

Abstract
Chronic activation of the sympathetic nervous system (SNS) is a key component of cardiac hypertrophy and fibrosis. However, previous studies have provided evidence to also implicate inflammatory cells, including mast cells, in the development of cardiac fibrosis. The current study investigated the potential interaction of cardiac mast cells with the SNS. Eight week old male SHR were sympathectomized to establish the effect of the SNS on cardiac mast cell density, myocardial remodeling and cytokine production in the hypertensive heart. Age-matched WKY served as controls. Cardiac fibrosis and hypertension were significantly attenuated and left ventricular mass normalized while cardiac mast cell density was markedly increased in sympathectomized SHR. Sympathectomy normalized myocardial levels of IFN-γ, IL-6 and IL-10, but had no effect on IL-4. The effect of norepinephrine and substance P on isolated cardiac mast cell activation was investigated as potential mechanisms of interaction between the two. Only substance P elicited mast cell degranulation. Substance P was also shown to induce the production of angiotensin II by a mixed population of isolated cardiac inflammatory cells, including mast cells, lymphocytes and macrophages. These results demonstrate the ability of neuropeptides to regulate inflammatory cell function, providing a potential mechanism by which the SNS and afferent nerves may interact with inflammatory cells in the hypertensive heart.
Keywords: sympathectomy, fibrosis, mast cell, cytokines, inflammation, substance P, sympathetic nervous system
Introduction
Up-regulation of the sympathetic nervous system (SNS) is involved in numerous cardiovascular disease processes and is responsible for alterations in normal myocardial structure and function, including the induction of arrhythmias, alterations in contractile properties and cardiac remodeling. 1 In an experimental setting, there is evidence that the SNS mediates hypertension-induced cardiac fibrosis and hypertrophy through α- and β-adrenergic receptors, respectively. 2 However, concomitant with elevated SNS activity, induction of an inflammatory infiltrate into the heart also occurs, which appears to mediate the development of hypertension-induced cardiac fibrosis, but not hypertrophy. This inflammatory component includes mast cells (MC), 3-6 macrophages 7,8 and possibly T cells. 9,10 MCs maintain a close anatomical association with nerves, 11,12 with sympathetic neurons able to alter rat basophilic leukemia (RBL-2H3) MC membrane resistance. 13 Further, other inflammatory cells express adrenergic receptors, with modulation of cytokine production being either enhanced or repressed by stimulation of these receptors. 14 Alternatively, we have shown that cardiac MCs can also be activated by neuropeptides such as substance P, 15 which is found mainly in afferent sensory nerves. Given the prominent involvement of both the SNS and inflammation in myocardial remodeling associated with hypertension, we sought to determine if SNS-mediated regulation of inflammation is a mechanism involved in myocardial remodeling secondary to hypertension and whether afferent nerves also have the potential to regulate inflammation in the hypertensive heart.
Methods
All in vivo experiments were performed using adult male spontaneously hypertensive rats (SHR, n=18) and Wistar Kyoto rats (WKY, n=13) housed under standard environmental conditions and maintained on commercial rat chow and tap water ad libitum. SHR were chosen for this study because they have been documented to have elevated SNS activity. 16 All studies conformed to the principles of the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Additionally, the protocol was approved by our Institution's Animal Care and Use Committee. All rats were anesthetized by intraperitoneal injection of sodium pentobarbital (50 mg/kg).
Experimental Protocol and Sympathectomy Surgery
Rats were divided into four groups: 1) untreated WKY; 2) sympathectomized WKY (WKY+Symp); 3) untreated SHR; and 4) sympathectomized SHR (SHR+Symp). Data reported herein for untreated WKY and SHR have been previously published. 6 Bilateral sympathectomy surgery was performed at 8 weeks of age, and all rats were sacrificed at 20 weeks of age. This time-point was chosen as it represents a steady state of hypertension as well as established LV hypertrophy and fibrosis in the SHR. The sympathectomy surgery was conducted by the attending veterinarian at Charles River Laboratories using the following procedure. A ventral midline incision was made in the neck allowing the exposed osmohyoid muscle to be retracted laterally to expose the carotid artery internal and external branche region. The superior cervical ganglion lying within this bifurcation was carefully excised. This procedure was repeated on the contralateral side. A successful sympathectomy was evidenced by the occurrence of Horner's syndrome. Adrenalectomy was not performed in these animals because the aim was to examine the specific interactions between the SNS and MCs, not the role of circulating norepinephrine.
Hemodynamic Parameters
At the experimental end-point, animals were anesthetized with sodium pentobarbital (50 mg/kg) and measurements of systolic, diastolic and mean arterial pressures (SBP, DBP, MAP) were made using a high fidelity Millar catheter inserted into the carotid artery and advanced into the aortic arch. Catheter data were analyzed using PVAN 3.5 software.
Histological Analysis
MC number and density were determined from pinacynol erythrosinate stained LV sections as previously described (please see http://hyper.ahajournals.org). 6,17 LV interstitial collagen volume fraction was determined by analysis of picrosirius red stained sections as previously described (please see http://hyper.ahajournals.org).6
Cytokine Characterization
IFN-γ, IL-4, IL-6 and IL-10 were quantified from frozen LV tissue (please see http://hyper.ahajournals.org) to determine the effects of sympathectomy, using commercial ELISA kits (Alpco Diagnostics, Salem, NH).
Cardiac Inflammatory Cell Isolation Procedure
Cardiac inflammatory cells were isolated as previously described. 15 Briefly, a thoracotomy exposed the intact pericardial sac. The tip of a teflon catheter sleeve, attached to a sterile syringe, was then inserted into the pericardial sac and the sac filled with Hanks balanced salt solution (HBSS, 7.4 pH) composed of: 1) Hanks calcium and magnesium free salt solution; 2) HEPES (13 mM); 3) 607 units/mL of deoxyribonuclease I (Sigma Chemical Co., St. Louis, MO); and 4) an antibiotic-antimycotic mixture of penicillin G sodium, 10,000 mg/mL of streptomycin sulfate, and 25 mg/mL of amphotericin B (Gibco-BRL, Life Technologies, Grand Island, NY). The buffer was then aspirated into another sterile syringe. This was repeated several times resulting in the collection of predominantly T cells, MCs and monocyte/macrophages (Figure S1; please see http://hyper.ahajournals.org). The cells were pelleted and resuspended in Hyclone buffer (HBSSMC; HBSS containing magnesium sulphate (1.1 mM), calcium chloride (1.3 mM), and phenol red; Hyclone, Logan, UT) before undergoing treatment.
Isolated Cardiac Mast Cell Studies
Cardiac MCs were obtained during the cardiac inflammatory cell isolation procedure described above. Cardiac MC responses were examined by incubating 4 × 103 MCs per treatment tube with norepinephrine or substance P at specific concentrations: 0, 1×10-12, 1×10-9, 1×10-6 and 1×10-4 M. The samples were incubated at 37 °C for 20 min and the post-treatment supernatants and pellets separated for subsequent analysis of histamine as a marker of MC degranulation. Percent histamine release was determined by dividing the histamine value from the supernatant by total histamine (supernatant plus pellet). Histamine concentration was measured using a Neogen® Veratox Histamine ELISA (Lexington, KY).
Substance P-mediated Production of Angiotensin II by Isolated Cardiac Inflammatory Cells
Isolated cardiac inflammatory cells (2 ×105/well) were maintained in DMEM containing 10% FBS and treated with substance P (1×10-4 M) for 20 hours. The media was collected and assayed for angiotensin II using a commercial enzyme immunoassay kit (SPI-BIO), which has cross-reactivities of 4, 36 and 33% with angiotensin I, angiotensin III and angiotensin 3-8, respectively.
Immunofluorescent Staining of α-Adrenergic Receptors
Please see http://hyper.ahajournals.org.
Statistical Analysis
All grouped data were expressed as mean ± standard deviation (SD) or standard error of the mean (SEM) as appropriate. Group comparisons were made by one-way ANOVA using SPSS 11.5 software (SPSS Inc., Chicago, IL). When a significant F test (p≤0.05) was obtained, inter-group comparisons were analyzed using Fisher's protected least significant difference post-hoc testing.
Results
Biometrics
Table 1 displays grouped biometric data. While no differences in body weight were observed between WKY, SHR and WKY+Symp, a significant increase was observed in sympathectomized SHR. Table 1 also includes LV weight and LV weight indexed to body weight as measures of hypertrophy. SHR developed significant LV hypertrophy in comparison to WKY by both measures, with sympathectomy preventing this hypertrophic response. There were no weight differences between WKY and WKY+Symp. Lung weights showed no significant differences. SHR showed marked increases in SBP, DBP and MAP compared to WKY. Sympathectomy in the SHR caused a significant attenuation of the blood pressure parameters, whereas sympathectomy had no effect on blood pressure in the WKY group.
Table 1.
Biometric data.
| Group | BW (g) | LV (mg) | LV/BW (mg/g BW) |
Lung Weights (mg) |
SBP (mmHg) |
DBP (mmHg) |
MAP (mmHg) |
|---|---|---|---|---|---|---|---|
| WKY | 328±28 | 786.8±54.0 | 2.24±0.11 | 1476±217 | 116±19 | 86±20 | 100±20 |
| SHR | 345±11 | 929.5±81.1* | 2.70±0.24* | 1445±122 | 229±26* | 167±13* | 193±17* |
| WKY+Symp | 332±14 | 755.8±45.3 | 2.28±0.06 | 1303±179 | 123±20 | 98±17 | 109±19 |
| SHR+Symp | 421±18*† | 715.8±26.9† | 1.70±0.04*† | 1511±154 | 147±16*† | 116±11*† | 130±13*† |
All values are mean ± SD. BW, body weight; LV, left ventricle; SBP, systolic blood pressure; DBP, diastolic blood pressure; MAP, mean arterial pressure.
p<0.05 versus WKY;
p<0.05 versus SHR.
Collagen Volume Fraction
Collagen volume fraction (CVF) results are displayed in Figure 1. LV CVF in the SHR was markedly increased in comparison with that in WKY. However, this increase was significantly attenuated in the sympathectomized SHR group. CVF was not affected by sympathectomy in the WKY.
Figure 1.

Left ventricular collagen volume fraction in WKY (n=4), SHR (n=4), WKY+Symp (n=4) and SHR+Symp (n=4). All values are mean ± SEM. * = p<0.05 versus WKY; † = p<0.05 versus SHR.
Cardiac Mast Cell Number and Density
A significant increase in the number of MCs occurred within the LV of SHR (WKY, 73.3±10.3 versus SHR 105.7±14.8 cells/section; p<0.05). However, due to the larger area of the SHR LV, there was no difference when expressed as MC density (Figure 2). In contrast, SHR that had undergone sympathectomy showed a dramatic increase in both LV MC number and density (SHR+Symp, 316.3±68.6 cells/section; p<0.05 versus WKY and SHR). In contrast, sympathectomy had no effect on MC number or density in WKY rats (WKY+Symp, 84.2±12.1 cells/section).
Figure 2.

Changes in left ventricular MC density in WKY (n=4), SHR (n=6), WKY+Symp (n=6) and SHR+Symp (n=4). All values are mean ± SD. * = p<0.05 versus WKY; † = p<0.05 versus SHR.
Myocardial Cytokine Profile
Elevated levels of IFN-γ in the SHR (Figure 3A) were normalized by sympathectomy. Sympathectomy had no effect in the WKY. Although myocardial IL-4 levels were also elevated in the SHR (Figure 3B) sympathectomy did not change these levels. However, sympathectomy in the WKY did cause a significant increase in IL-4. Myocardial IL-6 (Figure 3C) was found to be markedly lower in the SHR than the WKY, but was normalized by sympathectomy. Sympathectomy did not change IL-6 levels in the WKY. IL-10 was also found to be decreased in the SHR myocardium (Figure 3D). Sympathectomy resulted in increased amounts of IL-10 in both WKY and SHR.
Figure 3.

Changes in left ventricular levels of (A) IFN-γ, (B) IL-4, (C) IL-6 and (D) IL-10 in WKY (n=4), SHR (n=4), WKY+Symp (n=4) and SHR+Symp (n=4). All values are mean ± SEM. * = p<0.05 versus WKY; † = p<0.05 versus SHR.
Effect of SNS Products on Cardiac Mast Cell Activation and Angiotensin II Production
Incubation of isolated cardiac MCs with norepinephrine at concentrations of 1×10-12, 1×10-9, 1×10-6 and 1×10-4 M, did not induce degranulation (Figure 4A). In contrast, while no effects of substance P were seen at lower concentrations, 1×10-4 M did stimulate cardiac MC degranulation (Figure 4B). Also, this concentration of substance P stimulated the production of angiotensin II by a mixed population of isolated cardiac inflammatory cells (Figure 4C).
Figure 4.

Concentration-responses for histamine release by isolated cardiac MCs in response to norepinephrine (n=4) (A) and substance P (n=4) (B). Angiotensin II produced in response to stimulation with substance P (1 × 10-4 M) by a mixed population of isolated cardiac inflammatory cells, including MCs, lymphocytes and macrophages (C). p<0.05 versus control.
Identification of α-Adrenergic Receptors on Cardiac Mast Cells
Isolated cardiac mast cells were found to not possess α-adrenergic receptors (Figure S2; please see http://hyper.ahajournals.org).
Discussion
Chronic elevation of SNS activity in the myocardium is a key component of the altered signaling pathways that accompany cardiac remodeling resulting from hypertension. While there is evidence that the SNS mediates cardiac fibrosis in hypertension via α-adrenergic receptor stimulation, 2 the exact mechanisms have not been fully elucidated. However, it is likely to be more complex than direct activation of α-adrenergic receptors on cardiac fibroblasts. To this end, inflammation is also an important component in the development of hypertension-induced cardiac fibrosis. 3-5,7,8,18 More specifically, we recently demonstrated a causal role for MCs in regulating myocardial cytokines, macrophage recruitment and development of fibrosis in the hypertensive heart. 6 Given the critical roles of both SNS up-regulation and MCs in hypertension-induced myocardial remodeling, the current study aimed to explore the effects of surgical sympathectomy on cardiac MCs as well as specific cytokines in the hypertensive heart.
Following sympathectomy in SHR, we observed a dramatic almost 5-fold increase in cardiac MC density. While surgical removal of the superior cervical ganglion likely does not result in complete denervation of the heart, nerve fibers from this ganglion do project to the heart 19, and this striking change in MC density clearly demonstrates an impact of its removal on the heart. This same relationship is not present in the normal heart since such a change did not occur in sympathectomized WKY. Facoetti et al. 20 also found cardiac MC density to be unchanged in normal Sprague-Dawley rats following 10 weeks of chemical sympathectomy induced by 6-hydroxydopamine. These disparate effects on MC density are likely indicative of underlying differences in the local environment of normal versus hypertensive hearts. That fibrosis was attenuated in sympathectomized SHR despite MC density being elevated, suggests that either sympathectomy altered MCs to that of a non-fibrotic phenotype, or alternatively, MCs signal the SNS, but can no longer induce a pro-fibrotic response from the damaged SNS. Interestingly, in our isolated cardiac MC experiments, incubation with norepinephrine failed to induce degranulation, and we were unable to detect α-adrenergic receptors on cardiac mast cells using flow cytometry. This is consistent with a previous report that norepinephrine was not the mechanism by which contact with sympathetic neurons decreased membrane resistance in immortalized RBL-2H3 cells, a widely used model for MCs. 13 However, some afferent nerve fibers projecting from the heart, travel through the superior cervical ganglia, 21 and substance P, which is primarily located in afferent nerve fibers, has been demonstrated to be present in superior cervical ganglia. 22 Substance P-containing nerves are associated with numerous areas of the heart including the ventricle, atria, valves and connective linings. 23 Here we show that substance P was a strong inducer of isolated cardiac MC degranulation, consistent with what we have demonstrated previously 15 and suggestive of afferent activation of cardiac mast cells.
Suzuki et al., 24 using a co-culture system, showed that RBL-2H3 mast cells activated sympathetic neurons following IgE stimulation. Activation of sympathetic neurons did not occur if RBL-2H3 cells were not present. The most obvious mechanism for MC activation of neurons to occur is through release of MC tryptase acting on protease activated receptor-2 (PAR-2). Steinhoff et al. 25 recently detected PAR-2 on a large number of neurons containing substance P, and activation of these receptors by tryptase in dorsal horn slices initiated the release of substance P. However, cardiac MCs are also capable of producing renin and are, assumed to be involved in the local generation of angiotensin II in the heart. 12,26,27 Silver et al. 12 postulated that MC generated angiotensin II could then activate nerves through angiotensin receptors. We isolated a mixed population of inflammatory cells including lymphocytes, macrophages and MCs from rat hearts and found that incubation with substance P resulted in angiotensin II production. Thus, while in sympathectomized SHR, superficially it appears that the SNS is regulating cardiac MC density, MCs may actually be upstream of SNS activation and, therefore, responding to an underlying stimulus (e.g., increased wall stress) by unsuccessfully attempting to up-regulate SNS activity. That is, the increase in MC density may be an attempt to achieve increased SNS activity that cannot be elicited because of the sympathectomy. Therefore, we speculate, that a self-perpetuating relationship may exist, where cardiac MC tryptase could activate PAR-2 on afferent nerve fibers causing the release of substance P, which in turn induces angiotensin II production by inflammatory cells, that then acts on fibroblasts and also sympathetic nerves to release norepinephrine (Figure 5). Angiotensin II has been shown to induce myocardial necrosis through stimulation of sympathetic neurons. 28,29 In fact, a similar situation has been described in the rat knee joint, where it appears that sensory nerve activation of mast cells led to stimulation of the SNS and subsequently plasma extravasion. 30 Elaboration of norepinephrine by the SNS in response to stimulation by MCs would explain how α-adrenergic receptors mediate fibrosis 2 when cardiac MCs do not degranulate in response to norepinephrine in our study. This may also explain why sympathectomy does not result in increased cardiac MC density in the normal heart where there is no underlying pathologic stimulus driving MCs to up-regulate sympathetic activity.
Figure 5.
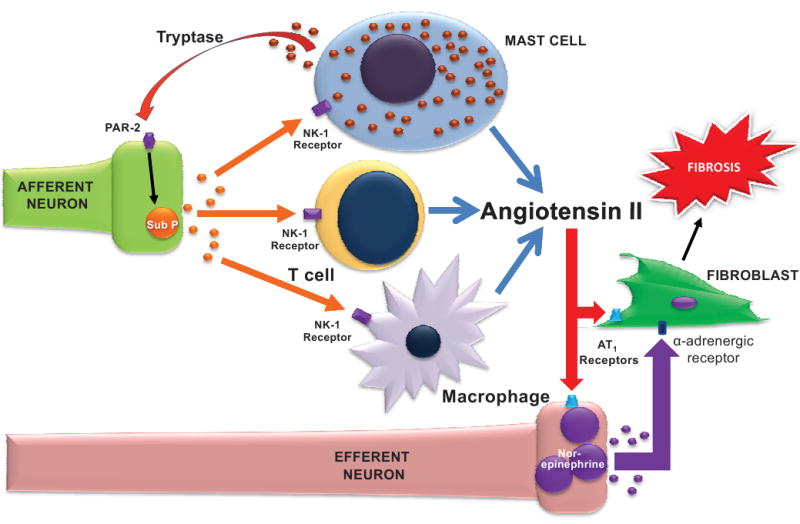
Schematic depicting a possible series of events in the development of fibrosis in the hypertensive heart. Cardiac mast cell tryptase may activate protease activated receptor-2 (PAR-2) on afferent nerve fibers resulting in the release of substance P, which in turn acts on the neurokinin-1 (NK-1) receptor to induce production of angiotensin II by inflammatory cells. Angiotensin II, in addition to activating fibroblasts, may also stimulate efferent nerves to produce norepinephrine, in turn further stimulating fibroblasts to produce collagen.
With this clear SNS-MC interaction, we sought to determine the effect of sympathectomy on selected myocardial cytokines that we have previously shown to be regulated, either directly or indirectly, by MCs in the hypertensive heart. 6 Both IFN-γ and IL-4 were elevated in the SHR, with sympathectomy normalizing IFN-γ, but not IL-4. IFN-γ is a marker of TH1 T cell phenotype 31 and its elevation in hypertension is consistent with the supposition advanced by Yu et al. 9 that TH1 cells may be pro-fibrotic. Interestingly, while sympathectomy had no effect on IFN-γ in the WKY, sympathectomy increased IL-4 levels in the WKY. Combined with our previous studies, 6 it would appear that, in the normal heart, the SNS has a role in regulating IL-4, but that role is shifted to MCs in the hypertensive heart. Although the levels of IL-6 and IL-10 were lower in the SHR myocardium compared to the WKY, sympathectomy returned them to normal. What is intriguing about these results is that, with the exception of IL-4, these changes are almost identical to what we have observed previously in the SHR following MC stabilization. 6 This similarity, together with the close spatial relationship between the nervous system and MCs, 11,12 is more evidence of an interplay between the SNS and MCs in the regulation of the inflammatory process in the hypertensive heart.
In addition to attenuating cardiac fibrosis, sympathectomy in the SHR also prevented myocardial hypertrophy. However, our previous finding that MC stabilization did not prevent hypertrophy in the hypertensive heart indicates that MCs are not involved in this modulation of hypertrophy.6 Instead, rather than acting via inflammatory cells to induce hypertrophy, the SNS most likely has direct effects on cardiomyocytes via β-adrenergic receptors. While it cannot be ruled out that the changes in these parameters and the assayed cytokines were the result of the sympathectomy-related attenuation of blood pressure, we believe the following considerations support the argument that the changes in cytokines and fibrosis were due to direct interactions between the SNS and MCs. Firstly, attenuation of hypertrophy by sympathectomy has been shown to be independent of systolic blood pressure, 2,32 and Perlini et al. 2 further demonstrated that chemical sympathectomy in hypertensive rats prevented cardiac fibrosis independent of blood pressure. Secondly, cardiac MC density was dramatically with sympathectomy in the SHR but not in the normal non-hypertensive heart. Thirdly, our in vitro studies demonstrate that substance P, found in afferent nerves, can stimulate cardiac MC activation and angiotensin II production from a mixed population of cardiac inflammatory cells. Finally, the cytokine profile reported herein following sympathectomy in the SHR is almost identical to that following MC stabilization in the SHR where blood pressure was unaffected. 6
Perspectives
The results of this study reiterate the importance of the SNS in the development of myocardial remodeling and provide evidence of a link between the neurohormonal and inflammatory paradigms of hypertension, by demonstrating the importance of SNS interactions with cardiac MCs and cytokine production. Furthermore, even though a number of inflammatory cell types have been implicated in the development of fibrosis, 7,9,10 our findings are consistent with the hypothesis that MCs not only trigger and orchestrate inflammatory responses, 33 but are also responsible for cardiac fibrosis. It is important to note that not all of the cardiac remodeling effects of the SNS are mediated via mast cells. Clearly hypertrophy does not appear to involve SNS interactions with mast cells, but instead is likely the result of direct stimulation of cardiomyocyte β-adrenergic receptors. The findings herein indicate the need for further investigation regarding the mechanisms mediating SNS, inflammatory cell and afferent nerve interactions in the hypertensive heart, with particular focus on the role of substance P.
Supplementary Material
Acknowledgments
We would like to thank Purnima Jani for her excellent technical assistance.
Sources of Funding: This study was supported in part by an American Heart Association Postdoctoral Fellowship 0825510E (SPL) as well as National Heart, Lung, and Blood Institute Grants R01-HL-62228 (JSJ) and R01-HL-073990 (JSJ).
Footnotes
Conflicts of Interest: None
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Reference List
- 1.Pepper GS, Lee RW. Sympathetic activation in heart failure and its treatment with β-blockade. Arch Intern Med. 1999;159:225–234. doi: 10.1001/archinte.159.3.225. [DOI] [PubMed] [Google Scholar]
- 2.Perlini S, Palladini G, Ferrero I, Tozzi R, Fallarini S, Facoetti A, Nano R, Clari F, Busca G, Fogari R, Ferrari AU. Sympathectomy or doxazosin, but not propranolol, blunt myocardial interstitial fibrosis in pressure-overload hypertrophy. Hypertension. 2005;46:1213–1218. doi: 10.1161/01.HYP.0000185689.65045.4c. [DOI] [PubMed] [Google Scholar]
- 3.Hara M, Ono K, Hwang MW, Iwasaki A, Okada M, Nakatani K, Sasayama S, Matsumori A. Evidence for a role of mast cells in the evolution to congestive heart failure. J Exp Med. 2002;195:375–381. doi: 10.1084/jem.20002036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Panizo A, Mindan FJ, Galindo MF, Cenarruzabeitia E, Hernandez M, Diez J. Are mast cells involved in hypertensive heart disease? J Hypertens. 1995;13:1201–1208. doi: 10.1097/00004872-199510000-00015. [DOI] [PubMed] [Google Scholar]
- 5.Shiota N, Rysa J, Kovanen PT, Ruskoaha H, Kokkonen JO, Lindstedt KA. A role for cardiac mast cells in the pathogenesis of hypertensive heart disease. J Hypertens. 2003;21:1823–1825. doi: 10.1097/00004872-200310000-00022. [DOI] [PubMed] [Google Scholar]
- 6.Levick SP, McLarty JL, Murray DB, Freeman RM, Carver WE, Brower GL. Cardiac mast cells mediate left ventricular fibrosis in the hypertensive rat heart. Hypertension. 2009;53:1041–1047. doi: 10.1161/HYPERTENSIONAHA.108.123158. [DOI] [PubMed] [Google Scholar]
- 7.Kuwahara F, Kai H, Tokuda K, Takeya M, Takeshita A, Egashira K, Imaizumi T. Hypertensive myocardial fibrosis and diastolic dysfunction: another model of inflammation? Hypertension. 2004;43:739–745. doi: 10.1161/01.HYP.0000118584.33350.7d. [DOI] [PubMed] [Google Scholar]
- 8.Kagitani S, Ueno H, Hirade S, Takahashi T, Takata M, Inoue H. Tranilast attenuates myocardial fibrosis in association with suppression of monocyte/macrophage infiltration in DOCA/salt hypertensive rats. J Hypertens. 2004;22:1007–1015. doi: 10.1097/00004872-200405000-00024. [DOI] [PubMed] [Google Scholar]
- 9.Yu Q, Watson RR, Marchalonis JJ, Larson DF. A role for T lymphocytes in mediating cardiac diastolic function. Am J Physiol Heart Circ Physiol. 2005;289:H643–H651. doi: 10.1152/ajpheart.00073.2005. [DOI] [PubMed] [Google Scholar]
- 10.Yu Q, Horak K, Larson DF. Role of t lymphocytes in hypertension-induced cardiac extracellular matrix remodeling. Hypertension. 2006;48:98–104. doi: 10.1161/01.HYP.0000227247.27111.b2. [DOI] [PubMed] [Google Scholar]
- 11.Arizono N, Matsuda S, Hattori T, Kojima Y, Maeda T, Galli SJ. Anatomical variation in mast cell nerve associations in the rat small intestine, heart, lung, and skin. Similarities of distances between neural processes and mast cells, eosinophils, or plasma cells in the jejunal lamina propria. Lab Invest. 1990;62:626–634. [PubMed] [Google Scholar]
- 12.Silver RB, Reid AC, Mackins CJ, Askwith T, Schaefer U, Herzlinger D, Levi R. Mast cells: A unique source of renin. PNAS. 2004;101:13607–13612. doi: 10.1073/pnas.0403208101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Blennerhassett MG, Janiszewski J, Bienenstock J. Sympathetic nerve contact alters membrane resistance of colls of the RBL-2H3 mucosal mast cell line. Am J Respir Cell Mol Biol. 1992;6:504–509. doi: 10.1165/ajrcmb/6.5.504. [DOI] [PubMed] [Google Scholar]
- 14.Madden KS, Sanders VM, Felten DL. Catecholamine influences and sympathetic neural modulation of immune responsiveness. Annu Rev Pharmacol Toxicol. 1995;35:417–448. doi: 10.1146/annurev.pa.35.040195.002221. [DOI] [PubMed] [Google Scholar]
- 15.Morgan LG, Levick SP, Voloshenyuk TG, Murray DB, Forman MF, Brower GL, Janicki JS. A novel technique for isolating functional mast cells from the heart. Inflamm Res. 2008;57:1–6. doi: 10.1007/s00011-007-7059-5. [DOI] [PubMed] [Google Scholar]
- 16.Provoost AP, De Jong W. Differential development of renal, DOCA-salt, and spontaneous hypertension in the rat after neonatal sympathectomy. Clin Exp Hypertens. 1978;1:177–189. doi: 10.3109/10641967809068603. [DOI] [PubMed] [Google Scholar]
- 17.Bensley SH. Pinacyanol erthrosinate as a stain for mast cells. Stain Technol. 1952;27:269–273. doi: 10.3109/10520295209105090. [DOI] [PubMed] [Google Scholar]
- 18.Tomita H, Egashira K, Kubo-Inoue M, Usui M, Koyanagi M, Shimokawa H, Takeya M, Yoshimura T, Takeshita A. Inhibition of NO synthesis induces inflammatory changes and monocyte chemoattractant protein-1 expression in rat hearts and vessels. Arteriosclerosis, Thrombosis, and Vascular Biology. 1998;18:1456–1464. doi: 10.1161/01.atv.18.9.1456. [DOI] [PubMed] [Google Scholar]
- 19.Kawashima T. The autonomic nervous system of the human heart with special reference to its origin, course, and peripheral distribution. Anat Embryol (Berl) 2005;209:425–438. doi: 10.1007/s00429-005-0462-1. [DOI] [PubMed] [Google Scholar]
- 20.Facoetti A, Fallarini S, Miserere S, Bertolotti A, Ferrero I, Tozzi R, Gatti C, Palladini G, Perlini S, Nano R. Histochemical study of cardiac mast cells degranulation and collagen deposition: interaction with the cathecolaminergic system in the rat. Eur J Histochem. 2006;50:133–140. [PubMed] [Google Scholar]
- 21.Meller ST, Gebhart GF. A critical review of the afferent pathways and the potential chemical mediators involved in cardiac pain. Neuroscience. 1992;48:501–524. doi: 10.1016/0306-4522(92)90398-l. [DOI] [PubMed] [Google Scholar]
- 22.Kessler JA, Adler JE, Bell WO, Black IB. Substance p and somatostatin metabolism in sympathetic and special sensory ganglia in vitro. Neuroscience. 1983;9:309–318. doi: 10.1016/0306-4522(83)90296-8. [DOI] [PubMed] [Google Scholar]
- 23.Furness JB, Costa M, Papka RE, Della NG, Murphy R. Neuropeptides contained in peripheral cardiovascular nerves. Clin Exp Hypertens A. 1984;6:91–106. doi: 10.3109/10641968409062553. [DOI] [PubMed] [Google Scholar]
- 24.Suzuki R, Furuno T, Teshima R, Nakanishi M. Bi-directional relationship of in vitro mast cell-nerve communication observed by confocal laser scanning microscopy. Biol Pharm Bull. 2001;24:291–294. doi: 10.1248/bpb.24.291. [DOI] [PubMed] [Google Scholar]
- 25.Steinhoff M, Vergnolle N, Young SH, Tognetto M, Amadesi S, Ennes HS, Trevisani M, Hollenberg MD, Wallace JL, Caughey GH, Mitchell SE, Williams LM, Geppetti P, Mayer EA, Bunnett NW. Agonists of proteinase-activated receptor 2 induce inflammation by a neurogenic mechanism. Nat Med. 2000;6:151–158. doi: 10.1038/72247. [DOI] [PubMed] [Google Scholar]
- 26.Mackins CJ, Kano S, Seyedi N, Schafer U, Reid AC, Machida T, Silver RB, Levi R. Cardiac mast cell-derived renin promotes local angiotensin formation, norepinephrine release, and arrhythmias in ischemia/reperfusion. J Clin Invest. 2006;116:1063–1070. doi: 10.1172/JCI25713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Reid AC, Silver RB, Levi R. Renin: at the heart of the mast cell. Immunol Rev. 2007;217:123–140. doi: 10.1111/j.1600-065X.2007.00514.x. [DOI] [PubMed] [Google Scholar]
- 28.Henegar JR, Brower GL, Kabour A, Janicki JS. Catecholamine response to chronic ANG II infusion and its role in myocyte and coronary vascular damage. Am J Physiol Heart Circ Physiol. 1995;269:H1564–H1569. doi: 10.1152/ajpheart.1995.269.5.H1564. [DOI] [PubMed] [Google Scholar]
- 29.Henegar JR, Schwartz DD, Janicki JS. ANG II-related myocardial damage: role of cardiac sympathetic catecholamines and beta-receptor regulation. Am J Physiol Heart Circ Physiol. 1998;275:H534–H541. doi: 10.1152/ajpheart.1998.275.2.H534. [DOI] [PubMed] [Google Scholar]
- 30.Coderre TJ, Basbaum AI, Levine JD. Neural control of vascular permeability: interactions between primary afferents, mast cells, and sympathetic efferents. J Neurophysiol. 1989;62:48–58. doi: 10.1152/jn.1989.62.1.48. [DOI] [PubMed] [Google Scholar]
- 31.Rogge L. A Genomic View of Helper T Cell Subsets. Ann NY Acad Sci. 2002;975:57–67. doi: 10.1111/j.1749-6632.2002.tb05941.x. [DOI] [PubMed] [Google Scholar]
- 32.Perlini S, Ferrero I, Palladini G, Tozzi R, Gatti C, Vezzoli M, Cesana F, Janetti MB, Clari F, Busca G, Mancia G, Ferrari AU. Survival benefits of different antiadrenergic interventions in pressure overload left ventricular hypertrophy/failure. Hypertension. 2006;48:93–97. doi: 10.1161/01.HYP.0000226859.42064.ea. [DOI] [PubMed] [Google Scholar]
- 33.Kinet JP. The essential role of mast cells in orchestrating inflammation. Immunol Rev. 2007;217:5–7. doi: 10.1111/j.1600-065X.2007.00528.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.