

Abstract
Diffusion tensor imaging (DTI) of the brain has become a mainstay in the study of normal aging of white matter, and only recently has attention turned to the use of DTI to examine aging effects in gray matter structures. Of the many changes in the brain that occur with advancing age is increased presence of iron, notable in selective deep gray matter structures. In vivo detection and measurement of iron deposition is possible with magnetic resonance imaging (MRI) because of iron's effect on signal intensity. In the process of a DTI study, a series of diffusion-weighted images (DWI) is collected, and while not normally considered as a major dependent variable in research studies, they are used clinically and they reveal striking conspicuity of the globus pallidus and putamen caused by signal loss in these structures, presumably due to iron accumulation with age. These iron deposits may in turn influence DTI metrics, especially of deep gray matter structures. The combined imaging modality approach has not been previously used in the study of normal aging. The present study used legacy DTI data collected in 10 younger (22–37 years) and 10 older (65–79 years) men and women at 3.0 T and fast spin-echo (FSE) data collected at 1.5 T and 3.0 T to derive an estimate of the field-dependent relaxation rate increase (the “FDRI estimate”) in the putamen, caudate nucleus, globus pallidus, thalamus, and a frontal white matter sample comparison region. The effect of age on the diffusion measures in the deep gray matter structures was distinctly different from that reported in white matter. In contrast to lower anisotropy and higher diffusivity typical in white matter of older relative to younger adults observed with DTI, both anisotropy and diffusivity were higher in the older than younger group in the caudate nucleus and putamen; the thalamus showed little effect of age on anisotropy or diffusivity. Signal intensity measured with DWI was lower in the putamen of elderly than young adults, whereas the opposite was observed for the white matter region and thalamus. As a retrospective study based on legacy data, the FDRI estimates were based on FSE sequences, which underestimated the classical FDRI index of brain iron. Nonetheless, the differential effects of age on DTI metrics in subcortical gray matter structures compared with white matter tracts appears to be related, at least in part, to local iron content, which in the elderly of the present study was prominent in the FDRI estimate of the putamen and visibly striking in the diffusion-weighted image of the basal ganglia structures.
Keywords: Brain, Aging, Iron, DTI, DWI, MRI, Diffusion
1. Introduction
The burgeoning use of magnetic resonance (MR) diffusion tensor imaging (DTI) in the study of normal aging has expanded the availability of diffusion-weighted images (DWI) of the brain. DWI, while not normally considered as a major dependent variable in research studies, are used clinically as a highly sensitive marker for acute ischemia (Moseley et al., 1990b; Moustafa and Baron, 2007; Srinivasan et al., 2006) and for the evaluation of stroke (Wang et al., 2006). Inspection of DWI, especially in advancing age, reveals a striking conspicuity of the globus pallidus and putamen caused by signal loss in these structures, presumably due to iron accumulation with age. These iron deposits may influence DTI metrics, especially of deep gray matter structures. Here, we present data on young and elderly healthy individuals who have had both brain DTI and MR-based brain iron assessment and examine the influence of brain iron on DTI.
Deep gray matter structures have a unique MR signal intensity in the elderly, with profound signal darkening on T2* and T2-weighted images in some regions, especially the putamen (Hiwatashi et al., 2003; Thomas et al., 1993). There is ample evidence indicating that these local age effects are due to age-related iron deposition (in vivo: 7,18,postmortem: Hallgren and Sourander, 1958), which may be a marker of occult vascular disease and a correlate of age-related decline in cognitive or motor functions associated with the iron-laden subcortical structures (Pujol et al., 1992). Iron effects on MR images are apparent even in young adults, for example, in substantia nigra, globus pallidus, and red nucleus, and consequently show little change with advancing age beyond adolescence (Pujol et al., 1992; Thomas et al., 1993). The thalamus and white matter contain little to no iron (Hallgren and Sourander, 1958) and therefore exhibit no T2* effect from iron (Bartzokis et al., 1993). Other structures, including the putamen and caudate, however, are vulnerable to further accumulation of iron with advancing age (Bartzokis et al., 1994, 2007b), and the iron accumulation is abnormally high in diseases of the basal ganglia, such as Parkinson's disease (Bartzokis et al., 1999; Michaeli et al., 2007; Pujol et al., 1992), striatonigral degeneration (Martin et al., 1998), and Huntington's disease (Bartzokis et al., 2007a).
Iron deposits in the brain cause loss of signal intensity with certain image acquisition types, and the effect is greater as the field strength increases (e.g., from 0.5 T to 1.5 T to 3.0 T). The signal loss effect is due to the fact that the iron deposits behave as little magnets when subjected to a strong magnetic field and cause protons in nearby water molecules to lose their coherence, and therefore magnetic resonance imaging (MRI) signal intensity, more quickly than they would if not in the vicinity of iron. This signal loss is usually referred to as an effect on transverse relaxation time, T2, or the corresponding rate, R2 = 1/T2, that characterize the time during which the proton spins interact with each other (spin–spin interactions) and the signal in the transverse plane is lost. Another source of signal loss is identified by a time constant T2*, or the corresponding rate R2* = 1/T2*, and characterizes the time during which the spinning protons lose their signal in the transverse plane because of local magnetic field inhomogeneity, e.g., caused by the iron molecules. The R2* signal loss can be recovered with certain imaging protocols, i.e., spin-echo sequences, whereas the R2 signal loss cannot be recovered.
Several methods have been proposed for detecting and quantifying iron in the brain non-invasively with MR-based techniques (reviewed by Haacke et al. (2005)). Among them are susceptibility-weighted imaging (Haacke et al., 2007), where the effect of magnetic susceptibility on signal phase accrual is used to estimate local iron concentrations. The difference in R2 and R2* has been proposed for iron measurements (Gelman and Guinto, 1992; Ma and Wehrli, 1996; Ordidge et al., 1994). Alternatively, iron measurement has been proposed based on a parametric model of signal decay in a series of measurements with incrementally asymmetric spin-echoes, in a procedure called magnetic field correlation imaging (Jensen et al., 2006).
The method used in the current work relies on acquiring MRI of each subject at two different field strengths to take advantage of iron's differential effect on transverse relaxation rates (R2) at different field strengths (Bizzi et al., 1992; Gomori and Grossman, 1993; Majumdar et al., 1989). This method has been calibrated for quantifying iron content, and relies on relatively simple processing of R2 measurements at the two field strengths. As described by Bartzokis et al. (1993, 1994, 2007b), the mainstay of brain iron is in ferritin molecules, which increase transverse relaxation rates (R2) linearly with increases in MR field strength (but see Bizzi et al., 1990), and can be quantified as the field-dependent R2 increase (FDRI) per unit Tesla, where higher FDRI indexes greater local concentration of iron. The FDRI measured regionally in vivo (Bartzokis et al., 2007b) correlated nearly perfectly with iron concentration measurements made postmortem (Hallgren and Sourander, 1958). A series of cross-sectional studies on regional differences in ferritin-bound iron measured with FDRI revealed significantly greater FDRI notable in caudate nucleus and putamen in older than younger healthy individuals, less of an FDRI-age effect in globus pallidus and hippocampus, and none in the thalamus. In contrast to accruing iron in deep gray matter structures with age, frontal white matter showed an age-related decrease in FDRI (Bartzokis et al., 2007b).
The local inhomogeneity caused by iron particles increases transverse relaxation rates contributing to signal darkening on conventional spin-echo MR images, notable in brain regions exhibiting aging effects (Raz et al., 2007). Co-occurring with advancing age, however, is a generalized decrease in transverse relaxation rates across the brain, which results in signal brightening, probably due to interstitial fluid accumulation. These two contributors to the relaxation processes compete, making the source of age-related R2-dependent MRI signal differences difficult to discern.
Additional information about the source of tissue quality differences with aging may be present in diffusion tensor imaging (DTI) and its metrics. DTI is sensitive to the intrinsic properties of water diffusion in tissue and provides information about tissue microstructural characteristics, based on two types of quantitative data: (1) diffusivity, which increases with extracellular fluid accumulation, decreases with gliosis, and can be expressed as mean diffusivity (MD); and (2) coherence of highly structured tissue (e.g., white matter bundles), which provides information about microstructural integrity (e.g., myelin) and can be expressed as fractional anisotropy (FA) (Basser and Pierpaoli, 1996; Le Bihan, 2003; Moseley et al., 1990a; Pfefferbaum and Sullivan, 2005a; Pierpaoli and Basser, 1996). DTI metrics have identified significant effects of adult aging that are now well described in regional white matter (for review, Sullivan and Pfefferbaum, 2006). Typically, with advancing age anisotropy declines and diffusivity increases more in anterior than posterior brain regions, whether measured with a region-of-interest approach (Head et al., 2004; Madden et al., 2004; O'Sullivan et al., 2001; Pfefferbaum et al., 2005; Pfefferbaum et al., 2006; Pfefferbaum and Sullivan, 2003; Pfefferbaum et al., 2000; Salat et al., 2005; Sullivan et al., 2001), voxel-based analysis (Grieve et al., 2007; Kochunov et al., 2007; Snook et al., 2007), or quantitative fiber tracking (Pfefferbaum et al., 2007; Sullivan et al., 2006, 2008). As observed postmortem, a predilection of age-related loss occurs for thin, unmyelinated fibers, which are in greatest abundance in the frontal lobes (Aboitiz et al., 1996; Bartzokis, 2004). Commonly but not necessarily, diffusivity negatively correlates with anisotropy within white matter samples (Chen et al., 2001; Engelter et al., 2000; Head et al., 2004; Helenius et al., 2002; Naganawa et al., 2003; Pfefferbaum et al., 2005; Pfefferbaum and Sullivan, 2005b, 2003), probably reflecting an age-related increase in unbound interstitial fluid (e.g., Norris et al., 1994; Peters and Sethares, 2003; Pfefferbaum and Sullivan, 2005b; Rumpel et al., 1998; Sehy et al., 2002; Silva et al., 2002). Signal loss attributable to iron deposition is especially conspicuous on diffusion-weighted spin-echo images (Hiwatashi et al., 2003). Although greater presence of free water should result in increased MD and reduced R2 (c.f., Pfefferbaum et al., 1999), these fluid-related age effects on MD may be more reflective of spin–spin R2 changes than are R2 estimates from multi-echo spin-echo MRI data.
Despite DTI's role in revealing age-related changes in quality of brain white matter and suggesting mechanisms of degradation, the utility of DTI metrics in furthering our understanding of age-related changes in gray matter is relatively unexplored, with a few exceptions (Huang et al., 2006; Kochunov et al., 2007; Liu et al., 2007). Theoretically, DTI could provide quantitative data about changes in magnitude and orientation of diffusivity resulting from compaction due to local tissue shrinkage or dilation of adjacent CSF-filled spaces, such as sulcal expansion on the cortex or ventricular enlargement on adjacent structures, such as the caudate or thalamus.
In the present study, motivated by the profound conspicuity of the globus pallidus from signal darkening on diffusion-weighted images (Fig. 1), we took advantage of legacy MRI and DTI data collected in a group of young adults and elderly adults at two field strengths, 1.5 T and 3.0 T, permitting calculation of FDRI, recognizing that the calculation of relaxation can be biased in an fast spin-echo (FSE) acquisition and herein after referred to the FDRI estimate. Our analysis focused on basal ganglia structures. From the FDRI estimate, we predicted a detectable iron presence in putamen, caudate, and globus pallidus but not in a subcortical gray matter comparison structure, the thalamus, or in a frontal white matter taken from the same imaging slices as the striatal samples. Despite age-related decrease in R2 relaxivity expected in all regions and tissue types, we predicted greater presence of iron in the putamen and caudate nucleus in older than younger individuals. We further examined the effects of the FDRI estimate on FA and MD, with the possibility that either measure could be higher or lower in the elderly depending on the amount of iron, the extent of tissue compaction, and the presence of interstitial fluid accumulation.
Fig. 1.
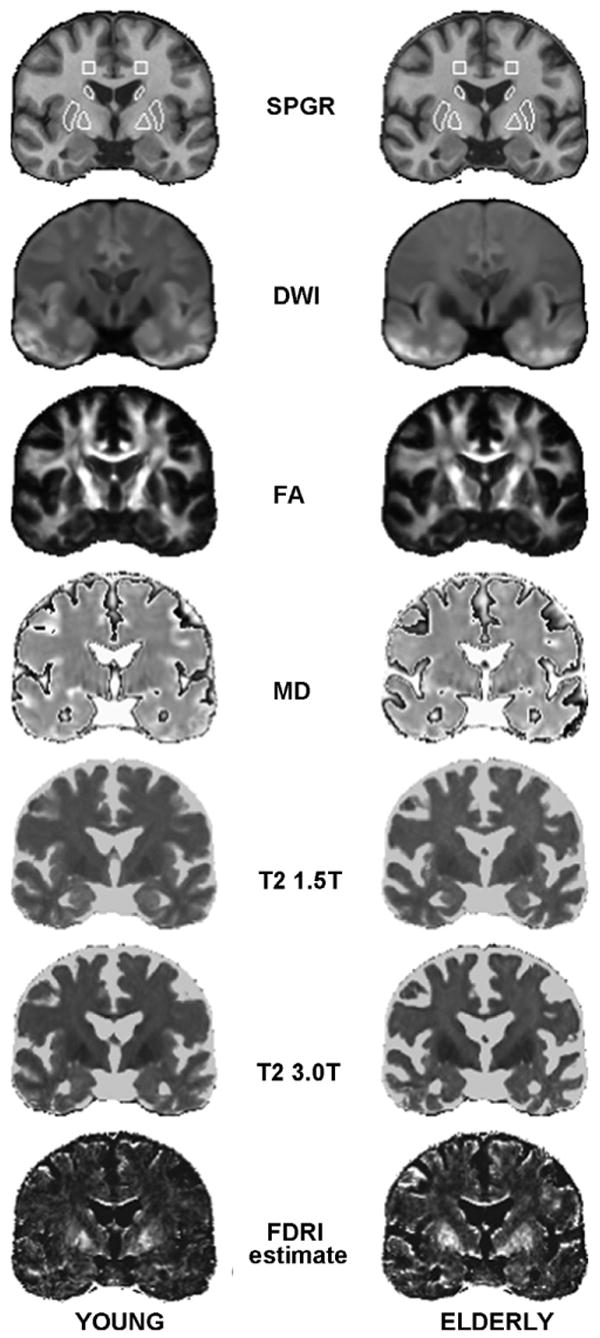
Structural MRI (SPGR), DTI (DWI, FA, and MD), source images for iron quantification (T2 at 1.5 T and T2 at 3.0 T), and images as estimated by FDRI from group averages in common anatomical space constructed separately for young and elderly participants. These mid-coronal images are at a level that includes caudate nucleus, globus pallidus, and putamen. The white matter, caudate nucleus, globus pallidus, and putamen regions of interest (ROIs) are outlined in white on the SPGR images; the slices containing the thalamus ROI are posterior to this slice.
2. Methods
2.1. Subjects
Two groups of healthy, highly educated adults were studied:10 younger (mean = 28.6, range = 22–37 years, 17.2 years of education; 5 men, 5 women) and 10 older (mean = 72.2, range = 65–79 years, 16.3 years of education; 5 men, 5 women). The younger subjects included laboratory members and men and women recruited from the local community. All older subjects were recruited from a larger ongoing study of normal aging and scored well within the normal range on dementia screening tests: Mini-Mental State Examination (Folstein et al., 1975) mean = 28.9, range = 27–30 out of 30 and the Dementia Rating Scale (Mattis, 1988) mean = 140.1, range = 138–143 out of 144. In addition to the DTI study at 3 T, 17 of the 20 subjects (10 older and 7 younger) also had been scanned in other protocols on a 1.5 T scanner within about year (mean = 213, range = 10–383 days). Appropriate approval and procedures were used concerning human subjects.
2.2. MRI and DTI acquisition protocol
The 3 T DTI-structural acquisition included a separate acquisition of a field map to be used for correction of spatial distortion due to main field (B0) inhomogeneity. MRI data were acquired on a 3-T General Electric (Milwaukee, WI) Signa human MRI scanner (gradient strength = 40 mT/m; slew rate = 150 T/m/s); software version VH3). Four axial sequences were collected: (1) Structural Fast Spin-Echo (FSE; FOV = 24cm, Tr = 10,000 ms TE = 14/98 ms, ETL = 8, thick = 2.5 mm, skip = 0 mm, slices = 62, echo train flip angle = 160°); (2) Inversion Recovery Prepared SPoiled Gradient Recalled echo (IRPrep-SPGR; FOV = 24cm, TI = 300 ms, TR/TE = 6.5/1.54 ms, thick = 1.25 mm, slices = 124); (3) Diffusion Tensor Images (DTI; FOV = 24cm, Tr = 11,000 ms TE = 97.6 ms, thick = 2.5 mm, skip = 0, slices = 62, b = 0 (8 NEX) + 6 non-collinear diffusion directions (4 NEX, gradient orientations +x + y, +y + z, +x + z, −x + y, −y + z, +x − z with a repeat of these six orientations with opposite gradient polarity, 1.5 Gauss/cm with 32 ms duration and 34 ms separation, resulting in a b-value of 860 s/mm2, x-dim = 96, y-dim = 96, 3472 total images); (4) Fieldmap (FOV= 24 cm, multi-slice, dual echo, multi-shot (16) spiral acquisition, x-dim = 128, y-dim =128). The SPGR data were aligned such that adjacent pairs of 1.25-mm thick SPGR slices subtended each 2.5-mm thick FSE and DTI slice using custom scanner prescription software, which computed precise slice locations. The data from the spiral acquisition for each echo were gridded and Fourier transformed, and a fieldmap was estimated from a complex difference image between the two echoes (Glover and Lai, 1998; Pfeuffer et al., 2002).
The 1.5 T structural data used for this analysis were coronally-acquired FSE images (FOV = 24 cm, Tr = 7500 ms TE = 14/98 ms, ETL = 8, thick = 4.0 mm, skip = 0 mm, slices = 47, and echo-train flip angle = 180°).
2.3. DTI analysis
DTI quantification was preceded by eddy current correction on a slice-by-slice basis by within-slice registration, which takes advantage of the symmetry of the opposing polarity acquisition (Bodammer et al., 2004). The reversing diffusion gradient polarity scheme also allowed for compensation of the diffusion effect created by the imaging gradients by averaging the opposite polarity data (Neeman et al., 1991), reducing the data to six non-collinear diffusion-weighted images per slice. Using the field maps, B0-field inhomogeneity induced geometric distortion in the eddy current-corrected images was corrected with PRELUDE (Phase Region Expanding Labeller for Unwrapping Discrete Estimates (Jenkinson, 2003)) and FUGUE (FMRIB's Utility for Geometrically Unwarping EPIs (Jenkinson, 2001)).
An average diffusion-weighted image was constructed from the average of the six diffusion-weighted images and signal intensity expressed in institutional units. From the b = 0 and 6 diffusion-weighted images, six maps of the apparent diffusion coefficient (ADC) were calculated. Solving six simultaneous equations with respect to ADCxx, ADCxy, etc. yielded the elements of the diffusion tensor. The diffusion tensor was then diagonalized, yielding eigenvalues λ1, λ2, λ3, as well as eigenvectors that define the predominant diffusion orientation. Based on the eigenvalues, FA, ADC, λL (λ1), and λT ((λ2 + λ3)/2) were calculated on a voxel-by-voxel basis (Basser and Jones, 2002; Basser and Pierpaoli, 1998; Pierpaoli and Basser, 1996).
2.4. Image registration
Voxel-by-voxel maps of R2 estimates were calculated by two-point log fit for the 3 T and 1.5 T FSE data separately. Because these maps were based on FSE rather than true spin-echo data, we refer to this measure as an R2 estimate. For the 3 T data, within each subject, after field map correction, the b = 0 EPI data were registered to the late echo FSE data with non-rigid registration (Rohlfing and Maurer, 2003) to compensate for residual uncorrected B0 distortion. The late echo FSE data were in turn registered to the SPGR data. Each subject's SPGR data were then registered to the laboratory SPGR atlas (Rohlfing et al., 2008) with non-rigid registration. These transformations were aggregated by concatenation and applied to the FA, MD, b = 0 EPI, DWI, R2 estimate, early and late echo FSE and SPGR data for each subject and all were reformatted, each with a single tri-cubic interpolation, into 1 mm isotropic data sets in atlas space.
The 1.5 T late echo FSE data were registered to the 3 T late echo FSE data, the transformation added by concatenation to the above transformation train and applied to the 1.5 T R2 estimate, which were also reformatted, each with a single tri-cubic interpolation, into 1 mm isotropic data sets in atlas space.
2.5. Region-of-interest analysis
Bilateral caudate, globus pallidus, putamen, thalamus and white matter sample regions of interest (ROI) were drawn (by A.P.) on the laboratory SPGR atlas presented in the coronal plane. The globus pallidus, putamen, caudate and white matter sample were drawn on 10 contiguous 1-mm thick slices at an anterior–posterior location that maximized the presence of all three basal ganglia structures (see Fig. 1). The thalamus was drawn on the next 10 contiguous slices posterior to the basal ganglia. The ROIs were used to quantify the various DTI metrics for each subject by analyzing their values within each ROI after transformation into laboratory atlas space. The caudate and thalamus ROIs were eroded one and two pixels respectively on a slice-by-slice basis to avoid partial voluming of CSF. The iron concentration was estimated by comparing the R2 estimates across the two field strengths (1.5 and 3 T) for each ROI (Bartzokis et al., 2007b). After maps of the R2 estimates were constructed, the median the R2 estimate for each ROI for each field strength was computed. The iron measure was computed as (R2@3T − R2@1.5 T)/1.5, and yielded a numerical range across all subjects of .07–4.1 s−1.
2.6. Statistical analysis
Two group (young vs. elderly) by two hemisphere (left vs. right) repeated measures analyses of variance (ANOVA) were conducted for each of the five bilateral brain regions (globus pallidus, putamen, caudate nucleus, thalamus, and the white matter sample) and each brain measure (FA, MD, DWI, R2 estimates at 1.5 T and 3 T, and FDRI estimate). We expected group effects in basal ganglia but not white matter or thalamic regions, but did not expect group-by-hemisphere interactions, that is, we predicted that in regions showing an age effect both hemispheres would be similarly affected by age. Additional repeated measures ANOVAs examined the profile of group differences across the five ROIs for each brain metric. Follow-up comparisons were based on within-subject and between-subject t-tests. Pearson product-moment correlations tested relations between DWI values and FDRI estimates and were confirmed with non-parametric Spearman Rank Order tests to minimize bias from outlying values.
3. Results
3.1. Effect of age on the five bilateral regions of interest
These analyses examined the differential effect of age on the imaging metrics focused on the globus pallidus, putamen, caudate nucleus, thalamus, and frontal white matter. The data were acquired at 3 T in the full group of 20 men and women, and the estimated field-dependent measure of iron content (FDRI estimate) was based on the 7 young and 10 elderly subjects with data acquired both at 1.5 T and 3 T. The ANOVA statistics are presented in Table 1 and described below. Significant group-by-hemisphere interactions were observed in six comparisons, but in none of these cases did the interaction determine the group effect. Consequently, the following analyses used the mean of the left and right hemisphere values for each imaging metric (Fig. 2).
Table 1.
Repeated measures ANOVAs: two groups by two hemispheres
| Frontal White Matter | Globus Pallidus | Thalamus | Caudate Nucleus | Putamen | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | ||
| FA (N = 20) | |||||||||||
| Group | 64.30 | 0.0001 | 2.97 | 0.102 | 0.06 | 0.82 | 23.47 | 0.0001 | 7.91 | 0.012 | |
| Hemisphere | 0.05 | 0.82 | 3.27 | 0.087 | 3.34 | 0.08 | 2.38 | 0.14 | 0.07 | 0.79 | |
| G × H | 1.38 | 0.26 | 0.61 | 0.446 | 0.55 | 0.47 | 0.18 | 0.68 | 0.23 | 0.64 | |
| MD (N = 20) | |||||||||||
| Group | 37.76 | 0.0001 | 17.75 | 0.0005 | 2.00 | 0.17 | 1.78 | 0.199 | 3.68 | 0.07 | |
| Hemisphere | 0.03 | 0.86 | 0.08 | 0.78 | 0.01 | 0.95 | 17.87 | 0.0005 | 0.02 | 0.91 | |
| G × H | 0.75 | 0.398 | 4.50 | 0.05 | 0.50 | 0.49 | 2.47 | 0.13 | 0.11 | 0.75 | |
| λL (N = 20) | |||||||||||
| Group | 0.95 | 0.34 | 4.89 | 0.0408 | 5.25 | 0.0343 | 20.14 | 0.0003 | 9.84 | 0.0057 | |
| Hemisphere | 0.21 | 0.65 | 1.66 | 0.21 | 1.80 | 0.20 | 5.80 | 0.027 | 0.10 | 0.75 | |
| G × H | 0.18 | 0.67 | 5.82 | 0.0267 | 0.30 | 0.59 | 5.12 | 0.0362 | 0.15 | 0.70 | |
| λT (N = 20) | |||||||||||
| Group | 84.87 | 0.0001 | 16.40 | 0.0008 | 0.82 | 0.38 | 0.13 | 0.73 | 1.62 | 0.22 | |
| Hemisphere | 0.19 | 0.67 | 0.03 | 0.87 | 0.60 | 0.45 | 19.03 | 0.0004 | 0.01 | 0.99 | |
| G × H | 1.42 | 0.25 | 3.30 | 0.0858 | 1.56 | 0.23 | 0.69 | 0.42 | 0.08 | 0.78 | |
| DWI (N = 20) | |||||||||||
| Group | 10.20 | 0.005 | 1.41 | 0.25 | 13.46 | 0.0018 | 1.17 | 0.29 | 16.13 | 0.0008 | |
| Hemisphere | 0.02 | 0.904 | 1.40 | 0.25 | 10.80 | 0.0041 | 7.97 | 0.0113 | 2.04 | 0.17 | |
| G × H | 0.45 | 0.51 | 3.85 | 0.0656 | 8.16 | 0.0105 | 0.03 | 0.87 | 5.49 | 0.031 | |
| R2 estimate @ 3 T (N = 20) | |||||||||||
| Group | 17.62 | 0.0005 | 1.66 | 0.214 | 42.64 | 0.0001 | 0.10 | 0.76 | 7.58 | 0.013 | |
| Hemisphere | 0.29 | 0.60 | 6.23 | 0.0225 | 2.45 | 0.14 | 3.21 | 0.09 | 0.24 | 0.629 | |
| GX H | 0.01 | 0.92 | 10.75 | 0.0042 | 0.86 | 0.36 | 0.15 | 0.7 | 0.78 | 0.39 | |
| R2 estimate @ 1.5 T (N = 17) | |||||||||||
| Group | 20.57 | 0.0004 | 2.79 | 0.12 | 24.07 | 0.0002 | 0.00 | 0.97 | 3.53 | 0.08 | |
| Hemisphere | 1.99 | 0.18 | 2.54 | 0.13 | 0.91 | 0.36 | 0.82 | 0.38 | 2.12 | 0.17 | |
| G × H | 1.26 | 0.28 | 2.48 | 0.14 | 0.26 | 0.62 | 0.97 | 0.34 | 1.47 | 0.24 | |
| FDRI estimate (N = 17) | |||||||||||
| Group | 0.10 | 0.76 | 1.25 | 0.28 | 14.79 | 0.0016 | 0.28 | 0.61 | 5.83 | 0.029 | |
| Hemisphere | 5.80 | 0.03 | 4.09 | 0.0612 | 1.02 | 0.33 | 0.77 | 0.39 | 1.72 | 0.209 | |
| G × H | 0.24 | 0.63 | 2.36 | 0.15 | 0.12 | 0.73 | 0.09 | 0.77 | 0.06 | 0.81 | |
Fig. 2.
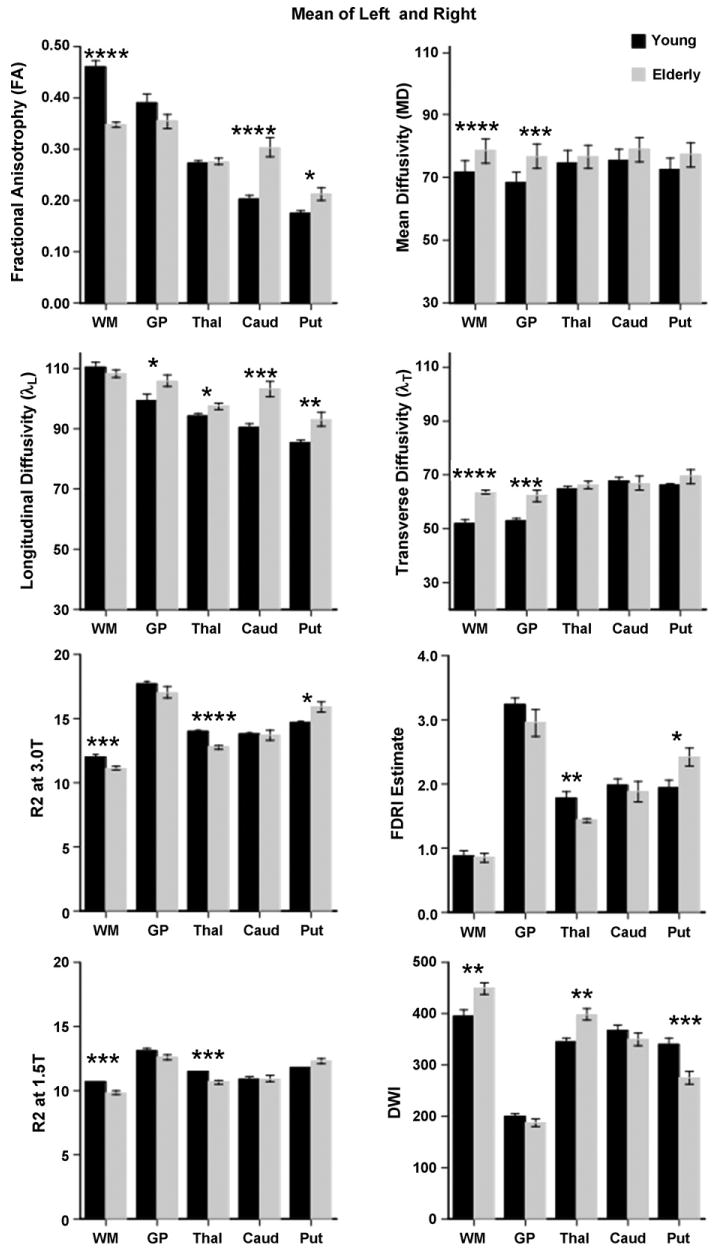
Mean ± S.E.M. of each bilateral DTI, MRI, and FDRI estimate metric for the young and elderly groups. *p ≤ .05, **p ≤ .01, ***p ≤ .001, and ****p ≤ .0001.
3.1.1. Anisotropy and magnitude of diffusion in gray matter and white matter
A repeated measures ANOVA across the five ROIs for FA yielded a significant interaction (p = .0001). Follow-up comparisons showed that FA was significantly higher in the elderly than young group in the putamen (p = .0115) and caudate nucleus (p = .0001) but significantly lower in the frontal white matter sample (p = .0001) and showed a trend to be lower in the globus pallidus (p = .1019). FA in the thalamus showed no age effect (p = .8165).
For MD, a repeated measures ANOVA across the five ROIs revealed higher MD in the elderly than the young group across the five brain regions (p = .0003). The group differences were significant in the globus pallidus (p = .0005) and white matter (p = .0001) but not in the putamen (p = .0712), caudate nucleus (p = .1992), or thalamus (p = .1749). The regional pattern of age effects was the same for transverse diffusivity and MD but different for longitudinal diffusivity. Although the group-by-region interaction was significant (p = .0007), the only region not to show an age effect was frontal white matter longitudinal diffusivity; thus, unlike transverse diffusivity, the older group had higher longitudinal diffusivity in the thalamus, caudate, and putamen as well as the globus pallidus.
For DWI, the ANOVA across the five ROIs revealed a significant group-by-region interaction (p = .0001) and opposite age effects in three ROIs. The elderly group had lower image intensity than the young group in the putamen (p = .0008) but higher intensity in the thalamus (p = .0105) and white matter (p = .005). Group differences were not significant in the globus pallidus (p = .2499) or caudate nucleus (p = .2929).
3.1.2. Indices of iron structures of the basal ganglia, thalamus, and frontal white matter
A repeated measures ANOVA for R2 identified interactions at both field strengths (p = .0001). The age effect on R2 relaxation rate at 3.0 T was significant in the putamen (p = .0136), where R2 was higher in the elderly than young group. By contrast, R2 was lower in the elderly in the thalamus (p = .0001) and white matter (p = .0005); although lower in the globus pallidus, the effect of age was not significant (p = .214). At 1.5 T field strength, the pattern of group differences was the same as at the higher field strength, but higher rate with age was only marginal in the putamen (p = .082). The mean R2 estimates were significantly lower at 1.5 T than at 3.0 T (p = .0001 for all five regions).
The FDRI estimated measure of iron was derived from R2 acquired at the two MR field strengths, 1.5 T and 3 T, available for 7 of the 10 younger subjects and all 10 elderly subjects. Two regions showed significant group effects with ANOVA: for the putamen, the elderly had higher FDRI estimates than the young (p = .029), whereas the opposite was the case for the thalamus (p = .0016). This age-related pattern difference across ROIs was confirmed with a repeated measures ANOVA across the five ROIs that yielded a significant interaction (p = .0066).
3.2. Correlations of FDRI with DTI metrics
The mean values of the left and right hemispheres were used to examine correlations among brain measures. Initial analyses examined the relation between the FDRI estimate and the five DTI metrics (FA, MD, λL, λT, and DWI). Three correlations met family-wise Bonferroni correction for five multiple comparisons, requiring p ≤ .01: Low diffusivity on DWI correlated with high FDRI estimates in the thalamus (r = −.62, p = .0088; Rho = −.53, p = .0342) and putamen (r = −.81, p = .0001; Rho = −79, p = .0016). Although high FA correlated with high FDRI estimates in the putamen (r = .62, p = .0079), this correlation was not confirmed with a non-parametric test because of an outlying value (Rho = .30, p = .2355) (Table 2 and Fig. 3).
Table 2.
Pearson correlations between regional FDRI estimates and DTI metrics (N = 17)
| Frontal white matter | Globus pallidus | Thalamus | Caudate nucleus | Putamen | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | r | p-Value | r | p-Value | |
| FA | 0.11 | 0.6893 | 0.33 | 0.2005 | 0.07 | 0.8035 | −0.12 | 0.6368 | 0.62 | 0.0079 |
| MD | −0.10 | 0.7053 | −0.57 | 0.0175 | −0.26 | 0.322 | −0.13 | 0.6312 | −0.33 | 0.1936 |
| λL | 0.09 | 0.7411 | −0.46 | 0.0661 | −0.33 | 0.1976 | −0.22 | 0.3993 | −0.18 | 0.4947 |
| λT | −0.08 | 0.7494 | −0.53 | 0.0288 | −0.23 | 0.3772 | −0.03 | 0.9083 | −0.37 | 0.1482 |
| DWI | 0.07 | 0.7797 | −0.35 | 0.1668 | −0.62 | 0.0088 | −0.26 | 0.316 | −0.81 | 0.0001 |
Family-wise Bonferroni corrected p = .01.
Fig. 3.

Significant correlations between regional FDRI and DTI metrics.
4. Discussion
This analysis was prompted by the observation based on retrospective analysis of legacy data that in a DTI study of young and old subjects there was a conspicuous decrease in signal intensity on the aggregated DW basis images used for computation of the quantitative diffusion metrics. The fact that these legacy data were available with accompanying R2 estimates at two field strengths, 1.5 T and 3.0 T, led us to the speculation that age-related iron deposition in the basal ganglia contributed to this observation. In spite of the plethora of limitations of the data set and the low statistical power, there are tantalizing suggestions that the quantitative metrics of diffusivity are indeed affected, that is, there are trends for negative correlations in the expected direction between FDRI estimates and the diffusivity measures in all regions examined. This phenomenon needs to be established with unbiased relaxivity estimates. If confirmed, it would suggest that iron deposition influences DTI and would provide further insight into a mechanistic explanation of brain changes with age and diseases of the basal ganglia.
4.1. Age, subcortical gray matter, and iron
The effect of age on the diffusion measures in the deep gray matter structures was distinctly different from that reported in white matter. In contrast to the notably lower FA and higher diffusivity especially prominent in frontal white matter in older than younger healthy adults (for review, Sullivan and Pfefferbaum, 2006), FA and diffusivity were higher, albeit insignificantly for diffusivity, in the older than younger group in the caudate nucleus and putamen. Consideration of the diffusivity components revealed another difference in age-related patterns between the white matter sample and the caudate and putamen. For white matter, the elderly had higher values than the young in transverse diffusivity (λT, a putative marker of myelin integrity) but not longitudinal diffusivity (λL, a putative marker of axonal integrity) (Song et al., 2003, 2002), whereas longitudinal rather than transverse diffusivity was greater in the four subcortical gray matter structures measured of the elderly than young. The thalamus showed little effect of age in anisotropy or diffusivity. The DWI metric showed a regionally distinct pattern of age-related differences not necessarily related to tissue type: DWI was lower in the putamen of the elderly relative to the young, whereas the opposite held true for the white matter region and thalamus, perhaps related to the high white matter presence in the thalamus. Thus, the differential effects of age on DTI metrics in subcortical gray matter structures compared with white matter tracts appears to be related, at least in part, to local iron content, which in the elderly of the present study was prominent in the FDRI estimates of the putamen and visibly striking in the diffusion-weighted image of the basal ganglia structures. We further note that in Table 2 all the diffusivity measures correlate negatively with FDRI estimates and although not individually significant, in the aggregate are supportive of our speculation that the greater presence of iron underlies this effect. Despite the regional pattern difference between age and FDRI, the MD was negatively correlated with FDRI estimates.
It has been speculated that age-related decline in the ferritin iron index of white matter and thalamus, which contains significant myelin, should be accompanied by myelin breakdown owing to oxidative stress caused by iron release (Bartzokis et al., 2007b). We tested this hypothesis by examining the correlation between FDRI estimates and transverse diffusivity, a diffusion index of myelin degradation. Although the hypothesis was not supported as tested, high FDRI estimates did relate to low DWI signal intensity in the thalamus and putamen, likely reflecting signal attenuation from iron. High FDRI estimates also correlated with high FA in putamen, the meaning of which is less clear but open to the speculation that high iron presence may be associated with extent of tissue compaction, and that concurrent high MD, notably longitudinal diffusivity, occurs in the presence of interstitial fluid accumulation in a region. Nonetheless, the interpretation of age-related changes in DTI metrics in deep gray matter structures appears to be different from that in white matter. In addition to the obvious differences in underlying microstructure, myelinated linear structures vs. predominantly neurons and glia, the presence of iron appears to have a greater effect on anisotropy than mean diffusivity.
The conspicuity of the R2-like increases due to iron deposition in the brain is enhanced in the presence of diffusion weighting gradients (i.e., DWI) in some regions, especially globus pallidus and putamen. The reasons for this are not immediately apparent. The spin–spin T2 (signal darkening) effects are on a molecular distance scale, over which the diffusion weighting gradients are negligible, but can affect the competing “T2 shine-through effect,” caused by signal brightening from the presence of interstitial fluid. Attenuation with diffusion weighting gradients of age-related T2 shine-through effect could uncover iron-dependent increase in the R2 estimate. Further, the magnitude of iron-induced signal loss may depend on the magnitude of local water diffusion near the iron molecules (Jensen et al., 2006) and the presence of diffusion weighting gradients could enhance the signal decay. The role of iron in increased anisotropy in deep gray matter structures is similarly puzzling and may be related to actual alterations in the condition of microstructural tissue.
4.2. Limitations
The retrospective nature of this study imposes limitations to acknowledge that by definition cannot be remedied. Firstly, the sample of younger and older healthy individuals with MR imaging data at two field strengths and with structural as well as diffusion tensor imaging data was small, thus limiting our conclusions, including those regarding functional correlations, and precluding secondary analyses, for example, examination of sex differences (c.f., Bartzokis et al., 2007b). Secondly, as a retrospective study, we did not have an optimized imaging acquisition protocol targeting iron. For example, slice thickness differed between acquisitions, being 4 mm for 1.5 T data and 2.5mm for 3 T data; thus, the 1.5 T data (with lower native resolution) required more interpolation than the 3 T data when the data were combined into isotropic space. Under ideal conditions of excitation homogeneity, the estimates of R2 can be made efficient by encoding FSE images with a train of refocusing pulses in a dual-echo sequence. While at 1.5 T, B1 excitation homogeneity is typically excellent on current clinical scanners, higher field strengths such as 3 T suffer from greater spatial variation in the excitation B1 fields. In addition, power deposition limits for in vivo scanning at high field are more likely to constrain the acquisition prescription than at 1.5 T. For these reasons, refocusing trains at 3 T may be implemented with flip angles lower than 180°, which substantially reduces peak and average power, but introduces a bias to the resulting R2 estimate. The bias term is more pronounced at the periphery of the axial slices of interest, whereas medial regions (such as the central gray matter) are less affected. Nonetheless, the imaging parameters and data quality were adequate to detect robust and regionally differential effects of iron, albeit underestimating FDRI because of the lower flip angles at 3 T, and suggest the need to consider age-related iron accumulation in the interpretation of diffusion tensor imaging studies.
4.3. Conclusions
Until very old age, white matter volume is more preserved relative to the profound degradation in microstructure detectable with DTI. By contrast, cortical gray matter as well as deep gray matter structures recede in volume with advancing age, yet the deep gray matter structures, like regional white matter, also exhibit age-related changes in anisotropy and diffusivity detectable with DTI. The fact that gray matter FA increases rather than decreases with age as it does in white matter warrants further investigation and may be influenced by or reflective of iron deposition. The regionally differential distribution of age-related iron accumulation in the brain may serve to explain certain age-related declines in function and may also have ramifications for studies of aging that rely on brain imaging protocols potentially affected by iron, such as functional MRI, which is founded on the T2*, blood oxygenation level dependent (BOLD) effect.
Acknowledgments
This work was supported by U.S. National Institutes of Health grants AG17919, AA05965, AA10723, and AA12388.
Footnotes
Disclosure statement: (a) There are no actual or potential conflicts of interest including any financial, personal or other relationships with other people or organizations within 3 years of beginning the work submitted that could inappropriately influence (bias) their work. No author's institution has contracts relating to this research through which it or any other organization may stand to gain financially now or in the future. There are no other agreements of authors or their institutions that could be seen as involving a financial interest in this work. (b) Appropriate approval and procedures were used concerning human subjects and obtained prior to obtaining data.
References
- Aboitiz F, Rodriguez E, Olivares R, Zaidel E. Age-related changes in fibre composition of the human corpus callosum: sex differences. Neuroreport. 1996;7(11):1761–1764. doi: 10.1097/00001756-199607290-00013. [DOI] [PubMed] [Google Scholar]
- Bartzokis G. Age-related myelin breakdown: a developmental model of cognitive decline and Alzheimer's disease. Neurobiol Aging. 2004;25(1):5–18. doi: 10.1016/j.neurobiolaging.2003.03.001. author reply 49–62. [DOI] [PubMed] [Google Scholar]
- Bartzokis G, Aravagiri M, Oldendorf WH, Mintz J, Marder SR. Field dependent transverse relaxation rate increase may be a specific measure of tissue iron stores. Magn Reson Med. 1993;29:459–464. doi: 10.1002/mrm.1910290406. [DOI] [PubMed] [Google Scholar]
- Bartzokis G, Cummings JL, Markham CH, Marmarelis PZ, Treciokas LJ, Tishler TA, Marder SR, Mintz J. MRI evaluation of brain iron in earlier- and later-onset Parkinson's disease and normal subjects. Magn Reson Imag. 1999;17(2):213–222. doi: 10.1016/s0730-725x(98)00155-6. [DOI] [PubMed] [Google Scholar]
- Bartzokis G, Lu PH, Tishler TA, Fong SM, Oluwadara B, Finn JP, Huang D, Bordelon Y, Mintz J, Perlman S. Myelin breakdown and iron changes in Huntington's disease: pathogenesis and treatment implications. Neurochem Res. 2007a;32(10):1655–1664. doi: 10.1007/s11064-007-9352-7. [DOI] [PubMed] [Google Scholar]
- Bartzokis G, Mintz J, Sultzer D, Marx P, Herzberg JS, Phelan CK, Marder SR. In vivo MR evaluation of age-related increases in brain iron. Am J Neuroradiol. 1994;15(6):1129–1138. [PMC free article] [PubMed] [Google Scholar]
- Bartzokis G, Tishler TA, Lu PH, Villablanca P, Altshuler LL, Carter M, Huang D, Edwards N, Mintz J. Brain ferritin iron may influence age- and gender-related risks of neurodegeneration. Neurobiol Aging. 2007b;28(3):414–423. doi: 10.1016/j.neurobiolaging.2006.02.005. [DOI] [PubMed] [Google Scholar]
- Basser PJ, Jones DK. Diffusion-tensor MRI: theory, experimental design and data analysis—a technical review. NMR Biomed. 2002;15(7–8):456–467. doi: 10.1002/nbm.783. [DOI] [PubMed] [Google Scholar]
- Basser PJ, Pierpaoli C. A simplified method to measure the diffusion tensor from seven MR images. Magn Reson Med. 1998;39(6):928–934. doi: 10.1002/mrm.1910390610. [DOI] [PubMed] [Google Scholar]
- Basser PJ, Pierpaoli C. Microstructural and physiological features of tissues elucidated by quantitative diffusion tensor MRI. J Magn Reson B. 1996;111:209–219. doi: 10.1006/jmrb.1996.0086. [DOI] [PubMed] [Google Scholar]
- Bizzi A, Armstrong M, Dietz M, Fulmah M, Frank J, DeChiro G, Alger J. Proton spectroscopic imaging of frontal and parietal lobes in normal human brain. Proceedings of the Annual Meeting of Society of Magnetic Resonance in Medicine; 1906.1992. [Google Scholar]
- Bizzi A, Brooks RA, Brunetti A, Hill JM, Alger JR, Miletich RS, Francavilla TL, Di Chiro G. Role of iron and ferritin in MR imaging of the brain: a study in primates at different field strengths. Radiology. 1990;177(1):59–65. doi: 10.1148/radiology.177.1.2399339. [DOI] [PubMed] [Google Scholar]
- Bodammer N, Kaufmann J, Kanowski M, Tempelmann C. Eddy current correction in diffusion-weighted imaging using pairs of images acquired with opposite diffusion gradient polarity. Magn Reson Med. 2004;51(1):188–193. doi: 10.1002/mrm.10690. [DOI] [PubMed] [Google Scholar]
- Chen ZG, Li TQ, Hindmarsh T. Diffusion tensor trace mapping in normal adult brain using single-shot EPI technique. A methodological study of the aging brain. Acta Radiol. 2001;42(5):447–458. doi: 10.1080/028418501127347160. [DOI] [PubMed] [Google Scholar]
- Engelter ST, Provenzale JM, Petrella JR, DeLong DM, MacFall JR. The effect of aging on the apparent diffusion coefficient of normal-appearing white matter. Am J Roentgenol. 2000;175(2):425–430. doi: 10.2214/ajr.175.2.1750425. [DOI] [PubMed] [Google Scholar]
- Folstein MF, Folstein SE, McHugh PR. Mini-mental state:a practical method for grading the cognitive state of patients for the clinician. J Psychiatr Res. 1975;12:189–198. doi: 10.1016/0022-3956(75)90026-6. [DOI] [PubMed] [Google Scholar]
- Gelman BB, Guinto FC. Morphometry, histopathology, and tomography of cerebral atrophy in the acquired immunodeficiency syndrome. Ann Neurol. 1992;32:31–40. doi: 10.1002/ana.410320107. [DOI] [PubMed] [Google Scholar]
- Glover GH, Lai S. Self-navigated spiral fMRI: interleaved versus single-shot. Magn Reson Med. 1998;39:361–368. doi: 10.1002/mrm.1910390305. [DOI] [PubMed] [Google Scholar]
- Gomori JM, Grossman RI. The relation between regional brain iron and T2 shortening. Am J Neuroradiol. 1993;14:1049–1050. [PMC free article] [PubMed] [Google Scholar]
- Grieve SM, Williams LM, Paul RH, Clark CR, Gordon E. Cognitive aging, executive function, and fractional anisotropy: a diffusion tensor MR imaging study. AJNR Am J Neuroradiol. 2007;28(2):226–235. [PMC free article] [PubMed] [Google Scholar]
- Haacke EM, Ayaz M, Khan A, Manova ES, Krishnamurthy B, Gollapalli L, Ciulla C, Kim I, Petersen F, Kirsch W. Establishing a baseline phase behavior in magnetic resonance imaging to determine normal vs abnormal iron content in the brain. J Magn Reson Imag. 2007;26(2):256–264. doi: 10.1002/jmri.22987. [DOI] [PubMed] [Google Scholar]
- Haacke EM, Cheng NY, House MJ, Liu Q, Neelavalli J, Ogg RJ, Khan A, Ayaz M, Kirsch W, Obenaus A. Imaging iron stores in the brain using magnetic resonance imaging. Magn Reson Imag. 2005;23:1–25. doi: 10.1016/j.mri.2004.10.001. [DOI] [PubMed] [Google Scholar]
- Hallgren B, Sourander P. The effect of age on the non-haemin iron in the human brain. J Neurochem. 1958;3(1):41–51. doi: 10.1111/j.1471-4159.1958.tb12607.x. [DOI] [PubMed] [Google Scholar]
- Head D, Buckner RL, Shimony JS, Williams LE, Akbudak E, Conturo TE, McAvoy M, Morris JC, Snyder AZ. Differential vulnerability of anterior white matter in nondemented aging with minimal acceleration in dementia of the Alzheimer type: evidence from diffusion tensor imaging. Cereb Cortex. 2004;14(4):410–423. doi: 10.1093/cercor/bhh003. [DOI] [PubMed] [Google Scholar]
- Helenius J, Soinne L, Perkio J, Salonen O, Kangasmaki A, Kaste M, Carano RA, Aronen HJ, Tatlisumak T. Diffusion-weighted MR imaging in normal human brains in various age groups. Am J Neuroradiol. 2002;23(2):194–199. [PMC free article] [PubMed] [Google Scholar]
- Hiwatashi A, Kinoshita T, Moritani T, Wang HZ, Shrier DA, Numaguchi Y, Ekholm SE, Westesson PL. Hypointensity on diffusion-weighted MRI of the brain related to T2 shortening and susceptibility effects. AJR Am J Roentgenol. 2003;181(6):1705–1709. doi: 10.2214/ajr.181.6.1811705. [DOI] [PubMed] [Google Scholar]
- Huang H, Zhang J, Wakana S, Zhang W, Ren T, Richards LJ, Yarowsky P, Donohue P, Graham E, van Zijl PC, Mori S. White and gray matter development in human fetal, newborn and pediatric brains. NeuroImage. 2006;33(1):27–38. doi: 10.1016/j.neuroimage.2006.06.009. [DOI] [PubMed] [Google Scholar]
- Jenkinson M. A fast, automated, N-dimensional phase unwrapping algorithm. J Magn Reson Med. 2003;49:193–197. doi: 10.1002/mrm.10354. [DOI] [PubMed] [Google Scholar]
- Jenkinson M. Human Brain Mapping – HBM200. 2001. Improved unwarping of EPI volumes using regularised B_0 Maps (abs) [Google Scholar]
- Jensen JH, Chandra R, Ramani A, Lu H, Johnson G, Lee SP, Kaczynski K, Helpern JA. Magnetic field correlation imaging. Magn Reson Med. 2006;55(6):1350–1361. doi: 10.1002/mrm.20907. [DOI] [PubMed] [Google Scholar]
- Kochunov P, Thompson PM, Lancaster JL, Bartzokis G, Smith S, Coyle T, Royall DR, Laird A, Fox PT. Relationship between white matter fractional anisotropy and other indices of cerebral health in normal aging: tract-based spatial statistics study of aging. NeuroImage. 2007;35(2):478–487. doi: 10.1016/j.neuroimage.2006.12.021. [DOI] [PubMed] [Google Scholar]
- Le Bihan D. Looking into the functional architecture of the brain with diffusion MRI. Nat Rev Neurosci. 2003;4(6):469–480. doi: 10.1038/nrn1119. [DOI] [PubMed] [Google Scholar]
- Liu T, Li H, Wong K, Tarokh A, Guo L, Wong ST. Brain tissue segmentation based on DTI data. NeuroImage. 2007;38(1):114–123. doi: 10.1016/j.neuroimage.2007.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma J, Wehrli FW. Method for image-based measurement of the reversible and irreversible contribution to the transverse-relaxation rate. J Magn Reson. 1996;111(1):61–69. doi: 10.1006/jmrb.1996.0060. [DOI] [PubMed] [Google Scholar]
- Madden DJ, Whiting WL, Huettel SA, White LE, MacFall JR, Provenzale JM. Diffusion tensor imaging of adult age differences in cerebral white matter: relation to response time. NeuroImage. 2004;21(3):1174–1181. doi: 10.1016/j.neuroimage.2003.11.004. [DOI] [PubMed] [Google Scholar]
- Majumdar S, Zoghbi S, Pope CF, Gore JC. A quantitative study of relaxation rate enhancement produced by iron oxide particles in polyacrylamide gels and tissue. Magn Reson Med. 1989;9(2):185–202. doi: 10.1002/mrm.1910090205. [DOI] [PubMed] [Google Scholar]
- Martin WR, Roberts TE, Ye FQ, Allen PS. Increased basal ganglia iron in striatonigral degeneration: in vivo estimation with magnetic resonance. Can J Neurol Sci. 1998;25(1):44–47. doi: 10.1017/s0317167100033473. [DOI] [PubMed] [Google Scholar]
- Mattis S. Dementia Rating Scale (DRS) Professional Manual. Psychological Assessment Resources, Inc.; Odessa, FL: 1988. [Google Scholar]
- Michaeli S, Oz G, Sorce DJ, Garwood M, Ugurbil K, Majestic S, Tuite P. Assessment of brain iron and neuronal integrity in patients with Parkinson's disease using novel MRI contrasts. Mov Disord. 2007;22(3):334–340. doi: 10.1002/mds.21227. [DOI] [PubMed] [Google Scholar]
- Moseley ME, Cohen Y, Kucharczyk J, Mintorovitch J, Asgari HS, Wendland MF, Tsuruda J, Norman D. Diffusion-weighted MR imaging of anisotropic water diffusion in cat central nervous system. Radiology. 1990a;176(2):439–445. doi: 10.1148/radiology.176.2.2367658. [DOI] [PubMed] [Google Scholar]
- Moseley ME, Kucharczyk J, Mintorovitch J, Cohen Y, Kurhanewicz J, Derugin N, Asgari H, Norman D. Diffusion-weighted MR imaging of acute stroke: correlation with T2-weighted and magnetic susceptibility-enhanced MR imaging in cats. Am J Neuroradiol. 1990b;11(3):423–429. [PMC free article] [PubMed] [Google Scholar]
- Moustafa RR, Baron JC. Clinical review: imaging in ischaemic stroke—implications for acute management. Crit Care. 2007;11(5):227. doi: 10.1186/cc5973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naganawa S, Sato K, Katagiri T, Mimura T, Ishigaki T. Regional ADC values of the normal brain: differences due to age, gender, and laterality. Eur Radiol. 2003;13(1):6–11. doi: 10.1007/s00330-002-1549-1. [DOI] [PubMed] [Google Scholar]
- Neeman M, Freyer JP, Sillerud LO. A simple method for obtaining cross-term-free images for diffusion anisotropy studies in NMR microimaging. Magn Reson Med. 1991;21(1):138–143. doi: 10.1002/mrm.1910210117. [DOI] [PubMed] [Google Scholar]
- Norris DG, Niendorf T, Leibfritz D. Healthy and infarcted brain tissues studied at short diffusion times: the origins of apparent restriction and the reudction in apparent diffusion coefficient. NMR Biomed. 1994;7(7):304–310. doi: 10.1002/nbm.1940070703. [DOI] [PubMed] [Google Scholar]
- O'Sullivan M, Jones D, Summers P, Morris R, Williams S, Markus H. Evidence for cortical “disconnection” as a mechanism of age-related cognitive decline. Neurology. 2001;57:632–638. doi: 10.1212/wnl.57.4.632. [DOI] [PubMed] [Google Scholar]
- Ordidge RJ, Gorell JM, Deniau JC, Knight RA, Helpern JA. Abstract assessment of relative brain iron concentrations using T2-weighted and T2*-weighted MRI at 3 Tesla. Magn Reson Med. 1994;32:335–341. doi: 10.1002/mrm.1910320309. [DOI] [PubMed] [Google Scholar]
- Peters A, Sethares C. Is there remyelination during aging of the primate central nervous system? J Comp Neurol. 2003;460(2):238–254. doi: 10.1002/cne.10639. [DOI] [PubMed] [Google Scholar]
- Pfefferbaum A, Adalsteinsson E, Sullivan EV. Frontal circuitry degradation marks healthy adult aging: evidence from diffusion tensor imaging. NeuroImage. 2005;26(3):891–899. doi: 10.1016/j.neuroimage.2005.02.034. [DOI] [PubMed] [Google Scholar]
- Pfefferbaum A, Adalsteinsson E, Sullivan EV. Supratentorial profile of white matter microstructural integrity in recovering alcoholic men and women. Biol Psychiatry. 2006;59(4):364–372. doi: 10.1016/j.biopsych.2005.06.025. [DOI] [PubMed] [Google Scholar]
- Pfefferbaum A, Rosenbloom MJ, Adalsteinsson E, Sullivan EV. Diffusion tensor imaging with quantitative fiber tracking in HIV infection and alcoholism comorbidity: synergistic white matter damage. Brain. 2007;130:48–64. doi: 10.1093/brain/awl242. [DOI] [PubMed] [Google Scholar]
- Pfefferbaum A, Sullivan EV. Diffusion MR imaging in neuropsychiatry and aging. In: Gillard J, Waldman A, Barker P, editors. Clinical MR Neuroimaging: Diffusion, Perfusion and Spectroscopy. Cambridge University Press; Cambridge: 2005a. pp. 558–578. [Google Scholar]
- Pfefferbaum A, Sullivan EV. Disruption of brain white matter microstructure by excessive intracellular and extracellular fluid in alcoholism: evidence from diffusion tensor imaging. Neuropsychopharmacology. 2005b;30:423–432. doi: 10.1038/sj.npp.1300623. [DOI] [PubMed] [Google Scholar]
- Pfefferbaum A, Sullivan EV. Increased brain white matter diffusivity in normal adult aging: relationship to anisotropy and partial voluming. Magn Reson Med. 2003;49:953–961. doi: 10.1002/mrm.10452. [DOI] [PubMed] [Google Scholar]
- Pfefferbaum A, Sullivan EV, Hedehus M, Adalsteinsson E, Lim KO, Moseley M. In vivo detection and functional correlates of white matter microstructural disruption in chronic alcoholism. Alcohol: Clin Exp Res. 2000;24(8):1214–1221. [PubMed] [Google Scholar]
- Pfefferbaum A, Sullivan EV, Hedehus M, Moseley M, Lim KO. Brain gray and white matter transverse relaxation time in schizophrenia. Schizophr Res: Neuroimag Sect. 1999;91:93–100. doi: 10.1016/s0925-4927(99)00023-2. [DOI] [PubMed] [Google Scholar]
- Pfeuffer J, Van de Moortele PF, Ugurbil K, Hu X, Glover GH. Correction of physiologically induced global off-resonance effects in dynamic echo-planar and spiral functional imaging. Magn Reson Med. 2002;47(2):344–353. doi: 10.1002/mrm.10065. [DOI] [PubMed] [Google Scholar]
- Pierpaoli C, Basser PJ. Towards a quantitative assessment of diffusion anisotropy. Magn Reson Med. 1996;36:893–906. doi: 10.1002/mrm.1910360612. [DOI] [PubMed] [Google Scholar]
- Pujol J, Junque C, Vendrell P, Grau JM, Marti-Vilalta JL, Olive C, Gili J. Biological significance of iron-related magnetic resonance imaging changes in the brain. Arch Neurol. 1992;49(7):711–717. doi: 10.1001/archneur.1992.00530310053012. [DOI] [PubMed] [Google Scholar]
- Raz N, Rodrigue KM, Haacke EM. Brain aging and its modifiers: insights from in vivo neuromorphometry and susceptibility weighted imaging. Ann NY Acad Sci. 2007;1097:84–93. doi: 10.1196/annals.1379.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rohlfing T, Maurer CR. Nonrigid image registration in shared-memory multiprocessor environments with application to brains, breasts, and bees. IEEE Trans Inf Technol Biomed. 2003;7(1):16–25. doi: 10.1109/titb.2003.808506. [DOI] [PubMed] [Google Scholar]
- Rohlfing T, Zahr NM, Sullivan EV, Pfefferbaum A. The SRI24 multi-channel brain atlas: construction applications. In: Reinhardt JM, Pluim JPW, editors. Medical Imaging 2008: Image Processing; Proceedings of SPIE; Bellingham, WA: 2008. p. 691409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rumpel H, Ferrini B, Martin E. Lasting cytotoxic edema as an indicator of irreversible brain damage: a case of neonatal stroke. Am J Neuroradiol. 1998;19:1636–1638. [PMC free article] [PubMed] [Google Scholar]
- Salat DH, Tuch DS, Greve DN, van der Kouwe AJW, Hevelone ND, Zaleta AK, Rosen BR, Fischl B, Corkin S, Rosas HD, Dale AM. Age-related alterations in white matter microstructure measured by diffusion tensor imaging. Neurobiol Aging. 2005;26:1215–1227. doi: 10.1016/j.neurobiolaging.2004.09.017. [DOI] [PubMed] [Google Scholar]
- Sehy JV, Ackerman JJ, Neil JJ. Evidence that both fast and slow water ADC components arise from intracellular space. Magn Reson Med. 2002;48(5):765–770. doi: 10.1002/mrm.10301. [DOI] [PubMed] [Google Scholar]
- Silva MD, Omae T, Helme RKG, Li F, Fisher M, Sotak CH. Separating changes in the intra- and extracellular water apparent diffusion coefficient following focal cerebral ischemia in the rat brain. Magn Reson Med. 2002;48(5):826–837. doi: 10.1002/mrm.10296. [DOI] [PubMed] [Google Scholar]
- Snook L, Plewes C, Beaulieu C. Voxel based versus region of interest analysis in diffusion tensor imaging of neurodevelopment. NeuroImage. 2007;34(1):243–252. doi: 10.1016/j.neuroimage.2006.07.021. [DOI] [PubMed] [Google Scholar]
- Song SK, Sun SW, Ju WK, Lin SJ, Cross AH, Neufeld AH. Diffusion tensor imaging detects and differentiates axon and myelin degeneration in mouse optic nerve after retinal ischemia. NeuroImage. 2003;20(3):1714–1722. doi: 10.1016/j.neuroimage.2003.07.005. [DOI] [PubMed] [Google Scholar]
- Song SK, Sun SW, Ramsbottom MJ, Chang C, Russell J, Cross AH. Dysmyelination revealed through MRI as increased radial (but unchanged axial) diffusion of water. NeuroImage. 2002;17(3):1429–1436. doi: 10.1006/nimg.2002.1267. [DOI] [PubMed] [Google Scholar]
- Srinivasan A, Goyal M, Al Azri F, Lum C. State-of-the-art imaging of acute stroke. Radiographics. 2006;26(Suppl 1):S75–95. doi: 10.1148/rg.26si065501. [DOI] [PubMed] [Google Scholar]
- Sullivan EV, Adalsteinsson E, Hedehus M, Ju C, Moseley M, Lim KO, Pfefferbaum A. Equivalent disruption of regional white matter microstructure in aging healthy men and women. Neuroreport. 2001;12(22):99–104. doi: 10.1097/00001756-200101220-00027. [DOI] [PubMed] [Google Scholar]
- Sullivan EV, Adalsteinsson E, Pfefferbaum A. Selective age-related degradation of anterior callosal fiber bundles quantified in vivo with fiber tracking. Cereb Cortex. 2006;16(7):1030–1039. doi: 10.1093/cercor/bhj045. [DOI] [PubMed] [Google Scholar]
- Sullivan EV, Pfefferbaum A. Diffusion tensor imaging and aging. Neurosci Biobehav Rev. 2006;30(6):749–761. doi: 10.1016/j.neubiorev.2006.06.002. [DOI] [PubMed] [Google Scholar]
- Sullivan EV, Rohlfing T, Pfefferbaum A. Quantitative fiber tracking of lateral and interhemispheric white matter systems in normal aging: relations to timed performance. Neurobiol Aging. 2008 doi: 10.1016/j.neurobiolaging.2008.04.007. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas LO, Boyko OB, Anthony DC, Burger PC. MR detection of brain iron. AJNR Am J Neuroradiol. 1993;14(5):1043–1048. [PMC free article] [PubMed] [Google Scholar]
- Wang C, Stebbins GT, Nyenhuis DL, deToledo-Morrell L, Freels S, Gencheva E, Pedelty L, Sripathirathan K, Moseley ME, Turner DA, Gabrieli JD, Gorelick PB. Longitudinal changes in white matter following ischemic stroke: a three-year follow-up study. Neurobiol Aging. 2006;27(12):1827–1833. doi: 10.1016/j.neurobiolaging.2005.10.008. [DOI] [PubMed] [Google Scholar]