

Abstract
Infants can anticipate the future location of a moving object and execute a predictive reach to intercept the object. When a moving object is temporarily hidden by darkness or occlusion, 6-month-old infants’ reaching is perturbed but performance on darkness trials is significantly better than occlusion trials. How does this reaching behavior change over development? Experiment 1 tested predictive reaching of 6- and 9-month-old infants. While there was an increase in the overall number of reaches with increasing age, there were significantly fewer predictive reaches during the occlusion compared to visible trials and no age-related changes in this pattern. The decrease in performance found in Experiment 1 is likely to apply not only to the object representations formed by infants but also those formed by adults. In Experiment 2 we tested adults with a similar reaching task. Like infants, the adults were most accurate when the target was continuously visible and performance in darkness trials was significantly better than occlusion trials, providing evidence that there is something specific about occlusion that makes it more difficult than merely lack of visibility. Together, these findings suggest that infants’ and adults’ capacities to represent objects have similar signatures throughout development.
Keywords: occlusion, infancy, reaching
1. Introduction
Occlusion happens. Whether you are searching for keys in a cluttered room or trying to track a ball while it passes behind your tennis partner, we are often maintaining expectations about objects that are temporarily hidden. This is a universal problem not specific to age, SES, culture, or gender. As we move around the environment many objects pass from visible to occluded and occluded to visible. Adults have extensive experience with these types of transitions and are able to maintain a coherent understanding of objects in the immediate environment and predict how objects should behave and interact. This knowledge allows us to organize our actions towards objects. The development of object knowledge has been the focus of research for over 50 years (Piaget, 1954). One general consensus that has emerged from the literature is that the relationship between object representation and action is complex (Gredeback & von Hofsten, 2007). There are two mountains of evidence concerning knowledge about occluded objects over development and surprisingly they point in opposite directions.
One set of findings suggests that the ability to represent objects that are occluded is evident in infancy and continuous through development. Two-month-old infants have expectations about the location, solidity, and persistence of hidden objects (Aguiar & Baillargeon, 1999; Hespos & Baillargeon, 2001b; Spelke, Breinlinger, Macomber, & Jacobson, 1992). Although these early representations are not mature, further research suggests that over the course of the first year infants gradually come to represent information about object width, height, shape, pattern, and color (Aguiar & Baillargeon, 2003; Hespos & Baillargeon, 2001a; Wang, Baillargeon, & Brueckner, 2004; Wilcox & Chapa, 2004; Wilcox & Schweinle, 2002). Representing occluded objects is not specific to humans. Non-human primates are able to track the number, location, and trajectory of hidden objects (Filion, Washburn, & Gulledge, 1996; Hauser, MacNeilage, & Ware, 1996a; Santos, 2004). These findings suggest that the ability to represent occluded objects is evident early and shared by other species, and that the developmental changes are processes of elaboration and refinement.
Experiments on object perception provide further support for this view and lend insight to the nature of their underlying mechanisms. Object perception has been well studied at the behavioral level in infants over the past 20 years (for reviews see, Baillargeon, 2004; Kellman & Arterberry, 1998; Spelke & Newport, 1998). This research has charted the developmental time course of a variety of central aspects of object perception including figure-ground organization (Termine, Hrynick, Kestenbaum, Gleitman, & Spelke, 1987), object and face discrimination (Bushnell, Sai, & Mullin, 1989), size and shape constancy (Slater, Mattock, & Brown, 1990), perception of shape over changes in location and size (Milewski, 1979), perception of partly occluded forms and objects (Kawabata, Gyoba, Inoue, & Ohtsubo, 1999; Kellman & Spelke, 1983) and categorization of objects into domains such as faces, animals, and artifacts (Mandler, 1992; Quinn & Eimas, 1998). In all these cases, studies of young infants (birth – 3 months) have found evidence for perceptual abilities that resemble those of adults. Moreover, studies of older infants have found progressive changes in the efficiency and sensitivity of their perceptual processing. The general conclusion gleaned from these studies is that the signatures of the infant and adult object perceptual systems are similar, providing evidence for a common underlying mechanism guiding behavior in these instances.
While the findings described above portray continuity in the ability to represent occluded objects through development, there is a different mountain of evidence that suggests discontinuity in actions toward occluded objects. Note that we draw a distinction between infants’ ability to represent and to act on an object that becomes hidden behind an occluder. Recent reports by Rosander & von Hofsten (2004) and von Hofsten, Kochukhova & Rosander (2007) suggest that under simple occlusion conditions, infants as young as 4 months engage in predictive tracking, taking into account the speed and path of the object to correctly anticipate its reappearance at the far edge of an occluder. However, there is no positive evidence that infants younger than 3 months consistently anticipate the reappearance of an object that moves behind an occluder (Rosander & von Hofsten 2004). Taken together there appears to be a discontinuity between the knowledge revealed in infants’ successful representation of occluded objects at 2 months of age and the inability to engage in predictive tracking of occluded objects until 4 months of age.
Further evidence of the discontinuity comes from action tasks requiring infants to retrieve hidden objects. For example, infants fail to retrieve hidden objects until about 9 months of age (Diamond & Lee, 2000; Piaget, 1954). Moreover, children perform surprisingly poorly on manual search tasks that were modeled after looking time studies. Two-year-old children viewed a ball rolling down a ramp toward a barrier. The ramp was mostly hidden behind a screen with 4 doors that could be opened to access the ball where it stopped against a barrier positioned on the ramp. The toddlers were encouraged to retrieve the ball by opening one of four doors in the screen. Surprisingly, toddlers choose among the doors at random, apparently oblivious to the relations between the ball, ramp, and barrier (Berthier, DeBlois, Poirier, Novak, & Clifton, 2000). This robust finding has been replicated numerous times in experiments that varied the direction of motion (e.g., vertical, left, and right)(Hood, Carey, & Prasada, 2000; Powell, Berthier, & Moore, 1979), used a transparent screen so that the children could see the object traverse between each door (Butler, Berthier, & Clifton, 2002), measured the amount of visual tracking during performance (Mash, Keen, & Berthier, 2003), and varied the proximity of a visual cue that reminded the participants where the object was located above the screen (Shutts, Keen, & Spelke, 2006). Toddlers’ failures to perform a successful reach were especially striking because two additional studies confirmed that, like infants, toddlers looked longer at an event that revealed the ball in an impossible position (Hood, Cole-Davies, & Dias, 2003; Mash, Novak, Berthier, & Keen, 2006).
If we evaluate knowledge about objects solely on these reaching studies, then it appears that there are qualitative differences in the cognitive capacities of children and adults. Data from these action tasks suggest that tracking occluded objects has a protracted development, emerges in a piecemeal fashion for most of the first year, and is still a fragile system in toddlers. Thus, there appears to be a discontinuity between infants’ ability to represent and to act on an object that becomes hidden behind an occluder.
1.1. The present research
These apparently inconsistent findings are puzzling in light of the fact that representing and acting on objects appears to require the same knowledge. Standing in the valley between these two mountains of evidence about objects we ask: Why do infants perform so badly on Piaget’s search tasks? Why do toddlers fail to retrieve a toy rolled down a ramp?
One possible answer to these questions is that object representations in infants are subject to two general limits found in studies of adults. According to the visibility hypothesis, one limit stems from effects of memory on representations: when an object ceases to be visible, its existence and location must be remembered, and the precision of this memory may decline as the length of occlusion increases. According to the competition hypothesis, a second limit stems from the effects of attention on representations: multiple objects in a scene may compete for attention, and this competition may reduce the precision with which each object is represented, whether it is visible or hidden. Infants and children may show evidence of representing occluded objects in looking time studies, but not reaching studies, because reaching requires a more precise representation of an object’s position, shape and motion (Goodale & Milner, 1995).
Jonsson and von Hofsten (2003) compared infants’ predictive reaching for objects that were hidden by darkness or occlusion for different durations of time. The experiment revealed three effects. First, infants reached most frequently and accurately when the object was continuously visible. Second, reaching was more impaired by longer than by shorter periods of hiding. Both of these findings are consistent with the visibility hypothesis. Third, reaching was more impaired by occlusion than by darkness. The competition hypothesis, motivated by studies of object-directed attention in adults (e.g., Alvarez & Franconeri, 2007), can account for this effect: representations of distinct objects may compete for attention, reducing the precision of each object representation as the number of other objects in the scene increases (Jonsson & von Hofsten, 2003; Munakata & Stedron, 2002). When an object is hidden behind an occluder, it may suffer a double loss of precision due both to its lack of visibility and to competition from its visible occluder. In contrast, an object that vanishes into darkness suffers a loss of precision only because of its lack of visibility and so should be represented more precisely than an occluded object, though less precisely than a visible one.
The visibility and competition hypotheses support a key developmental prediction: occlusion and darkness should have the same effects on object representations at all ages. To test this prediction, we created a situation that distinguished representational abilities from the abilities involved in trying to coordinate an action toward an object. The experiments presented a reaching task that required representing occluded objects but did not require execution of a complex motor response. We presented this task to infants who straddle the ages over which Piagetian search tasks revealed developmental discontinuities. In addition, we also tested adults who are well beyond the age of the discontinuous changes found in children. If there is a dramatic improvement in reaching for an occluded object, relative to reaching for a visible object between 6 and 9 months of age, that finding would suggest that older infants and adults process these events in qualitatively different ways from younger infants. However, difficulty with occlusion trials at 9 months and in adults, would support the view that there are common mechanisms for representing hidden objects across development.
In these experiments, we used a predictive reaching task. We discriminate between a predictive reach (one initiated before the object enters the reaching space) and a reactive reach (one initiated after the object enters the reaching space). Von Hofsten (1980) was the first to show that young infants can make a predictive reach for moving objects. When a continuously visible, out-of-reach object begins to move smoothly toward them, infants as young as 4 months typically will attempt to grab the object, initiating their action before the object enters their reaching space and aiming ahead of the object’s current position so as to intercept it when it comes within their range (von Hofsten, 1980; von Hofsten, Vishton, Spelke, Feng, & Rosander, 1998).
Predictive reaching experiments can serve to dissociate two effects of an occluder that otherwise are confounded: its effects on the visibility of an object and its effects on the actions needed to obtain the object. Spelke and von Hofsten (2001) therefore compared infants’ predictive reaching with and without an occluder placed over a portion of the object’s trajectory, such that the object was hidden briefly before it entered infants’ reaching space. Because the occluder was positioned outside the reaching area, it did not serve as a physical barrier to infants’ reaching: The action demands were identical in the visible and occluded trials. Moreover, infants’ head tracking in trials with the occluder suggested that they tracked the hidden object, because their head moved in anticipation of the object emerging from the other side of the occluder (Jonsson & von Hofsten, 2003; Spelke & von Hofsten, 2001). Nevertheless, the presence of the occluder interfered with young infants’ ability to perform a predictive reach. Predictive reaching was impaired by an occluder that blocked infants’ view of the object, even though it did not affect the motor actions needed to retrieve the object.
2. Experiment 1
We replicated Spelke & von Hofsten’s (2001) findings with young infants and extended them to older infants. We coded predictive and reactive reaches. The reactive reaches provided a measure of overall engagement in the task. Our principal analyses focused on predictive reaches, however, because they are anticipatory acts and hence more likely to be influenced by object representations. To encourage predictive reaching, we moved the object at a quick speed such that a reactive reach, initiated once the object was within reach, was unlikely to intercept the object. The independent variable that we manipulated was whether the object was visible or occluded during the critical window when the predictive reach needed to begin.
Infants aged 6 and 9 months were seated in an infant seat in front of a large white board (Fig. 1). A small toy with a rattle inside was given to the infants to explore. After a short time, the toy was taken and placed on the board out of the infant’s reach. The infant’s attention was drawn to the toy and the toy moved in a diagonal, linear trajectory across the infant’s reaching space in the fronto-parallel plane. The object came within reach during a short period when it was right in front of the infant. The infant was encouraged to reach/catch the moving object and remove it from the board. The experiment included 24 trials. During the first and last 6 trials the object was visible throughout its motion to establish baseline reaching activity. During the 12 middle trials, the object was temporarily hidden when it passed under a tunnel. The interval of non-visibility was 600 ms. Piaget’s search task where infants retrieve a hidden object suggests that there is a qualitative shift in performance between 6 and 9 months. If there is discontinuity in infants’ performance when an object becomes temporarily occluded, then the older infants should show improved performance in occlusion trials relative to the younger infants. However, if failures in reaching performance are due to the nature of the representation then both age groups should have fewer predictive reaches on trials with occlusion.
Fig. 1. Schematic of the set up used in Experiment 1.

The infant seat was centered in front of a large white board. The yellow circle signifies the toy. The solid line is the toy’s trajectory during the trial and the dotted line was the path the toy took between trials. The pink shaded region represents the infant’s reaching space and the black rectangle represents the size and position of the occluder.
2.1. Method
2.1.1. Participants
The participants were 41 healthy, full-term infants (20 male, 21 female). There were 17 6-month-olds (range = 6 months, 8 days to 7 months, 2 days, M = 6 months, 19 days), and 24 9-month-olds (range = 8 months, 11 days to 10 months, 20 days M = 9 months, 6 days). The children were recruited by mail and came from the surrounding area. The parents were offered $5 reimbursement for their travel expenses. Eleven additional infants were tested but eliminated from the final sample, 5 because of fussiness, 4 because the machine that moved the toy malfunctioned, and 2 refused to reach on any trial.
As stated above we were particularly interested in infants who performed predictive reaches instead of reactive reaches. Thirty infants (out of the entire sample of 41 infants) performed a predictive reach on at least one trial. There were 13 6-month-olds (range = 6 months, 8 days to 7 months, 2 days, M = 6 months, 20 days), and 17 9-month-olds (range = 8 months, 11 days to 10 months, 20 days, M = 9 months, 11 days). The average number of predictive reaches per child was 3 (2.07 for 6-month-old and 3.79 for the 8-month-old infants)
2.1.2. Display and apparatus
To be able to produce linear motion on a relatively large surface with precision, we used a computer-controlled plane plotter (Roland DPX-4600) whose pen was replaced with a small magnet (von Hofsten, Vishton, Spelke, Feng, & Rosander, 1998). The plotting area was topped with a sheet of aluminum that was painted white, coated with a silicone lubricant and placed on a supporting structure such that it tilted 15 degrees forward from the vertical position (this was identical to the degree that the infant was reclined in the infant seat and allowed the board to be at a comfortable position in front of the infant). The aluminum sheet served as the background for a toy, which was supported by a 10 cm wooden dowel attached to a magnet. When the magnet on the toy’s supporting rod was placed on the aluminum sheet directly over the plotter magnet, the combined attraction held the toy in place and caused it to undergo whatever motion was produced by the plotter. By using the commands originally intended to direct the motion of the plotter pen, this apparatus enabled us to direct the motion of the toy very precisely.
The toy always moved in a linear diagonal path with constant velocity (30 cm/s) from the infant’s upper left to their lower right (Fig. 1). The toy was a plush bear, approximately 12 cm tall and 5 cm wide. On trials involving occlusion, the occluder was positioned above and to the left of the infant’s reaching space. The occluder was a table made out of black foam core. It was 15.8 × 16.71 cm, and 15 cm from the board. There were dowels in each of the 4 corners with white suction cups at the base that allowed them to remain secure against the white board.
To code the infant’s actions we recorded two separate video images from perpendicular angles. The sagittal view was centered above the infant’s midline to capture the left to right sweep of the reaching space and the lateral view was to the infant’s right side to capture a top to bottom view of the reaching space. Using a video mixer the images from both cameras were recorded side by side on a single screen.
2.1.3. Design and procedure
Each infant did 24 trials. The experiment was divided into 4 blocks, of 6 trials each. During the first and last block the toy was fully visible. For the two middle blocks the toy was occluded during part of its motion. This ABBA design allowed us to test for learning effects by comparing reaching during the first and second half of the experiment to gage overall engagement in the task.
When parents arrived with their child, the procedure was explained to them and they signed a written consent. The infant was given several minutes to play with the toy attached to the magnet and to become accustomed to the new surroundings. The infant was placed in a standard infant chair (Mothercare Inc.) approximately 25 cm from the white board, and the toy was positioned on the board directly in front of the infant. In order to make the task more attractive, the infants were given the opportunity to retrieve the toy from the white board twice before the experiment started. The toy was then moved to the far upper left position of the screen and the infant’s attention was drawn to the toy, after which the experimenter stepped back and pressed a computer key to initiate the toy’s motion.
During the experiment the toy always started out of reach in the upper left corner with respect to the infant. The toy moved down and to the right on a diagonal trajectory at a constant speed of 30 cm/s (Fig. 1). The motion path was 128 cm long and measured 83 cm in the vertical dimension and 97 cm in the horizontal dimension. The periods of non-visibility by occlusion started approximately 37 cm into the trajectory. The toy always became visible again after 18 cm of occlusion. The toy passed in front of the infant’s midpoint 64 cm into the trajectory (at this point the toy had been in the infant’s reaching space for about13 cm). The pink portion of the Fig. 1 represents the infant’s reaching space. The black rectangle represents the occluder. If the infant retrieved the object, it was gently removed from the infant’s hand and manually repositioned at the same starting position. The plotter made a noise during motion but this noise did not originate from the moving object but rather from the stationary motors of the plotter. It was not possible to determine the position of the moving object from the noise made by the plotter.
2.1.4. Coding
During the experiment a trained observer watched the infant and recorded whether the infant made any action toward the object at any time during the diagonal portion of the trajectory. These behaviors were used as a general measure of engagement in the task.
Identification of predictive reaches was made from frame-by-frame analyses of the video using the same technique as the other studies that have measured predictive reaching (Jonsson & von Hofsten, 2003; Spelke & von Hofsten 2001; von Hofsten, et al., 1998). A reach takes at least 300 ms to prepare and at least another 300 ms to carry out, reaches arriving at the object on the left side of the midline had to be planned before the object reappeared. A predictive reach was coded when the hand came within 7.5 cm of the object in all three dimensions of space for at least one frame during the period from when it had reappeared in the occlusion condition until it passed the midline. The 7.5 cm distance was the maximum distance that an infant could be from the object with hopes of intercepting it while it was within reach. For the purpose of comparison, the same criteria were applied when coding reaches in Block 1 and 4 where the object was continuously visible.
For coding purposes, two circles with their centers marked were drawn on a sheet of transparent plastic. The radius of the circles corresponded to 7.5 cm in each of the two video projection planes (lateral and sagittal) and when the hand was within both of these circles it was closer than 7.5 cm in all dimensions of space. To judge whether a reach was within 7.5 cm of the object in each of the two projection planes, the plastic sheet was placed over the video screen with the center of the appropriate circle positioned at the center of the object seen on the video. If the hand was within the circle on each of the video projections, a predictive reach was coded. All trials were coded by two people independently. Cohen’s kappa was found to be .92.
2.2. Results
The data were analyzed for the total number of reaches (predictive and reactive, n = 41) across trials to assess overall engagement in the task and learning effects over trials. We found no significant difference in number of reaches comparing the first half of the experiment (M = 64% of trials) to the second half of the experiment (M = 56% of trials, t (81) = 1.60, p = .11) suggesting that infants maintained interest in the task throughout the experiment.
Next we analyzed the predictive reaching data (n = 30) by analysis of variance (ANOVA) with the within-subject factors of condition (visible or occluded) and block (first or second) and age as a between-subject variable (6 or 9 months). The analysis revealed a significant main effect for condition (F (1,28) = 21.87, p < .001, η2 = .44) demonstrating that infants made more predictive reaches during visible (M = 3.05) compared to occluded (M = 1.78) trials (Fig. 2). There was a significant main effect of age (F(1, 28) = 8.63, p = .007 η2 = .24) suggesting that as infants got older they reached more often regardless of condition (M for 6 months = 1.52, M for 9 months = 3.10). The interaction between condition and age was not significant (F(1, 28) < 1) (Fig. 3).
Fig. 2. Results from Experiment 1.
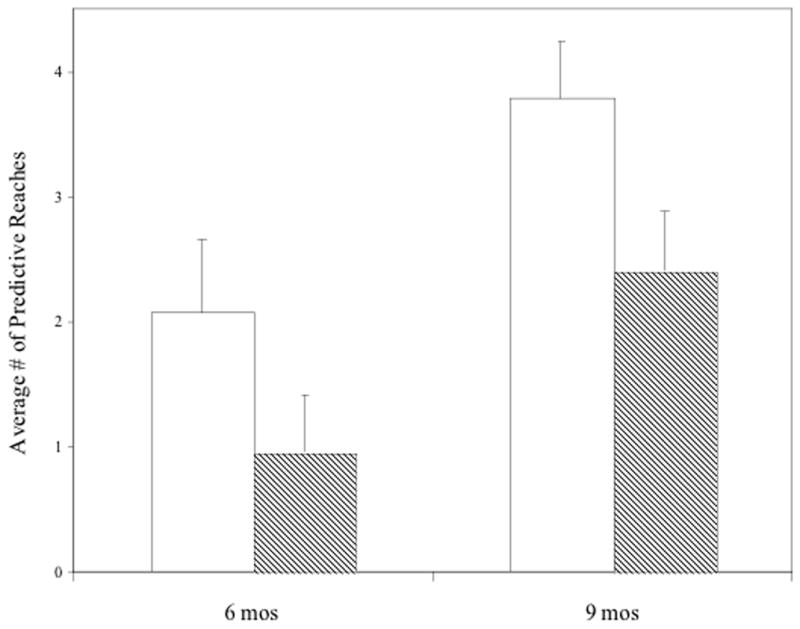
Average number of predictive reaches during visible (white) and occlusion (lines) conditions. Error bars represent standard error.
Fig. 3. Results from Experiment 1.
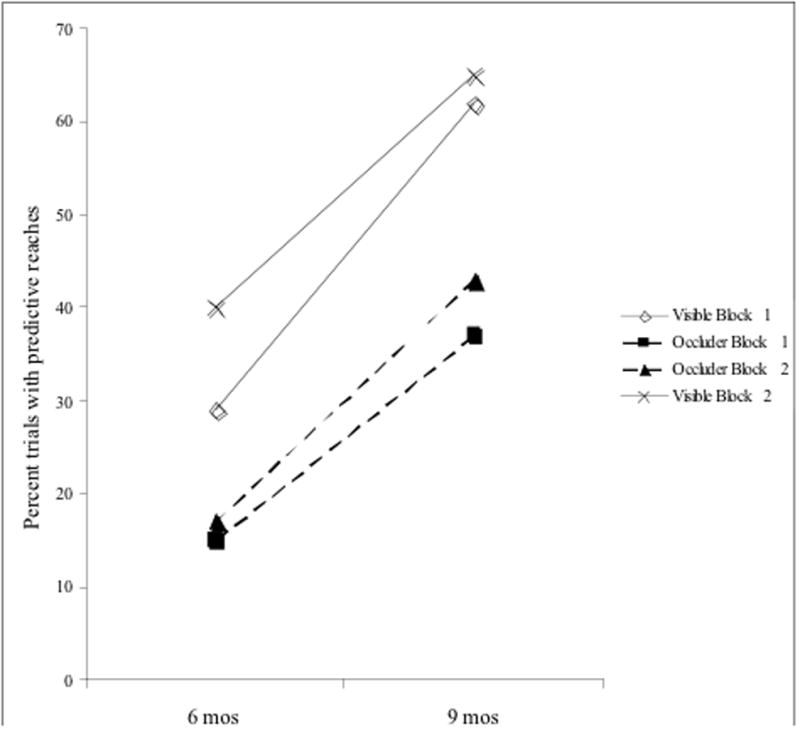
Percentage of trials that had predictive reaching. Solid lines represent visible trials and dashed lines were occlusion trials.
Examination of the participants’ individual responses revealed that 22 out of 30 infants had more predictive reaches during the visible than the occluded trials (cumulative binomial probability, p = .008). Further analysis revealed that the pattern of decreased reaching during the occluded compared to the visible trials was upheld in each age group independently: for 6-month-olds F(1,12) = 9.67, p = .009, η2 = .45, for 9-month-olds F(1,16) = 13.41, p = .002, η2 = .46.
2.3. Discussion
There were two main findings from this experiment. First, there was a significant increase in the number or reaches over development. Regardless of whether the object was visible or occluded, 9-month-old infants reached significantly more often than 6-month-old infants. The second main finding was that as a group and as individuals, infants showed a significant decrease in reaching during occlusion trials. More specifically, as infants get older there is an increase in the number of predictive reaches, but the influence of occlusion remained constant.
These findings contribute new insights to the mechanisms that guide our ability to coordinate actions toward moving objects. We created a situation that distinguished representation abilities from the abilities involved in trying to coordinate an action toward an object. These data provide evidence that occlusion imposed a representational cost (e.g., that there is significantly less reaching during occlusion compared to visible trials). Our findings replicate and extend research by Spelke and von Hofsten (2001) and Jonsson and von Hofsten (2003) to 9-month-old infants, an age where infants typically succeed in the Piagetian search tasks for hidden objects. Furthermore these findings complement the research from Gredebäck and von Hofsten (2004) on visual tracking of occluded objects. Gredebäck and von Hofsten did a longitudinal study on infants between 6 and 12 months of age and found no qualitative shifts in their ability to make predictive eye movements in anticipation of an object emerging from the side of an occluder.
The visibility hypothesis, that infants’ object representations are more precise when objects are visible than when they are hidden, could explain why infants reach predictively for moving, visible objects and look predictively at moving, temporarily occluded objects, but fail to reach predictively for moving, temporarily occluded objects. To catch a moving object, one must represent considerable information about the object, including its size, shape, path, and speed of motion. When the object is continuously visible, infants’ representations evidently are adequate to guide appropriate reaching. However, when the object is hidden, their representation of its properties may become too imprecise to represent and/or the cost associated with guiding effective attempts to intercept it overwhelm their ability. The visibility hypothesis is similar to the graded representation hypothesis (Shinskey & Munakata, 2003, 2005) in that both center on the idea that representations are noisy but become less noisy over development.
3. Experiment 2
Because our analysis of object representations in infants is based on properties of object representations first uncovered in adults, this analysis should apply not only to the object representations formed by infants but also to those formed by adults. To test this prediction, we conducted an experiment on adults, modeled on the studies of predictive reaching in infants. We measured participants actions toward a moving object under 3 conditions: continuously visible, hidden by darkness, or hidden by occlusion. By presenting the same adults with two manners of hiding the object (darkness and occlusion), we build a bridge to the infant studies where performance in predictive reaching during darkness is significantly better than predictive reaching during occlusion (Jonsson & von Hofsten, 2003; Spelke & von Hofsten, 2001). The significant difference between darkness and occlusion is likely to be due to the nature of the representation because the hidden portion of the trajectory is matched across conditions in terms of position and duration. In addition, the same action is required in both conditions. Based on the notion that there are similarities in the nature of the representation over development, we predict that adults should show a similar pattern of results, namely performance in visible trials will be the best and performance in darkness will be significantly better than the occlusion condition.
The adults stood on the right side of the display used in the reaching studies with infants. Because adults were likely to perform at ceiling on the conditions presented to infants, three changes were made to increase the task difficulty: we doubled the speed of object motion (60 cm/s), increased the duration of interrupted visibility (1 s) and showed a variety of linear trajectories (6). Adults were randomly assigned to one of two conditions. One task involved predictive reaching - to mark a moving target with a marker held in their hand (Fig. 4), and the other task involved predicting the trajectory of a linear path after viewing only a portion of the trajectory (Fig. 5). We had identical predictions for the target and trajectory conditions. The rationale for the adult study was to test for parallels in infants’ and adults’ abilities to coordinate actions and intercept a moving target. If there are similar signatures, then adults’ performance should be the best in the visible trials, less accurate in the blackout trials, and the least accurate in the occlusion trials.
Fig. 4. Schematic of the set up used in Experiment 2 for the target task.

The adults stood at the right side of the apparatus with their hand holding a marker under the white cloth that hung over the right portion of the stage. The concentric circles are the target that participants tried to mark. The diagonal red arrow represents 1 of the 6 possible linear trajectories that the target traversed. In the blackout condition, the vertical red line represents the light beam that extinguished the room lights for 1 s when the target passed through the beam. In the occlusion condition, the black square represents the size and position of the occluder.
Fig. 5. Schematic of the set up used in Experiment 2 for the trajectory task.

This condition was similar to the target task in every respect except on the right side of the board the cloth was replaced with white tape and participants were asked to mark a spot on the tape where they predicted the trajectory would cross the tape.
In the target task, the toy used in the infant studies was replaced with a platform that was parallel to the white board, on the platform we glued a piece of paper that had concentric circles like a bulls-eye target. The participant’s task was to stand on the right side of the white board with a pen in hand, view the linear path of the object, and tap the target in the center when it reached the right side of the board. The task was challenging because the target was constantly moving and therefore available only briefly. We tallied the amount of ink marks on the target for each condition.
In the trajectory task, a separate group of participants viewed the linear trajectory and marked the point at the right edge of the board where the object should intercept the tape stuck to the board. We tallied the distance between the predicted and the actual position of the trajectory.
3.1. Methods
3.1.1. Participants
The participants were 28 adults 12 male and 16 female (range = 18 years to 43 years, M = 22 years). Twelve of the adults were assigned to the target condition the remaining 16 were assigned to the trajectory condition. Adults were paid $5 for their participation.
3.1.2. Display and apparatus
We used the same apparatus as Experiment 1. The toy that the infants reached for was replaced with a foam core platform that was attached to a 10 cm wooden dowel. The platform was circular and 11 cm in diameter with concentric circles on it.
The objects always moved in a linear diagonal path with constant velocity (60 cm/s). There were 6 different paths presented in a random order. The first path was identical to the path described in Experiment 1. The subsequent 5 paths had start points on the left side of the board 14, 28, 56, 73, and 83 cm below the original. The endpoints were 14, 28, 56, 73, and 83 cm above the original respectively. Three of the paths started in upper half of the board and ended in the lower half the other three started on the lower half of the board and ended in the upper half. A photo-cell switch triggered the extinction of room light with a timer set to the blackout period. The lights went out instantaneously. The occluder was 86 cm tall and 76 cm wide made out of black foam core. The occluder and blackout portions were set up so that the object became hidden at exactly the same time and place across conditions so the only difference across these conditions was the manner of hiding. The period of hiding started approximately 40 cm into the trajectory.
In the target condition, there was a 20 cm wide white cloth that was attached to the top and bottom edge of the white board. The participants held the marker in their hand under the cloth so that they were forced to rely on the visible portion of the trajectory not the endpoint on the right side of the board. (Pilot studies revealed that performance without the white cloth was at ceiling.) The target was under the white cloth and hence available to marking for 600 ms. A different color pen was used on each trial in a block to insure that two marks made on a single trial were not counted twice. The paper with the bulls-eye was replaced after each block.
In the trajectory condition, there was a white piece of masking tape (5 cm wide) affixed to the right side of the board so the participant could mark the tape where they predicted the linear trajectory would intercept. The tape was replaced after each block to ensure that guesses from prior conditions did not influence performance. During the visible trials, the linear trajectory was 64 cm long and the experimenter removed the target as soon as it stopped because pilot studies revealed that performance was at ceiling when the adults saw more of the trajectory.
3.1.3. Design and procedure
The experimental design was the same for the target and trajectory conditions. Each adult completed 36 trials presented in 6 blocks. There were 3 conditions (visible, blackout, and occlusion) each presented 2 times. Within each block we presented 6 different linear trajectories. The order of the 6 trajectories was randomized. The blocks were presented in a ABCCBA order to assess learning over the course of the experiment. The visible condition was always presented first and last. The presentation of the two hidden conditions (blackout or occlusion) was counterbalanced across subjects.
3.1.4. Coding
For the target condition, the coding was simply a tally of the amount of pen marks on the target across conditions subtracted from the total possible (6) to yield the number of misses. For the trajectory condition the coding was a measurement of the absolute distance between the mark on the tape and the correct intercept. The absolute difference did not take into account whether the mistake was above or below the correct location. All of the coding was verified by a second person independently and discrepancies were coded another time until agreement was obtained.
3.2. Results
Figures 6 and 7 reveal that the results went in the predicted direction across conditions in both the target and trajectory conditions. The target data were analyzed by repeated measures ANOVA with within-subject factors of condition (visible, blackout, and occluder) and block (first and second). There was a significant main effect for condition (F(2, 30) = 14.14, p < .001, η2 = .49) demonstrating that the lowest number of misses were in the visible condition (M = .78) followed by the blackout condition (M = 1.63) and then the occlusion (M = 2.34) condition. Further t-tests revealed that all conditions were significantly different from each other (visible vs. blackout t(15) = 3.01, p < .009; visible vs. occlusion t(15) = 5.17, p < .001; blackout vs. occlusion t(15) = 2.40, p < .03).
Fig. 6. Results from Experiment 2 target task.
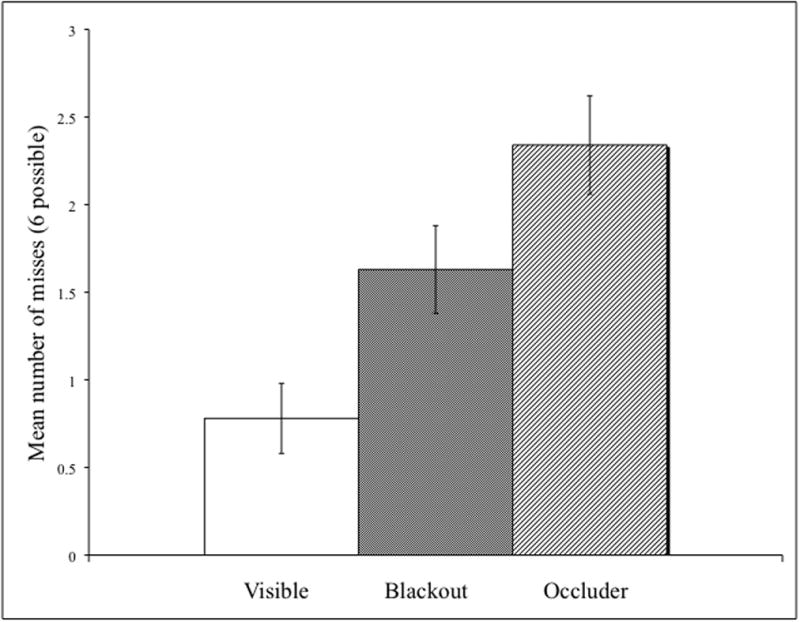
Average number of misses for visible (white), blackout (shaded), and occlusion (lines) conditions. Error bars represent standard error.
Fig. 7. Results from Experiment 2 trajectory task.
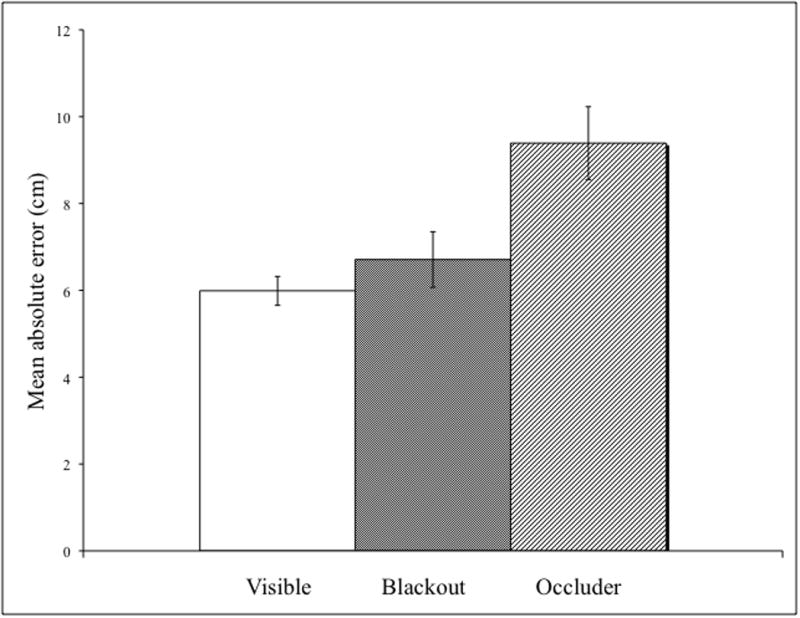
Average absolute error for visible (white), blackout (shaded), and occlusion (lines) conditions. Error bars represent standard error.
The trajectory data were analyzed in the same manner as the target condition. There was a significant main effect for condition (F(2,22) = 13.27, p < .001, η2 = .55) demonstrating that the least amount of error was in the visible condition (M = 5.99), next was the blackout condition (M = 6.71) and the most error was in the occlusion (M = 9.39) condition. Further t-tests revealed that the occlusion condition was significantly different from the visible and blackout conditions (visible vs. occlusion t(11) = 4.62, p < .001; blackout vs. occlusion t(11) = 3.65, p < .004). The visible and blackout conditions were not significantly different (visible vs. blackout t(11) = 1.19, p = .26). Surprisingly, there was a significant main effect for block (F(1,11) = 22.64 p < .001, η2 = .67) demonstrating that performance got worse from the first block (M = 6.26) to the second block (M = 8.46).
3.3. Discussion
There were two main findings from this experiment. First, adults had better performance when the target was visible compared to when the target was hidden for a portion of its trajectory. Second, there were significant differences in behavior depending on whether the object was hidden by darkness or occlusion. In both the target and trajectory conditions, the performance in the blackout condition was significantly better than performance in the occlusion condition.1 Since the conditions were matched for duration and position the difference in performance must be related to the manner in which the object became hidden. This is the same pattern of results observed in experiments testing infants and adults using multiple object tracking paradigms (Franconeri, Pylyshyn, & Scholl, 2008; Jonsson & von Hofsten, 2003; Scholl & Pylyshyn, 1999; Spelke & von Hofsten, 2001).
This finding supports the visibility and competition hypothesis, during the occlusion condition the representation suffered a double loss of precision due both to its lack of visibility and to competition from the visible occluder. In contrast, the object that was hidden by darkness alone suffered loss of precision only because of its lack of visibility.
4. General Discussion
The main conclusions from Experiment 1 are that infants’ ability to reach for a moving object is perturbed when the object becomes briefly occluded. Experiment 2 complemented these results by showing that even adults reveal a disruption in performance during visible trials compared to trials when the object became briefly hidden. Between the two types of hidden trials, adult performance was significantly better in the blackout condition than the occlusion condition. These findings suggest that there is something specific about the manner in which an object is hidden that predicts performance. Together these findings suggest that infants’ and adults’ capacities to represent objects have similar signatures over development, in particular that representing an object hidden by occlusion is harder than representing an object hidden by darkness.
There is developmental change in the frequency of predictive reaching between 6 and 9 months:--the younger infants made fewer predictive reaches than the older infants—but this change does not appear to indicate a qualitative shift. The effect of the occlusion did not change across development, because all infants made fewer predictive reaches during occlusion compared to visible trials. Together these findings suggest continuity across development in terms of representing hidden objects and coordinating actions to intercept them. In terms of measurement equivalence, we had to make the infant and adult tasks different so that the adults did not perform at ceiling levels. However the fact that both infants and adults perform significantly better in darkness compared to occlusion is striking and confirms that there are similar signatures in this representational ability over development.
Our findings accord with the analyses of predictive reaching of von Hofsten and his collaborators (Jonsson & von Hofsten, 2003; Spelke & von Hofsten, 2001). Indeed, a wide range of studies are consistent with the thesis that object-directed reaching requires precise representations, that the precision of object representations increases with age, and that precision declines at all ages when objects are out of view (the visibility hypothesis) and when other objects compete for attention (the competition hypothesis). These interpretations are consistent with research on reaching for stationary objects as well. In search tasks involving stationary objects, there is typically no time pressure therefore one would expect the ability to coordinate an action would play a smaller role. Indeed, a variety of reaching tasks involving stationary objects reveal cognitive performance at levels equivalent to looking paradigms (Hespos & Baillargeon, 2006; 2008).
Our data revealed a decrement in performance when other objects competed for attention. These findings are consistent with a theory proposed by Keen and her colleagues, whereby children’s failures at object-directed reaching tasks increase with the overall cognitive load that the tasks impose (Berthier et al., 2001; Boudreau & Bushnell, 2000; Keen & Berthier, 2004; Keen, Carrico, Sylvia, & Berthier, 2003). They propose that infants have limited information processing resources and succeed in an action task only when the combined demands of the task do not exceed the resources. Task demands depend on the difficulties of the physical reasoning involved as well as on the difficulties of the actions involved. Applying this account to our findings, suggests that the difference between conditions must result from variation in representational demands, because the action demands were identical across visible, blackout, and occlusion conditions. The data reported here nevertheless are consistent with Keen’s account, because inter-object competition is one form of cognitive load, and indeed it impairs both infants’ and adults’ performance on our task.
Our findings accord with the evidence that human infants have a capacity to represent occluded objects as early as two months of age (Hespos & Baillargeon, 2001b; Spelke et al. 1992), and that the capacity undergoes no qualitative reorganization as children grow. The evidence for ontogenetic continuity complements evidence for phylogenetic continuity in the capacity to represent objects. In particular, non-human primates represent objects similarly to human infants both in preferential looking and in object search tasks (Filion, Washburn, & Gulledge, 1996; Hauser, MacNeilage, & Ware, 1996b; Santos, 2004). These findings mesh well with the view that basic mechanisms of object representation are constant over much of evolution and ontogeny.
Acknowledgments
This research was supported by grants from NIH to the first (HD-08124) and fourth authors (R37-HD23103). We would like to thank the parents of the infants and the adults who participated in this study. We thank Kirsten Condry for help in getting the machinery and software to cooperate. We thank the members of the SALLY group for their insightful comments.
Footnotes
It is interesting to note that in the trajectory task the cost of hiding the object in the blackout condition was minimal because the performance was not significantly different between the visible and blackout conditions.
References
- Aguiar A, Baillargeon R. 2.5-month-old infants’ reasoning about when objects should and should not be occluded. Cognitive Psychology. 1999;39(2):116–157. doi: 10.1006/cogp.1999.0717. [DOI] [PubMed] [Google Scholar]
- Aguiar A, Baillargeon R. Perseverative responding in a violation-of-expectation task in 6.5-month-old infants. Cognition. 2003;88(3):277–316. doi: 10.1016/s0010-0277(03)00044-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alvarez GA, Franconeri SL. How many objects can you track?: Evidence for a resource-limited attentive tracking mechanism. Journal of Vision. 2007;7(3):1 – 10. doi: 10.1167/7.13.14. [DOI] [PubMed] [Google Scholar]
- Baillargeon R. Infants’ physical world. Current Directions in Psychological Science. 2004;13(3):89–94. [Google Scholar]
- Berthier NE, Bertenthal BI, Seaks JD, Sylvia MR, Johnson RL, Clifton RK. Using object knowledge in visual tracking and reaching. Infancy. 2001;2(2):257–284. [Google Scholar]
- Berthier NE, DeBlois S, Poirier CR, Novak MA, Clifton RK. Where’s the ball? Two- and three-year-olds reason about unseen events. Developmental Psychology. 2000;36(3):394–401. doi: 10.1037//0012-1649.36.3.394. [DOI] [PubMed] [Google Scholar]
- Boudreau JP, Bushnell EW. Spilling thoughts: Configuring attentional resources in infants’ goal-directed actions. Infant Behavior & Development. 2000;23(3–4):543–566. [Google Scholar]
- Bushnell IWR, Sai F, Mullin JT. Neonatal recognition of the mother’s face. British Journal of Developmental Psychology. 1989;7(1):3–15. [Google Scholar]
- Butler SC, Berthier NE, Clifton RK. Two-year-olds’ search strategies and visual tracking in a hidden displacement task. Developmental Psychology. 2002;38(4):581–590. doi: 10.1037//0012-1649.38.4.581. [DOI] [PubMed] [Google Scholar]
- Diamond A, Lee E. Inability of five-month-old infants to retrieve a contiguous object: A failure of conceptual understanding or of control of action? Child Development. 2000;71(6):1477–1494. doi: 10.1111/1467-8624.00241. [DOI] [PubMed] [Google Scholar]
- Filion CM, Washburn DA, Gulledge JP. Can monkeys (Macaca mulatta) represent invisible displacement? Journal of Comparative Psychology. 1996;110(4):386–395. doi: 10.1037/0735-7036.110.4.386. [DOI] [PubMed] [Google Scholar]
- Franconeri SL, Pylyshyn ZW, Scholl BJ. Spatiotemporal cues for tracking multiple objects through occlusion. 2008. Manuscript under review. [DOI] [PubMed] [Google Scholar]
- Goodale MA, Milner AD. The Visual Brain in Action. New York: Oxford University Press; 1995. [Google Scholar]
- Gredeback G, von Hofsten C. Infants’ evolving representations of object motion during occlusion: A longitudinal study of 6-to 12-month-old infants. Infancy. 2004;6(2):165–184. doi: 10.1207/s15327078in0602_2. [DOI] [PubMed] [Google Scholar]
- Gredeback G, von Hofsten C. Taking an action perspective on infant’s object representations. In: von Hofsten C, Rosander K, editors. Progress in Brain Research. Vol. 164. B.V.: Elsevier; 2007. pp. 265–282. [DOI] [PubMed] [Google Scholar]
- Hauser MD, MacNeilage P, Ware M. Numerical representations in primates. Proceedings of the National Academy of Sciences of the United States of America. 1996a;93(4):1514–1517. doi: 10.1073/pnas.93.4.1514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauser MD, MacNeilage P, Ware M. Numerical representations in primates. Paper presented at the Proceedings of the National Academy of Sciences of the United States of America.; 1996b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hespos SJ, Baillargeon R. Infants’ knowledge about occlusion and containment events: A surprising discrepancy. Psychological Science. 2001a;12(2):141–147. doi: 10.1111/1467-9280.00324. [DOI] [PubMed] [Google Scholar]
- Hespos SJ, Baillargeon R. Reasoning about containment events in very young infants. Cognition. 2001b;78(3):207–245. doi: 10.1016/s0010-0277(00)00118-9. [DOI] [PubMed] [Google Scholar]
- Hespos SJ, Baillargeon R. Decalage in infants’ reasoning about occlusion and containment events: Converging evidence from action tasks. Cognition. 2006;99:B31–B41. doi: 10.1016/j.cognition.2005.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hespos SJ, Baillargeon R. Young infants’ actions reveal their developing knowledge of support variables: Converging evidence for violation-of-expectation findings. Cognition. 2008;107(1):304–316. doi: 10.1016/j.cognition.2007.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hood BM, Carey S, Prasada S. Predicting the Outcomes of Physical Events: Two-Year-Olds Fail to Reveal Knowledge of Solidity and Support. Child Development. 2000;71(6):1540–1554. doi: 10.1111/1467-8624.00247. [DOI] [PubMed] [Google Scholar]
- Hood BM, Cole-Davies V, Dias M. Looking and search measures of object knowledge in preschool children. Developmental Psychology. 2003;39(1):61–70. [PubMed] [Google Scholar]
- Jonsson B, von Hofsten C. Infants’ ability to track and reach for temporarily occluded objects. Developmental Science. 2003;6(1):86–99. [Google Scholar]
- Kawabata H, Gyoba J, Inoue H, Ohtsubo H. Visual completion of partly occluded grating in infants under 1 month of age. Vision Research. 1999;39(21):3586–3591. doi: 10.1016/s0042-6989(99)00060-7. [DOI] [PubMed] [Google Scholar]
- Keen RE, Berthier NE. Continuities and Discontinuities in Infants’ Representation of Objects and Events. In: Kail RV, editor. Advances in child development and behavior. Vol. 32. San Diego, CA: Elsevier Academic Press; 2004. pp. 243–279. [DOI] [PubMed] [Google Scholar]
- Keen RE, Carrico RL, Sylvia MR, Berthier NE. How infants use perceptual information to guide action. Developmental Science. 2003;6(6):221 – 231. [Google Scholar]
- Kellman PJ, Arterberry ME. The cradle of knowledge. Cambridge, MA: MIT Press; 1998. [Google Scholar]
- Kellman PJ, Spelke ES. Perception of Partly Occluded Objects in Infancy. Cognitive Psychology. 1983;15(4):483–524. doi: 10.1016/0010-0285(83)90017-8. [DOI] [PubMed] [Google Scholar]
- Mandler JM. How to Build a Baby II. Conceptual Primitives. Psychological Review. 1992;99(4):587–604. doi: 10.1037/0033-295x.99.4.587. [DOI] [PubMed] [Google Scholar]
- Mash C, Keen RE, Berthier NE. Visual Access and Attention in Two-Year-Olds’ Event Reasoning and Object Search. Infancy. 2003;4(3):371–388. [Google Scholar]
- Mash C, Novak E, Berthier NE, Keen RE. What do two-year-olds understand about hidden-object events? Developmental Psychology. 2006;42(2):263–271. doi: 10.1037/0012-1649.42.2.263. [DOI] [PubMed] [Google Scholar]
- Milewski AE. Visual discrimination and detection of configurational invariance in 3-month infants. Developmental Psychology. 1979;15(4):357–363. [Google Scholar]
- Munakata Y, Stedron JM. Memory for hidden objects in early infancy: Behavior, theory, and neural network simulation. In: Fagen JW, Hayne H, editors. Progress in infancy research. Vol. 2. Mahwah, NJ: Lawrence Erlbaum Associates, Publishers; 2002. pp. 25–69. [Google Scholar]
- Piaget J. The construction of reality in the child. Oxford, England: Basic Books; 1954. [Google Scholar]
- Powell GM, Berthier NE, Moore JW. Efferent neuronal control of the nictitating membrane response in rabbit (Oryctolagus cuniculus): A reexamination. Physiology & Behavior. 1979;23(2):299–308. doi: 10.1016/0031-9384(79)90371-8. [DOI] [PubMed] [Google Scholar]
- Quinn PC, Eimas PD. Evidence for a global categorical representation of humans by young infants. Journal of Experimental Child Psychology. 1998;69(3):151–174. doi: 10.1006/jecp.1998.2443. [DOI] [PubMed] [Google Scholar]
- Santos LR. ‘Core Knowledges’: A dissociation between spatiotemporal knowledge and contact-mechanics in a non-human primate? Developmental Science. 2004;7(2):167–174. doi: 10.1111/j.1467-7687.2004.00335.x. [DOI] [PubMed] [Google Scholar]
- Scholl BJ, Pylyshyn ZW. Tracking multiple items through occlusion: Clues to visual objecthood. Cognitive Psychology. 1999;38(2):259–290. doi: 10.1006/cogp.1998.0698. [DOI] [PubMed] [Google Scholar]
- Shutts K, Keen RE, Spelke ES. Object boundaries influence toddlers’ performance in a search task. Developmental Science. 2006;9(1):97–107. doi: 10.1111/j.1467-7687.2005.00466.x. [DOI] [PubMed] [Google Scholar]
- Slater A, Mattock A, Brown E. Size constancy at birth - Newborn-infants responses to retinal and real size. Journal of Experimental Child Psychology. 1990;49(2):314–322. doi: 10.1016/0022-0965(90)90061-c. [DOI] [PubMed] [Google Scholar]
- Spelke ES, Breinlinger K, Macomber J, Jacobson K. Origins of knowledge. Psychological Review. 1992;99(4):605–632. doi: 10.1037/0033-295x.99.4.605. [DOI] [PubMed] [Google Scholar]
- Spelke ES, Newport EL. Nativism, empiricism, and the development of knowledge. In: Damon W, Lerner RM, editors. Handbook of child psychology: Volume 1: Theorectical models of human development. 5. Hoboken, NJ: John Wiley & Sons, Inc; 1998. pp. 275–340. [Google Scholar]
- Spelke ES, von Hofsten C. Predictive reaching for occluded objects by 6-month-old infants. Journal of Cognition and Development. 2001;2(3):261–281. [Google Scholar]
- Termine N, Hrynick T, Kestenbaum R, Gleitman H, Spelke ES. Perceptual completion of surfaces in infancy. Journal of Experimental Psychology-Human Perception and Performance. 1987;13(4):524–532. doi: 10.1037//0096-1523.13.4.524. [DOI] [PubMed] [Google Scholar]
- von Hofsten C. Predictive reaching for moving objects by human infants. Journal of Experimental Child Psychology. 1980;30(3):369–382. doi: 10.1016/0022-0965(80)90043-0. [DOI] [PubMed] [Google Scholar]
- von Hofsten C, Vishton P, Spelke ES, Feng Q, Rosander K. Predictive action in infancy: tracking and reaching for moving objects. Cognition. 1998;67(3):255–285. doi: 10.1016/s0010-0277(98)00029-8. [DOI] [PubMed] [Google Scholar]
- Wang SH, Baillargeon R, Brueckner L. Young infants’ reasoning about hidden objects: evidence from violation-of-expectation tasks with test trials only. Cognition. 2004;93(3):167–198. doi: 10.1016/j.cognition.2003.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilcox T, Chapa C. Priming infants to attend to color and pattern information in an individuation task. Cognition. 2004;90(3):265–302. doi: 10.1016/s0010-0277(03)00147-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilcox T, Schweinle A. Object individuation and event mapping: Developmental changes in infants’ use of featural information. Developmental Science. 2002;5(1):132–150. [Google Scholar]