

Abstract
The hemagglutinin (HA) envelope protein of influenza viruses mediates essential viral functions, including receptor binding and membrane fusion, and is the major viral antigen for antibody neutralization. The 1957 H2N2 subtype (Asian flu) was one of the three great influenza pandemics of the last century and caused 1 million deaths globally from 1957 to 1968. Three crystal structures of 1957 H2 HAs have been determined at 1.60 to 1.75 Å resolutions to investigate the structural basis for their antigenicity and evolution from avian to human binding specificity that contributed to its introduction into the human population. These structures, which represent the highest resolutions yet recorded for a complete ectodomain of a glycosylated viral surface antigen, along with the results of glycan microarray binding analysis, suggest that a hydrophobicity switch at residue 226 and elongation of receptor-binding sites were both critical for avian H2 HA to acquire human receptor specificity. H2 influenza viruses continue to circulate in birds and pigs and, therefore, remain a substantial threat for transmission to humans. The H2 HA structure also reveals a highly conserved epitope that could be harnessed in the design of a broader and more universal influenza A virus vaccine.
Influenza (flu) is an infection of the respiratory tract that affects millions of people every year. In addition to the seasonal toll, three flu pandemics in the past century caused millions of deaths worldwide in relatively short time periods (27). In April 2009, a novel strain of influenza A virus H1N1 (S-OIV) with swine origin emerged in North America and has become the first influenza pandemic in 4 decades. To date, this new H1N1 pandemic has spread globally and caused at least 7,800 deaths (World Health Organization, http://www.who.int).
Hemagglutinin (HA) is the major surface envelope glycoprotein on influenza virus, and responsible for essential viral functions, such as binding to host receptors, viral entry, and membrane fusion (31). A key factor that determines the host range, restriction, and transmission of influenza virus is the specificity of HA for binding glycan receptors comprising terminal sialic acids linked to a vicinal galactose residue. HAs in avian viruses are specific for sialic acids with an α2,3-linkage, whereas in humans, the specificity is for sialic acids with an α2,6-linkage (Fig. 1a). This simple linkage difference likely contributes to the inability of most avian influenza viruses to become established and transmit in the human population (26). Influenza pandemics in humans are generally associated with nonhuman viruses of novel antigenicity acquiring specificity for human receptors. HA is also the principal antigen of influenza viruses and the main target for neutralizing antibodies.
FIG. 1.

Crystal structure of H2 HA. (a) Chemical structures of α2,3- and α2,6-linked glycans, with the terminal sialic acid and galactose shown here. (b) Overview of the 1957 H2 trimer. One of the monomers is highlighted in green (HA1) and blue (HA2), respectively. Five potential glycosylation sites are found on each monomer (as labeled). Glycans in the density map are shown in orange. (c) Receptor binding site of H2. Residues involved in receptor binding, as suggested by the H3 structures, are shown in sticks. Aromatic residues comprising the base of the binding site are absolutely conserved in various HA subtypes. Residues from the 220 loop and position 190 are critical for the receptor specificity switch in H1, H2, and H3.
Although future influenza pandemics seem inevitable, predicting the potential HA subtypes that will emerge remains a daunting task (41). To date, 16 HA subtypes have been identified and classified based on their antigenic properties (1). Theoretically, all influenza viruses new to the immune system of the human population today possess the potential to initiate a flu pandemic if their ability to enter human cells and transmit efficiently evolves. Historically, however, only viruses of three HA subtypes have acquired the ability to efficiently transmit from human to human, and these were responsible for the influenza pandemics of the last century: 1918 (H1N1), 1957 (H2N2), 1968 (H3N2), and 2009 (H1N1). In recent years, viruses of other HA subtypes (H5, H7, and H9) of avian origin have infected humans in sporadic cases and occasionally with very high mortality, such as H5N1 (2, 4, 10). A key barrier to avian flu becoming a human pandemic is its inefficient human-to-human transmission, which requires a switch of receptor specificity from α2,3- to α2,6-linked receptors. Although the H2 subtype has disappeared from the human population since 1968, it has reemerged in swine in the United States (19). Preparedness for future pandemics can be best addressed by rigorous characterization of the HA subtypes that have already caused pandemics, as well as development of therapeutic reagents that broadly target multiple influenza subtypes.
Here, we present three crystal structures of human H2 HA from the 1957 pandemic at resolutions of 1.60, 1.73, and 1.75 Å. These structures, which differ only by one or two residues in the receptor-binding site, represent the evolution of binding specificity for human-like receptors of avian origin during the 1957 H2N2 pandemic. Structural comparisons among the structures, along with glycan array binding studies, have shed new light on the requirements for avian H2 HA to adapt for human transmission.
MATERIALS AND METHODS
The gene corresponding to the ectodomain of HA from A/Japan/305/57 was inserted into a baculovirus expression vector modified from pAcGP67A (BD Biosciences Pharmingen) (35) and expressed as described previously (36). Mutations were generated by site-directed mutagenesis. Purified HA was digested with TPCK (l-1-tosylamide-2-phenylmethyl chloromethyl ketone)-treated trypsin (New England Biolab) and concentrated to 12 mg/ml in 20 mM Tris-HCl-50 mM NaCl (pH 8) and crystallized by sitting-drop vapor diffusion method at 22°C. The reservoir contains 25 to 28% polyethylene glycol 3000 (PEG 3000) and 0.1 M Tris (pH 7.8 to 8.5). Crystals were flash cooled in liquid nitrogen after a brief soaking in reservoir solution plus 10% ethylene glycol. Data sets were collected at synchrotron radiation sources and processed with HKL2000 (25). The structure of the H2 avian HA was solved by molecular replacement using Phaser (22) and a monomer of H5 HA (1JSM). The initial model was automatically built in ARP/wARP (18). The structure was then adjusted using COOT (9) and refined with REFMAC (23). Mutant structures of H2 HA were solved by molecular replacement using the avian H2 structure and were rebuilt in COOT. Statistics for data collection and structure refinement are presented in Table 1.
TABLE 1.
Data collection and refinement statistics
| Statistics | Value(s)a |
||
|---|---|---|---|
| H2-human | H2-226L/228G | H2-avian | |
| Data statistics | |||
| Space group | P63 | P63 | P63 |
| Unit cell parameters | |||
| a | 70.69 | 70.48 | 70.25 |
| b | 70.69 | 70.48 | 70.25 |
| c | 236.75 | 236.84 | 236.88 |
| Beamline | APS-23ID-B | ALS 5.0.3 | SSRL-11-1 |
| Mosaicity (o) | 0.30 | 0.49 | 0.43 |
| Reflection | 67,767 | 66,916 | 86,866 |
| Redundancy | 6.5 (5.9) | 5.6 (4.9) | 13.1 (7.0) |
| Completeness (%) | 97.7 (99.0) | 99.9 (99.6) | 100 (100) |
| Rmerge (%)b | 8.6 (53.4) | 7.1 (33.0) | 6.2 (50.4) |
| I/σI | 13.1 (3.1) | 16.4 (3.5) | 39.1 (3.7) |
| Refinement | |||
| Resolution (Å) | 40-1.73 | 45-1.75 | 30.2-1.60 |
| Reflection (test) | 64,327 (3,427) | 63,446 (3,385) | 82,347 (4,317) |
| Rcryst (%)c | 18.8 | 19.1 | 20.2 |
| Rfree (%)d | 22.2 | 22.2 | 23.2 |
| No. of atoms | 4656 | 4613 | 4623 |
| Avg B value | 26.1 | 29.9 | 14.0 |
| rmsd bonds (Å) | 0.013 | 0.014 | 0.012 |
| rmsd angles (o) | 1.44 | 1.47 | 1.44 |
| Ramachandran plot (% favored/% outliers)e | 97.6/0.0 | 96.7/0.2 | 97.2/0.2 |
| PDB code | 3KU5 | 3KU6 | 3KU3 |
Values in the parentheses correspond to the highest resolution shell.
Rmerge = Σ|I −<I> |/Σ |I|.
Rcryst = (Σ| |Fo| − |Fc| |)/Σ|Fo|.
Five percent of all reflections were randomly selected as test set.
Calculated using Molprobity (http://molprobity.biochem.duke.edu).
Protocols for microarray printing and recombinant HA analysis were as described previously (3, 34, 35). The imprinted slides used were obtained from the Consortium for Functional Glycomics (CFG) (glycan microarray version 3.2; see supplemental material for glycans used for analyses in these experiments). Briefly, for analysis using recombinant HA, HA-antibody complexes were prepared by mixing 15 μg of recombinant HA, mouse anti-penta-His Alexa Fluor 488 (Qiagen, Valencia, CA), and anti-mouse IgG Alexa Fluor 488 (Invitrogen, Carlsbad, CA) in a molar ratio of 4:2:1, respectively. These prepared mixtures of complexes were incubated for 15 min on ice, diluted to 1 ml with phosphate-buffered saline containing 3% (wt/vol) bovine serum albumin (PBS-BSA) and incubated on the microarray slide in a humidified chamber for 1 h. Slides were subsequently washed by successive rinses with PBS with 0.05% Tween 20 (PBS-T), PBS, and deionized water. To detect biotin-labeled grid markers, slides were subjected to a secondary incubation with 0.4 μg/ml streptavidin-Alexa Fluor 488 (Invitrogen, Carlsbad, CA), incubated and rinsed as described above, and then immediately subjected to imaging as described previously (3). Only results for glycans carrying 2,3- or 2,6-linked sialic acids (73 out of 406) are presented. The complete list of glycans and binding data for experiments described here can be found at http://www.functionalglycomics.org.
Protein structure accession number.
The atomic coordinates and structure factors for avian, human, and 226L/228G H2 HAs were deposited in the Protein Data Bank (PBD; http://www.rcsb.org) under identification (ID) codes 3KU3, 3KU5, and 3KU6, respectively.
RESULTS
H2 HA structure.
The ectodomain of the H2 HA gene from influenza virus A/Japan/305/57 was overexpressed in a baculovirus expression system as previously described (36). The soluble H2 HA trimer was cleaved into its mature form by trypsin digestion. Crystals for the three different HAs were obtained from similar crystallization conditions in the same crystal lattice. The structures were solved by molecular replacement, and the crystallographic data and statistics are summarized in Table 1.
H2 HA is a homomeric trimer (∼200 kDa) with each monomer containing two polypeptides (HA1, HA2) (Fig. 1b). Each HA trimer comprises a large, membrane-distal, globular domain that binds to receptors on host cells and an elongated membrane-proximal domain (stem region) dominated by intertwined and interconnecting α-helices. The membrane-distal globular domain is formed solely by HA1, whereas the stem region is comprised of HA2 and the N- and C-terminal segments of HA1. H2 HA has a structural fold highly similar to that of other HA subtypes. The closest structural homolog is the avian H5 structure of A/Duck/Singapore/3/1997 (Sing97; PDB ID, 1JSM) (12), as expected from sequence identity. The root-mean-square deviation (rmsd) between H2 and H5 Sing97 is only 0.87 Å for the HA monomer but is substantially greater (>2.8 Å) for the more distantly related H3 and H7 HAs, mainly due to different orientations of the membrane-distal domain in relation to the stem region, consistent with previously described clade-specific traits (28) (see Table S1 in the supplemental material). The receptor-binding site is located near the top of the HA1 globular head in a shallow cavity surrounded by residues from four secondary structure elements: the 190 helix (residues 190 to 198), 220 loop (221 to 228), 130 loop (134 to 138), and Thr155 (Fig. 1c). Several conserved aromatic residues, including Tyr98, Trp153, and His183, form the bottom of the depression. Residues surrounding the binding site tend to be more variable across different HA subtypes than those at the base. Mutations in these peripheral residues often shift or alter receptor-binding specificity. In H1, H2, and H3, as few as two mutations in the receptor-binding site, are sufficient to switch specificity from avian to human receptors (6, 21). In H2 and H3, Q226L and G228S mutations correlate with this switch in specificity (6, 21), whereas two different mutations, E190D and G225D, interconvert the specificity of 1918 H1 HA from avian α2,3- to human α2,6-linked sugars and vice versa (21, 35).
Receptor-binding specificities of H2 HAs.
In the 1957 pandemic, early viral isolates from humans could be assigned to three subpopulations: avian-like viruses containing 226Q and 228G, classic human viruses with 226L and 228S, and atypical 226L/228G mutants (Fig. 2a) (6, 21). In subsequent years, all H2 viruses acquired the human-specific 226L and 228S combination. To understand the basis for the binding specificity shift, three H2 HA constructs of A/Japan/305/57 were studied by glycan microarray analysis (3, 35). The 226Q/228G construct (hereafter referred to as H2-avian) showed a clear preference for α2,3-linked sialosides, with negligible binding to α2,6-linked glycans (Fig. 2b). H2-avian recognizes most glycans that contain Neu5Acα2-3Gal except for α2,3-linked glycans replaced by sulfate (glycans 2, 3, and 4) or fucose (glycans 31 to 40), or glycans with sialic acid attached to an internal (branched) galactose (glycans 41 to 46) (see supplemental material for list of glycans). The 226L/228S construct (hereafter referred to as H2-human) displayed markedly reduced binding to α2,3-linked glycans and preferential binding to α2,6-linked sialosides (Fig. 2d). In general, both biantennary and linear α2,6-linked glycans bind well to H2-human HA. Several α2,3-linked sialosides also bind, including some fucosylated sialosides (glycans 33 and 35) that show significant binding to H2-human but not to H2-avian for reasons not currently understood. The 226L/228G single mutant (hereafter referred to as H2-226L/228G) exhibited weak binding to both α2,3- and α2,6-linked glycans (Fig. 2c). Thus, the precise combination of residues at positions 226 and 228 dictates H2 glycan-binding preferences. Similar receptor-binding properties in other H2 viral isolates have also been observed by others (21).
FIG. 2.

Switch of binding specificity in H2 HA. (a) Amino acids at positions 226 and 228 in selected H2 HAs of avian, human, and swine origins. (b to d) Residues at positions 226 and 228 dictate binding specificity in H2. Sialosides are grouped into α2,3 (residues 1 to 46)- and α2,6 (residues 47 to 73)-linked glycans for interrogation on the array. In each graph, binding to α2,3- and α2,6-linked glycans are shown as green and red bars, respectively. Glycans are also color-coded in subgroups along the x axis and in the glycan list in the supplemental material (orange, α2,3 sulfated sialosides; blue, α2,3 branched disialosides; green, α2,3 linear sialosides; red, α2,3 fucosylated sialosides; light blue, α2,3 internal sialosides; yellow, α2,6 sulfated sialosides; purple, α2,6 branched di-sialosides; magenta, α2,6 linear sialosides; brown, α2,6 internal sialosides). H2-human HA (d) reveals classic α2,6 specificity, while H2-avian HA (b) binds only α2,3-linked glycans. A single mutant H2-226L/228G (H2-LG) (c) shows relatively low binding to both α2,3 and α2,6 sialosides.
Structural basis of binding specificity.
Comparison of the H2-avian and H2-human structures reveals that the 226 and 228 substitutions slightly elongate the receptor-binding site in H2-human HA (Fig. 3). Residue 226 Cα shifts 0.5 Å between the two structures. In addition, neighboring residues also move slightly in response to changes at position 226. Most notably, Tyr98 shifts slightly away from the 220 loop. The separation between Tyr98 and the 220 loop resulted from a hydrophobic stacking of Leu226 side chain on the aromatic ring of Tyr98, along with van der Waals contacts between Ser228 and the hydroxyl group of Tyr98. The shift of the 220 loop is maximal when both Q226L and G228S substitutions are present, as the single mutant H2-226L/228G is intermediate, between H2-avian and H2-human. Similar structural differences were also observed between H3-avian and H3-human structures (13). The rationale behind H3 receptor specificity has been well documented through structural studies of H3 in complexes with receptor analogs (7, 13, 29). Aside from changes in hydrophobicity and size at residue 226, affinity differences result from variation in shape complementarity of the various HA binding sites with the host glycan receptors. Sialic acids with α2,3- or α2,6-linkage are bound differently in HA crystal complexes (7, 11). Avian-like α2,3 glycans adopt a trans conformation of the sialoside linkage, whereas human-like α2,6 glycans are cis (7). In the H3 complexes, the hydrophilic Q226 in avian HA interacts with the galactose glycosidic oxygen (Gal-O3) of α2,3-linked glycans (13), whereas mutation to Leu226 in human HA positions the aliphatic side chain beneath the galactose C6 methylene group (Gal-C6) of α2,6-linked glycans (7). Selective binding, therefore, results from the hydrophilic/hydrophobicity switch and the differential geometric constraints at the contact surface. These clear, but subtle, differences in receptor binding correlate with small differences in intrinsic affinity for the receptor glycans. Direct nuclear magnetic resonance (NMR) affinity measurements of the human virus H3 HA for α2,6- and α2,3-linked glycans revealed Kd (dissociation constant) values of 2.1 and 3.2 mM, respectively, while an avian variant with Gln at position 226 exhibited corresponding values of 5.9 and 2.9 mM (30). Such small differences in absolute affinity emphasize the importance of multivalency for increasing avidity that translates into biologically distinct binding properties for recognition of α2,6- and α2,3-linked glycans, as exemplified by the glycan array analysis results in Fig. 2. Our structures suggest that H2 and H3 viruses both successfully exploit these differences in avian/human glycan linkages via the same mechanism. Adaptation to humans and birds is highly efficient since only two mutations are required to switch the binding specificity for both H2 and H3 subtypes.
FIG. 3.

Structural comparison of H2 mutants in a stereo representation. H2-avian (orange), H2-human (cyan), and H2-226L/228G (gray). The binding pocket is extended by about 0.5 Å by mutations Q226L/G228S from H2-avian to H2-human. The 220 loop is shown in Cα trace for clarity.
Antigenic structure of H2 HA.
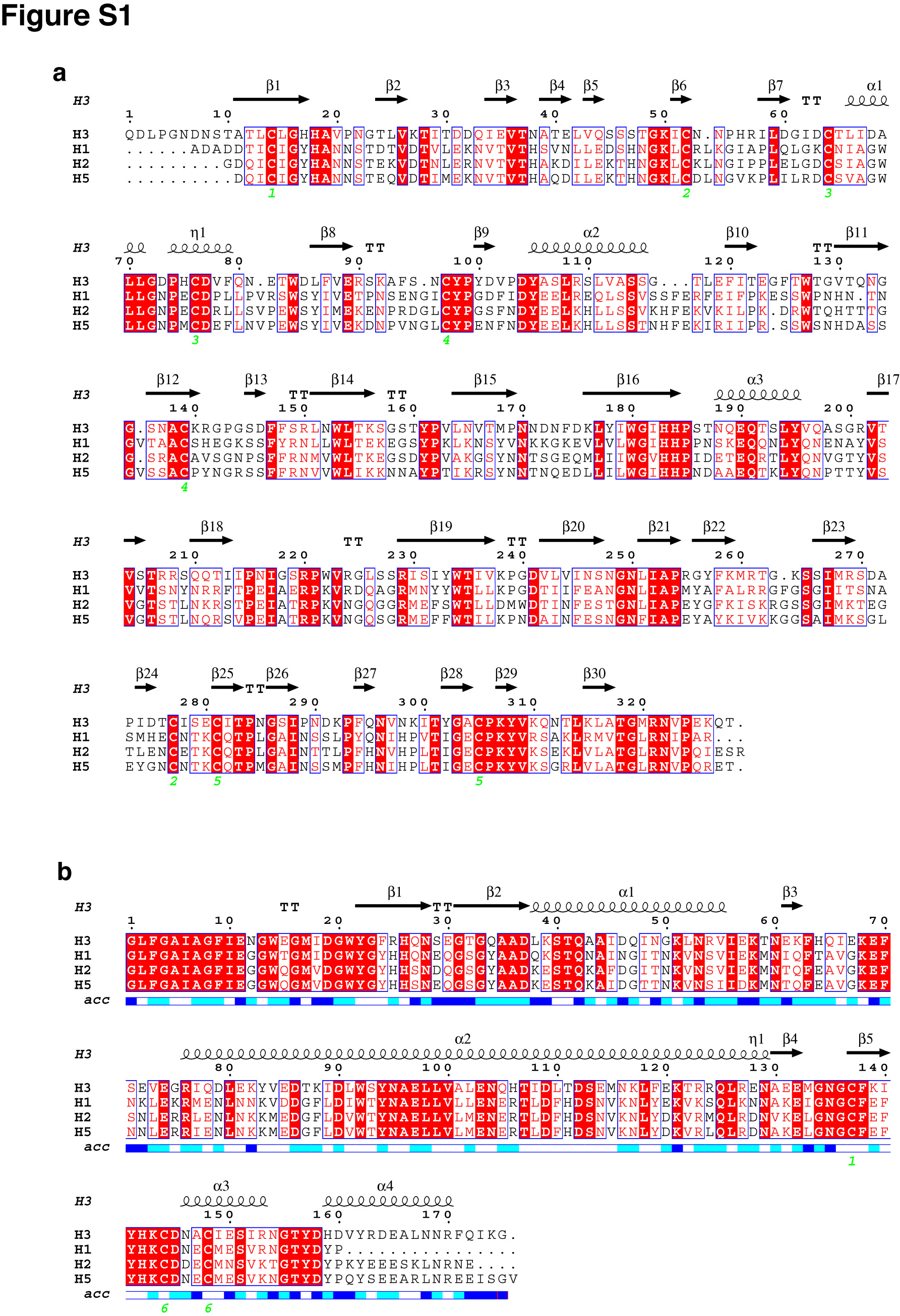
The antigenic structure of H2 HA was previously studied by the analysis of escape mutants from monoclonal antibodies (40). Most escape mutants cluster in the membrane-distal domain, coincident with the antigenic sites in H3 (Fig. 4a and b). However, one escape mutant revealed a unique antigenic site in the stem region by the mutation of Lys40 in HA2 (40). Independently, Okuno et al. identified a monoclonal antibody, C179, from mice immunized with an H2N2 strain (A/Okuda/57) (24) that targets a common neutralizing epitope in the stem region shared by multiple influenza A subtypes (24, 32). The epitope includes part of helix 1 (residues 47 to 58) in HA2 that is highly conserved in the clades of H1, H2, and H5 (Fig. 4c) and part of HA1 in the stem region (24, 32). Escape mutants were mapped to Thr318 of HA1 or Val52 of HA2. Together with Lys40, they revealed a conserved cavity in the stem region of H2 HA (Fig. 4d). An overlay of different HA subtypes shows nearly identical secondary structures with minor variations in side chains across subtypes, suggesting an ideal epitope for broad neutralization. Antigenic epitopes of various HA subtypes have been well studied, and most epitopes are located in the membrane-distal globular domain (5, 16, 17, 40, 42). Those variable and exposed epitopes in the membrane-distal domain can be readily mutated without functional consequences. Compared with the membrane-distal domain, the stem region of HA is highly conserved in the primary sequences and is the key region for mediating membrane fusion. Thus, influenza viruses with the ability to shift host antigenic attention to the membrane-distal domain likely are at an advantage evolutionarily. The so-called “deceptive imprinting” allows many pathogens, including human immunodeficiency virus type 1 (HIV-1) and flu virus, to protect conserved and functionally critical regions from host immune attacks (39). Influenza virus HA shields its conserved, membrane-proximal stem region, likely either through crowding of the flu virus HA spikes on the viral surface or through shielding by glycosylation. Stem regions of HAs are usually heavily glycosylated, and four out of the five N glycosylation sites in H2 HA are located in this region (Fig. 1a). The relative inability of H2 to continue to evade the human immune system, as suggested by the stem region antibodies, may have resulted in its eventual elimination from the human population.
FIG. 4.

Antigenic structure of H2. Positions of escape mutants are mapped on one monomer, in side view (a) and top view (b). Mutations that add a glycosylation site are colored in green, whereas other single residue mutations are colored in blue. A mutation on the stem region (Lys40; shown in red) reveals a highly conserved epitope for antibody binding. (c) Sequence alignment of the conserved epitope within the stem domain (HA2, residues 38 to 57) from the four structures is shown. Residues conserved in all four structures are highlighted in red, with residues conserved only in the H2 clade colored in green. (d) Comparison of the conserved epitope from various HA subtypes. Helix 1 from H1 (1ru7) (gray), H3 (2hmg) (blue), and H5 (1jsm) (yellow) are superimposed on the same segment of H2 (green). Positions of escape mutants in this region, identified in two independent studies, are shown in sticks and labeled in red.
DISCUSSION
Our structural and functional analyses suggest that H2 viruses of nonhuman origin readily adapted to human receptor specificity with only two mutations during the 1957 influenza pandemic. The conversion of binding specificity relies on precise interactions between HA and glycan receptors. A slight elongation of the binding pocket between H2-avian and H2-human, along with the hydrophobicity switch at residue 226, dictates drastically different glycan binding profiles. The seemingly stringent requirement for binding site length is suggested by the unique binding properties of the intermediate mutant H2-226L/228G. The intermediate mutant carries a leucine residue at position 226, the same as H2-human, yet still lacks binding affinities for α2,6-linked glycans. Preferential binding, thus, is achieved by exact positioning of interacting residues.
These structural comparisons elucidate the structural impact of the receptor-altering mutations Q226L/G228S at very high resolution. These three H2 structures were generated from protein with the same genetic background, ensuring that any structural and functional effects observed are strictly a result of the two key residues. Furthermore, the proteins were crystallized under the same conditions and within the same crystal lattice. That, along with the highest resolutions among known influenza virus HA structures, allows us to interpret these subtle structural differences that have such a dramatic impact on HA receptor binding.
The recent emergence of the 2009 H1N1 influenza pandemic highlights the social impacts of human pandemics and calls for improved surveillance of zoonotic influenza virus strains with pandemic potential. It has been 40 years since the H2 pandemic, ensuring the continual growth of a largely naïve human population for initiating and sustaining a new H2 pandemic (41). Avian flu viruses possessing H2 HAs closely related to human H2 viruses are currently circulating in wild birds (20). Recently isolated H2N3 strains from swine in the United States carry Leu226, instead of the avian-like Gln226, with increased preference for α2,6-linked receptors (19). Given the likelihood of pigs acting as potential intermediate hosts (33) and the relative ease for these viruses to transfer into human population, the current circulating H2 viruses warrant close attention and should be considered a high-risk subtype for future pandemics.
Meanwhile, the conserved antigenic site, originally identified on the stem region of H2, could have inadvertently revealed the Achilles' heel for influenza viruses. Currently, the most effective way to combat flu is to use vaccines that elicit a neutralizing antibody response against the HA. Unfortunately, the variable epitopes on the receptor binding domain are almost always targeted by the immune system and, thus, and only neutralize the immunizing virus or very closely related strains (14). Recently, exciting new studies have identified antibodies with broad neutralizing activity against H1 and H5 (15, 37, 38). Crystal structures of antibody-HA complexes reveal that these antibodies recognize a conserved epitope in the stem region (8, 37) that likely overlaps the H2 stem epitope. Notwithstanding, it still remains a significant obstacle to focus the host immune response on such conserved epitopes in order to take advantage of this perceived weakness in H2, as well as in H1 and H5, viruses to design a better and more universal flu vaccine.
Supplementary Material
Acknowledgments
The work was supported in part by NIAID grant AI058113 (I.A.W.), the Skaggs Institute for Chemical Biology, and the Scripps DNA-Protein Microarray Core Facility. Glycan microarrays were provided by the Consortium for Functional Glycomics, funded by NIGMS grant GM62116 (J.C.P.).
X-ray diffraction data sets were collected at the Stanford Synchrotron Radiation Lightsource beamline 11-1, the Advanced Photon Source beamline 23ID-B, and the Advanced Light Source beamline 5.0.3, as listed in Table 1. We thank X. Dai (The Scripps Research Institute) for expert technical assistance and D. Ekiert and X. Zhu (The Scripps Research Institute) for helpful discussions.
This is publication 19951 from The Scripps Research Institute.
Footnotes
Published ahead of print on 9 December 2009.
Supplemental material for this article may be found at http://jvi.asm.org/.
REFERENCES
- 1.Bao, Y., P. Bolotov, D. Dernovoy, B. Kiryutin, L. Zaslavsky, T. Tatusova, J. Ostell, and D. Lipman. 2008. The influenza virus resource at the National Center for Biotechnology Information. J. Virol. 82:596-601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Beigel, J. H., J. Farrar, A. M. Han, F. G. Hayden, R. Hyer, M. D. de Jong, S. Lochindarat, T. K. Nguyen, T. H. Nguyen, T. H. Tran, A. Nicoll, S. Touch, and K. Y. Yuen. 2005. Avian influenza A (H5N1) infection in humans. N. Engl. J. Med. 353:1374-1385. [DOI] [PubMed] [Google Scholar]
- 3.Blixt, O., S. Head, T. Mondala, C. Scanlan, M. E. Huflejt, R. Alvarez, M. C. Bryan, F. Fazio, D. Calarese, J. Stevens, N. Razi, D. J. Stevens, J. J. Skehel, I. van Die, D. R. Burton, I. A. Wilson, R. Cummings, N. Bovin, C. H. Wong, and J. C. Paulson. 2004. Printed covalent glycan array for ligand profiling of diverse glycan binding proteins. Proc. Natl. Acad. Sci. U. S. A. 101:17033-17038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Butt, K. M., G. J. Smith, H. Chen, L. J. Zhang, Y. H. Leung, K. M. Xu, W. Lim, R. G. Webster, K. Y. Yuen, J. S. Peiris, and Y. Guan. 2005. Human infection with an avian H9N2 influenza A virus in Hong Kong in 2003. J. Clin. Microbiol. 43:5760-5767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Caton, A., G. Brownlee, J. Yewdell, and W. Gerhard. 1982. The antigenic structure of the influenza virus A/PR/8/34 hemagglutinin (H1 subtype). Cell 31:417-427. [DOI] [PubMed] [Google Scholar]
- 6.Connor, R., Y. Kawaoka, R. Webster, and J. Paulson. 1994. Receptor specificity in human, avian, and equine H2 and H3 influenza virus isolates. Virology 205:17-23. [DOI] [PubMed] [Google Scholar]
- 7.Eisen, M., S. Sabesan, J. Skehel, and D. Wiley. 1997. Binding of the influenza A virus to cell-surface receptors: structures of five hemagglutinin-sialyloligosaccharide complexes determined by X-ray crystallography. Virology 232:19-31. [DOI] [PubMed] [Google Scholar]
- 8.Ekiert, D. C., G. Bhabha, M. A. Elsliger, R. H. Friesen, M. Jongeneelen, M. Throsby, J. Goudsmit, and I. A. Wilson. 2009. Antibody recognition of a highly conserved influenza virus epitope. Science 324:246-251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Emsley, P., and K. Cowtan. 2004. Coot: model-building tools for molecular graphics. Acta Crystallogr. D 60:2126-2132. [DOI] [PubMed] [Google Scholar]
- 10.Fouchier, R. A., P. M. Schneeberger, F. W. Rozendaal, J. M. Broekman, S. A. Kemink, V. Munster, T. Kuiken, G. F. Rimmelzwaan, M. Schutten, G. J. Van Doornum, G. Koch, A. Bosman, M. Koopmans, and A. D. Osterhaus. 2004. Avian influenza A virus (H7N7) associated with human conjunctivitis and a fatal case of acute respiratory distress syndrome. Proc. Natl. Acad. Sci. U. S. A. 101:1356-1361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gamblin, S., L. Haire, R. Russell, D. Stevens, B. Xiao, Y. Ha, N. Vasisht, D. Steinhauer, R. Daniels, A. Elliot, D. Wiley, and J. Skehel. 2004. The structure and receptor binding properties of the 1918 influenza hemagglutinin. Science 303:1838-1842. [DOI] [PubMed] [Google Scholar]
- 12.Ha, Y., D. Stevens, J. Skehel, and D. Wiley. 2002. H5 avian and H9 swine influenza virus haemagglutinin structures: possible origin of influenza subtypes. EMBO J. 21:865-875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ha, Y., D. Stevens, J. Skehel, and D. Wiley. 2003. X-ray structure of the hemagglutinin of a potential H3 avian progenitor of the 1968 Hong Kong pandemic influenza virus. Virology 309:209-218. [DOI] [PubMed] [Google Scholar]
- 14.Johansson, B., and I. Brett. 2007. Changing perspective on immunization against influenza. Vaccine 25:3062-3065. [DOI] [PubMed] [Google Scholar]
- 15.Kashyap, A. K., J. Steel, A. F. Oner, M. A. Dillon, R. E. Swale, K. M. Wall, K. J. Perry, A. Faynboym, M. Ilhan, M. Horowitz, L. Horowitz, P. Palese, R. R. Bhatt, and R. A. Lerner. 2008. Combinatorial antibody libraries from survivors of the Turkish H5N1 avian influenza outbreak reveal virus neutralization strategies. Proc. Natl. Acad. Sci. U. S. A. 105:5986-5991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kaverin, N., I. Rudneva, N. Ilyushina, A. Lipatov, S. Krauss, and R. Webster. 2004. Structural differences among hemagglutinins of influenza A virus subtypes are reflected in their antigenic architecture: analysis of H9 escape mutants. J. Virol. 78:240-249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kaverin, N., I. Rudneva, N. Ilyushina, N. Varich, A. Lipatov, Y. Smirnov, E. Govorkova, A. Gitelman, D. Lvov, and R. Webster. 2002. Structure of antigenic sites on the haemagglutinin molecule of H5 avian influenza virus and phenotypic variation of escape mutants. J. Gen. Virol. 83:2497-2505. [DOI] [PubMed] [Google Scholar]
- 18.Lamzin, V., A. Perrakis, and K. Wilson. 2001. The ARP/WARP suite for automated construction and refinement of protein models. In M. G. Rossmann and E. Arnold (ed.), International tables for crystallography, vol. F. Crystallography of biological macromolecules. Kluwer Academic Publishers, Dordrecht, The Netherlands.
- 19.Ma, W., A. Vincent, M. Gramer, C. Brockwell, K. Lager, B. Janke, P. Gauger, D. Patnayak, R. Webby, and J. Richt. 2007. Identification of H2N3 influenza A viruses from swine in the United States. Proc. Natl. Acad. Sci. U. S. A. 104:20949-20954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Makarova, N., N. Kaverin, S. Krauss, D. Senne, and R. Webster. 1999. Transmission of Eurasian avian H2 influenza virus to shorebirds in North America. J. Gen. Virol. 80:3167-3171. [DOI] [PubMed] [Google Scholar]
- 21.Matrosovich, M., A. Tuzikov, N. Bovin, A. Gambaryan, A. Klimov, M. Castrucci, I. Donatelli, and Y. Kawaoka. 2000. Early alterations of the receptor-binding properties of H1, H2, and H3 avian influenza virus hemagglutinins after their introduction into mammals. J. Virol. 74:8502-8512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.McCoy, A. J., R. W. Grosse-Kunstleve, L. C. Storoni, and R. J. Read. 2005. Likelihood-enhanced fast translation functions. Acta Crystallogr. D 61:458-464. [DOI] [PubMed] [Google Scholar]
- 23.Murshudov, G. N., A. A. Vagin, and E. J. Dodson. 1997. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. D 53:240-255. [DOI] [PubMed] [Google Scholar]
- 24.Okuno, Y., Y. Isegawa, F. Sasao, and S. Ueda. 1993. A common neutralizing epitope conserved between the hemagglutinins of influenza A virus H1 and H2 strains. J. Virol. 67:2552-2558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Otwinowski, Z., and W. Minor. 1997. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 276:307-326. [DOI] [PubMed] [Google Scholar]
- 26.Parrish, C. R., and Y. Kawaoka. 2005. The origins of new pandemic viruses: the acquisition of new host ranges by canine parvovirus and influenza A viruses. Annu. Rev. Microbiol. 59:553-586. [DOI] [PubMed] [Google Scholar]
- 27.Potter, C. 2001. A history of influenza. J. Appl. Microbiol. 91:572-579. [DOI] [PubMed] [Google Scholar]
- 28.Russell, R., S. Gamblin, L. Haire, D. Stevens, B. Xiao, Y. Ha, and J. Skehel. 2004. H1 and H7 influenza haemagglutinin structures extend a structural classification of haemagglutinin subtypes. Virology 325:287-296. [DOI] [PubMed] [Google Scholar]
- 29.Sauter, N., J. Hanson, G. Glick, J. Brown, R. Crowther, S. Park, J. Skehel, and D. Wiley. 1992. Binding of influenza virus hemagglutinin to analogs of its cell-surface receptor, sialic acid: analysis by proton nuclear magnetic resonance spectroscopy and X-ray crystallography. Biochemistry 31:9609-9621. [DOI] [PubMed] [Google Scholar]
- 30.Sauter, N. K., M. D. Bednarski, B. A. Wurzburg, J. E. Hanson, G. M. Whitesides, J. J. Skehel, and D. C. Wiley. 1989. Hemagglutinins from two influenza virus variants bind to sialic acid derivatives with millimolar dissociation constants: a 500-MHz proton nuclear magnetic resonance study. Biochemistry 28:8388-8396. [DOI] [PubMed] [Google Scholar]
- 31.Skehel, J., and D. Wiley. 2000. Receptor binding and membrane fusion in virus entry: the influenza hemagglutinin. Annu. Rev. Biochem. 69:531-569. [DOI] [PubMed] [Google Scholar]
- 32.Smirnov, Y., A. Lipatov, A. Gitelman, Y. Okuno, R. Van Beek, A. Osterhaus, and E. Claas. 1999. An epitope shared by the hemagglutinins of H1, H2, H5, and H6 subtypes of influenza A virus. Acta Virol. 43:237-244. [PubMed] [Google Scholar]
- 33.Smith, G. J., J. Bahl, D. Vijaykrishna, J. Zhang, L. L. Poon, H. Chen, R. G. Webster, J. S. Peiris, and Y. Guan. 2009. Dating the emergence of pandemic influenza viruses. Proc. Natl. Acad. Sci. U. S. A. 106:11709-11712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Stevens, J., O. Blixt, L. Glaser, J. Taubenberger, P. Palese, J. Paulson, and I. Wilson. 2006. Glycan microarray analysis of the hemagglutinins from modern and pandemic influenza viruses reveals different receptor specificities. J. Mol. Biol. 355:1143-1155. [DOI] [PubMed] [Google Scholar]
- 35.Stevens, J., O. Blixt, T. Tumpey, J. Taubenberger, J. Paulson, and I. Wilson. 2006. Structure and receptor specificity of the hemagglutinin from an H5N1 influenza virus. Science 312:404-410. [DOI] [PubMed] [Google Scholar]
- 36.Stevens, J., A. Corper, C. Basler, J. Taubenberger, P. Palese, and I. Wilson. 2004. Structure of the uncleaved human H1 hemagglutinin from the extinct 1918 influenza virus. Science 303:1866-1870. [DOI] [PubMed] [Google Scholar]
- 37.Sui, J., W. C. Hwang, S. Perez, G. Wei, D. Aird, L. M. Chen, E. Santelli, B. Stec, G. Cadwell, M. Ali, H. Wan, A. Murakami, A. Yammanuru, T. Han, N. J. Cox, L. A. Bankston, R. O. Donis, R. C. Liddington, and W. A. Marasco. 2009. Structural and functional bases for broad-spectrum neutralization of avian and human influenza A viruses. Nat. Struct. Mol. Biol. 16:265-273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Throsby, M., E. van den Brink, M. Jongeneelen, L. L. Poon, P. Alard, L. Cornelissen, A. Bakker, F. Cox, E. van Deventer, Y. Guan, J. Cinatl, J. ter Meulen, I. Lasters, R. Carsetti, M. Peiris, J. de Kruif, and J. Goudsmit. 2008. Heterosubtypic neutralizing monoclonal antibodies cross-protective against H5N1 and H1N1 recovered from human IgM+ memory B cells. PLoS One 3:e3942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Tobin, G., J. Trujillo, R. Bushnell, G. Lin, A. Chaudhuri, J. Long, J. Barrera, L. Pena, M. Grubman, and P. Nara. 2008. Deceptive imprinting and immune refocusing in vaccine design. Vaccine 26:6189-6199. [DOI] [PubMed] [Google Scholar]
- 40.Tsuchiya, E., K. Sugawara, S. Hongo, Y. Matsuzaki, Y. Muraki, Z. Li, and K. Nakamura. 2001. Antigenic structure of the haemagglutinin of human influenza A/H2N2 virus. J. Gen. Virol. 82:2475-2484. [DOI] [PubMed] [Google Scholar]
- 41.Webster, R. 1997. Predictions for future human influenza pandemics. J. Infect. Dis. 176(Suppl. 1):S14-S19. [DOI] [PubMed] [Google Scholar]
- 42.Wiley, D., I. Wilson, and J. Skehel. 1981. Structural identification of the antibody-binding sites of Hong Kong influenza haemagglutinin and their involvement in antigenic variation. Nature 289:373-378. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}