

Abstract
In all mammals, the sensory epithelium for audition is located along the spiraling organ of Corti that resides within the conch shaped cochlea of the inner ear (fig 1). Hair cells in the developing cochlea, which are the mechanosensory cells of the auditory system, are aligned in one row of inner hair cells and three (in the base and mid-turns) to four (in the apical turn) rows of outer hair cells that span the length of the organ of Corti. Hair cells transduce sound-induced mechanical vibrations of the basilar membrane into neural impulses that the brain can interpret. Most cases of sensorineural hearing loss are caused by death or dysfunction of cochlear hair cells.
An increasingly essential tool in auditory research is the isolation and in vitro culture of the organ explant 1,2,9. Once isolated, the explants may be utilized in several ways to provide information regarding normative, anomalous, or therapeutic physiology. Gene expression, stereocilia motility, cell and molecular biology, as well as biological approaches for hair cell regeneration are examples of experimental applications of organ of Corti explants.
This protocol describes a method for the isolation and culture of the organ of Corti from neonatal mice. The accompanying video includes stepwise directions for the isolation of the temporal bone from mouse pups, and subsequent isolation of the cochlea, spiral ligament, and organ of Corti. Once isolated, the sensory epithelium can be plated and cultured in vitro in its entirety, or as a further dissected micro-isolate that lacks the spiral limbus and spiral ganglion neurons. Using this method, primary explants can be maintained for 7-10 days. As an example of the utility of this procedure, organ of Corti explants will be electroporated with an exogenous DsRed reporter gene. This method provides an improvement over other published methods because it provides reproducible, unambiguous, and stepwise directions for the isolation, microdissection, and primary culture of the organ of Corti.
Keywords: Neuroscience, Issue 36, hearing, mice, cochlea, organ of Corti, organotypic, culture, hair cell, stem cell, gene expression, in vitro
Protocol
Day 1. Sterilization and coating of glass coverslips.
Dry sterilize glass microscope coverslips in an autoclave.
Place the sterilized coverslips into two wells of a pre-sterilized four-well cell culture dish.
- Coat the coverslips with 1:1 poly-L-ornithine and laminin supplemented with 20% Fetal Bovine Serum (FBS) overnight at 4 °C.
- 400 μL poly-L-ornithine (0.01% solution stored at 4 °C)
- 400 μL laminin (50 μg/mL stock solution stored in aliquots at -20 °C)
- 200 μL FBS (stored in aliquots at -20 °C)
Heat-sterilize the dissection tools overnight in a 150°C incubator.
- Make culture medium containing 10% serum and 10 mg /mL ampicillin.
- 90 mL Dulbecco's Modified Eagle Medium
- 5 mL FBS (stored in aliquots at -20 °C)
- 5 mL horse serum (stored in aliquots at -20 °C)
- 10 μL ampicillin (10 mg/mL stock solution stored at 4 °C )
Day 2. Isolation of the organ of Corti.
- Sterilize the positive flow dissection hood.
- turn on UV light for 20 min
- spray all surfaces with 70% ethanol and wait 5 min before use.
Decapitate mouse pup (P4) at the base of the foramen magnum using operating scissors.
Briefly rinse the head in 10 cm dish containing 70% ethanol.
Remove the epidermis using a scalpel blade.
Open the cranium along the sagittal suture using a scalpel blade and then bisect the forebrain. Retain the caudal forebrain for further dissection.
Remove the forebrain, cerebellum and brainstem using blunt dissection.
Remove the temporal bones (fig 2A), dip them briefly in 70% ethanol, and transfer them to a 3 mm dish containing sterile HBSS.
Using forceps, remove the bulla and surrounding tissue from the petrous portion of the temporal bone.
Locate the conch shaped cochlea (fig 2B) and separate it from the vestibular system using forceps.
At this stage of development, the bony labyrinth is not completely calcified and is easily dissected using forceps. Remove the bony labyrinth of the cochlea by careful separation starting at the basal end and moving apically using forceps.
The spiral ligament and attached organ of Corti is coiled along the spiral of the modiolus (fig 2C). Carefully remove the organ of Corti by securing the spiral ligament at the hook region of the base using forceps and unwinding it as you move apically.
Starting at the base, remove the spiral ligament from the organ of Corti using #55 fine forceps (fig 2D). Micro-isolation of the organ of Corti sensory epithelium (Optional).
Remove the organ of Corti hook region of the base using two ½ cc Insulin Syringes with permanently attached U-100 28G½ needles as forceps.
Starting at the apex, remove the spiral limbus from the row of inner hair cells and proceed basally (fig 2E-F). Plating the organ of Corti explant
Remove the poly-L-ornithine/laminin/FBS solution from the culture wells and add 130 μl of culture medium.
Transfer the dissected organ of Corti to the coated glass coverslip in the culture well and orient the explant so that the cilia of the hair cells are pointing up.
Remove the medium in the culture well using a 200 μL pipette. Make sure that the basilar membrane makes solid contact with the coated glass coverslip.
Carefully add 130 μL of culture medium to the organ of Corti using a 200 mL pipette. Apply two drops onto the surface of the organ of Corti and then slowly add the remaining volume to the side of the coverslip. Make sure that the organ of Corti does not float in the culture media, but remains affixed to the coverslip.
Incubate overnight at 37 °C in the presence of 5% CO2.
Day 3. Electroporation of the reporter gene into cultured organ of Corti.
Remove the culture medium from the organ of Corti culture.
Add 130 μL H2O for 1 min and then remove using a 200 μL pipette.
Add 30 μL of DsRed reporter plasmid (2mg /mL H2O stored at -20 °C).
Advance the electrodes of the electroporator using a micromanipulator so that the anode and cathode are on either side of the culture.
- Generate a pulse (27V, 30 ms duration, 10 pulse trains) to electroporate the reporter gene into the organ of Corti explant culture.
- Optional: reverse the polarity of the pulse to ensure electroporation of the transgene on both the modiolar and spiral ligament sides of the explant.
Wait 5 min.
- Add 130 μL of the Fugene 6: DNA solution (3 parts Fugene to 2 parts DNA). This solution should be prepared prior to the start of the electroporation procedure (step 20) using a 3 mL round bottom polystyrene test tube in a laminar flow hood. To make this solution:
- Add 2.4 μL of Opti-MEM (stored at 4 °C) to the test tube.
- Add 0.6 μL of Fugene 6 reagent (stored at 4 °C) to the test tube. Make sure to add the Fugene directly to the Opti-MEM and avoid direct contact with the sides of the test tube.
- Vortex for 1 second.
- Incubate 5 min at room temperature.
- Add 2.0 μL DsRed reporter plasmid double stranded DNA (stored at -20 °C at 100 μg DNA/mL H2O aliquots).
- Vortex for 1 second.
- Incubate 15 min at room temperature.
- Add 200 μL of culture medium.
- Vortex for 1 second.
Incubate overnight at 37 °C in 5% CO2.
Add 2 mL culture medium to the culture well and incubate for 37 °C for up to 10 days.
Representative Results
We present a method for the isolation of the organ of Corti from a perinatal mouse. The procedure can be used for mice as young as embryonic day 16 up to about postnatal day 6, at which point the bony labyrinth becomes sufficiently calcified to render the dissection cumbersome. Once the organ of Corti is dissected, it may be plated and cultured either in its entirety (fig 3) or as micro-isolated sensory epithelium (fig 4). We have further presented a technique to express exogenous genes in the cultured organ of Corti. The organotypic culture is useful for many other types of study, such as the analysis of organ of Corti gene expression using RT-PCR or in situ hybridization, organ of Corti co-culture with spiral ganglion cells or exogenous stem cells using the micro-isolate3, or in vitro analysis of hair cell death and regeneration.
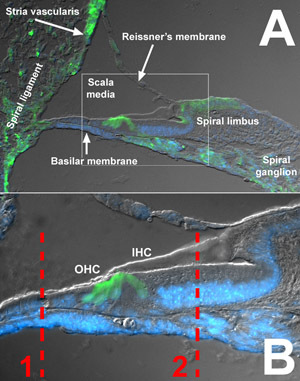 Figure 1. Cross section of the P4 murine organ of Corti. (A) A cross section from the basal turn of a cryosectioned cochlea obtained from a P4 mouse illustrates the general structures of the murine cochlea described in this protocol. The scala media is bordered by the spiral ligament and stria vascularis laterally, the Reissner s membrane superiorly, the spiral limbus medially, and the basilar membrane inferiorly. The box indicated the region expanded in B. (B) The organ of Corti is located on the superior side of the basilar membrane and includes one row of inner hair cells, three rows of outer hair cells, and their respective supporting cells. Dashed line 1 indicates the location along the basilar membrane that is removed during this organ of Corti dissection. Dashed line 2 indicates the location along the basilar membrane that is removed during the micro-isolation procedure. Green indicates immunohistochemical labeling of calbindin, which labels interdental cells of the spiral limbus, cochlear hair cells, spiral ganglion neurons, as well as cells of the spiral ligament and stria vascularis 7. DAPI labeling of nuclei is in blue. To see a larger version of this image please click here.
Figure 1. Cross section of the P4 murine organ of Corti. (A) A cross section from the basal turn of a cryosectioned cochlea obtained from a P4 mouse illustrates the general structures of the murine cochlea described in this protocol. The scala media is bordered by the spiral ligament and stria vascularis laterally, the Reissner s membrane superiorly, the spiral limbus medially, and the basilar membrane inferiorly. The box indicated the region expanded in B. (B) The organ of Corti is located on the superior side of the basilar membrane and includes one row of inner hair cells, three rows of outer hair cells, and their respective supporting cells. Dashed line 1 indicates the location along the basilar membrane that is removed during this organ of Corti dissection. Dashed line 2 indicates the location along the basilar membrane that is removed during the micro-isolation procedure. Green indicates immunohistochemical labeling of calbindin, which labels interdental cells of the spiral limbus, cochlear hair cells, spiral ganglion neurons, as well as cells of the spiral ligament and stria vascularis 7. DAPI labeling of nuclei is in blue. To see a larger version of this image please click here.
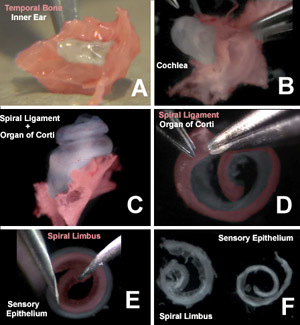 Figure 2. Organ of Corti dissection. Images from the accompanying video of the organ of Corti dissection highlight A) the cochlea and vestibular system located within the isolated temporal bone (red), B) the bony labyrinth of the cochlea, C) the spiral ligament and attached organ of Corti after removal of the bony labyrinth, D) removal of the spiral ligament and stria vascularis (red) from the organ of Corti, E) micro-isolation of the sensory epithelium from the spiral limbus (red), and F) the isolated spiral limbus (left) and sensory epithelium (right). To see a larger version of this image please click here.
Figure 2. Organ of Corti dissection. Images from the accompanying video of the organ of Corti dissection highlight A) the cochlea and vestibular system located within the isolated temporal bone (red), B) the bony labyrinth of the cochlea, C) the spiral ligament and attached organ of Corti after removal of the bony labyrinth, D) removal of the spiral ligament and stria vascularis (red) from the organ of Corti, E) micro-isolation of the sensory epithelium from the spiral limbus (red), and F) the isolated spiral limbus (left) and sensory epithelium (right). To see a larger version of this image please click here.
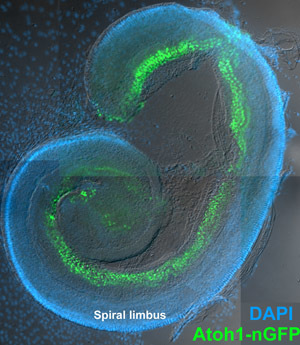 Figure 3. Cultured organ of Corti explant. DIC image showing the organ of Corti of a P4 Atoh1-nGFP mouse that was isolated, plated, and cultured for five days as described. This mouse has been genetically engineered so that cells that express the pro-hair cell gene Atonal homolog 1 (Atoh1 aka Math1) exhibit a green fluorescent protein that is localized to the nucleus8. The organs of Corti from these mice exhibit a nuclear GFP label in all hair cell nuclei and therefore allow for easy visualization of the sensory epithelium using an epifluorescent microscope. The relatively large spiral limbus can be seen lateral to the sensory epithelium. Mesenchymal cells that have originated from the organ of Corti have migrated away from the explant. Blue nuclear label is DAPI. To see a larger version of this image please click here.
Figure 3. Cultured organ of Corti explant. DIC image showing the organ of Corti of a P4 Atoh1-nGFP mouse that was isolated, plated, and cultured for five days as described. This mouse has been genetically engineered so that cells that express the pro-hair cell gene Atonal homolog 1 (Atoh1 aka Math1) exhibit a green fluorescent protein that is localized to the nucleus8. The organs of Corti from these mice exhibit a nuclear GFP label in all hair cell nuclei and therefore allow for easy visualization of the sensory epithelium using an epifluorescent microscope. The relatively large spiral limbus can be seen lateral to the sensory epithelium. Mesenchymal cells that have originated from the organ of Corti have migrated away from the explant. Blue nuclear label is DAPI. To see a larger version of this image please click here.
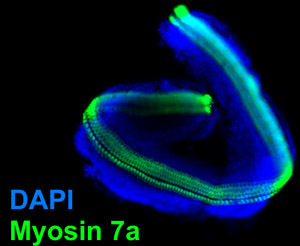 Figure 4. Micro-isolated sensory epithelium. Epifluorescent image obtained from the sensory epithelium that has been isolated from a P4 murine organ of Corti as described and cultured overnight. The micro-isolate was then fixed in 4% paraformaldehyde and processed for immunolabeling of Myosin 7a which labels cochlear hair cells. Note the absence of the comparatively larger spiral limbus from the figure 3. Blue nuclear label is DAPI. To see a larger version of this image please click here.
Figure 4. Micro-isolated sensory epithelium. Epifluorescent image obtained from the sensory epithelium that has been isolated from a P4 murine organ of Corti as described and cultured overnight. The micro-isolate was then fixed in 4% paraformaldehyde and processed for immunolabeling of Myosin 7a which labels cochlear hair cells. Note the absence of the comparatively larger spiral limbus from the figure 3. Blue nuclear label is DAPI. To see a larger version of this image please click here.
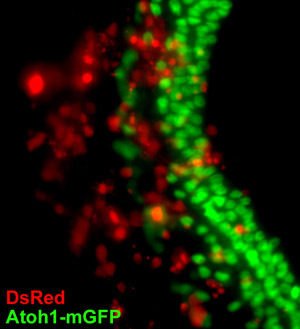 Figure 5. Electroporation of DsRed reporter gene into the cultured organ of Corti. Whole organs of Corti from P4 Atoh1-nGFP mouse pups were isolated, plated, and then electroporated with the DsRed reporter gene as described and then viewed under an epifluorescent microscope. Cells of the organ of Corti explant that expressed the transgenic DsRed reporter protein exhibit a red fluorescence and endogenous cells of the sensory epithelium exhibit a green nuclear fluorescence. Transgenic cells can be seen throughout the spiral limbus and sensory epithelium. To see a larger version of this image please click here.
Figure 5. Electroporation of DsRed reporter gene into the cultured organ of Corti. Whole organs of Corti from P4 Atoh1-nGFP mouse pups were isolated, plated, and then electroporated with the DsRed reporter gene as described and then viewed under an epifluorescent microscope. Cells of the organ of Corti explant that expressed the transgenic DsRed reporter protein exhibit a red fluorescence and endogenous cells of the sensory epithelium exhibit a green nuclear fluorescence. Transgenic cells can be seen throughout the spiral limbus and sensory epithelium. To see a larger version of this image please click here.
Discussion
There are several details that are critical for the success of this procedure. The shorter the time from temporal bone isolation to organ of Corti incubation, the greater the chance that the organs will attach to the coverslip and result in viable organ cultures. Therefore, it is important to limit the amount of time between dissection and placing the organs in the incubator. The choice of antibiotic is also crucial, since many aminoglycoside antibiotics are ototoxic and will result in hair cell death. Although it is preferable to forgo the use of antibiotics altogether, this leaves open the possibility for contamination. Therefore, we suggest the use of 10 μg /mL ampicillin as a general rule to overcome potential contamination problems.
The most troublesome aspect of this, and other procedures for the primary culture of the organ of Corti, is tendency of the organs to float off the plates during incubation. Although floating organ cultures can remain viable for 5-7 days, there are drawbacks of culturing floating organs. For instance, floating organ cultures often fold onto themselves after 4-5 days rendering microscopy problematic. Subsequently, the structural integrity of the organ can become compromised when compared to organs that have been affixed to the coverslip. We have found that the following techniques help to ensure that the organ of Corti does not float in the culture media, but remains affixed to the coverslip. First, coat the glass coverslips in 1:1 polyornithine/laminin supplemented with 20% FBS as described. The overnight incubation called for in this protocol is the minimum time that the plate should be coated. In our laboratory, we often coat all of the plates that we need for one week and keep them at 4 °C until the day prior to use when we move them to the incubator for an overnight incubation. Second, after the organs are transported to the coated coverslips, orient the explant so that the cilia of the hair cells face up. This orientation will facilitate the adherence of the basilar membrane to the culture dish. Third, remove the media within the well to affix the explant to the coated dish. This will ensure contact between the culture dish and the basilar membrane and enhance the ability of the explants to adhere to the glass. This will also assist in maintaining the structural integrity of the rows of hair cells. Lastly, carefully drip 2 drops of culture medium onto the surface of the organ of Corti using a 200 μL capacity pipette and then slowly fill the well by dripping the remaining volume (of the total 130 μL) on the side of the coverslip. It is important to work quickly to assure the attachment of the organ of Corti to the coated coverslip. From the initiation of the dissection to the incubation of the explants, it typically takes 10 minutes for a practiced operator to complete this organ of Corti isolation procedure.
In this protocol, we also present a method for the micro-isolation of the sensory epithelium from the spiral limbus of the organ of Corti. In this procedure, the spiral limbus is dissected away from the sensory epithelium using 28G½ insulin needles as dissection tools. The resulting micro-isolate consists of the rows of hair cells and their corresponding supporting cells (fig 3). The isolated sensory epithelium can then be cultured as described in this protocol. This micro-isolation procedure should be compared to the enzymatic separation of the sensory epithelia from surrounding tissue4. In mammals, as well as non-mammalian species such as chickens, thermolysin digestion of isolated vestibular organs results in the isolation of sensory epithelia from the basement mesenchymal cells4. In the rat cochlea, thermolysin digestion results in the separation of the greater epithelial ridge, lesser epithelial ridge and accompanying sensory epithelia from the basement membrane5. However, it is unclear whether the cochlear sensory epithelium can attach to the coated plates without the accompanying mesenchymal cells. While both the mechanical micro-isolation method and enzymatic digestion method result in the separation of the sensory epithelia from the spiral limbus, advantages of the micro-isolate dissection over the thermolysin digestion include a relatively shorter protocol, cheaper reagents, and potentially less stress to the explant due to enzymatic effects of the digestion. Additionally, the basement membrane is left intact in this approach, which may enhance attachment of the sensory epithelium to the culture plate. Disadvantages of this method include the need to develop the skills for this delicate dissection and potential mechanical damage to the sensory epithelium resulting from the micro dissection.
As an example of the utility of this procedure, we also present one example of the use of the organ of Corti cultures; the electroporation of exogenous genes into the explant culture. The electroporation procedure described above is based on previous methods of organ of Corti electroporation. Notably, Zheng and Gao (2000) describe the electroporation of isolated rat organs of Corti where the explants are held in place for electroporation by a molded groove of agarose and then plated on collagen coated 8-well LabTek slide in serum-free medium2. An advantage of their approach is that the organs are oriented so that the top surfaces of the explants face the cathode, which should theoretically result in an even distribution of electroporated cells across the explant. In our hands however, the method that we describe improved on this procedure because the organ of Corti remained affixed to the coverslip throughout the procedure thereby reducing the manipulation of the explant after electroporation. Additionally, a higher percentage of organs of Corti remained attached to the coverslips using this presented method. Our method is taken from Jones, et al. (2006) which uses the addition of the Fugene 6 reagent to increase the efficiency of gene expression after the electroporation. In the Jones et al. (2006) protocol, the organs are electroporated, incubated for 5 minutes with 100 μL of Fugene 6 transfection reagent, and plated 6. Our method differs in the use of a 3:2 ratio of Fugene 6 reagent to plasmid DNA, which we found empirically to provide optimal transgenic expression with minimum organ toxicity. We do not use undiluted Fugene 6 reagent, which can result in toxicity to the cultures. The electrode configuration in our protocol, as well as Jones et al. (2006), results in primary gene expression in either the spiral limbus or sensory epithelium depending upon the position of the cathode. Although there are DsRed positive cells on the side of the culture distant to the cathode, there is a higher concentration of transgenic cells closer to the cathode. To ensure a complete expression of the transgene in both sides of the organ of Corti explant, the current can be reversed for a second pulse train by simply inverting the leads. The presented protocol results in robust expression of the transgene throughout the organ of Corti (fig 5).
Acknowledgments
The authors would like to thank Demêmes Danielle and Douglas Cotanche and for their efforts in teaching us the methods for the isolation of the organ of Corti. Additionally, we would like to thank Ishmael Stefanov-Wagner for engineering the electroporator electrodes; Sherry Lin for her contributing artwork to the video animation; and Matthew Chana, Jason Meeker, and Kendra Marshall (www.goodfightproductions.com) for producing the video. This work was funded by grants (R03DC010065-Parker; RO1DC007174-Edge; P30DC05209- MEEI Core Support for Hearing Research) from the NIDCD.
References
- Sobkowicz HM, Bereman B, Rose JE. Organotypic Development of the Organ of Corti in Culture. Journal of Neurocytology. 1975;119(4):543–543. doi: 10.1007/BF01351537. [DOI] [PubMed] [Google Scholar]
- Zheng JL, Gao WQ. Overexpression of Math1 induces robust production of extra hair cells in postnatal rat inner ears. Nature Neuroscience. 2000;3(6):580–580. doi: 10.1038/75753. [DOI] [PubMed] [Google Scholar]
- Martinez-Monedero R. THE POTENTIAL ROLE OF ENDOGENOUS STEM CELLS IN REGENERATION OF THE INNER EAR. J Neurobiol. 2006;66(4):319–319. doi: 10.1016/j.heares.2006.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saffer LD, Gu R, Corwin JT. An RT-PCR analysis of mRNA for growth factor receptors in damaged and control sensory epithelia of rat utricles. Hearing Research. 1996;94(1,2):14–14. doi: 10.1016/0378-5955(95)00228-6. [DOI] [PubMed] [Google Scholar]
- Zhang Y. Isolation, growth and differentiation of hair cell progenitors from the newborn rat cochlear greater epithelial ridge. Journal of Neuroscience Methods. 2007;164(2):271–271. doi: 10.1016/j.jneumeth.2007.05.009. [DOI] [PubMed] [Google Scholar]
- Jones JM. Inhibitors of Differentiation and DNA Binding (Ids) Regulate Math1 and Hair Cell Formation during the Development of the Organ of Corti. J. Neurosci. 2006;26(2):550–550. doi: 10.1523/JNEUROSCI.3859-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daniela B, Josef S. Calbindin and S100 protein expression in the developing inner ear in mice. The Journal of Comparative Neurology. 2009;513(5):469–469. doi: 10.1002/cne.21967. [DOI] [PubMed] [Google Scholar]
- Helms AW. Autoregulation and multiple enhancers control Math1 expression in the developing nervous system. Development. 2000;127(6):1185–1185. doi: 10.1242/dev.127.6.1185. [DOI] [PubMed] [Google Scholar]
- Zheng JL, Wei-Qiang G. Differential Damage to Auditory Neurons and Hair Cells by Ototoxins and Neuroprotection by Specific Neurotrophins in Rat Cochlear Organotypic Cultures. European Journal of Neuroscience. 1996;8(9):1897–1897. doi: 10.1111/j.1460-9568.1996.tb01333.x. [DOI] [PubMed] [Google Scholar]