cohort studies at or near these research sites have ranged from 0.77 to 1.8 per 100 person-years observation (M. Hoelscher, personal communication;[20]).
Vaccinations were well tolerated (supplemental figure 1). Solicited local and systemic adverse events were commonly reported by both vaccine (88% for both) and placebo recipients (80% for both). The frequency and severity of local and systemic solicited adverse reactions was not different between the high and low dose of rAd5 either alone (p=0.440 and 0.367 for local and systemic adverse events, respectively) or in the prime-boost regimen (p=0.458 and 0.803 for local and systemic adverse events, respectively). The presence and titer of pre-existing antibody to Ad5 did not alter the frequency or severity of solicited local or systemic adverse events among vaccine recipients (data not shown, p=0.767 and p=0.938 respectively). There were no differences between vaccine and placebo recipients in the frequency or severity of routinely collected clinical laboratory measures (data not shown).
A total of 1446 AE were reported among 309 of 324 participants. Infectious diseases were the predominate adverse events with upper respiratory tract infections most common followed by malaria, urinary tract infections and gastroenteritis. Blood disorders were the second most common diagnoses led by neutropenia and thrombocytopenia. Adverse event rates were similar among placebo and vaccine participants. 62 adverse events were graded as severe or life-threatening and malaria and neutropenia were the most common diagnoses. The proportion of severe or life-threatening adverse events among vaccine and placebo participants were similar (4.9% versus 3.4%, p =0.159). Four of these, all occurring in vaccine recipients, were considered possibly or probably related to vaccine. These were two cases of generalized urticaria, an episode of neutropenia and an episode of malaise. There was no relationship between the frequency or severity of adverse events and the titer of pre-existing adenovirus antibody for either placebo or vaccine recipients (data not shown).
Immunogenicity Assessment Results
Responses to HIV antigens were measured using IFN-γ ELISpot prior to and 6 weeks after either a single rAd5 injection or the DNA/rAd5 regimen. Results from groups 3 and 5 were combined (3/5) as they evaluated the same regimen. Fifteen study participants who did not complete vaccination as assigned per protocol were excluded from analysis and in some cases, insufficient cells for analysis resulted in further missing data points.
IFN-γ ELISpot response frequencies post-vaccination for each peptide pool and for any positive response are shown for placebo and vaccine recipients by study group in Table 2. False positive rates from placebo (7/121;6%) and baseline samples were low for all peptide pools evaluated together (9/275; 3%, data not shown). The response rates to any peptide pool for the low and high dose rAd5 alone were 74% and 52% respectively (p=0.221). Similarly, response rates for DNA prime and low or high dose rAd5 boost were 63% and 60% respectively (p=0.806). Positive IFN-γ ELISpot responses to any Env, Gag or Pol peptide pool at 6 weeks following rAd5 alone (groups 1 and 2) and DNA/rAd5 (groups 3, 4 and 5) were 29 of 46 (63%) and 78 of 125 (62%), respectively and were not significantly different. Response rates for each Env peptide pool were not different between the Ad5 alone and prime-boost groups. The Env pools matching the vaccine amino acid sequence elicited better response rates compared to the subtype C Env peptide pool regardless of study group. The frequency of Gag responses in the rAd5 alone group was significantly less than that in the DNA/rAd5 groups, (3/45 versus 41/124; p < 0.001). Pol responses were more common in the rAd5 alone groups compared to the DNA/rAd5 groups, (15/45 versus 19/120; p=0.018).
Table 2.
Frequency (percentage) of subjects with detectable T cell responses to HIV antigens by treatment group, as measured by interferon-gamma enzyme-linked immunospot (ELISpot) assay in CD4+ or CD8+ T cells, 6 weeks after immunization.
| HIV-1 Peptide Pool | |||||||
|---|---|---|---|---|---|---|---|
| Env A, B or C | Env A | Env B | Env C | Gag | Pol | Any | |
| All Placebos | 5/121 (4) | 1/120 (1) | 2/117 (2) | 3/110 (3) | 1/119 (1) | 1/117 (1) | 7/121 (6) |
| rAd 5 1010 PU | 15/23 (65) | 10/23 (43) | 14/22 (64) | 3/21 (14) | 2/22 (9) | 10/22 (45) | 17/23 (74) |
| rAd 5 1011 PU | 11/23 (48) | 7/23 (30) | 10/23 (43) | 2/21 (10) | 1/23 (4) | 5/23 (22) | 12/23 (52) |
| rAd 5 single dose total | 26/46 (57) | 17/46 (37) | 24/45 (53) | 5/42 (12) | 3/45 (7) | 15/45 (33) | 29/46 (63) |
| D/D/D/rAd5 1010 PU | 58/105 (55) | 46/105 (44) | 48/104 (46) | 19/101 (19) | 34/105 (32) | 14/102 (14) | 66/105 (63) |
| D/D/D/rAd5 1011 PU | 12/20 (60) | 11/20 (55) | 8/18 (44) | 4/16 (25) | 7/19 (37) | 5/18 (28) | 12/20 (60) |
| rAd 5 prime boost total | 70/125 (56) | 57/125 (46) | 56/122 (46) | 23/117 (20) | 41/124 (33) | 19/120 (16) | 78/125 (62) |
Note: rAd5, recombinant Adenovirus 5 vectors (VRC-HIVADV014-00-V); D, DNA plasmids (VRC-HIVDNA016-00-VP); PU, particle units. Data are no. of positive responses/no. of subjects tested (%).
There was no difference in the magnitude of response measured by IFN-γ ELISpot among the four regimens examined in the study (Figure 1, p=0.254). The median (range) responses were 127 (60–493), 141 (70–463), 163 (55–2103) and 236 (58–1007) SFC/106 PBMC in Groups 1, 2, 3/5 and 4, respectively. There was no difference between low and high dose rAd5 alone, (arm 1 versus arm 2) or in a prime/boost format between arms 3/5 and 4 in terms of response frequency or magnitude. Analyzing these data with stratification for pre-existing Adenovirus antibody titers shows no significant differences but these results must be interpreted with caution given the small numbers of such a comparison (data not shown). The proportions of volunteers with responses to combinations of Env, Gag and/or Pol peptides are shown in Figure 2. The frequency of positive responses was predominantly to Env, followed by Pol or Gag, regardless of immunization regimen. Among the four regimen groups, roughly equal proportions recognized two antigens (20%–26%). Group 4, with the high dose rAd5 boost had the highest frequency of responders to all three antigens tested (20%, p = 0.137).
Figure 1.
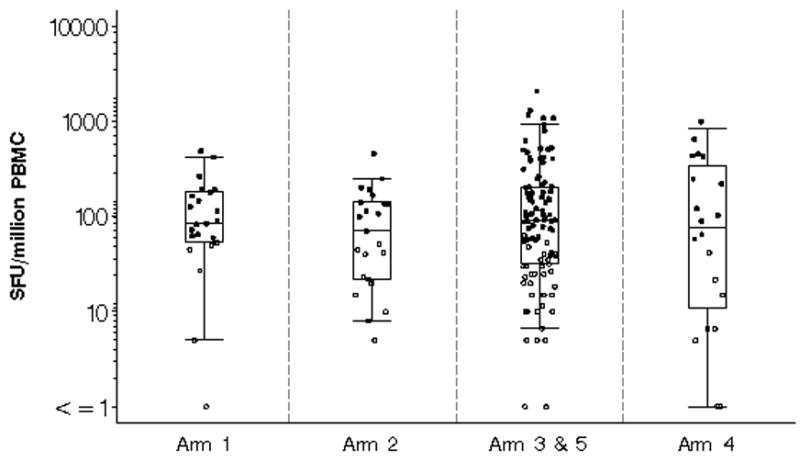
The maximum magnitude of T cell responses measured by interferon-γ ELISpot expressed as spot forming units (SFU) per one million peripheral blood mononuclear cells (PBMC) in vaccinees to either Env, Pol or Gag by immunization regimen 6 weeks after immunization. Open circles represent positive responses and black circles represent negative responses. A positive response was defined as at least 55 spot forming cells/106PBMC in peptide pools and 4 X background number of spot forming cells/106PBMC (media only). The box plots indicate the median, 25th and 75th percentile for each group and the error bars show the 5th and 95th percentiles. Placebo and baseline responses are not shown.
Figure 2.
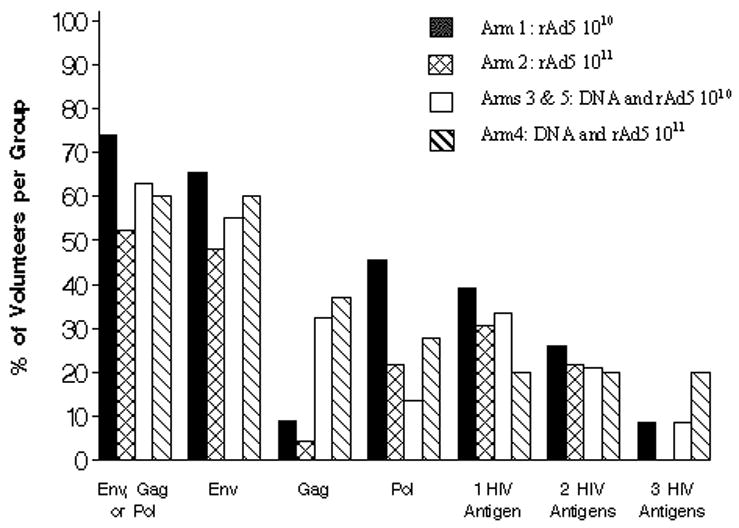
Breadth of T cell responses to HIV antigens. Frequencies of vaccinated subjects with detectable T cell responses to HIV antigens 6 weeks after immunization. Responses were measured by the interferon (IFN)-γ ELISpot assay. A positive response was defined as at least 55 spot forming cells/106PBMC in peptide pools and 4 X background number of spot forming cells/106PBMC (media only). The columns represent the percent volunteers with a positive IFN-γ ELISpot response for each study group represented by different colored bars for Env Gag or Pol peptides alone, Env alone, either Gag or Pol peptide pools, and for response rates to single, any two or three HIV antigens.
Ad5 Nab was present at baseline in 175 of 186 vaccine recipients (94%) with a median titer of 2385. After substituting a value of 8750 for titers reported as >8748, the corresponding median and geometric mean were 3521 and 1407, respectively. Positive IFN-γ ELISpot response rates to HIV Env, Pol or Gag peptide pools among vaccine recipients only, stratified by Ad5 Nab titer, are shown in Table 3. The frequency of response to any HIV antigen after rAd5 immunization alone was 3 of 4 (75%) for participants with negative Ad5 Nab titers, 14 of 18 (78%) for participants with Ad5 Nab titers of <1000, 7 of 12 (58%) for those with titers 1000 to 5000, and 5 of 12 (42%) for those with titers > 5000 (p=0.048). Pre-existing immunity to Ad5 had no effect (p=0.252) on HIV-specific response rates in prime and boost recipients where 6 of 7 (86%) with negative Ad5 Nab titers had a positive IFN-γ ELISpot response, compared with 24 of 39 (62%) with Ad5 Nab < 1000, 27 of 41 (66%) of those with titers 1000 to 5000, and 21 of 38 (55%) of those with titers over 5000. The magnitude of IFN-γ ELISpot responses for groups 1 and 2 combined and groups 3, 4 and 5 combined and stratified by baseline Ad5 Nab titer is shown in Figure 3. A modest reduction in magnitude with increasing Ad5 Nab titer is apparent, though not statistically significant for both single rAd5 vaccination (p=0.184) and the prime boost regimens (p=0.595).
Table 3.
Effect of pre-existing Adenovirus 5 neutralizing antibody titer on HIV-1 specific interferon-gamma ELISpot reactivity
| Immunization regimen Responders, no. (%) | |||
|---|---|---|---|
| HIV-1 Peptide pool | Adenovirus 5 Nab titer | rAd5 only | DNA prime/rAd5 boost |
| ENV A | <12 | 2 (50) | 5 (71) |
| <1000 | 9 (50) | 17 (44) | |
| 1000–5000 | 3 (25) | 21 (51) | |
| >5000 | 3 (25) | 14 (37) | |
| ENV B | <12 | 3 (75) | 4 (57) |
| <1000 | 13 (76) | 17 (44) | |
| 1000–5000 | 6 (50) | 19 (49) | |
| >5000 | 2 (17) | 16 (43) | |
| ENV C | <12 | 1 (25) | 2 (33) |
| <1000 | 2 (13) | 4 (11) | |
| 1000–5000 | 0 (0) | 10 (27) | |
| >5000 | 2 (17) | 7 (18) | |
| GAG | <12 | 0 (0) | 4 (57) |
| <1000 | 2 (12) | 16 (41) | |
| 1000–5000 | 1 (8) | 13 (33) | |
| >5000 | 0 (0) | 8 (21) | |
| POL | <12 | 3 (75) | 4 (57) |
| <1000 | 7 (41) | 5 (13) | |
| 1000–5000 | 2 (17) | 6 (16) | |
| >5000 | 3 (25) | 4 (11) | |
| Any | <12 | 3 (75) | 6 (86) |
| <1000 | 14 (78) | 24 (62) | |
| 1000–5000 | 7 (58) | 27 (66) | |
| >5000 | 5 (42) | 21 (55) | |
Figure 3.
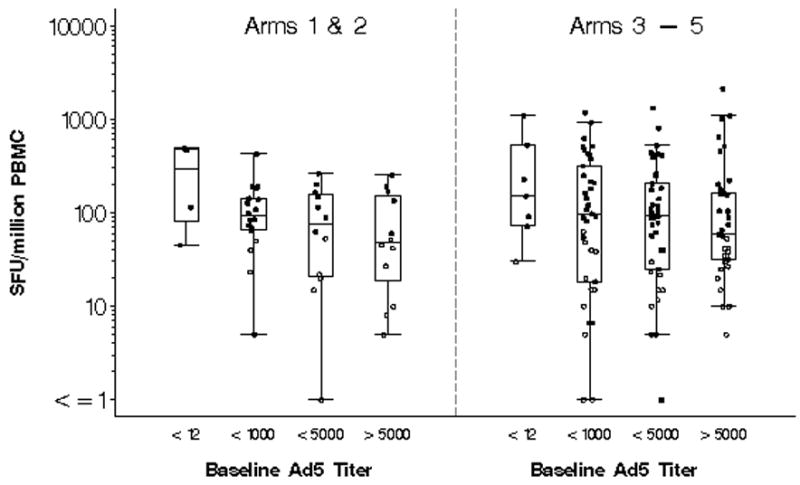
The maximum magnitude of IFN-γ ELISpot responses stratified by baseline Adenovirus type 5 neutralizing antibody titer of <12, 12–1000, 1000–5000 and > 5000. Results are expressed as spot forming units (SFU) per one million peripheral blood mononuclear cells (PBMC), for groups 1 and 2 combined and groups 3–5. Open circles represent positive responses and black circles represent negative responses. A positive response was defined as at least 55 spot forming cells/106PBMC in peptide pools and 4 X background number of spot forming cells/106PBMC (media only). The box plots indicate the median, 25th and 75th percentile for each group and the error bars show the 5th and 95th percentiles. Placebo and baseline responses are not shown.
DISCUSSION
The HIV vaccine field has achieved only incremental progress, which has been in the form of negative studies defining conceptual approaches that do not lower acquisition risk or reduce viral load. The benefit of a vaccine that would modestly reduce viral load, even if it confers no protection from acquisition could be significant. In addition to extending the interval before initiating anti-viral therapy, decreasing host viral burden is likely to reduce secondary HIV transmission [21, 22]. The current study examines the clinical and immunological characteristics of a novel candidate DNA plasmid prime/rAd5 vector boost vaccine combination that is distinct from the vaccines that have failed in test of concept efficacy trials [4–6].
The VaxGen monomeric gp120 vaccine was able to induce neutralizing antibody to T cell line adapted variants of HIV but unable to generate antibody responses to the most commonly transmitted variants of HIV [5, 6]. The Merck vaccine was designed to elicit HIV specific T cell immunity and produced HIV-specific responses which were predominantly CD8+ T cells and only 31% of recipients showing both CD8+ and CD4+ T cell responses [23]. In non-human primates, the Merck vaccine produced marginal reduction of viral load following SIV challenge [14]. In contrast, the VRC candidate reported here, is a heterologous vector strategy employing a DNA prime and rAd5 boost and has shown both a reduced level of post-challenge viremia and a survival benefit in non-human primate SIV challenge studies. Interestingly, this survival benefit was correlated to induction of total HIV specific T cell responses, CD8+ T cell responses, CD4+ T HIV-specific IFN-γ ELISpot responses and preservation of central memory CD4+ T cells [11, 13]. Similar to observations in the non-human primate challenge study, intracellular cytokine staining of a subset of volunteers in this study identifies a balanced CD4+ and CD8+ T cell response to rAd5 following DNA priming [12, 24].
The data presented here, show that the VRC rAd5, and DNA/rAd5 HIV vaccines are safe and well tolerated in a population within resource-constrained settings experiencing a significantly greater burden of endemic illness. Vaccination with rAd5 alone or in a prime-boost combination with DNA was highly immunogenic despite very high prevalence and titer of antibody to Ad5 at study entry. A similar study in the United States using the identical rAd5 immunogen, dose and regimen described here for groups 1 and 2 demonstrated nearly identical IFN-γ ELISpot response frequencies to HIV antigens as this study, supporting the use of this immunogen in a global setting [15]. A diminution in the magnitude of the ELISpot responses with increasing Ad5 Nab has been previously reported for rAd5 vaccines [15, 23]. While the overall frequency of responders to all HIV antigens was similar between the rAd5 only and prime-boost recipients, there was a reduction in response frequency in the rAd5 only groups with rising baseline Ad5 titer. The prime-boost strategy induced a higher frequency of responders than the rAd5 group in subjects with rAd5 Nab titers ≥1000. Surprisingly, there was no evidence of benefit to the higher 1011 dose of Ad5 vector compared to the lower 1010 dose in either the rAd5 only or the prime-boost combination. In terms of overall frequency and magnitude of cellular immune responses as measured by IFN-γ ELISpot, the rAd5 alone performed as well as the prime boost regimen, with the exception of Gag responses, where the frequency of responders in the prime-boost regimen was almost 5 times that of the single rAd5 immunization.
It has been argued that a more rigorous animal model would utilize a challenge that is genetically divergent from the strain used to design the test vaccine. However, it is not known whether the non-human primate model will be predictive of protection in humans and if so, under what experimental conditions. It remains necessary to examine the efficacy of products, which are safe and distinct from previous vaccine candidates as a complimentary avenue of discovery to validate pre-clinical testing paradigms. In view of the Step study results, the VRC DNA prime and rAd5 boost vaccine is expected to enter efficacy testing in the US only among circumcised males without evidence of pre-existing immunity to Ad5. Although the population under evaluation in this study is not part of the initial test of concept efficacy trial in view of the low rates of circumcision and high prevalence of Ad5 antibody, it will be an important component of other studies should evidence of efficacy be demonstrated.
Supplementary Material
Acknowledgments
We wish to thank the study volunteers, the study staff and Alberto Ortiz, Dr. Chuen-Yen Lau, Dr. Alan Fix and Dr. Robert Gramzinski.
This work was supported by a cooperative agreement (W81XWH-07-2-0067) between the Henry M. Jackson Foundation for the Advancement of Military Medicine, Inc., and the U.S. Department of Defense (DOD). The study was funded and sponsored by the Division of AIDS, NIAID, NIH, DHHS. The views expressed are those of the authors and should not be construed to represent the positions of the U.S. Department of Defense.
Footnotes
The authors do not have a commercial or other association that might pose a conflict of interest.
This data was presented in part as abstract P13-23 at “AIDS Vaccine 2008”, 13–16 October, 2008 in Capetown, South Africa.
http://clinicaltrials.gov - registry number NCT00123968
References
- 1.UNAIDS/WHO. Report on the Global AIDS Epidemic. Geneva: UNAIDS; 2008. [Google Scholar]
- 2.Gray RH, Wawer MJ, Serwadda D, Kigozi G. The role of male circumcision in the prevention of human papillomavirus and HIV infection. J Infect Dis. 2009;199:1–3. doi: 10.1086/595568. [DOI] [PubMed] [Google Scholar]
- 3.Karim SA, Coletti A, Richardson B, Ramjee G, Hoffman I, Chirenje M, Taha T, Maslankowski L, Soto-Torres L. Safety and Effectiveness of Vaginal Microbicides BufferGel and 0.5% PRO 2005/5 Gel for the Prevention of HIV Infection in Women: Results of the HPTN 035 Trial. Conference on Retroviruses and Opportunistic Infections; Montreal, Canada. 2009. [Google Scholar]
- 4.Buchbinder SP, Mehrotra DV, Duerr A, et al. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): a double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet. 2008;372:1881–93. doi: 10.1016/S0140-6736(08)61591-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Flynn NM, Forthal DN, Harro CD, Judson FN, Mayer KH, Para MF. Placebo-controlled phase 3 trial of a recombinant glycoprotein 120 vaccine to prevent HIV-1 infection. J Infect Dis. 2005;191:654–65. doi: 10.1086/428404. [DOI] [PubMed] [Google Scholar]
- 6.Pitisuttithum P, Gilbert P, Gurwith M, et al. Randomized, double-blind, placebo-controlled efficacy trial of a bivalent recombinant glycoprotein 120 HIV-1 vaccine among injection drug users in Bangkok, Thailand. J Infect Dis. 2006;194:1661–71. doi: 10.1086/508748. [DOI] [PubMed] [Google Scholar]
- 7.Johnston MI, Fauci AS. An HIV vaccine--challenges and prospects. N Engl J Med. 2008;359:888–90. doi: 10.1056/NEJMp0806162. [DOI] [PubMed] [Google Scholar]
- 8.Walker BD, Burton DR. Toward an AIDS vaccine. Science. 2008;320:760–4. doi: 10.1126/science.1152622. [DOI] [PubMed] [Google Scholar]
- 9.Watkins DI, Burton DR, Kallas EG, Moore JP, Koff WC. Nonhuman primate models and the failure of the Merck HIV-1 vaccine in humans. Nat Med. 2008;14:617–21. doi: 10.1038/nm.f.1759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nabel GJ. Mapping the future of HIV vaccines. Nat Rev Microbiol. 2007;5:482–4. doi: 10.1038/nrmicro1713. [DOI] [PubMed] [Google Scholar]
- 11.Letvin NL, Mascola JR, Sun Y, et al. Preserved CD4+ central memory T cells and survival in vaccinated SIV-challenged monkeys. Science. 2006;312:1530–3. doi: 10.1126/science.1124226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mattapallil JJ, Douek DC, Buckler-White A, et al. Vaccination preserves CD4 memory T cells during acute simian immunodeficiency virus challenge. J Exp Med. 2006;203:1533–41. doi: 10.1084/jem.20060657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sun Y, Schmitz JE, Buzby AP, et al. Virus-specific cellular immune correlates of survival in vaccinated monkeys after simian immunodeficiency virus challenge. J Virol. 2006;80:10950–6. doi: 10.1128/JVI.01458-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Casimiro DR, Wang F, Schleif WA, et al. Attenuation of simian immunodeficiency virus SIVmac239 infection by prophylactic immunization with dna and recombinant adenoviral vaccine vectors expressing Gag. J Virol. 2005;79:15547–55. doi: 10.1128/JVI.79.24.15547-15555.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Catanzaro AT, Koup RA, Roederer M, et al. Phase 1 safety and immunogenicity evaluation of a multiclade HIV-1 candidate vaccine delivered by a replication-defective recombinant adenovirus vector. J Infect Dis. 2006;194:1638–49. doi: 10.1086/509258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Catanzaro AT, Roederer M, Koup RA, et al. Phase I clinical evaluation of a six-plasmid multiclade HIV-1 DNA candidate vaccine. Vaccine. 2007;25:4085–92. doi: 10.1016/j.vaccine.2007.02.050. [DOI] [PubMed] [Google Scholar]
- 17.Kostense S, Koudstaal W, Sprangers M, et al. Adenovirus types 5 and 35 seroprevalence in AIDS risk groups supports type 35 as a vaccine vector. Aids. 2004;18:1213–6. doi: 10.1097/00002030-200405210-00019. [DOI] [PubMed] [Google Scholar]
- 18.Currier JR, Kuta EG, Turk E, et al. A panel of MHC class I restricted viral peptides for use as a quality control for vaccine trial ELISPOT assays. J Immunol Methods. 2002;260:157–72. doi: 10.1016/s0022-1759(01)00535-x. [DOI] [PubMed] [Google Scholar]
- 19.Fu TM, Dubey SA, Mehrotra DV, et al. Evaluation of cellular immune responses in subjects chronically infected with HIV type 1. AIDS Res Hum Retroviruses. 2007;23:67–76. doi: 10.1089/aid.2006.0114. [DOI] [PubMed] [Google Scholar]
- 20.Guwatudde D, Wabwire-Mangen F, Eller LA, et al. Relatively low HIV infection rates in rural Uganda, but with high potential for a rise: a cohort study in Kayunga District, Uganda. PLoS ONE. 2009;4:e4145. doi: 10.1371/journal.pone.0004145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Letvin NL. Progress and obstacles in the development of an AIDS vaccine. Nat Rev Immunol. 2006;6:930–9. doi: 10.1038/nri1959. [DOI] [PubMed] [Google Scholar]
- 22.Quinn TC, Wawer MJ, Sewankambo N, et al. Viral load and heterosexual transmission of human immunodeficiency virus type 1. Rakai Project Study Group. N Engl J Med. 2000;342:921–9. doi: 10.1056/NEJM200003303421303. [DOI] [PubMed] [Google Scholar]
- 23.McElrath MJ, De Rosa SC, Moodie Z, et al. HIV-1 vaccine-induced immunity in the test-of-concept Step Study: a case-cohort analysis. Lancet. 2008;372:1894–905. doi: 10.1016/S0140-6736(08)61592-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Schuetz A, Haule A, Schunk M, et al. Cellular Immune Responses in HIV-1 Uninfected Adult Tanzanian Volunteers Enrolled in a Phase I/II Multiclade HIV-1 DNA Plasmid Vaccine (VRC-HIVDNA016-00-VP)/Adenovirus-5 Vector (VRC-HIVADV014-00-VP) Boost Vaccine Trial. Conference on Retroviruses and Opportunistic Infections; Boston, Massachusettes. 2008. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.