

Abstract
Elimination of tuberculosis (TB) largely depends upon definitive rapid diagnosis and treatment. Widely used diagnostic tests do not qualify for use in a developing country due to lack of either desired accuracy or their cost. In the present study an enzyme-linked immunosorbent assay was used to evaluate the diagnostic potential of an immuno-dominant 30/32-kDa mycolyl transferase complex (Ag85 complex) and Mycobacterium tuberculosis-specific proteins (ESAT-6 and CFP-10) of the RD1 region. Higher sensitivity (84.1%) with Ag85 complex was observed compared with ESAT-6 (64.9%) and CFP-10 (66%), with almost similar specificity (Ag85: 85.2%, ESAT-6: 88.9%, CFP-10: 85.2%), whereas the individual components of Ag85 complex, i.e. Ag85A, Ag85B, and Ag85C, showed sensitivities of 44.6, 34, and 80.9% and specificities of 55.6, 74.1, and 40.7% respectively. A cocktail of Ag85 complex, ESAT-6, CFP-10, Ag85A, Ag85B, and Ag85C antigens also could not help in increasing either sensitivity (51.1%) or specificity (85.2%). Furthermore, immunoblot analysis using clinical isolates as well as a standard strain (H37Rv) of M. tuberculosis also showed strong reactivity of sera from TB patients to Ag85 complex and, to a lesser extent, also to ESAT-6. To conclude, use of Ag85 complex along with ESAT-6 and CFP-10 seems to be promising in minimizing the heterogeneous sero-responses of adult TB cases.
Keywords: Secretory proteins, 30/32-kDa Mycolyl transferase complex, ESAT-6/CFP-10, Serodiagnosis, Tuberculosis
Introduction
Tuberculosis (TB) is a growing health problem in the developing world. Nine million new cases of TB and three million deaths are reported every year around the globe (WHO 2007). In asymptomatic individuals, TB infection can be diagnosed with the help of the intra-dermal skin test using purified protein derivative (PPD). It is impossible to distinguish between present and past infection on the basis of a PPD test and this test is also not specific in BCG-vaccinated individuals (Huebner et al. 1993). Furthermore, conversion of the PPD reactivity from negative to positive challenges the clinician (Fine et al. 1999).
The specificity of acid-fast bacilli (AFB) staining is typically >99% and the sensitivity of this test varies between 25 and 75% (American Thoracic Society 2000). A positive AFB test and negative culture result may be caused by nonviable bacilli, when a person is receiving anti-TB drugs. The culture technique has an advantage in distinguishing the morphology of Mycobacterium tuberculosis from those of some non-tuberculous mycobacteria. However, it requires more time for results (3–4 weeks) (Thornton et al. 1998). Advanced molecular techniques such as PCR and BACTEC are, although rapid, sensitive, and specific (Hawkinds et al. 1991; Noordhoek et al. 1994), are not available everywhere and are also not cost effective.
Active pulmonary TB is contagious and leads to the spread of M. tuberculosis. Therefore, rapid and early diagnosis is needed for a TB control program. Various studies have been undertaken to develop a serodiagnostic test using secretory proteins of M. tuberculosis as these are known as immuno-dominant and early markers for TB (Kanaujia et al. 2003; Malen et al. 2008; Samanich et al. 2000). Among these proteins, ESAT-6 (Rv3875) and CFP-10 (Rv3874) are known as M. tuberculosis-specific antigens as they are absent in BCG and in most of the nonpathogenic mycobacteria (Arend et al. 2000; Harboe et al. 1996); therefore these antigens could differentiate infected and BCG-vaccinated individuals with TB. The role of these antigens in the early diagnosis and latent TB was previously reported (Silva et al. 2003). A well-defined fraction of the secretory proteins is Ag85 complex (30/32-kDa mycolyl transferases) abundantly found in culture filtrate of M. tuberculosis (Wiker and Harboe 1992). This is a family of three closely related proteins (Ag85A, Ag85B, and Ag85C) found in all mycobacteria (Wiker et al. 1986). Ag85 complex was recently reported to be an immuno-dominant marker for TB (Malen et al. 2008).
In our recent report we showed the potential of Ag85C in childhood TB cases (Kumar et al. 2008). The present study was conducted to find out the diagnostic potential of immuno-dominant Ag85 complex as well as its individual components and highly specific (ESAT-6, CFP-10) secretory antigens using enzyme-linked immunosorbent assay (ELISA) for the serodiagnosis of adult TB in a tuberculosis and leprosy endemic country, India. Furthermore, the study was also extended using a cocktail of all these antigens. The reactivity of sera was also evaluated using immunoblot analysis.
Materials and Methods
Study Subjects
Serum samples of 86 confirmed active TB patients were obtained from the State Tuberculosis Demonstration Centre, Agra, and the Department of Tuberculosis and Chest Diseases, S. N. Medical College, Agra. The patients were from the following well-defined categories.
Of the total pulmonary TB cases (n = 76), fresh TB cases (n = 48) comprised patients who had infection with M. tuberculosis bacilli for the first time and had no history of TB treatment. Defaulter TB cases (n = 20) comprised TB patients who had not taken the complete course of antitubercular treatment and the symptoms reoccurred. Relapse TB cases (n = 8) comprised TB patients who were treated earlier for TB but symptoms reoccurred after completion of the treatment (Table 1). All these patients were examined microbiologically (Ziehl Nielsen staining/culture) with clinical evaluations. All the pulmonary TB patients were smear positive for AFB. Extra-pulmonary TB patients (n = 10) had TB involvement in organs other than the lungs in which the disease was confirmed by tuberculin skin test and suggestive clinical findings (Table 1). The history of tuberculosis contact was available for all the TB patients. Bacteriologically negative, partially or fully treated cases of TB (n = 10) were also included in this study. All patients were categorized according to the guideline of the Revised National Tuberculosis Control Program, Central Tuberculosis Division, Government of India.
Table 1.
Details of different groups of patient with tuberculosis and control subjects studied in this study
| Study subjects | Total No. | Sex | Mean age | ||
|---|---|---|---|---|---|
| Male | Female | Year | Range | ||
| 1. Tuberculosis | 86 | 53 | 33 | ||
| i. Pulmonary cases | 48 | 32 | 16 | 32.6 ± 13.2 | 18–65 |
| ii. Extra-pulmonary cases | 10 | 3 | 7 | 31.0 ± 14.0 | 18–65 |
| iii. Defaulter TB cases | 20 | 10 | 10 | 28.7 ± 8.8 | 18–50 |
| iv. Relapse TB cases | 8 | 8 | 0 | 32.0 ± 10.8 | 24–55 |
| 2. Healthy controls | 29 | 21 | 8 | 25.9 ± 6.7 | 22–52 |
| 3. Leprosy controls | 18 | 12 | 6 | 33.3 ± 6.8 | 25–60 |
| 4. Anti tuberculosis treated cases | 10 | 8 | 2 | 35.3 ± 12.8 | 18–58 |
Healthy controls (n = 29) included in this study were staff members or temporary students working in our Institute who had no disease involvement or any family contact history with TB/leprosy patients. Eighteen well-confirmed leprosy controls (7 lepromatous, 8 borderline tuberculoid, 2 mid-borderline, and 1 tuberculoid leprosy) were also included. These leprosy cases were taken from the outpatient department (OPD) of the National JALMA Institute for Leprosy and Other Mycobacterial Diseases (ICMR), Agra. Two untreated active TB patients, two healthy BCG-vaccinated individuals, and one contact of TB were selected for Western blot analysis on the basis of their antibody response shown in ELISA.
All the patients and controls included in the present study were over 18 and under 65 years of age. Human immunodeficiency virus (HIV) testing was performed for all the cases and HIV-positive cases were excluded from the study. The study was approved by the institutional ethics committee and informed consent was obtained from each patient.
Antigen and Antibody
Purified r-ESAT-6, r-CFP-10, native Ag85 complex, its individual components (Ag85A, Ag85B, Ag85C), monoclonal antibody (CS-90) to Ag85 complex, and polyclonal antibody to ESAT-6 and CFP-10 were obtained from Colorado State University (Colorado, USA) through a TB Research Materials and Vaccine Testing Contract (NIH Contract HHSN266200400091C/ADB Contract NOI-AI-40091).
Enzyme-Linked Immunosorbent Assay
One hundred μl of the antigens ESAT-6 (12.5 ng/ml), CFP-10 (12.5 ng/ml), Ag85 complex (12.5 ng/ml), Ag85A (50 ng/ml), Ag85B (6.25 ng/ml), Ag85C (25 ng/ml), and a cocktail of all these antigens having the same concentration as described above was coated on a 96-well ELISA plate (flat bottom, Nunc Maxisorp, Roskilde, Denmark) in 0.05 M carbonate bicarbonate buffer, pH 9.6. The working dilution of each antigen was standardized using checkerboard titration as described earlier (Kumar et al. 2008). After overnight incubation at 37°C, the contents of all the wells were aspirated and the plate was washed three times with 150 mM PBS, pH 7.4. The plate was blocked with 2% BSA (Sigma, St. Louis, MO, USA) in PBS (200 μl/well) for 1 h. After washing with PBST (with 0.05% Tween-20), serum diluted 1:200 in assay diluent (1% BSA in PBS with 0.05% Tween-20) was added in duplicate wells and incubated at 37°C for 2 h. Anti-human IgG peroxidase-conjugated antibody (Sigma, St. Louis, MO, USA) diluted 1:10,000 in assay diluent was added to each well. After incubation for 1 h, 100 μl of substrate solution (orthophenylene diamine (OPD) tablet of 5 mg dissolved in 10 ml of distilled water and 50 μl H2O2) was added to each well and kept at room temperature for 20 min in the dark. The reaction was stopped by adding 50 μl of 7% H2SO4 (stop solution) to each well and the absorbance was measured at 492 nM using a Spectramax-M2 Reader (Molecular Devices, Sunnyvale, CA, USA). All the experiments were carried out at least two times.
Preparation of Whole-Cell Extracts
The growth of M. tuberculosis culture was harvested at the late log phase (3 weeks) from Sauton’s medium and washed twice with PBS (150 mM, pH 7.4). Bacterial growth was then re-suspended (0.2 g growth/ml) in lysis buffer (50 mM Tris, 10 mM MgCl2, 1 mM EGTA, 1 mM PMSF, pH 7.4) and subjected to sub-cellular fractionation (Brodie et al. 1979) using a Vibra-Cell probe ultrasonicator for a total of 20 min using 50% output control (100% = 475 W) and 50% duty cycle (on/off) at 4°C. The extracts were then centrifuged at 23,000×g for 30 min to remove debris and the supernatant was collected. The protein concentration of each sample was determined using Bradford’s method (Bradford 1976). These extracts were stored at –20°C until used.
Immunoblot Analysis
Polyacrylamide gel electrophoresis (PAGE) was done under reducing conditions (Laemmli 1970) using 12% (w/v) resolving gel in a Mini-Protean gel apparatus (Bio-Rad Laboratories, Hercules, CA, USA) by loading 20 μg/lane of protein extract. The molecular mass marker was obtained from Bio-Rad. The resolved proteins were transferred (Towbin et al. 1979) onto a nitrocellulose membrane (0.45-μm pore size; Sigma, St. Louis, MO, USA) using a TransBlot apparatus (Bio-Rad Laboratories, Hercules, CA, USA). The membrane was washed with PBS and blocked with 1% BSA (Sigma, St. Louis, MO, USA). The membrane was probed with sera diluted 1:400 in assay diluent (1% BSA in PBS containing 0.05% Tween-20) overnight at 4°C. The blots were washed with PBS containing 0.05% Tween-20 and incubated with anti-human IgG peroxidase-conjugated antibody (Sigma, St. Louis, MO, USA) diluted 1:5,000 in assay diluent for 1 h. After final washing, the color was developed with diamino benzedine (Sigma, St. Louis, MO, USA) in citrate phosphate buffer (0.5 M, pH 5). When reactivity was observed, the reaction was stopped by rinsing the membrane with distilled water. Data were analyzed by the Gel Documentation system (Bio-Rad Laboratories, Segrate (Milan), Italy) using Quantity One Software.
Statistical Analysis
Receiver operating characteristic (ROC) curves were constructed using Software Stata-7 (Strata Corporation, College Station, Texas, USA) to determine the cutoff, sensitivity, and specificity. Scattergrams were plotted using GraphPad prism software version 3.02 (San Diego, CA, USA).
Results
The focus of the present study was to identify the serodiagnostic potential of various secretory antigens of M. tuberculosis (ESAT-6, CFP-10, Ag85 complex as well as its individual components (A, B, C), and a cocktail of all these antigens) using ELISA. The antibody response to each antigen was analyzed in 76 well-characterized patients with active pulmonary TB (48 fresh TB, 20 defaulters, 8 relapsed TB), 10 extra-pulmonary TB, 10 partially/fully treated smear-negative cases of TB, 29 healthy controls, and 18 leprosy controls. Furthermore, the reactivity of sera against clinical isolates of M. tuberculosis was also evaluated by immunoblotting.
Sensitivity and specificity for a particular antigen were determined using ROC analysis. For all the antigens, the same numbers of TB cases, healthy individuals, and leprosy patients were taken. The cutoff was selected at the point which showed the best accuracy (correctly classifying individuals to their groups), sensitivity, and specificity by ROC. A predictive value (PV) to define the probability of a disease is very important as it provides the significance of a disease to characterize a patient for the particular disease from the patient’s population (+PV), and a high −PV is also needed to exclude the disease (Table 2); hence these parameters were also analyzed. Furthermore, concordance of IgG response to antigens that show good results in terms of sensitivity, specificity, and accuracy was determined to assess the chance of agreement (Table 3).
Table 2.
Immunoglobulin G reactivity against various secretory antigens of M. tuberculosis at cut off decided by ROC analysis
| Antigen | Cutoff | Sensitivity (%) | Specificity (%) | +PV (%) | −PV (%) | +LR | −LR | ROC area |
|---|---|---|---|---|---|---|---|---|
| ESAT-6 | 0.331 | 64.9 | 88.9 | 95.3 | 42.1 | 5.84 | 0.39 | 0.743 |
| CFP-10 | 0.293 | 66.0 | 85.2 | 93.9 | 41.8 | 4.45 | 0.40 | 0.746 |
| Ag85 complex | 0.150 | 84.1 | 85.2 | 95.2 | 60.5 | 5.67 | 0.19 | 0.879 |
| Ag85A | 0.119 | 44.6 | 55.6 | 77.4 | 23.2 | 1.01 | 0.10 | 0.496 |
| Ag85B | 0.163 | 34.0 | 74.1 | 82.8 | 24.4 | 1.31 | 0.89 | 0.545 |
| Ag85C | 0.072 | 80.9 | 40.7 | 82.6 | 37.9 | 1.36 | 0.47 | 0.603 |
| Cocktail | 0.114 | 51.1 | 85.2 | 92.3 | 33.3 | 3.45 | 0.57 | 0.685 |
Table 3.
Correlation of antibody reactivity of various secretory antigens
| ESAT-6–CFP-10+/Total patients | CFP-10–ESAT-6+/Total patients | Ag85–ESAT-6+/Total patients | Ag85–CFP-10+/Total patients | Ag85–Ag85A+/Total patients | Ag85–Ag85B+/Total patients | Ag85–Ag85C+/Total patients | Ag85–Cocktail+/Total patients |
|---|---|---|---|---|---|---|---|
| 7/76 | 7/76 | 1/76 | Nil/76 | 5/76 | 4/76 | 7/76 | 4/76 |
Antibody Response of Different Groups of TB Patients and Controls to the Various Secretory Antigens
We observed antibody reactivity of 64.9% and specificity of 88.9% for ESAT-6 using the cutoff determined by ROC (Table 2). Positivity at this cutoff was 68.8% for fresh TB, 70% for defaulters, 62.5% for relapse, and 70% for extra-pulmonary TB. None of the leprosy patients were positive for this antigen, whereas only 30% of the treated TB cases were found to be positive for it (Fig. 1).
Fig. 1.
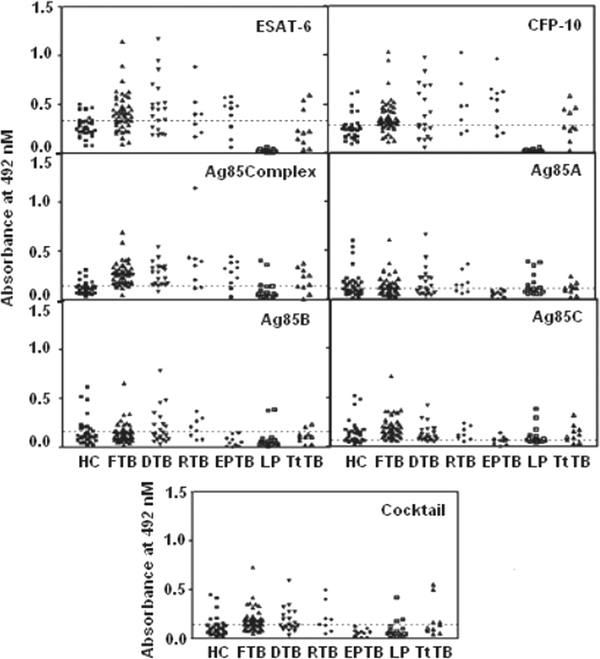
Antibody reactivity to various secretory antigens of M. tuberculosis with sera derived from different clinical groups (FTB: fresh untreated, DTB: defaulters, RTB: relapsed, EPTB: extra-pulmonary TB) of tuberculosis patients and controls (HC: healthy controls, LP: leprosy patients). TtTB denotes the treated cases of tuberculosis. Each scatter represents a tested sample of serum, the dotted line denotes the cutoff decided by ROC
We noted slightly increased reactivity to CFP-10 antigen in comparison with ESAT-6 when sera from fresh TB (72.8%), relapse cases (75%), and treated cases (40%) were measured. But this antigen was less reactive (55%) with the sera of defaulter cases of TB. Reactivity of extra-pulmonary TB was found to be similar (70%) to that of ESAT-6. We did not note any reactivity of the sera of leprosy patients to this antigen (Fig. 1). The overall sensitivity to this antigen was 66.0% and specificity 85.2% (Table 2).
The reactivity for Ag85 complex was the highest (84.1%) of all the antigens used in this study, with a specificity of 85.2% (Table 2). We observed 88.9% reactivity in the fresh TB group, 90% in defaulters, 75% in relapse cases, 80% in extra-pulmonary TB, 60% in treated cases of TB, and 6.89% in leprosy patients to this antigen (Fig. 1).
The reactivities to the individual components Ag85A, Ag85B, and Ag85C of Ag85 complex were 44.6, 34.0, and 80.9% and specificity 55.6, 74.1, and 40.7%, respectively (Table 2). Furthermore, 36.6, 35.4, and 93.7% of fresh TB cases were reactive to the Ag85A, B, and C components respectively. The observed reactivities of sera from the defaulter cases were 55, 45, and 80%, respectively. We noted 62.5% reactivity with Ag85A, 62.5% with Ag85B, and 87.5% with Ag85C in the sera of relapse cases. None of the extra-pulmonary TB cases showed antibody response to Ag85A or Ag85B, although 30% reactivity was noted using Ag85C. The leprosy controls reacted more (44.4%) with the Ag85C component than to Ag85A (33.3%) and Ag85B (5.5%). Reactivities in the treated TB cases were only 40, 20, and 70% for Ag85A, Ag85B, and Ag85C, respectively (Fig. 1).
We observed lower reactivity (51.1%) using the cocktail of all these antigens, but the specificity was the same (85.2%) as that reported for the Ag85 complex (85.2%) (Table 2). However, antibody reactivity in fresh, relapse, and defaulter TB cases, treated cases of TB, and leprosy cases was observed to be 64.6, 75, 50, 40, and 16.6%, respectively. No reactivity was noted with this antigen in the extra-pulmonary TB patients (Fig. 1). Interestingly, four of the healthy individuals were reactive to all the antigens used in this study.
Correlation of Reactivity of Individual Patients to Various Secretory Antigens
The antibody responses to the various secretory antigens were also correlated. We noted antibody response to ESAT-6 in seven patients who did not show antibody to CFP-10 and, similarly, antibody reactivity to CFP-10 was noted in seven patients who were negative for ESAT-6. All the patients negative for Ag85 complex were also negative for CFP-10 and ESAT-6, except one patient who was positive for ESAT-6. However, five patients who showed antibody reactivity to Ag85A were not positive for Ag85 complex, and 4 patients who were negative for Ag85 complex were positive for both Ag85B and the cocktail. Using Ag85C, we were able to detect seven more positive patients who were negative for Ag85 complex (Table 3). Furthermore, the areas under the ROC curves were compared for all these antigens (Table 2 and Fig. 2); it was highest for Ag85 complex (0.879), followed by CFP-10 (0.746) and ESAT-6 (0.743).
Fig. 2.
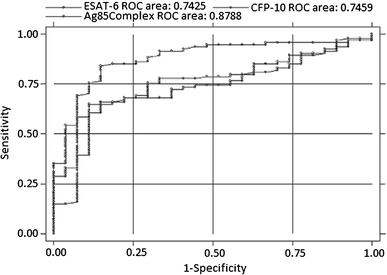
Area under ROC of three significant antigens with sera from tuberculosis patients
Sero-reactivity of Ag85 Complex, ESAT-6, and CFP-10 by Employing Western Blotting with H37Rv and Clinical Isolates of M. tuberculosis
We further analyzed the antibody reactivity to 39 clinical isolates from different regions of India using the pooled sera from fresh TB patients. Reactivity to the Ag85 complex was observed in almost all the isolates as well as in H37Rv (data not shown). Finally, five clinical isolates were randomly selected from all these clinical isolates, taking their reactivity and growth profile into consideration. Using these isolates, the seroreactivities of the individual sera were evaluated (Fig. 3).
Fig. 3.
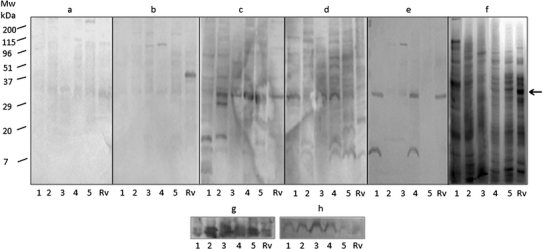
Lanes 1–5 of whole-cell extracts (WCEs) of clinical isolates of M. tuberculosis. a, b Blotting with BCG-vaccinated healthy individual’s sera. c–d Reactivity with individual tuberculosis patient’s sera. e Reactivity pattern of contact with tuberculosis. f 12% SDS–PAGE gel pattern of WCEs of clinical isolates and H37Rv. g Hybridization of polyclonal antibody against ESAT-6 and CFP-10 with whole-cell extracts. h Hybridization of monoclonal antibody (CS-90) against Ag85 complex with WCEs of clinical isolates and H37Rv. Rv: M. tuberculosis laboratory reference strain H37Rv; arrow shows the reactivity pattern of Ag85 complex
Significant reactivity at the 30-kDa region (Ag85 complex) in H37Rv as well as in all five clinical isolates was observed in both the TB patients used in the study (Fig. 3c, d) compared with two BCG-vaccinated individuals (Fig. 3a, b). However, only one patient was reactive to the 6-kDa region (ESAT-6) in 4 clinical isolates and to the 10-kDa region (CFP-10) in 3 isolates, but the reactivity was low (Fig. 3c, d). Interestingly, one person with contact with TB exhibited reactivity to the 30-kDa region (Ag85 complex) in H37Rv and in 2 clinical isolates and to the 6-kDa region (ESAT-6) in two clinical isolates (Fig. 3e). The reactivity to Ag85 complex in the clinical isolates was observed to be slightly greater than that in H37Rv when all these strains were hybridized with antibody against Ag85 complex (Fig. 3h). Reactivity to ESAT-6 and CFP-10 in the clinical isolates was shown after treating the transferred proteins with anti-ESAT-6 and -CFP-10 antibody (Fig. 3g).
Discussion
The management of tuberculosis in the regions of developing countries urgently needs an efficient diagnostic test that should be simple to perform with limited cost. Extensive studies have been done using various antigens of M. tuberculosis (Abebe et al. 2007; Mishra et al. 2008). The antigens selected for the current study have been reported to be strong targets for humoral and cell-mediated immune response (Barnes 2004; Kanaujia et al. 2003; Kanaujia et al. 2004; Mori et al. 2004; Samanich et al. 2001; Van Vooren et al. 1992).
Studies using ESAT-6 suggested its role as a marker of risk (Davidow et al. 2005; Silva et al. 2003) and progressive TB (Demissie et al. 2006). Berthet et al. reported a highly specific antigen present in the RD1 region, and CFP-10 is found in the same strongly stimulatory fraction of culture filtrate as ESAT-6, with limited distribution outside the TB complex (Berthet et al. 1998). In our study, very little difference between ESAT-6 (64.9%) and CFP-10 (66.0%) reactivity was observed. This finding was in support of a study in which some relatedness between ESAT-6 and CFP-10 at the amino-acid level was reported (Skjot et al. 2000). Interaction between ESAT-6 and CFP-10 genes and the co-transcription of these antigens were also demonstrated (Berthet et al. 1998). However, in our study, antibody to ESAT-6 was observed in seven TB patients but not to CFP-10 and vice versa (Table 3). This finding suggests a sequence difference between CFP-10 and ESAT-6.
India is a known endemic country for tuberculosis and leprosy and a study documented the presence of orthologues of ESAT-6 and CFP-10 in other mycobacteria, including M. leprae (Gey Van Pittius et al. 2001). Therefore, control sera from leprosy patients were also included in the present study to determine the cross-reactivity of antigens in leprosy and tuberculosis. To our surprise, none of the leprosy patients were found to have antibody response against either ESAT-6 or CFP-10. We noted reactivity in healthy individuals, due to which a higher cutoff was chosen for these two antigens, which could be one of the reasons for the low positivity among the leprosy patients. Genes of these proteins are reported to be present in other pathogenic mycobacteria, for example M. africanum, M. kansasii, M. marinum, M. szulgai (Harboe et al. 1996; Sorensen et al. 1995), and M. bovis (Harboe et al. 1996), as well as the slow-growing nonpathogenic mycobacterium M. gastri (Colangeli et al. 2000) and the fast-growing nonpathogenic environmental species M. flavesens (Harboe et al. 1996). All the healthy individuals were either staff or students working at our institute, hence higher reactivity to ESAT-6 could be due to the presence of cross-reactive antibodies to these mycobacteria. In our previous study we noted overall sensitivity and specificity of 53.01 and 91%, respectively, for ESAT-6 in pediatric TB patients (Dayal et al. 2006).
Although Ag85 complex is reported to be cross-reactive with other mycobacteria as well (Rinke de Wit et al. 1993), we observed the highest sensitivity (84.0%) and 85.2% specificity (Table 1) using Ag85 complex. In an earlier report, Ag85 complex was reported to be 72% sensitive and 100% specific in Mexican Totonac Indians with pulmonary tuberculosis (Sanchez-Rodriguez et al. 2002). Similar findings have been confirmed by others (Abdelnour et al. 2000). Furthermore, we observed high antibody (60%) against Ag85 complex in patients undergoing anti-TB treatment, which suggests persistence of this antigen for longer duration. A recent report also indicates the immuno-dominant nature of Ag85 complex (Malen et al. 2008).
The seroreactivity of Ag85A in our study was 44.68% in TB patients and it was only 55.66% specific. This antigen was also noted to be highly reactive with the sera of leprosy patients (33.33%). Antibody reactivity to Ag85B was only 33.0% and specificity 74.1% (Table 2). Antibody response to different components of the Ag85 complex was also reported earlier (Lim et al. 1999; Samanich et al. 2000; Van Vooren et al. 1992). Van Vooren et al. suggested that Ag85B was the most useful component of the Ag85 complex for the serodiagnosis of the active form of TB (Van Vooren et al. 1991). This was further supported by Lim et al., who showed that the antibody responses to Ag85B and Ag85A in patients were significantly greater than those to the Ag85C protein (Lim et al. 1999). Ag85B (Rv1886c) was reported to be 41–94% sensitive in various studies (Raja et al. 2002, 2005; Steingart et al. 2009; Uma Devi et al. 2003; Vikerfors et al. 1993). In contrast, McDonough et al. failed to detect antibodies against Ag85B (McDonough et al. 1992). We noted antibody to Ag85B in four more patients compared with the Ag85 complex (Table 2). However, antibodies to Ag85A and Ag85B have been reported to appear in extensive disease state (Sada et al. 1990). Ag85C has also been identified as an early marker for TB diagnosis; reactivity to this antigen has been reported in 36% of patients and it was reported to be 100% specific in HIV-positive smear-positive cases of TB (Samanich et al. 2000). We noted sensitivity of 80.9% and specificity of only 40.7% using this antigen. In contrast, Samanich et al., in their study on HIV-negative smear-positive TB patients, showed 80% reactivity to this antigen (Samanich et al. 2000). In our study, Ag85C was found to be highly reactive in leprosy patients (44.4%) among all the antigens. In a very recent study we reported strong reactivity with high specificity to Ag85C in pediatric TB patients (Kumar et al. 2008), which could be possible because of less exposure of pediatric patients to environmental mycobacteria. Homologues of Ag85 complex are also present in nonpathogenic mycobacteria (Rinke de Wit et al. 1993) and, this being an endemic country, it is possible that the controls taken for our study were already exposed and had antibody to these antigens. Despite the presence of cross-reactive epitope sequences, the stronger reactivity of the sera from TB patients to Ag85 complex suggests that this antigen also has some specific immuno-dominant sequences for the diagnosis of TB.
Further in our study, the antibody reactivity of two patients to all the clinical isolates is suggestive of similar expression of Ag85 complex genes and a relative immuno-dominant nature of this antigen complex. Reactivity to ESAT-6 in H37Rv and four clinical isolates was observed in one TB patient. Furthermore, a blot with a contact of TB also showed strong reactivity against Ag85 complex in H37Rv as well as two clinical isolates and with ESAT-6 in two clinical isolates (out of 5) (Fig. 3e). These results highlight the importance of Ag85 complex and ESAT-6 in minimizing the heterogeneous immune response of TB patients (Fig. 3c, d). Although the reactivity was analyzed with only one contact of TB, this finding indicates that exposure status could be identified using Ag85 complex and ESAT-6. Furthermore, studies taking more contacts of tuberculosis would be helpful for a strong conclusion. However, the reactivity with CFP-10 was low.
Several reports indicate that the pattern of antigen reactivity to various antibodies in serum varied greatly from patient to patient and no antigen alone can perform significantly for the diagnosis of TB (Abebe et al. 2007; Lyashchenko et al. 1998). Therefore, a cocktail of these antigens was made to minimize the possible heterogeneity of antigen recognition by patients. The lower sensitivity reported for the cocktail in the present study might be due to steric hindrance or masking of some of the dominant epitopes in the cocktail, as observed in our previous study of pediatric patients with TB (Kumar et al. 2008).
This is the first report from northern India to show a response to highly specific (ESAT-6, CFP-10) and immuno-dominant antigen 85 in complex form as well as its individual components in adult patients with tuberculosis. The strong antibody responses against Ag85 complex (despite the presence of cross-reacting epitopes in BCG and other non-tuberculous mycobacteria), ESAT-6, and CFP-10 in all the categories of active TB suggest the potential of these antigens. These antigens could be more useful in a high-prevalence area of TB to develop an array for serodiagnosis. Therefore, detailed epitope-based studies of these secretory antigens might be promising to provide specific sequences for the serodiagnosis of TB. Collectively, these results also indicate that an array of antigens would be more useful for developing a simple and inexpensive serodiagnostic test for the diagnosis of adult cases of tuberculosis than a mixture of various antigens.
Acknowledgments
We are grateful to Dr. John T. Belisle, Colorado State University, (Colorado, USA) for providing the recombinant and native antigens of M. tuberculosis used in the present study. We thank Dr. B. K. Girdhar, NJIL & OMD, Agra, for providing well-diagnosed serum samples of leprosy patients, Ms Rajni Upadhyay and Ms Bhavyata Dua, NJIL & OMD, for their help in preparing this manuscript. The authors acknowledge ICMR and DBT, New Delhi, for financial support and UGC-CSIR and ICMR, New Delhi, for providing fellowships to GK and PKD, respectively. Help given by Mr. M. M. Alam, Lab Technician, Immunology Department, NJIL & OMD, Agra, for sample collection and Mr. Navneet Kulshreshtha, Lab Technician, Microbiology & Molecular Biology Department, NJIL & OMD, for mycobacterial culture is also acknowledged.
References
- Abdelnour AM, Khaled G, Matar GM, et al. Evaluation of a serological test for the diagnosis of tuberculosis using the 38 kDa antigen. East J Medi. 2000;5:10–12. [Google Scholar]
- Abebe F, Holm-Hansen C, Wiker HG, et al. Progress in serodiagnosis of Mycobacterium tuberculosis infection. Scan J Immunol. 2007;66:176–191. doi: 10.1111/j.1365-3083.2007.01978.x. [DOI] [PubMed] [Google Scholar]
- American Thoracic Society Diagnostic standards and classification of tuberculosis in adults and children. Am J Respir Crit Care Med. 2000;161:1376–1395. doi: 10.1164/ajrccm.161.4.16141. [DOI] [PubMed] [Google Scholar]
- Arend SM, Andersen P, Meijgaarden KE, et al. Detection of active tuberculosis infection by T cell responses to early secreted antigen target 6 kDa protein and culture filtrate protein 10. J Infect Dis. 2000;181:1850–1854. doi: 10.1086/315448. [DOI] [PubMed] [Google Scholar]
- Barnes PF. Diagnosing latent tuberculosis infection, turning glitter to gold. Am J Respir Crit Care Med. 2004;170:5–6. doi: 10.1164/rccm.2404004. [DOI] [PubMed] [Google Scholar]
- Berthet FX, Rasmussen PB, Rosenkrands I, et al. A Mycobacterium tuberculosis operon encoding ESAT-6 and a novel low-molecular-mass culture filtrate protein (CFP-10) Microbiology. 1998;144:3195–3203. doi: 10.1099/00221287-144-11-3195. [DOI] [PubMed] [Google Scholar]
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- Brodie AF, Kalra VK, Lee SH, et al. Properties of energy-transducing systems in different types of membrane preparations from Mycobacterium phlei—preparation, resolution, and reconstitution. Methods Enzymol. 1979;55:175–199. doi: 10.1016/0076-6879(79)55024-1. [DOI] [PubMed] [Google Scholar]
- Colangeli R, Spencer JS, Bifani P, et al. MTSA-10, the product of the Rv3874 gene of Mycobacterium tuberculosis, elicits tuberculosis-specific, delayed-type hypersensitivity in guinea pigs. Infect Immun. 2000;68:990–993. doi: 10.1128/IAI.68.2.990-993.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davidow A, Kanaujia GV, Shi L, et al. Antibody profiles characteristic of M. tuberculosis infection state. Infect Immun. 2005;73:6846–6851. doi: 10.1128/IAI.73.10.6846-6851.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dayal R, Sirohi G, Singh MK, et al. Diagnostic value of ELISA serological tests in childhood tuberculosis. J Trop Pediatr. 2006;52:433–437. doi: 10.1093/tropej/fml047. [DOI] [PubMed] [Google Scholar]
- Demissie A, Leyten EM, Abebe M, et al. Recognition of stage-specific mycobacterial antigens differentiates between acute and latent infections with Mycobacterium tuberculosis. Clin Vaccine Immunol. 2006;13:179–186. doi: 10.1128/CVI.13.2.179-186.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fine PE, Bruce J, Ponnighaus JM, et al. Tuberculin sensitivity: conversions and reversions in a rural African population. Int J Tuberc Lung Dis. 1999;3:962–975. [PubMed] [Google Scholar]
- Gey Van Pittius NC, Gamieldien J, Hide W et al (2001) The ESAT-6 gene cluster of Mycobacterium tuberculosis and other high G + C Gram-positive bacteria. Genome Biol 2:research0044.1–0044.18 [DOI] [PMC free article] [PubMed]
- Harboe M, Oettinger T, Wiker HG, et al. Evidence for occurrence of the ESAT-6 protein in Mycobacterium tuberculosis and virulent Mycobacterium bovis and for its absence in Mycobacterium bovis BCG. Infect Immun. 1996;64:16–22. doi: 10.1128/iai.64.1.16-22.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawkinds JE et al (1991) Antibacterial susceptibility tests. In: Manual of clinical microbiology, 5 edn. American Society for Microbiology, Washington DC, pp 1138–1152
- Huebner RE, Schein MF, Bass JB., Jr The tuberculin skin test. Clin Infect Dis. 1993;17:968–975. doi: 10.1093/clinids/17.6.968. [DOI] [PubMed] [Google Scholar]
- Kanaujia GV, Garcia MA, Bouley DM, et al. Detection of early secretory antigenic target-6 antibody for diagnosis of tuberculosis in non-human primates. Comp Med. 2003;53:602–606. [PubMed] [Google Scholar]
- Kanaujia GV, Motzel S, Garcia MA, et al. Recognition of ESAT-6 sequences by antibody in sera of tuberculosis nonhuman primates. Clin Diag Lab Immunol. 2004;11:222–226. doi: 10.1128/CDLI.11.1.222-226.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar G, Dagur PK, Singh M, et al. Diagnostic potential of Ag85C in comparison to various secretory antigens for childhood tuberculosis. Scand J Immunol. 2008;68:177–183. doi: 10.1111/j.1365-3083.2008.02133.x. [DOI] [PubMed] [Google Scholar]
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- Lim JH, Park JK, Jo EK, et al. Purification and immunoreactivity of three components from the 30/32-kilodalton antigen 85 complex in Mycobacterium tuberculosis. Infect Immun. 1999;67:6187–6190. doi: 10.1128/iai.67.11.6187-6190.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyashchenko K, Colangeli R, Houde M, et al. Heterogeneous antibody responses in tuberculosis. Infect Immun. 1998;66:3936–3940. doi: 10.1128/iai.66.8.3936-3940.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malen H, Softeland T, Wiker HG. Antigen analysis of Mycobacterium tuberculosis H37Rv culture filtrate proteins. Scan J Immunol. 2008;67:245–252. doi: 10.1111/j.1365-3083.2007.02064.x. [DOI] [PubMed] [Google Scholar]
- McDonough JA, Sada DE, Sippola AA, et al. Microplate and dot immunoassays for the serodiagnosis of tuberculosis. J Lab Clin Med. 1992;120:318–322. [PubMed] [Google Scholar]
- Mishra KC, Chastellier C, Narayana Y, et al. Functional role of the PE domain and immunogenicity of the Mycobacterium tuberculosis triacylglycerol hydrolase LipY. Infect Immun. 2008;76:127–140. doi: 10.1128/IAI.00410-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mori T, Sakatani M, Yamagishi F, et al. Specific detection of tuberculosis infection: an interferon-gamma-based assay using new antigens. Am J Respir Crit Care Med. 2004;170:59–64. doi: 10.1164/rccm.200402-179OC. [DOI] [PubMed] [Google Scholar]
- Noordhoek GT, Kolk AH, Bjune G, et al. Sensitivity and specificity of polymerase chain reaction for detection of M. tuberculosis, a blind comparison study among seven laboratories. J Clin Microbiol. 1994;32:277–284. doi: 10.1128/jcm.32.2.277-284.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raja A, Uma Devi KR, Ramalingam B, et al. Immunoglobulin G, A, and M responses in serum and circulating immune complexes elicited by the 16-kilodalton antigen of Mycobacterium tuberculosis. Clin Diagn Lab Immunol. 2002;9:308–312. doi: 10.1128/CDLI.9.2.308-312.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raja A, Uma Devi KR, Ramalingam B, et al. Improved diagnosis of pulmonary tuberculosis by detection of free and immune complex-bound anti-30 kDa antibodies. Diagn Microbiol Infect Dis. 2005;50:253–259. doi: 10.1016/j.diagmicrobio.2004.08.010. [DOI] [PubMed] [Google Scholar]
- Rinke de Wit TF, Bekelie S, Osland A, et al. The Mycobacteriumleprae antigen 85 complex gene family: identification of the gene for the 85A, 85C, and related MPT51 proteins. Infect Immun. 1993;61:3642–3647. doi: 10.1128/iai.61.9.3642-3647.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sada E, Ferguson LE, Daniel TM. An ELISA for the serodiagnosis of tuberculosis using a 30,000-Da native antigen of Mycobacterium tuberculosis. J Infect Dis. 1990;162:928–931. doi: 10.1093/infdis/162.4.928. [DOI] [PubMed] [Google Scholar]
- Samanich KM, Keen MA, Vissa VD, et al. Serodiagnostic potential of culture filtrate antigens of Mycobacterium tuberculosis. Clin Diagn Lab Immunol. 2000;7:662–668. doi: 10.1128/cdli.7.4.662-668.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samanich K, Belisle JT, Laal S. Homogeneity of antibody responses in tuberculous patients. Infect Immun. 2001;69:4600–4609. doi: 10.1128/IAI.69.7.4600-4609.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez-Rodriguez C, Estrada-Chavez C, Garcia-Vigil J, et al. An IgG antibody response to the antigen 85 complex is associated with good outcome in Mexican Totonaca Indians with pulmonary tuberculosis. Int J Tuberc Lung Dis. 2002;6:706–712. [PubMed] [Google Scholar]
- Silva VM, Kanaujia G, Gennaro ML, et al. Factors associated with humoral response to ESAT-6, 38 kDa and 14 kDa in patients with a spectrum of tuberculosis. Int J Tuberc Lung Dis. 2003;7:478–484. [PubMed] [Google Scholar]
- Skjot RLV, Oettinger T, Rosenkrands I, et al. Comparative evaluation of low-molecular-mass proteins from Mycobacterium tuberculosis identifies members of the ESAT-6 family as immunodominant T-cell antigens. Infect Immun. 2000;68:214–220. doi: 10.1128/IAI.68.1.214-220.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sorensen AL, Nagai S, Houen G, et al. Purification and characterization of a low-molecular-mass T-cell antigen secreted by Mycobacterium tuberculosis. Infect Immun. 1995;63:1710–1717. doi: 10.1128/iai.63.5.1710-1717.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steingart KR, Dendukuri N, Henry M, et al. Performance of purified antigens for serodiagnosis of pulmonary tuberculosis: a meta-analysis. Clin Vaccine Immunol. 2009;16:260–276. doi: 10.1128/CVI.00355-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thornton CG, MacLellan KM, Brink TL, Jr, et al. In vitro comparison of NALC-NaOH, tween 80, and C18-carboxypropylbetaine for processing of specimens for recovery of mycobacteria. J Clin Microbiol. 1998;36:3558–3566. doi: 10.1128/jcm.36.12.3558-3566.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA. 1979;76:4350–4354. doi: 10.1073/pnas.76.9.4350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uma Devi KR, Ramalingam B, Raja A. Antibody response to Mycobacterium tuberculosis 30 and 16 kDa antigens in pulmonary tuberculosis with human immunodeficiency virus coinfection. Diagn Microbiol Infect Dis. 2003;46:205–209. doi: 10.1016/S0732-8893(03)00047-6. [DOI] [PubMed] [Google Scholar]
- Vooren JP, Drowart A, Cock M, et al. Humoral immune response of tuberculous patients against the three components of the Mycobacterium bovis BCG 85 complex separated by isoelectric focusing. J Clin Microbiol. 1991;29:2348–2350. doi: 10.1128/jcm.29.10.2348-2350.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vooren JP, Drowart A, Bruyn J, et al. Humoral responses against the 85A and 85B antigens of Mycobacterium bovis BCG in patients with leprosy and tuberculosis. J Clin Microbiol. 1992;30:1608–1610. doi: 10.1128/jcm.30.6.1608-1610.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vikerfors T, Olcen P, Wiker H, et al. Serological response in leprosy and tuberculosis patients to the 18-kDa antigen of Mycobacterium leprae and antigen 85B of Mycobacterium bovis BCG. Int J Lepr Other Mycobact Dis. 1993;61:571–580. [PubMed] [Google Scholar]
- Wiker HG, Harboe M. The antigen 85 complex, a major secretion product of Mycobacterium tuberculosis. Microbiol Rev. 1992;56:648–661. doi: 10.1128/mr.56.4.648-661.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiker HG, Harboe M, Nagai S, et al. MPB59, a widely cross-reacting protein of Mycobacterium bovis BCG. Int Arch Allergy Appl Immunol. 1986;81:307–314. doi: 10.1159/000234154. [DOI] [PubMed] [Google Scholar]
- World Health Organization, WHO Report (2007) Global tuberculosis control, surveillance, planning, financing. WHO, Geneva, 277 pp