

Abstract
Background
Across multiple mental health-related measures, a polymorphism (5-HTTLPR) within the promoter of the serotonin transporter gene has been associated with differential psychological sensitivity to stressful experiences. Yet, the specific mechanisms by which this polymorphism contributes to risk for psychological dysfunction is unclear. Therefore, we investigated cortisol reactivity to psychosocial stress as a potential intermediate phenotype that may predispose to such risk.
Methods
A psychologically healthy sample of 182 young adults were genotyped for the 5-HTTLPR. Each participant delivered a speech and performed mental arithmetic in one of three audience conditions: A critical evaluative audience, a supportive evaluative audience, or no audience. Salivary cortisol was sampled at baseline and at 20, 40, and 75 minutes after stressor onset.
Results
The two evaluative audience conditions elicited similar, significant increases in cortisol that were significantly greater than in the no audience control. The evaluative audience conditions revealed a significant relationship between cortisol reactivity and the 5-HTTLPR, with the short/short genotype showing the greatest reactivity. Internal analyses revealed that the 5-HTTLPR was significantly associated with cortisol reactivity in the negative audience condition only, suggesting that short/short individuals may be especially vulnerable to social threat.
Conclusions
The short/short genotype of the 5-HTTLPR is associated with greater cortisol reactivity to social threat. When short/short individuals experience stressful life events, they may be at greater risk for the adverse psychological and physical health consequences associated with heightened cortisol exposure.
Keywords: Stress, Serotonin, Cortisol, Mental Health Disorders
Introduction
Although it is well-documented that exposure to stressful life experiences increases risk for adverse physical and mental health outcomes (1), this stress-related increase in risk is not uniform. Accumulating evidence indicates that genetic variation is a key contributor to individual variability in response to stressful life events (2). One marker of such variation is a polymorphism (5-HTTLPR) within the promoter of the serotonin transporter gene (SLC6A4). At this locus, there are two principal alleles, short and long, that differ in length by 43 base pairs (3, 4).
A growing body of research suggests that the 5-HTTLPR moderates the effects of adverse life experiences on the probability and severity of a diverse array of mental health related conditions and constructs, including depression (5, 6), suicide (7, 8), anxiety (9, 10), and substance abuse (11), to name a few. Across these diverse stress-related psychological disorders, the majority of studies associate the 5-HTTLPR short allele, and particularly the short/short genotype, with greater psychological sensitivity to stress (12), although there is variability in the success of replicating such findings (13). One potential explanation for the variability in results is that the 5-HTTLPR is more closely associated with the regulation of the hypothalamic-pituitary-adrenal (HPA) axis response to stress than it is with any particular psychological state that is influenced by stress. Thus, the cortisol response to stressful life events may represent an intermediate phenotype between the 5-HTTLPR and psychological disorder.
Alterations in HPA axis activity are present in subgroups of a broad array of psychological disorders (14). For example, hypercortisolemia is characteristic of certain depressed patients, which has led to speculation that heightened cortisol levels may actually be causal in the onset of the disorder (15). The serotonin system is ideally positioned to moderate cortisol reactivity to stressors via its ability to influence neural activity at suprahypothalamic levels as well as at each level of the hypothalamic-pituitary-adrenal axis (16, 17). In particular, serotonin fibers activate the hypothalamic corticotrophin-releasing factor containing neurons (18, 19) that initiate the hormonal cascade leading to cortisol release. Therefore, variation in serotonin system functioning, for which the 5-HTTLPR is a marker, may be an important link between exposure to stressful life events and the altered HPA axis activity seen in some patient subgroups. However, studying the relationship between the 5-HTTLPR and cortisol reactivity in clinical samples is complicated by the effects of the disease state on both the HPA axis and the serotonin system (20).
Therefore, in this study, the goal was to determine if the 5-HTTLPR is associated with cortisol responses to laboratory-based psychosocial stressors in a healthy adult sample. An association of the 5-HTTLPR with cortisol reactivity to psychosocial stress would suggest that one means by which the 5-HTTLPR influences mental health and well-being may be via differential cortisol exposure from experiencing stress.
Methods and Materials
Participants
Participants responded to a poster offering $120 compensation for participation in the study. Prospective participants were screened during a telephone interview, and those who were currently being treated by a mental health professional, had mental or physical health problems (including post-traumatic stress disorder), or using mental-health related medications (e.g. Prozac) or other medications affecting cardiovascular or endocrine function were excluded. In addition, pregnant or lactating women were excluded. All procedures were approved by the Institutional Review Board from the University of California at Los Angeles. The final sample consisted of 185 participants (39% male; 61% female; age range:18 to 35). As participants were affiliated with the University as students, employees, or both, the sample reflects these demographics and was 37% Asian-American, 22% European-American, 16% Latino, 23% of “mixed” ethnicity and 2% African American (These last three groups are designated “other” in the analyses to be reported).
Trier Social Stress Task
Participants reported to the University's Clinical Research Center in the mid to late afternoon in order to control for diurnal variation in cortisol (21). After arrival, participants were further screened for: 1) blood pressure less than 140/90 mmHg, 2) a resting pulse between 60 and 100, 3) normal lung auscultation, and 4) a normal cardiac exam (no evidence of congestive heart failure or arrythmia). Participants then provided a baseline saliva sample from which cortisol concentrations were measured. The nurse then inserted an indwelling catheter and performed a blood draw to assay gonadal hormone and peptide levels (data not reported here). Blood pressure and heart rate were assessed every three minutes via a Critikon Dinamap Vital Signs Monitor Model 1846SX (data not reported here). After 30 minutes of screening for medical problems, the second saliva sample was taken. At this time, participants completed the Positive and Negative Affect Schedule (PANAS; 22), which served as a baseline for self-reported response to the stress task. Cortisol samples at 60 and 30 minutes prior to the onset of the TSST were averaged to form the baseline measure.
Each participant then took part in the Trier Social Stress Task (TSST), a widely-used laboratory stress challenge known to elicit autonomic and HPA axis stress responses (23). Participants performed the TSST in front of a video camera. The experimenter sat off to the side and out of direct view of the participant and did not give any verbal or nonverbal indications of positive or negative reactions to the participant's performance.
Participants were given five minutes to prepare a five-minute speech on why they would be a good administrative assistant, a popular campus job for students and employees. They then delivered the speech under one of three audience conditions. In the negative audience condition, the participant delivered his/her speech to two members of an evaluative panel who gave nonverbal indications of frustration over the quality of the speech. They displayed non-verbal signs of boredom and exchanged glances with each other that communicated mutual negative assessments. This manipulation represents a somewhat stronger version of the standardized audience condition for the TSST. In the positive audience condition, the two audience members leaned forward, smiled, and gave non-verbal indications of approval. They occasionally exchanged glances with each other that communicated mutual positive assessments and when not explicitly communicating positive assessments, their demeanor communicated interest in what the participant was saying. The two audience conditions mirrored each other precisely in terms of the timing and type of feedback, with the exception that the nonverbal feedback was positive in one condition and negative in the other. All panels included one man and one woman, and measures were taken to ensure the participant and audience members were unacquainted. The no audience control condition did not include an audience.
After the speech, participants performed challenging mental arithmetic tasks for 5 minutes that required counting backwards by 7s and by 13s from 2,935 out loud, during which time they were urged by the experimenter to try to go faster. For the participants in the two audience conditions, these math problems were completed in front of the audience as well.
Twenty minutes after the commencement of the TSST, participants provided a third saliva sample. This time lag falls within the recommended window for observing stress-related increases in cortisol (24). Participants then rated their affective state on the PANAS. At 40 minutes after the onset of the TSST, the fourth cortisol sample was taken. A 35-minute recovery period then took place after which the fifth (recovery) cortisol sample was taken. This time lag is typically associated with significant declines in cortisol levels from peak stress, although not always full return to baseline (25). At the conclusion of the experiment, the participant was debriefed and then dismissed.
Genotyping
DNA was collected from saliva using Oragene kits (DNA Genotek, Ottawa, Ontario, Canada) and extracted according to the manufacturer's recommendations. The 5-HTTLPR was genoptyed as described previously (5) using a protocol modified from (4). Briefly, the forward primer was 5′-GGC GTT GCC GCT CTG AAT GC-3′ (labeled with 6-carboxyfluorescein fluorophore), and the reverse primer was 5′- GAG GGA CTG AGC TGG ACA ACC AC-3′, which yielded 486-bp (short) and 529-bp (long) fragments. Polymerase chain reaction (PCR) was performed in a total volume of 25μl, containing 100 ng of DNA, 160 nM of each primer, 1mM Tris-HCL (pH 8.3), 5mM KCl, 1.5 mM MgCl2, 2% DMSO (v/v), 2.5 U Amplitaq Gold DNA polymerase (Applied Biosystems, Foster City, CA), 200 μM of dATP, dCTP, dTTP, and 100 μM of dGTP, and 7-deaza-2′-dGTP. After an initial denaturation at 94°C for 5 min, 35 cycles of denaturation (94°C for 30 sec), annealing (63°C for 30 sec), and extension (72°C for 1 min) was performed followed by a final extension at 72°C for 20 min. The PCR products were electrophoresed on an ABI 3730 DNA analyzer (Applied Biosystems) with a Mapmaker size standard (Bioventures, Murfreesboro, TN). Data collection and analysis used GeneScan and Genotyper software (Applied Biosystems).
Salivary Cortisol Assay Procedures
Saliva was collected using the passive drool method into a 2.0ml Corning cryovial (Corning, Inc., Corning, NY), which was immediately placed on ice and then transferred to a - 20°C freezer for storage. The concentrations of cortisol within these samples were quantified by Salimetrics LLC (State College, PA). The radioimmunoassay was run in duplicate with 25-μl samples using the HS-cortisol High Sensitivity Salivary Cortisol Enzyme Immunoassay Kit (Salimetrics LLC, State College, PA). The HS-Cortisol Assay allows for robust results when the saliva samples have a pH within the range of 3.5-9.0. All samples were within this pH range. The inter- and intra-assay coefficients of variation were 6.1% and 5.0%, respectively.
Statistical Analyses
Tests of Hardy–Weinberg equilibrium for the entire sample and each ethnic grouping were conducted using the software program Haploview v3.32 (http://www.broad.mit.edu/mpg/haploview/; 26). Remaining statistical analyses were performed using SPSS 14.0 (Chicago, IL). Chi-square tests were used to assess allele distributions (2n). For assessment of cortisol responses to the TSST, repeated measures ANOVAs were used with cortisol sample timepoint as a within subjects variable and genotype as a between subjects variable. Greenhouse-Geisser corrections for repeated measures were used due to the violation of sphericity assumptions. For significant results, planned pair-wise comparisons used the Bonferroni correction for multiple comparisons, unless otherwise noted. When ethnicity was used as a covariate, two dummy variables were created (i.e. East Asian = 1, all others = 0; and European-American = 1, all others = 0). Due to the skewed distributions, cortisol values were log transformed prior to analysis. All statistical tests were two-tailed with alpha set to p < 0.05.
Results
One participant could not be genotyped, and two participants did not have the complete time course of cortisol samples, leaving a sample of 182 participants. Across the three experimental conditions, there were no differences in the distribution of gender (χ2(2,N=182)= .05, p = .98), ethnicity (χ2(4,N=182)= 1.69, p = .79), or 5-HTTLPR genotype (χ2(4,N=182) = 2.19, p = .70).
As expected (27), 5-HTTLPR alleles were differentially distributed as a function of ethnic grouping (χ2(2,N=364)= 12.42, p = .002). Among participants of East Asian ancestry, the 5-HTTLPR allele frequency was 70% for the short allele and 30% for the long allele. This allele distribution was significantly different from those declaring European ancestry (Short allele: 47.6%; χ2(1,N=222)= 11.02, p = .001) or Other ancestry (Short allele: 54.1%; χ2(1,N=286)= 7.65, p = .006). There were no differences in allele distribution between the group with European ancestry and the group with “Other” ancestry (χ2(1,N=228)= .9, p = .34). Hardy-Weinberg equilibrium calculations showed no significant deviation from equilibrium for each ethnic grouping (all p's > 0.4), as was the case for the calculation using the entire sample (p = 0.19).
Preliminary Analyses
Across all experimental conditions, cortisol levels showed a significant rise after the onset of the public speaking task and then recovered close to baseline (F(3,546) =56.1, p < .001, η2 = .24). There was a significant interaction between audience condition and time (F(4.29, 386.08) =6.31, p < 0.001, η2 = .066). Although there were significant increases in cortisol levels in each of the experimental conditions (the no audience condition (F(1.96,125.27) =12.94, p < 0.001, η2 = .17); the supportive evaluative audience condition (F(1.76,102.3) =27.95, p < 0.001 η2 = .33); and the negative evaluative audience condition (F(2.46,142.72) =22.99, p < 0.001, η2 = .28), post-test comparisons revealed that both the positive evaluative audience (p < .001) and the negative evaluative audience (p = .045) were significantly different from the no audience condition. The two evaluative audience conditions were not significantly different from each other (p = .175), prompting analysis of the two evaluative audience conditions together.
With respect to ethnicity, there were no significant ethnic differences in cortisol responses in the no audience condition (F(3.91, 121.07)=.30, p = .88) or in the combined evaluative audience conditions (F(4.31,248)= .48, p = .82). When each evaluative audience condition was assessed separately there were also no significant ethnic differences in cortisol response (supportive evaluative audience condition (F(3.51, 98.4)=.11, p = .97); negative evaluative audience condition (F(4.85,135.76)=.7, p = .62)). Likewise, there were no significant gender differences in response to the stressor in the no audience condition (F(1.95,123.07)= .25, p = .77) or in the combined evaluative audience conditions (F(2.19,253.95)= 1.13, p = .33). Similarly, there were no significant effects of gender on cortisol reactivity in the supportive evaluative audience condition: F(1.77, 101.11)= .62, p = .52) and the negative evaluative audience condition (F(2.47, 140.93)= .66, p = .55).
The 5-HTTLPR and Cortisol Response to Psychosocial Stress
To assess genetic moderation of cortisol reactivity, we first assessed the relationship between the 5-HTTLPR and reactivity to social evaluation (i.e., positive and negative audience conditions combined). The analyses revealed a significant relationship between the 5-HTTLPR and cortisol reactivity (F(4.38,251.68)=2.78, p = .024, η2 = .046). Specifically, as Figure 1 shows, the short/short individuals exhibited the strongest cortisol responses to social evaluation. These results were unchanged when gender and ethnicity were entered as covariates (F(4.37,244.24)=2.67, p = .029,η2 = .051).
Figure 1.
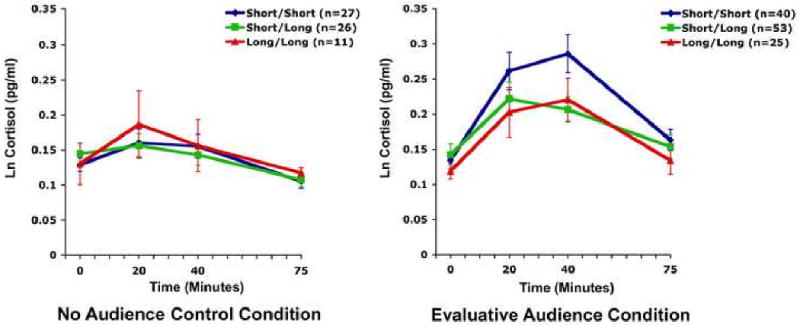
Mean (±SEM) cortisol response to the Trier Social Stress Test as a function of 5-HTTLPR in the no audience condition and the evaluative audience conditions.
With respect to the specific time points, one-way ANOVAs revealed no differences in cortisol levels by allele distribution at baseline (F(2,115)= .57, p = .57), 20 minutes post-stressor onset (F(2,115)= 1.05, p = .35), or recovery (F(2,115)= .54, p = .59), but there were significant differences in cortisol levels at 40 minutes post-stressor onset (F(2,115)= 3.34, p = .04, η2 = .06) with the short/short individuals having the highest cortisol levels.
We next conducted follow-up internal analyses to examine the relationship between 5-HTTLPR genotype and cortisol response in each audience condition. In the no audience condition, there was no relationship between the 5-HTTLPR and cortisol response (F(3.88, 118.44)= .68, p = .6). In the supportive evaluative audience condition, the relationship between the 5-HTTLPR and cortisol response was not significant (F(3.59,100.46)= 1.41, p = .24). However, in the negative evaluative audience condition, there was a significant relationship between the 5-HTTLPR and the cortisol response (F(4.94, 138.17)= 3.37, p = .007, η2 = .28). The short/short individuals had the strongest response and the long/long individuals, the least response (p = .042, LSD), with the short/long individuals falling inbetween. These results remained significant when ethnicity and gender were entered as covariates (F(5.1, 129.98)= 3.42, p = .006, η2 = .11).
To ensure that these significant results were not due to population stratification, the relationship between the 5-HTTLPR and cortisol response was examined separately for each ethnic category. Although the comparisons with this subdivided sample were not significant due to reduced power, the qualitative pattern of cortisol response was the same in each ethnic group, with a graded genotype-dependent decrease in cortisol response, with the short/short individuals being the highest. Thus, among those of European descent (M = 0.25, SD = 0.08), those of East Asian descent (M = 0.26, SD = 0.13) and those of Other ancestry (M = 0.37, SD = 0.21), the mean peak cortisol response of the short/short individuals was greater than that in the short/long individuals (European ancestry: M = 0.2, SD = 0.11; East Asian ancestry, M = 0.19, SD = 0.08; Other ancestry, M = 0.21, SD = 0.13). In turn, the short/long heterozygotes had higher peak cortisol responses than the long/long individuals in both the European ancestry (M = 0.19, SD = 0.13) and Other ancestry (M = 0.13, SD = 0.13) group.
Consistent with findings from a meta-analysis of laboratory stress studies (25), self-reported negative affect did not correlate with the changes in cortisol. Thus, changes in negative affect between the pre and post stressor ratings were not signfiicanly different among the three audience conditions (F(2,176) = .23, p = .8). There was also no relationship between the 5-HTTLPR and changes in negative affect in any of the three audience conditions (no audience: F(2,58)= .096, p = .91; positive audience: F(2,53)= 1.06, p = .35; negative audience F(2,53)= .41, p = .67).
Discussion
The results of this investigation reveal a significant relationship between the 5-HTTLPR and cortisol reactivity to laboratory stressors. Those with the short/short genotype had the greatest increase in cortisol levels followed by those with the short/long genotype, followed by those with the long/long genotype. In addition, the relationship between genotype and cortisol reactivity was strongest when individuals were exposed to a negative evaluative audience, indicating that the short/short genotype appears to confer sensitivity to social threat.
This greater sensitivity in the short allele homozygotes is consistent with other recent studies using stress paradigms similar to the one used here. In a sample of girls, a portion of whom were at familial risk for depression, short/short individuals had the greatest cortisol response to psychosocial stress (28). Similarly, in an adult sample that included healthy participants, depressed patients, and individuals at familial risk for depression, women with the short/short genotype had the greatest cortisol response to psychosocial stress (29). Thus, across studies, a consistent picture is emerging indicating that the short/short genotype of the 5-HTTLPR is associated with higher cortisol response to psychosocial stress. The consistency in the direction of effects across these studies indicates that the study of intermediate phenotypes such as cortisol reactivity may be of great utility in identifying genetic associations, relative to studies of broader phenotypes such as psychiatric diagnoses.
The presence or absence of an evaluative audience in the present study also helps to provide psychological insight into the cognitive processes influenced by the 5-HTTLPR. According to a recent meta-analysis (25), two principal psychological factors mediate the effect of psychosocial stress on cortisol response: A lack of control and threat to self-esteem or status (social evaluative threat). Compared to the no audience condition, both of these factors were accentuated in the evaluative audience conditions, especially in the negative evaluation condition. Lack of control associated with the inability to avoid social evaluation may be akin to the uncontrollable stressors used in animal studies, which show greater activation of the serotonin system compared to controllable stressors (30).
The association of the 5-HTTLPR and cortisol reactivity seen in the present research is consistent with studies that have focused on amygdala reactivity as an intermediate phenotype associated with the 5-HTTLPR. In a small study comparing the neural correlates of a speech delivered in the presence or absence of an audience, short allele carriers were found to have greater amygdala activation than long allele homozygotes when an audience was present (31). Due to the confines imposed by the scanning environment, the participants in this study could not see the audience, indicating that the mere awareness of being evaluated, irrespective of the reaction of the audience, can lead to differential amygdala activation as a function of 5-HTTLPR genotype. With respect to the present study, although the pressure of social evaluation appears to be a main activator of the cortisol response, the significant association of cortisol reactivity with the 5-HTTLPR in the negative evaluative audience condition may be due to the greater sensitivity of short/short individuals to the threatening, critical, and disapproving feedback in this condition. This would be consistent with functional neuroimaging studies that have shown greater amygdala activity in short allele carriers when viewing threatening faces (32); meta-analysis: (33). As stimulation of the amygdala can trigger HPA axis activation (34) and lesions to the amygdala prevent serotonin neurons from activating the HPA axis (35), the amygdala is likely to be a critical site mediating the greater cortisol reactivity of short/short individuals to the threatening social cues in the negative audience condition in the present study and potentially the threat of social evaluation more generally.
In addition to a role in activating the HPA axis, the serotonin system can also influence feedback inhibition of the HPA axis, particularly via the hippocampus (17). The time point showing the greatest differentiation between short/short individuals and the other groups was 40 minutes after stressor onset (Figure 1), reflecting persisting activation of the HPA axis in short/short individuals relative to those with the long allele. Thus, the heightened cortisol response in these individuals may not only be due to greater HPA axis activation, but also to impaired feedback inhibition. This would be consistent with findings in the serotonin transporter knockout mouse (36) as well as human data showing that serotonin transporter inhibitor treatment influences feedback inhibition (37). Further studies will be needed to address this issue definitively.
Limitations
One limitation of this study is the heterogeneous ethnic composition of the sample, which raises concerns over potential population stratification artifacts (38). However, in spite of the differences in allele frequency between ethnic groups, the qualitative relationship between the 5-HTTLPR and cortisol responses was similar. That the 5-HTTLPR appears to function similarly across ethnic groups is further evidenced by a recent study in an exclusively Korean sample showing that carriers of the short allele were at higher risk for depression when exposed to stressful life events than were long allele homozygotes (39), which is akin to the original finding of Caspi et al., (6) in a Caucasian sample. This is consistent with a meta-analysis of replicated disease-gene association studies that showed the vast majority of associations are similar across ethnic groups (40).
The specific focus on the 5-HTTLPR in this study assumes that it is a functional polymorphism affecting serotonin transporter gene expression. Although expression assays using cells from the periphery indicate this to be the case (4), data from neural tissue has provided less clarity (41, 42). Thus, greater consistency across studies could potentially be obtained with more detailed assessment of variation in the vicinity of the 5-HTTLPR (e.g. rs25531; 43) or the SLC6A4 locus in general (44).
It should be noted that the serotonin system can influence the HPA axis at the hypothalamic, pituitary and adrenal levels (16). Thus, with only cortisol measurements, which reflect the output of the adrenal gland, it is not possible to delineate the specific level at which the 5-HTTLPR is altering responsivity to social stress. Although the most parsimonious explanation is that the 5-HTTLPR is affecting HPA axis activity via influence at the hypothalamic level, it could also be associated with modulation of ACTH release from the pituitary or cortisol release from the adrenal gland.
The findings reported here are based on reactions to laboratory stressors rather than naturalistic stressors. Although this methodological choice confers some potential disadvatanges, it may have benefits as well. The ability to experimentally control the magnitude and duration of the stressor may reduce some of the variance associated with measurement of exposure to stressors in daily life (45).
As this sample excluded individuals with any current or past mental health problems, it precludes making direct applications to clinical disorders for which disease-related processes may influence responsivity of the HPA axis. Nonetheless, the fact that the 5-HTTLPR is associated with cortisol reactivity in this nonclinical sample broadens the significance of the 5-HTTLPR's relation to cortisol reactivity, suggesting a pervasive risk factor for a broad array of stress-related disorders.
Conclusion
We report a significant relationship between the 5-HTTLPR and cortisol response to laboratory stressors. The heightened cortisol reactivity of short/short individuals, especially in the negative audience condition, indicates that these individuals are especially sensitive to social evaluation, and short/short individuals may have greater vulnerability to stress-related mental health outcomes as a result.
Acknowledgments
This research was supported by a grant from the National Institute on Aging (AG030309). This work was supported in part by a NIMH fellowship to B.M.W. as part of the UCLA Health Psychology program as well as by a NARSAD Young Investigator Award. We would also like to thank the UCLA Genotyping facility and particularly Uma Dandekar for her assistance.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Taylor SE. Health Psychology. Boston, Mass.: McGraw-Hill Higher Education; 2009. [Google Scholar]
- 2.Kendler KS, Prescott CA. Genes, environment, and psychopathology : understanding the causes of psychiatric and substance use disorders. New York: Guilford Press; 2006. [Google Scholar]
- 3.Wendland JR, Martin BJ, Kruse MR, Lesch KP, Murphy DL. Simultaneous genotyping of four functional loci of human SLC6A4, with a reappraisal of 5-HTTLPR and rs25531. Mol Psychiatry. 2006;11:224–226. doi: 10.1038/sj.mp.4001789. [DOI] [PubMed] [Google Scholar]
- 4.Lesch KP, Bengel D, Heils A, Sabol SZ, Greenberg BD, Petri S, et al. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science. 1996;274:1527–1531. doi: 10.1126/science.274.5292.1527. [DOI] [PubMed] [Google Scholar]
- 5.Taylor SE, Way BM, Welch WT, Hilmert CJ, Lehman BJ, Eisenberger NI. Early family environment, current adversity, the serotonin transporter promoter polymorphism, and depressive symptomatology. Biological Psychiatry. 2006;60:671–676. doi: 10.1016/j.biopsych.2006.04.019. [DOI] [PubMed] [Google Scholar]
- 6.Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H, et al. Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science. 2003;301:386–389. doi: 10.1126/science.1083968. [DOI] [PubMed] [Google Scholar]
- 7.Roy A, Hu XZ, Janal MN, Goldman D. Interaction between childhood trauma and serotonin transporter gene variation in suicide. Neuropsychopharmacology. 2007;32:2046–2052. doi: 10.1038/sj.npp.1301331. [DOI] [PubMed] [Google Scholar]
- 8.Retz W, Freitag CM, Retz-Junginger P, Wenzler D, Schneider M, Kissling C, et al. A functional serotonin transporter promoter gene polymorphism increases ADHD symptoms in delinquents: Interaction with adverse childhood environment. Psychiatry Research. 2008;158:123–131. doi: 10.1016/j.psychres.2007.05.004. [DOI] [PubMed] [Google Scholar]
- 9.Stein MB, Schork NJ, Gelernter J. Gene-by-environment (serotonin transporter and childhood maltreatment) interaction for anxiety sensitivity, an intermediate phenotype for anxiety disorders. Neuropsychopharmacology. 2007;33:312–319. doi: 10.1038/sj.npp.1301422. [DOI] [PubMed] [Google Scholar]
- 10.Gunthert KC, Conner TS, Armeli S, Tennen H, Covault J, Kranzler HR. Serotonin transporter gene polymorphism (5-HTTLPR) and anxiety reactivity in daily life: A daily process approach to gene-environment interaction. Psychosomatic Medicine. 2007;69:762–768. doi: 10.1097/PSY.0b013e318157ad42. [DOI] [PubMed] [Google Scholar]
- 11.Covault J, Tennen H, Armeli S, Conner TS, Herman AI, Cillessen AHN, et al. Interactive effects of the serotonin transporter 5-HTTLPR polymorphism and stressful life events on college student drinking and drug use. Biological Psychiatry. 2007;61:609–616. doi: 10.1016/j.biopsych.2006.05.018. [DOI] [PubMed] [Google Scholar]
- 12.Uher R, McGuffin P. The moderation by the serotonin transporter gene of environmental adversity in the aetiology of mental illness: review and methodological analysis. Molecular Psychiatry. 2007;13:131–146. doi: 10.1038/sj.mp.4002067. [DOI] [PubMed] [Google Scholar]
- 13.Risch N, Herrell R, Lehner T, Liang KY, Eaves L, Hoh J, et al. Interaction between the serotonin transporter gene (5-HTTLPR), stressful life events, and risk of depression: a meta-analysis. Journal of the American Medical Association. 2009;301:2462–2471. doi: 10.1001/jama.2009.878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dinan TG. Stress: the shared common component in major mental illnesses. European Psychiatry. 2005;20:326–328. doi: 10.1016/s0924-9338(05)80184-1. [DOI] [PubMed] [Google Scholar]
- 15.Van Praag HM. Can stress cause depression? Progress in Neuropsychopharmacology & Biological Psychiatry. 2004;28:891–907. doi: 10.1016/j.pnpbp.2004.05.031. [DOI] [PubMed] [Google Scholar]
- 16.Contesse V, Lefebvre H, Lenglet S, Kuhn JM, Delarue C, Vaudry H. Role of 5-HT in the regulation of the brain-pituitary-adrenal axis: effects of 5-HT on adrenocortical cells. Canadian Journal of Physiology and Pharmacology. 2000;78:967–983. [PubMed] [Google Scholar]
- 17.Lowry CA. Functional subsets of serotonergic neurones: Implications for control of the hypothalamic-pituitary-adrenal axis. Journal of neuroendocrinology. 2002;14:911. doi: 10.1046/j.1365-2826.2002.00861.x. [DOI] [PubMed] [Google Scholar]
- 18.Liposits ZS, Phelix C, Paull WK. Synaptic interaction of serotonergic axons and corticotropin releasing factor (CRF) synthesizing neurons in the hypothalamic paraventricular nucleus of the rat. Histochemistry and Cell Biology. 1987;86:541–549. doi: 10.1007/BF00489545. [DOI] [PubMed] [Google Scholar]
- 19.Heisler LK, Pronchuk N, Nonogaki K, Zhou L, Raber J, Tung L, et al. Serotonin activates the hypothalamic-pituitary-adrenal axis via serotonin 2C receptor stimulation. Journal of Neuroscience. 2007;27:6956–6964. doi: 10.1523/JNEUROSCI.2584-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lanfumey L, Mongeau R, Cohen-Salmon C, Hamon M. Corticosteroid–serotonin interactions in the neurobiological mechanisms of stress-related disorders. Neuroscience and Biobehavioral Reviews. 2008;32:1174–1184. doi: 10.1016/j.neubiorev.2008.04.006. [DOI] [PubMed] [Google Scholar]
- 21.Van Cauter E, Leproult R, Kupfer DJ. Effects of gender and age on the levels and circadian rhythmicity of plasma cortisol. Journal of Clinical Endocrinology & Metabolism. 1996;81:2468–2473. doi: 10.1210/jcem.81.7.8675562. [DOI] [PubMed] [Google Scholar]
- 22.Watson D, Clark L, Tellegen A. Development and validation of brief measures of positive and negative affect: The PANAS scales. Journal of Personality and Social Psychology. 1988;54:1063–1070. doi: 10.1037//0022-3514.54.6.1063. [DOI] [PubMed] [Google Scholar]
- 23.Kirschbaum C, Pirke K, Hellhammer D. The ‘Trier Social Stress Test’-a tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology. 1993;28:76–81. doi: 10.1159/000119004. [DOI] [PubMed] [Google Scholar]
- 24.Kirschbaum C, Pirke KM, Hellhammer DH. The ‘Trier Social Stress Test’-a tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology. 1993;28:76–81. doi: 10.1159/000119004. [DOI] [PubMed] [Google Scholar]
- 25.Dickerson SS, Kemeny ME. Acute stressors and cortisol responses: A theoretical integration and synthesis of laboratory research. Psychological Bulletin. 2004;130:355–391. doi: 10.1037/0033-2909.130.3.355. [DOI] [PubMed] [Google Scholar]
- 26.Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 2005;21:263–265. doi: 10.1093/bioinformatics/bth457. [DOI] [PubMed] [Google Scholar]
- 27.Gelernter J, Kranzler H, Cubells JF. Serotonin transporter protein (SLC6A4) allele and haplotype frequencies and linkage disequilibria in African-and European-American and Japanese populations and in alcohol-dependent subjects. Human Genetics. 1997;101:243–246. doi: 10.1007/s004390050624. [DOI] [PubMed] [Google Scholar]
- 28.Gotlib IH, Joormann J, Minor KL, Hallmayer J. HPA axis reactivity: A mechanism underlying the associations among 5-HTTLPR, stress, and depression. Biological Psychiatry. 2008;63:847–851. doi: 10.1016/j.biopsych.2007.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Jabbi M, Kema IP, van der Pompe G, te Meerman GJ, Ormel J, den Boer JA. Catechol-o-methyltransferase polymorphism and susceptibility to major depressive disorder modulates psychological stress response. Psychiatric Genetics. 2007;17:183–193. doi: 10.1097/YPG.0b013e32808374df. [DOI] [PubMed] [Google Scholar]
- 30.Maier SF, Watkins LR. Stressor controllability and learned helplessness: the roles of the dorsal raphe nucleus, serotonin, and corticotropin-releasing factor. Neuroscience and Biobehavioral Reviews. 2005;29:829–841. doi: 10.1016/j.neubiorev.2005.03.021. [DOI] [PubMed] [Google Scholar]
- 31.Furmark T, Tillfors M, Garpenstrand H, Marteinsdottir I, Långström B, Oreland L, et al. Serotonin transporter polymorphism related to amygdala excitability and symptom severity in patients with social phobia. Neuroscience Letters. 2004;362:189–192. doi: 10.1016/j.neulet.2004.02.070. [DOI] [PubMed] [Google Scholar]
- 32.Hariri AR, Drabant EM, Munoz KE, Kolachana BS, Mattay VS, Egan MF, et al. A susceptibility gene for affective disorders and the response of the human amygdala. Arch Gen Psychiatry. 2005;62:146–152. doi: 10.1001/archpsyc.62.2.146. [DOI] [PubMed] [Google Scholar]
- 33.Munafò MR, Brown SM, Hariri AR. Serotonin transporter (5-HTTLPR) genotype and amygdala activation: a meta-analysis. Biological Psychiatry. 2008;63:852–857. doi: 10.1016/j.biopsych.2007.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Feldman S, Conforti N, Siegel RA. Adrenocortical responses following limbic stimulation in rats with hypothalamic deafferentations. Neuroendocrinology. 1982;35:205. doi: 10.1159/000123382. [DOI] [PubMed] [Google Scholar]
- 35.Weidenfeld J, Newman ME, Itzik A, Gur E, Feldman S. The amygdala regulates the pituitary-adrenocortical response and release of hypothalamic serotonin following electrical stimulation of the dorsal raphe nucleus in the rat. Neuroendocrinology. 2002;76:63–69. doi: 10.1159/000064430. [DOI] [PubMed] [Google Scholar]
- 36.Jiang X, Wang J, Luo T, Li Q. Impaired hypothalamic-pituitary-adrenal axis and its feedback regulation in serotonin transporter knockout mice. Psychoneuroendocrinology. 2009;34:317–331. doi: 10.1016/j.psyneuen.2008.09.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Pariante CM, Lightman SL. The HPA axis in major depression: classical theories and new developments. Trends in Neurosciences. 2008;31:464–468. doi: 10.1016/j.tins.2008.06.006. [DOI] [PubMed] [Google Scholar]
- 38.Hutchison KE, Stallings M, McGeary J, Bryan A. Population stratification in the candidate gene study: fatal threat or red herring? Psychol Bull. 2004;130:66–79. doi: 10.1037/0033-2909.130.1.66. [DOI] [PubMed] [Google Scholar]
- 39.Kim JM, Stewart R, Kim SW, Yang SJ, Shin IS, Kim YH, et al. Interactions between life stressors and susceptibility genes (5-HTTLPR and BDNF) on depression in Korean elders. Biological Psychiatry. 2007;62:423–428. doi: 10.1016/j.biopsych.2006.11.020. [DOI] [PubMed] [Google Scholar]
- 40.Ioannidis JPA, Ntzani EE, Trikalinos TA. ‘Racial’ differences in genetic effects for complex diseases. Nature genetics. 2004;36:1312–1318. doi: 10.1038/ng1474. [DOI] [PubMed] [Google Scholar]
- 41.Lim JE, Papp A, Pinsonneault J, Sadee W, Saffen D. Allelic expression of serotonin transporter (SERT) mRNA in human pons: lack of correlation with the polymorphism SERTLPR. Mol Psychiatry. 2006;11:649–662. doi: 10.1038/sj.mp.4001797. [DOI] [PubMed] [Google Scholar]
- 42.Mann JJ, Huang YY, Underwood MD, Kassir SA, Oppenheim S, Kelly TM, et al. A serotonin transporter gene promoter polymorphism (5-HTTLPR) and prefrontal cortical binding in major depression and suicide. Arch Gen Psychiatry. 2000;57:729–738. doi: 10.1001/archpsyc.57.8.729. [DOI] [PubMed] [Google Scholar]
- 43.Hu XZ, Lipsky RH, Zhu G, Akhtar LA, Taubman J, Greenberg BD, et al. Serotonin transporter promoter gain-of-function genotypes are linked to obsessive-compulsive disorder. The American Journal of Human Genetics. 2006;78:815–826. doi: 10.1086/503850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wendland JR, Moya PR, Kruse MR, Ren-Patterson RF, Jensen CL, Timpano KR, et al. A novel, putative gain-of-function haplotype at SLC6A4 associates with obsessive-compulsive disorder. Human Molecular Genetics. 2008;17:717–723. doi: 10.1093/hmg/ddm343. [DOI] [PubMed] [Google Scholar]
- 45.Monroe SM, Reid MW. Gene-environment interactions in depression research: Genetic polymorphisms and life-stress polyprocedures. Psychological Science. 2008;19:947–956. doi: 10.1111/j.1467-9280.2008.02181.x. [DOI] [PubMed] [Google Scholar]