

Abstract
Neuroligins (NLs) are a family of neural cell-adhesion molecules that are involved in excitatory/inhibitory synapse specification. Multiple members of the NL family (including NL1) and their binding partners have been linked to cases of human autism and mental retardation. We have now characterized NL1-deficient mice in autism- and mental retardation-relevant behavioral tasks. NL1 knock-out (KO) mice display deficits in spatial learning and memory that correlate with impaired hippocampal long-term potentiation. In addition, NL1 KO mice exhibit a dramatic increase in repetitive, stereotyped grooming behavior, a potential autism-relevant abnormality. This repetitive grooming abnormality in NL1 KO mice is associated with a reduced NMDA/AMPA ratio at corticostriatal synapses. Interestingly, we further demonstrate that the increased repetitive grooming phenotype can be rescued in adult mice by administration of the NMDA receptor partial coagonist d-cycloserine. Broadly, these data are consistent with a role of synaptic cell-adhesion molecules in general, and NL1 in particular, in autism and implicate reduced excitatory synaptic transmission as a potential mechanism and treatment target for repetitive behavioral abnormalities.
Introduction
Neuroligins (NLs) are a family of neuronal postsynaptic cell adhesion molecules (Ichtchenko et al., 1995, 1996). NLs are differentially localized to excitatory and inhibitory synapses (Ichtchenko et al., 1995, 1996; Song et al., 1999; Graf et al., 2004; Varoqueaux et al., 2004; Budreck and Scheiffele, 2007). Neuroligin-1 (NL1) is enriched preferentially at excitatory synapses (Song et al., 1999), neuroligin-2 (NL2) is enriched at inhibitory synapses (Graf et al., 2004; Varoqueaux et al., 2004), and neuroligin-3 (NL3) appears to be present at both (Budreck and Scheiffele, 2007). Expression of NLs in vitro has been shown to induce presynaptic specializations and increase synaptic density (Scheiffele et al., 2000; Dean et al., 2003; Graf et al., 2004; Prange et al., 2004; Boucard et al., 2005; Chih et al., 2005; Chubykin et al., 2005, 2007; Levinson et al., 2005; Nam and Chen, 2005). However, the synapse-increasing activities of NLs in culture do not reflect a requirement for NLs in initial synapse formation in vivo but rather a role of NLs in synapse specification and modulation (Varoqueaux et al., 2006; Chubykin et al., 2007).
Understanding NL function in vivo is not only critical for a basic understanding of synapse function but is also relevant to human autism spectrum disorders (ASDs). Indeed, mutations in members of the NL family and its associated binding partners, including the NL1 binding partners neurexin-1 and shank3, have been implicated in human autism and mental retardation (Jamain et al., 2003; Zoghbi, 2003; Chih et al., 2004; Comoletti et al., 2004; Laumonnier et al., 2004; Yan et al., 2005; Feng et al., 2006; Durand et al., 2007; Szatmari et al., 2007; Yan et al., 2008a,b). Chromosomal rearrangements in regions that harbor the NL1 and NL2 genes and single nucleotide polymorphisms in the gene encoding NL1 have been associated directly with human ASDs (Konstantareas and Homatidis, 1999; Zoghbi, 2003; Yan et al., 2004; Ylisaukko-oja et al., 2005; Südhof, 2008). More recently, a genome-wide copy number variation analysis also implicated NL1 among several candidate genes in ASD susceptibility (Glessner et al., 2009), further suggesting a direct link between NL1 and human autism.
In light of the link between NLs and autism, we predicted that NL1 knock-out (KO) mice might exhibit autism- or mental retardation-relevant behavioral abnormalities. Consistent with our hypothesis, NL1 KO mice displayed deficits in hippocampus-dependent spatial memory along with impaired hippocampal long-term potentiation. NL1 KO mice also exhibited increased repetitive grooming behavior, which may be relevant to the increased repetitive behavior seen in autism (American Psychiatric Association, 2000), along with a reduced NMDA/AMPA ratio at corticostriatal synapses. Furthermore, we demonstrate that the autism-related repetitive grooming phenotype can be rescued by systemic d-cycloserine (DCS) in adult mice. Overall, these data are consistent with the hypothesis that NL1 dysfunction can lead to autism- and mental retardation-related behavioral abnormalities in part via alteration of NMDA receptor (NMDAR) function.
Materials and Methods
Genetic manipulations
NL1 KO mice were generated as described previously (Varoqueaux et al., 2006). To reduce genetic and experimental variability, the NL1 mice studied were sex-matched, littermate products of heterozygous mating on a hybrid 129S6/SvEvTac/C57BL/6J background. In all studies, experimenters were blind to genotype of the animals.
Western blot
Protein compositions were determined by immunoblotting on brain tissues homogenized in PBS, 10 mm EDTA, and proteinase inhibitors from four pairs of P40 littermate mice per genotype. Proteins (40 μg) were loaded per lane and blotted with antibodies for synaptic proteins and internal controls (β-actin or GDP dissociation inhibitor). Blots were reacted with 125I-labeled secondary antibodies, followed by PhosphoImager (STORM 860; GE Healthcare) detection.
Morphological analyses
NL1 KO and wild-type (WT) littermate control mice were anesthetized and perfusion fixed with 4% fresh paraformaldehyde and cryoprotected with 30% sucrose. Sections (30 μm) were blocked with 3% goat serum/0.3% Triton X-100 in PBS and incubated with anti-synaptophysin monoclonal antibody (Millipore Corporation), anti-vesicular glutamate transporter 1 (vGlut1) monoclonal antibody (Synaptic Systems), and/or anti-vesicular GABA transporter (VGAT) polyclonal antibody (Millipore Corporation) overnight at 4°C, followed by incubation with Alexa Fluor 488 or 633 goat anti-mouse IgG (Invitrogen). Sections were transferred onto SuperFrost slides and mounted under glass coverslips with Vectashield with 4′,6′-diamidino-2-phenylindole (Vector Laboratories). Sections of the CA1 and CA3 subfields of the hippocampus were imaged with a Leica TCS2 laser-scanning confocal microscope at 63× and magnified fivefold. For each experimental series, all images were acquired with identical settings for laser power, photomultiplier gain, and offset with a pinhole diameter. Images were imported into NIH ImageJ software, and synaptic densities and sizes were analyzed under fixed thresholds across all slides. Thresholds were chosen within the range that allowed outlining as many immunopositive puncta as possible throughout all images. The number and size of puncta were detected using the “analyze particle” module of the program. The average number and size of puncta were normalized with data from wild type to determine synaptic density and size, respectively. Statistical significance was determined by Student's t test. All of the data shown are means ± SEM.
Electrophysiological techniques
Hippocampus.
Transverse hippocampal slices were prepared from 4- to 8-week- old mice as described previously (Volk et al., 2007). In brief, mice were anesthetized with isoflurane and decapitated, and the brain was quickly isolated into ice-cold dissecting solution (in mm: 222 sucrose, 11 glucose, 26 NaHCO3, 1 NaH2PO4, 3 KCl, 7 MgCl2, and 0.5 CaCl2). Slices, 400 μm thick, were made using a Leica VT1200s and allowed to recover for at least 1.5 h in artificial CSF (ACSF) [in mm: 126 NaCl, 3 KCl, 1.25 NaH2PO4, 1.3 MgSO4 26 NaHCO3, 25 d-glucose, and 2.5 CaCl2, pH 7.4 (saturated with 95%O2/5%CO2)] before recording. The slices were then placed in a submerged chamber at 28–30°C and allowed to equilibrate for 15 min before recording. Extracellular recordings were performed in area CA1 of the hippocampus. Recording electrodes (3–5 MΩ) were filled with ACSF and placed in the stratum radiatum of area CA1. Basal field EPSPs (fEPSPs) were evoked at 0.067 Hz with a concentric bipolar electrode placed in the stratum radiatum. Input–output curves were constructed from the average of 10 traces at each stimulus intensity with the amplitude of the presynaptic fiber volley measured relative to the slope of the fEPSP.
For paired-pulse and long-term potentiation (LTP) protocols, stimulus strengths were adjusted to produce responses 40% of maximum fEPSP. The LTP protocol used was a theta burst stimulation (TBS) pattern consisting of five bursts of four pulses at 100 Hz with an interburst interval of 0.2 s. Baseline and post-induction responses were sampled at 0.033 Hz. Baseline recordings for LTP experiments were performed for >10 min, and slices were rejected if baseline was unstable.
Corticostriatal synapses.
Horizontal-oblique slices were prepared and recordings were performed as described previously (Ding et al., 2008). Briefly, wild-type and NL1 KO littermate mice were anesthetized with isoflurane and decapitated, and the brain was quickly isolated into dissecting solution. Slices 350–400 μm thick were prepared at ice-cold temperature from juvenile mice at postnatal day 15 (P15) to P38. Average age was 26.9 ± 1.8 d for wild-type and 26.1 ± 1.6 d for NL1 KO. The dissecting solution contained the following (in mm): 54 NaCl, 100 sucrose, 3 KCl, 1.25 NaH2PO4, 10 Mg Cl2, 26 NaHCO3, 10 dextrose, and 0.5 CaCl2. The bathing solution contained the following (in mm): 126 NaCl, 3 KCl, 1.25 NaH2PO4, 1 MgCl2, 26 NaHCO3, 10 dextrose, and 2 CaCl2 (saturated with 95% O2/5% CO2). Slices were incubated in the bathing solution at 32°C for 1 h. Afterward, slices were kept at room temperature until transferred to a submersion-type recording chamber. Whole-cell patch recordings on medium spiny neurons were done using micropipettes (3–5 MΩ) made from 1.1/1.5 mm borosilicate glass (Sutter Instruments). Recording pipettes were filled with the following solution (in mm): 117 Cs-methanesulphonate, 15 CsCl, 8 NaCl, 10 tetraethylammonium-Cl, QX-314 [N-(2,6-dimethylphenylcarbamoylmethyl)triethylammonium chloride], 0.2 EGTA, 2 ATP-Mg, 0.3 GTP, and 10 HEPES-CsOH, pH 7.25 (290–295 mOsm). A theoretical junction potential of 12, calculated using the corresponding function in Clampfit, was used to correct voltages post hoc. Access resistance was frequently checked to be <25 MΩ and stable (<20% of variability). Recordings were obtained using the 700B Multiclamp amplifier (Molecular Devices), and neurons were visualized using a Carl Zeiss Axioexaminer D1 scope equipped with infrared differential interference contrast visualization through microscope and a CCD camera and DOT optics.
EPSCs were evoked by stimulating corticostriatal projections with 0.2 ms current injections (commonly between 0.1 and 1 mA) at the boundary between neocortical layer VI and the corpus callosum by using concentric bipolar electrodes (FHC Inc.) and an A365 battery-driven stimulus isolator (WPI). Evoked NMDAR/AMPAR ratios were determined using standard, published methods (Myme et al., 2003). In the presence of picrotoxin, AMPAR currents are measured at the peak and at a voltage of −80 mV, at which most NMDAR currents are expected to be blocked by Mg2+. In the same cell, NMDAR currents are measured in a 2 ms window 50 ms after spike onset at a voltage of +40 mV.
All responses were digitized at 10 kHz and filtered at 1 kHz. Data were analyzed offline using pClamp and Microsoft Excel. Student's t test was used to evaluate significance of all analyses. Experimenters were blind to genotype.
Behavioral overview
Mice were age/sex-matched littermate progeny of heterozygous/heterozygous (NL1 KO) matings tested behaviorally in four groups. Experimenters were blind to genotype. The first cohort of mice for behavioral studies included 23 NL1 KO littermate pairs (total of 46 mice), except when noted. For shock threshold, nesting, and visible water maze, there were 11 littermate pairs (22 mice total), and, for the nonsocial test of olfaction, there were 10 littermate pairs (20 mice total), because some of the mice were removed from the original cohort for histological studies. Less stressful behaviors were tested first, with more stressful procedures at the end. The order of tests for the first cohort of mice was as follows: locomotor, dark/light box, open field, elevated plus maze, accelerating rotarod, social interaction with a juvenile, social learning, social versus inanimate preference test, preference for social novelty test, social interaction with an adult caged conspecific, fear conditioning, Morris water maze, prepulse inhibition, startle amplitude, nesting, olfaction for a nonsocial stimulus (peanut butter cookie), and shock threshold. A second cohort of mice (22 littermate pairs, 44 mice total) was tested for grooming, interaction with a social smell, and marble burying. A third cohort of mice (17 littermate pairs, 34 mice total) was combined with the second cohort to test grooming behavior after administration of d-cycloserine (or vehicle). A fourth cohort of mice (21 littermate pairs, 42 mice total) was reexamined in anxiety-related tasks in a different order (elevated plus maze, dark/light, open field) and then tested for hotplate sensitivity.
The dark/light, open field, and elevated plus mazes were used as measures of anxiety-like behavior. Social interaction with a juvenile, social learning, the social versus inanimate preference test, the preference for social novelty test, and social interaction with an adult caged conspecific were all used as measures of social behavior, and nesting behavior was tested because of its association with social and affiliative behaviors. Interaction with a social smell and olfaction for a nonsocial stimulus (peanut butter cookie) were both used as controls to test for normal olfactory function. Locomotor activity, startle amplitude, prepulse inhibition, shock threshold, and hotplate sensitivity were tested to examine basic neurologic function, including sensitivity to auditory and sensory stimuli. Grooming was observed to examine repetitive, stereotyped behavior. The Morris water maze was conducted to examine spatial learning and memory, and fear conditioning was conducted to examine fear learning and memory.
Within each cohort, all mice ranged from 2 to 8 months of age during the behavioral testing. Mice were moved within the animal facility to the testing room and allowed to habituate to the new location for at least 1 h before behavioral testing. All statistical analyses were conducted using Statistica software (StatSoft), and significance was taken as p < 0.05 for all experiments.
Morris water maze
The Morris water maze and visible platform tests were performed essentially as described previously (Powell et al., 2004) except a probe trial was performed only on day 12. Briefly, a 4-feet-diameter, white, plastic, circular pool was filled to a depth of 13 inches with 22 ± 1°C water made opaque with gothic white, nontoxic, liquid tempera paint in a room with prominent extramaze cues. Mice were placed in one of four starting locations facing the pool wall and allowed to swim until they found a 10-inch-diameter, white platform submerged by 1 cm, or until a maximum of 60 s had elapsed. On finding the platform, mice remained on the platform for 15 s before being removed to the home cage. If mice did not find the platform within 60 s, they were guided to the platform by the experimenter where they remained for 15 s before being removed to the home cage. Latency to reach the platform, distance traveled to reach the platform, swim speed, and percentage thigmotaxis (time spent near the wall of the pool) were measured using automated video-tracking software from Noldus (Ethovision 2.3.19). Mice were trained with 4 trials/d with an intertrial interval of 1–1.5 min for 11 consecutive days between 8:00 A.M. and 1:00 P.M. A probe trial (free swim with the submerged platform removed) was performed as the first trial of the day on day 12. The percentage time spent in the target quadrant and the number of platform location crossings was calculated using Ethovision 2.3.19. Percentage time spent in all quadrants, latency to platform, distance to platform, swim speed, and percentage thigmotaxis were analyzed with a three-way mixed ANOVA. Training in the visible water maze task was conducted in the same manner as the main Morris water maze except that a visible cue (black foam cube) was placed on top of the platform, mice were placed in the same location for each trial, and the platform was moved to a new, random location for each trial. Mice were trained with 6 trials/d for 2 consecutive days, and the latency to reach the visible platform was analyzed with a three-way mixed ANOVA.
Social behavior
Direct social interaction with a juvenile took place in a novel, empty, clear, plastic mouse cage under red light, as described previously (Kwon et al., 2006; Tabuchi et al., 2007). After a 15 min habituation in the dark, the experimental and target mice were placed in the neutral cage for 2 min and allowed to directly interact. Time spent interacting with the juvenile was scored by an observer blind to genotype. Social learning was assessed 3 d later by allowing mice to interact with the same juvenile for an additional 2 min. Again, time spent interacting with the juvenile was scored. Data were analyzed with a three-way mixed ANOVA.
Caged adult social interaction tests were performed in a 48 × 48 cm2 white plastic arena under red light using a 6.0 × 9.5 cm wire mesh rectangular cage containing an unfamiliar adult mouse, allowing olfactory, visual, and minimal tactile interaction (Kwon et al., 2006; Tabuchi et al., 2007). Mice were first placed in the arena for 5 min with an empty wire mesh cage. Then mice were allowed to interact with a novel caged social target for another 5 min. Time spent in the interaction zone was obtained using Noldus software (Ethovision 2.3.19). The box was wiped with 70% ethanol and air dried between mice. Data were analyzed with a three-way mixed ANOVA.
Social versus inanimate preference and preference for social novelty analyses were modified from previous descriptions (Moy et al., 2004; Nadler et al., 2004) as described in detail previously (Kwon et al., 2006; Tabuchi et al., 2007). Data were analyzed with a three-way mixed ANOVA. Nesting behavior was performed as described previously (Lijam et al., 1997) and analyzed with a three-way mixed ANOVA.
Interaction with a social smell was performed similar to interaction with a caged adult. Initially, mice were placed in a 48 × 48 cm2 white plastic arena for 5 min with a slide containing a nonsocial smell (rubbed with distilled water). Immediately after, mice were allowed to interact with a slide containing a “social” smell (slide rubbed on the anogenital region of an unfamiliar C57BL/6J WT mouse) for another 5 min. Time in the interaction zone was obtained using Noldus software (Ethovision 2.3.19). The box was wiped with 70% ethanol and air dried between mice. Data were analyzed with a three-way mixed ANOVA.
Olfaction for a nonsocial stimulus was measured as described previously (Moretti et al., 2005) except animals were food deprived overnight before the test and a peanut butter cookie was used as the olfactory treat. Data were analyzed with a two-way ANOVA.
Grooming
Mice were habituated to a novel home cage for 10 min. Immediately thereafter, total time spent grooming the face, head, or body was measured for 10 min. Grooming behavior was analyzed using a two-way ANOVA.
Marble burying
Similar to a previous description (Deacon, 2006), empty home cages were filled with bedding up to 5 cm from the cage floor, and 20 black marbles were placed evenly throughout the cage. Mice were allowed to freely explore the cage (and marbles) for 30 min, and, afterward, the number of successfully buried marbles was counted. A marble was defined as “buried” when <25% of the marble was visible. Data were analyzed with a two-way ANOVA.
Fear conditioning
Cued and contextual fear conditioning was performed essentially as described previously (Powell et al., 2004). Briefly, mice were habituated to the shock context for 2 min, during which the level of “pretraining” freezing was measured. Then, a 30 s, 90 dB tone coterminating with a 2 s, 0.5 mA footshock was delivered twice with a 1 min interstimulus interval. Mice remained in the context for 2 min before returning to their home cage. Freezing behavior (motionless except respirations) was monitored at 10 s intervals by an observer blind to genotype. To test for contextual learning 24 h after training, mice were placed into the same training context for 5 min and scored for freezing behavior every 10 s. To assess cue-dependent fear conditioning, mice were placed in a novel environment 3 h after the context test. Freezing behavior was assessed during a 3 min baseline, followed by a 3 min presentation of the tone. Cue-dependent fear conditioning was determined by subtracting baseline freezing from freezing during the tone. Both cued and contextual fear conditioning data were analyzed with a two-way ANOVA.
Hotplate sensitivity
Mice were placed on a black, anodized, constant-temperature plate of 52°C (IITC model 39 hotplate) covered with a Plexiglas enclosure. Latency to lick or shake the hindpaw was measured, and mice were removed after the first lick or shake of a hindpaw or after 30 s if no response was elicited. The plate was cleaned with water between mice and allowed to return to baseline temperature. Data were analyzed with a two-way ANOVA.
Footshock sensitivity
Pain sensitivity during footshock was measured as described previously (Powell et al., 2004). Briefly, mice were placed in a conditioning chamber and allowed to habituate for 2 min. Then, a series of 2 s footshocks was delivered; the initial shock was delivered at 0.05 mA, and the current increased by 0.05 mA every trial with a 20 s intertrial interval. The current required to elicit flinching, jumping, and vocalizing was recorded by an observer. Data were analyzed with a two-way ANOVA.
Grooming plus d-cycloserine or vehicle
The same protocol for assessing grooming behavior was used as described above except that half the mice (NL1 KO and WT) were injected with DCS (20 mg/kg, i.p.; Sigma) and the other half were injected with vehicle (saline) 20 min before placement in the novel home cage. Treatment groups were counterbalanced for the total time spent grooming during a baseline observation. Data were analyzed with a three-way ANOVA.
Results
NL1 deletion results in a decrease in neurexin levels
To determine whether NL1 deletion causes changes in other synaptic proteins, we examined protein levels of 26 presynaptic and postsynaptic proteins in brains of NL1 KO and littermate control mice by Western blot (Table 1). For the majority of proteins examined, these experiments revealed only subtle changes in the gross levels of synaptic markers as a result of NL1 deletion. In particular, it should be noted that no significant change was observed in NMDAR subunit protein levels, which is of interest given the decreased NMDA/AMPA ratio observed in NL1 KO mice (Chubykin et al., 2007; Kim et al., 2008b). However, NL1 KO mice exhibited a 30% increase in the expression of NL3 as well as a 20% decrease in both α and β neurexin levels (Table 1). NL1 KO mice also exhibited a significant increase in the expression of synapsin 1a as well as decreases of ∼15–20% in the levels of several presynaptic proteins: liprin, CSP, and munc-18 (Table 1). Given the direct interactions between NL1 and neurexins as well as the link between neurexin-1 copy number and human cases of autism, the reduction of neurexin levels in NL1 KO mice further increases their potential relevance to autism.
Table 1.
Synaptic protein composition in NL1 KO brain
| % | SEM | p value | |
|---|---|---|---|
| β-Catenin | 106.6754 | 7.863655 | 0.5225598 |
| CaMKIIα | 89.19236 | 9.984238 | 0.5153004 |
| Complexin1 | 106.1328236 | 6.209750233 | 0.428415248 |
| CSP | 86.68285 | 4.591786 | 0.0326469* |
| GABAα-Rα | 96.37907 | 8.532568 | 0.7916562 |
| GluR1 | 103.8289703 | 8.288285636 | 0.813943664 |
| Liprin | 88.46195 | 2.025309 | 0.02345723* |
| Munc-18 | 78.83607 | 2.23259 | 0.003578847** |
| NL3 | 129.3825882 | 6.80079828 | 0.02431589* |
| NL2 | 104.98536 | 7.8880185 | 0.8019042 |
| NR1 | 78.09982304 | 19.44676586 | 0.45496166 |
| NR2A | 89.75655 | 8.760029 | 0.388649 |
| NR2B | 93.48802144 | 6.362418868 | 0.509075447 |
| NSF | 91.73566 | 7.769968 | 0.403433 |
| Nrx-α | 82.330652 | 3.2919612 | 0.0354927* |
| Nrx-β | 80.442576 | 5.5239526 | 0.0325197* |
| PSD-95 | 114.2735874 | 8.599706997 | 0.240376141 |
| Rab3A | 76.61192 | 8.136521 | 0.3786824 |
| Rabphilin | 94.49966 | 5.816795 | 0.5273652 |
| SCAMP | 106.7596 | 12.57834 | 0.7664692 |
| SNAP-25 | 98.74584 | 2.942259 | 0.7681499 |
| Synaptobrevin-2 | 107.7806 | 7.415825 | 0.6034307 |
| Synapsin-1a/b | 118.6078871 | 5.154650933 | 0.038798877* |
| Synapsin-2b | 109.4151858 | 8.029006022 | 0.440344385 |
| Synaptophysin | 102.6427 | 3.131189 | 0.5091538 |
| Synaptotagmin-1 | 99.25104 | 1.845906 | 0.794641 |
| VAChT | 121.0259 | 7.899216 | 0.2178337 |
| VGAT | 102.2081 | 16.37049 | 0.9298164 |
| VGlut1 | 95.76447 | 12.95471 | 0.8080348 |
Protein levels (percentage of wild-type) in P40 NL1 KO brain homogenate, SEM, and p value (Student's t test) are listed (wild type, n = 4; KO, n = 4). Nrx, Neurexin.
*p < 0.05;
**p < 0.01 compared to WT.
NL1 KO mice do not exhibit global behavioral deficits
Because NL1 is a ubiquitously expressed, excitatory synaptic cell adhesion molecule, one might expect widespread CNS dysfunction; on the contrary, NL1 KO mice showed normal anxiety-like behavior, locomotor activity, motor coordination/learning, auditory startle responses, and sensitivity to sensory stimuli. Anxiety-like behaviors were normal on three anxiety tests: elevated plus maze (supplemental Fig. 1A,B, available at www.jneurosci.org as supplemental material), dark/light box (supplemental Fig. 1C,D, available at www.jneurosci.org as supplemental material), and open field (supplemental Fig. 1E,F, available at www.jneurosci.org as supplemental material). Indeed, these tests of anxiety were repeated later in a naive cohort of 21 littermate pairs in a completely different testing order (elevated plus maze, dark/light box, followed by open field) with the same lack of effect observed (data not shown). Locomotor activity was also normal in NL1 KO mice when tested under four different conditions. First, locomotor activity in an open-field arena was normal in NL1 KO mice (supplemental Fig. 2A,B, available at www.jneurosci.org as supplemental material). This was also true of locomotor activity in the dark/light apparatus (supplemental Fig. 2D, available at www.jneurosci.org as supplemental material), with only a small decrease in distance moved in the elevated plus maze (supplemental Fig. 2C, available at www.jneurosci.org as supplemental material) [two-way ANOVA, main effect of genotype (between-subjects factor), F(1,42) = 4.09, p < 0.049; main effect of sex (between-subjects factor), F(1,42) = 0.70, p = 0.41; genotype × sex interaction, F(1,42) = 0.42, p = 0.52]. NL1 KO mice also exhibited normal locomotor activity and habituated at a similar rate compared with controls in a 2 h novel-cage locomotor task (supplemental Fig. 2E, available at www.jneurosci.org as supplemental material). For the complete statistical analyses of all behavioral tests, see Table 2.
Table 2.
Additional statistical analyses for behavioral tests
| n | Test variant | Parameter | Comparison | Results |
|---|---|---|---|---|
| Tests of sensory sensitivity(Fig. 1) | ||||
| 11 pairs | Shock threshold | Jump | Genotype and sex | Two-way ANOVA: genotype, F(1,18)= 0.94, p = 0.34; sex, F(1,18)= 0.26, p = 0.62; genotype × sex interaction, F(1,18)= 0.61, p = 0.45 |
| Flinch | Genotype and sex | Two-way ANOVA: genotype, F(1,18)= 0.081, p = 0.78; sex, F(1,18)= 0.39, p = 0.54; genotype × sex interaction, F(1,18)= 1.17, p = 0.29 | ||
| Three-box social interaction test (Fig. 2) | ||||
| 23 pairs | Social versus inanimate preference | Time interaction | Genotype, sex, and interaction target | Three-way mixed ANOVA: genotype, F(1,42)= 1.56, p = 0.22; sex, F(1,42)= 2.05, p = 0.16; target, F(1,42)= 3.11, p = 0.08; genotype × sex interaction, F(1,42)= 1.23, p = 0.27; genotype × target interaction, F(1,42)= 0.002, p = 0.97; genotype × sex × target interaction, F(1,42)= 0.06, p = 0.81 |
| 23 pairs | Familiar versus novel preference | Time interaction | Genotype, sex, and interaction target | Three-way mixed ANOVA: genotype, F(1,42)= 0.74, p = 0.39; sex, F(1,42)= 0.46, p = 0.50; target, F(1,42)= 10.59, p < 0.003; genotype × sex interaction, F(1,42)= 1.06, p = 0.31; genotype × target interaction, F(1,42)= 0.72, p = 0.40; sex × target interaction, F(1,42)= 0.03, p = 0.87; genotype × sex × target interaction, F(1,42)= 0.55, p = 0.46 |
| Social interaction with a juvenile (Fig. 2) | ||||
| 23 pairs | Social interaction with a juvenile | Time interaction | Genotype, sex, and trial | Three-way mixed ANOVA: genotype, F(1,42)= 0.74, p = 0.39; sex, F(1,42)= 4.47, p < 0.041; trial, F(1,42)= 13.61, p < 0.0007; genotype × sex interaction, F(1,42)= 0.34, p = 0.56; genotype × trial interaction, F(1,42)= 0.08, p = 0.78; sex × trial interaction, F(1,42)= 3.37, p = 0.07; genotype × sex × trial interaction, F(1,42)= 1.47, p = 0.23 |
| Genotype (WT vs KO) within each trial | Planned comparisons: trial 1 only, F(1,42)= 0.53, p = 0.47; trial 2 only, F(1,42)= 0.80, p = 0.38 | |||
| Trial (1st vs 2nd) within each genotype | Planned comparisons: WT only, F(1,42)= 5.81, p < 0.021; NL1 KO only, F(1,42)= 7.88, p < 0.008 | |||
| Morris water maze (Fig. 3; supplemental Fig. 2, available at www.jneurosci.org as supplemental material) | ||||
| 23 pairs | Initial training | Latency to platform | Genotype, sex, and day | Three-way mixed ANOVA: genotype, F(1,42)= 0.54, p = 0.47; sex, F(1,42)= 0.21, p = 0.65; day, F(10,420) = 8.34, p < 0.000001; genotype × sex interaction, F(1,42)= 0.11, p = 0.74; genotype × day interaction, F(10,420)= 0.43, p = 0.93; sex × day interaction, F(10,420) = 0.43, p = 0.11; genotype × sex × day interaction, F(10,400) = 0.94, p = 0.50 |
| Percentage thigmotaxis | Genotype, sex, and day | Three-way mixed ANOVA: genotype, F(1,42)= 0.10, p = 0.95; sex, F(1,42)=0.055, p = 0.82; day, F(10,420) = 11.89, p < 0.000001; genotype × sex interaction, F(1,42)= 0.000001, p = 0.99; genotype × day interaction, F(10,420) = 0.83, p = 0.60; sex × day interaction, F(10,420) = 0.64, p = 0.78; genotype × sex × day interaction, F(10,420) = 0.38, p = 0.96 | ||
| 23 pairs | Initial probe | Percentage time in quadrant | Genotype, sex, and quadrant | Three-way mixed ANOVA: sex, F(1,42)= 1.57, p = 0.22; genotype, F(1,42)= 0.04, p = 0.84; quadrant, F(3,126) = 11.69, p < 0.000001; sex × genotype interaction, F(1,42)= 0.04, p = 0.84; sex × quadrant interaction, F(3,126) = 0.74 p = 0.53; genotype × quadrant interaction, F(3,126) = 3.74, p < 0.02; sex × genotype × quadrant interaction, F(3,126) = 0.09, p = 0.96 |
| 11 pairs | Visible platform task (supplemental Fig. 2, available at www.jneurosci.org as supplemental material) | Latency to platform | Genotype, sex, and trial | Three-way mixed ANOVA: sex, F(1,17) = 0.81, p = 0.38; genotype, F(1,17) = 0.01, p = 0.91; trial, F(11,187) = 8.01, p < 0.000001; sex × genotype interaction, F(1,17) = 0.005, p = 0.95; sex × trial interaction, F(11,187) = 0.52, p = 0.89; genotype × trial interaction, F(11,187) = 1.49, p = 0.14; sex × genotype × trial interaction, F(11,187) = 0.93, p = 0.51 |
| Grooming plus DCS (Fig. 7) | ||||
| Vehicle, 20 pairs; DCS, 19 pairs | Grooming 30 min after drug treatment | Time grooming | Genotype, sex, and treatment | Three-way ANOVA: sex, F(1,40)= 0.57, p = 0.45; genotype, F(1,40)= 9.25, p < 0.004; treatment, F(1,40)= 6.01, p < 0.02; sex × genotype interaction, F(1,40)= 0.51, p = 0.48; sex × treatment, F(1,40)= 0.37, p = 0.54; genotype × treatment interaction, F(1,40)= 2.98, p = 0.09; sex × genotype × treatment interaction, F(1,40)=0.85, p = 0.36 |
| Genotype and treatment | Post hoc Tukey's honestly significant difference test: KO + vehicle versus KO + DCS, p < 0.02; KO + vehicle versus WT + DCS, p < 0.002; KO + DCS versus WT + vehicle, p = 0.97; WT + vehicle versus WT + DCS, p = 0.95 | |||
| Anxiety-like behavior (supplemental Figs. 1, 2, available at www.jneurosci.org as supplemental material) | ||||
| 23 pairs | Elevated plus maze | Frequency to enter open arms/frequency to enter all arms | Genotype and sex | Two-way ANOVA: genotype, F(1,42)= 0.00034, p = 0.99; sex, F(1,42)= 0.25, p = 0.62; genotype × sex interaction, F(1,42)= 2.80, p < 0.021; no significant comparisons with Tukey's post hoc test for the interaction |
| Time in open arms/time in all arms | Genotype and sex | Two-way ANOVA: genotype, F(1,42)= 0.92, p = 0.34; sex, F(1,42)= 1.47, p = 0.23; genotype × sex interaction, F(1,42)= 4.52, p < 0.039; no significant comparisons with Tukey's post hoc test for the interaction | ||
| 23 pairs | Dark/light box | Latency to enter light side | WT versus NL1 KO | Two-way ANOVA: genotype, F(1,42)= 1.00, p = 0.32; sex, F(1,42)= 0.066, p = 0.79; genotype × sex interaction, F(1,42)= 4.736, p < 0.035; no significant comparisons with Tukey's post hoc test for the interaction |
| Time in light side | WT versus NL1 KO | Two-way ANOVA: genotype, F(1,42)= 3.32, p = 0.077; sex, F(1,42)= 0.13, p = 0.72; genotype × sex interaction, F(1,42)= 0.29, p = 0.59 | ||
| Total activity | WT versus NL1 KO | Two-way ANOVA: genotype, F(1,42)= 3.37, p = 0.073; sex, F(1,42)= 0.011, p = 0.92; genotype × sex interaction, F(1,42)= 0.12, p = 0.73 | ||
| 23 pairs | Open field | Time in center/time in periphery | WT versus NL1 KO | Two-way ANOVA: genotype, F(1,42)= 0.53, p = 0.47; sex, F(1,42)= 1.93, p = 0.17; genotype × sex interaction, F(1,42)= 1.13, p = 0.29 |
| Frequency in center | WT versus NL1 KO | Two-way ANOVA: genotype, F(1,42)= 0.52, p = 0.47; sex, F(1,42)= 4.09, p = 0.05; genotype × sex interaction, F(1,42)= 0.74, p = 0.40 | ||
| Distance traveled | WT versus NL1 KO | Two-way ANOVA: genotype, F(1,42)= 1.59, p = 0.21; sex, F(1,42)= 0.17, p = 0.68; genotype × sex interaction, F(1,42)= 0.15, p = 0.70 | ||
| Velocity | WT versus NL1 KO | Two-way ANOVA: genotype, F(1,42)= 1.59, p = 0.21; sex, F(1,42)= 0.17, p = 0.68; genotype × sex interaction, F(1,42)= 0.15, p = 0.70 | ||
| Motor coordination (supplemental Fig. 2, available at www.jneurosci.org as supplemental material) | ||||
| 23 pairs | Rotarod | Time to fall off | Genotype, sex, and trial | Three-way mixed ANOVA: genotype, F(1,42)= 0.17, p = 0.69; sex, F(1,42)= 4.86, p < 0.033; trial, F(26,1092)= 6.24, p < 0.00001; no genotype × sex interaction, F(1,42)= 0.35, p = 0.56; no genotype × trial interaction, F(26,1092)= 0.78, p = 0.78; no sex × trial interaction, F(26,1092)= 0.92, p = 0.59; no genotype × sex × trial interaction, F(26,1092)= 0.56, p = 0.96 |
| Locomotor activity (supplemental Fig. 2, available at www.jneurosci.org as supplemental material) | ||||
| 23 pairs | Locomotor activity in a novel home cage | Locomotor activity | Genotype, sex, and trial | Three-way mixed ANOVA: genotype, F(1,30)= 0.043, p = 0.84; sex, F(1,30)= 6.56, p < 0.016; trial, F(23,690)= 41.22, p < 0.00001; no genotype × sex interaction, F(1,30)= 0.016, p = 0.90; no genotype × trial interaction, F(23,690)= 0.58, p = 0.94; no sex × trial interaction, F(23,690)= 0.39, p = 0.99; no genotype × sex × trial interaction, F(23,690)= 0.89, p = 0.62 |
| Startle response (supplemental Fig. 2, available at www.jneurosci.org as supplemental material) | ||||
| 23 pairs | Startle response | Baseline startle amplitude | Genotype and sex | Two-way ANOVA: genotype, F(1,40)= 0.85, p = 0.36; sex, F(1,40)= 0.90, p = 0.35; genotype × sex interaction, F(1,40)=0.097, p = 0.76 |
| Prepulse inhibition | Genotype, sex, and decibel | Three-way mixed ANOVA: genotype, F(1,40)= 0.064, p = 0.80; sex, F(1,40)= 3.50, p = 0.069; decibel, F(2,80)= 66.99, p < 0.000001; genotype × sex interaction, F(1,40)= 0.85, p = 0.36; genotype × decibel interaction, F(2,80)= 0.35, p = 0.71; sex × decibel interaction, F(2,80)= 2.01, p = 0.14; genotype × sex × decibel interaction, F(2,80)= 0.58, p = 0.56 |
Details and results of statistical analyses conducted for behavioral tests. Mixed ANOVA, ANOVA with a repeated measure (i.e., within-subjects factor). Repeated measures used in the analyses were target (for the 3-box social interaction test), trial (for social interaction with a juvenile, visible platform task, rotarod, and locomotor activity), day (for Morris water maze), quadrant (for Morris water maze probe trial), and decibel (for prepulse inhibition). F(x,y), F ratio statistic is used to determine whether the variances in two independent samples are equal; x,y, degrees of freedom.
On the accelerating rotarod, NL1 KO mice showed normal motor coordination and motor learning as measured over 27 trials (supplemental Fig. 2F, available at www.jneurosci.org as supplemental material). Compared with their WT littermates, NL1 KO mice also exhibited normal prepulse inhibition (supplemental Fig. 2G, available at www.jneurosci.org as supplemental material) as well as a normal baseline startle amplitude in response to an acoustic tone (supplemental Fig. 2H, available at www.jneurosci.org as supplemental material).
NL1 KO mice also exhibited normal fear learning and memory. During a test for contextual fear memory, both genotypes spent a similar percentage of time freezing [NL1 KO (n = 23), 54.91 ± 3.52%; WT (n = 23), 63.60 ± 3.81%, mean ± SEM; two-way ANOVA, main effect of genotype (between-subjects factor), F(1,42) = 2.71, p = 0.11; main effect of sex (between-subjects factor), F(1,42) = 0.42, p = 0.52; genotype × sex interaction, F(1,42) = 0.01, p = 0.91]; NL1 KO and WT mice also exhibited comparable levels of cue-dependent fear memory [NL1 KO (n = 23), 32.50 ± 5.10%; WT (n = 23), 39.64 ± 4.17%, mean ± SEM; two-way ANOVA, main effect of genotype (between-subjects factor), F(1,42) = 0.90, p = 0.35; main effect of sex (between-subjects factor), F(1,42) = 0.09, p = 0.76; genotype × sex interaction, F(1,42) = 0.69, p = 0.41].
Sensitivity to painful sensory stimuli was measured in two different tests. In a test of footshock sensitivity, a series of footshocks were delivered through a metal grid floor at increasing currents. Both WT and NL1 KO mice required similar current thresholds to elicit flinching and jumping behaviors (Fig. 1A). Compared with their WT littermates, NL1 KO mice required a higher current threshold to elicit audible vocalizations (Fig. 1) [two-way ANOVA (n = 11 pairs), main effect of genotype (between-subjects factor), F(1,18) = 6.47, p < 0.20; main effect of sex (between-subjects factor), F(1,18) = 9.29, p < 0.0069; genotype × sex interaction, F(1,18) = 1.78, p = 0.20], suggesting that, if anything, NL1 KO mice are slightly less sensitive to footshock. In a second test, mice were placed on a hotplate at 52°C, and NL1 KO mice exhibited a shorter latency to lick or shake their hindpaw compared with WT mice, suggesting that they are slightly more sensitive to heat [NL1 KO (n = 21), 11.79 ± 0.81 s; WT (n = 21), 14.33 ± 0.80 s, mean ± SEM; two-way ANOVA, main effect of genotype (between-subjects factor), F(1,38) = 4.90, p < 0.04; main effect of sex (between-subjects factor), F(1,38) = 2.26, p = 0.14; genotype × sex interaction, F(1,38) = 1.09, p = 0.30]. Although NL1 deletion appears to have mixed effects on nociception depending on the specific stimulus modality, the balance of the data suggest that, in general, NL1 deletion does not cause nonspecific, global behavioral dysfunction. For the complete statistical analyses of all behavioral tests, see Table 2.
Figure 1.

NL1 KO mice exhibit mixed responses to painful stimuli. When footshocks were delivered through a metal grid floor at increasing currents, the amount of current required to elicit flinching [two-way ANOVA, genotype (between-subjects factor), p = 0.78; sex (between-subjects factor), p = 0.54; genotype × sex interaction, p = 0.29] and jumping [two-way ANOVA, genotype (between-subjects factor), p = 0.34; sex (between-subjects factor), p = 0.62; genotype × sex interaction, p = 0.45] behaviors was similar for both WT and NL1 KO mice. Compared with WT mice, NL1 KO mice required a higher current threshold to elicit vocalizations [two-way ANOVA, genotype (between-subjects factor), *p < 0.0.020; sex (between-subjects factor), p < 0.0069; genotype × sex interaction, p = 0.20]. n = 11 littermate pairs. Data represent means ± SEM.
NL1 KO mice exhibit minimal deficits in social behavior
Because NL1 and neurexin 1 mutations in humans have been linked to autism spectrum disorders (Jamain et al., 2003; Chih et al., 2004; Comoletti et al., 2004; Laumonnier et al., 2004; Feng et al., 2006; Szatmari et al., 2007) and because there is a significant decrease in neurexin levels in NL1 KO mice, we tested NL1 KO mice in several tests of social behavior. NL1 KO mice exhibited a social interaction abnormality in only one of several tasks, showing decreased interaction with a caged, adult target mouse (Fig. 2A) [planned comparison (contrast analysis) of the effect of genotype within the social target only, F(1,42) = 4.64, p < 0.04; initial three-way mixed ANOVA (n = 23 pairs), main effect of genotype (between-subjects factor), F(1,42) = 4.76, p < 0.04; main effect of sex (between-subjects factor), F(1,42) = 0.001, p = 0.97; main effect of target (within-subjects factor), F(1,42) = 1.00, p = 0.76; genotype × sex interaction, F(1,42) = 1.17, p = 0.29; genotype × target interaction, F(1,42) = 0.80, p = 0.38; sex × target interaction, F(1,42) = 0.79, p = 0.38; genotype × sex × target interaction, F(1,42) = 0.30, p = 0.59] in a task that has been validated as a measure of social approach/avoidance in several previous publications (Berton et al., 2006; Kwon et al., 2006; Tsankova et al., 2006; Krishnan et al., 2007; Tabuchi et al., 2007; Lutter et al., 2008). It is important to note that interaction with an inanimate, empty cage in the same apparatus, under the same conditions, was normal (Fig. 2A) [planned comparison (contrast analysis) of the effect of genotype within the inanimate target only, F(1,42) = 2.43, p = 0.13], indicating specificity for social interaction. In addition, the total distance moved during the test of interaction with a social target was similar between genotypes [trial with a social interaction target: NL1 KO, 2035.52 ± 129.71 cm; WT, 2490.96 ± 96.50 cm; trial with an inanimate interaction target: NL1 KO, 1581.90 ± 100.18 cm; WT, 1573.97 ± 78.26 cm, mean ± SEM; three-way mixed ANOVA (n = 23 pairs), main effect of genotype (between-subjects factor), F(1,42) = 2.74, p = 0.11; main effect of sex (between-subjects factor), F(1,42) = 0.10, p = 0.76; main effect of target (within-subjects factor), F(1,42) = 112.26, p < 0.000001; genotype × sex interaction, F(1,42) = 0.12, p = 0.73; genotype × target interaction, F(1,42) = 10.36, p < 0.003; sex × target interaction, F(1,42) = 2.70, p = 0.11; genotype × sex × target interaction, F(1,42) = 4.97, p < 0.04].
Figure 2.
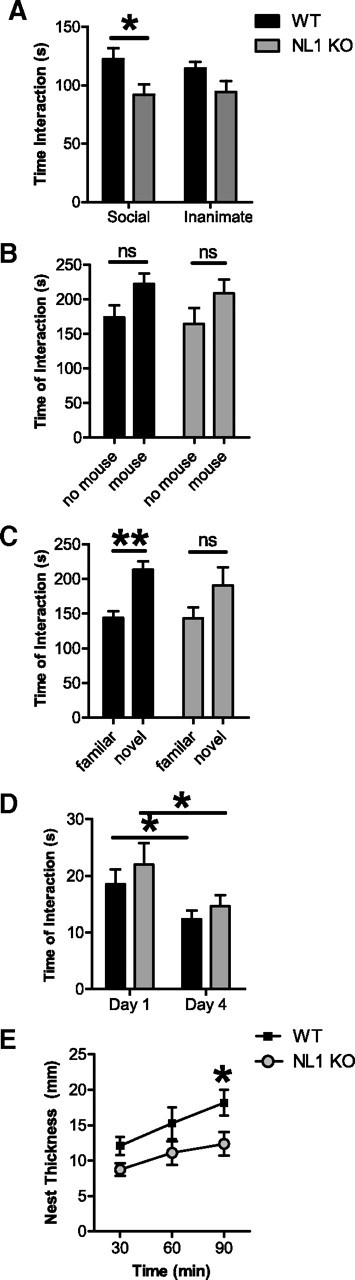
NL1 KO mice exhibit minimal social behavior abnormalities. A, NL1 WT and KO mice were allowed to interact sequentially with a novel inanimate target (i.e., an empty cage) and a social target (i.e., a caged adult conspecific mouse) in an open field. An initial three-way mixed ANOVA found a significant main effect of genotype [three-way mixed ANOVA, genotype (between-subjects factor), p < 0.04; sex (between-subjects factor), p = 0.97; target (within-subjects factor), p = 0.76; genotype × sex interaction, p = 0.29; genotype × target interaction, p = 0.38; sex × target interaction, p = 0.38; genotype × sex × target interaction, p = 0.59]. Additional planned comparisons found that NL1 WT mice spent less time interacting with the social target than controls (left bars, *p < 0.04, planned comparisons). Both NL1 WT and KO mice spent similar amounts of time interacting with the novel, inanimate target (right bars, p = 0.13, planned comparisons). n = 23 littermate pairs. Legend in A applies to A–E. B, Time spent in interactions using the social versus inanimate preference test. Mice were simultaneously exposed to a novel inanimate target (i.e., an empty cage) and a novel social target (i.e., a novel, caged adult conspecific mouse). The time spent interacting with inanimate and social targets was not different between NL1 KO mice and controls, and neither WT or NL1 KO mice preferred the social target over the inanimate target [three-way mixed ANOVA, genotype (between-subjects factor), p = 0.22; sex (between-subjects factor), p = 0.16; target (within-subjects factor), p = 0.08; genotype × sex interaction, p = 0.27; genotype × target interaction, p = 0.97; genotype × sex × target interaction, p = 0.81]. ns, Not significant. n = 23 littermate pairs. C, Time spent in interactions using the social novelty preference test. Mice were simultaneously exposed to a novel social target (i.e., a novel, caged adult conspecific mouse) and a familiar social target (i.e., a familiar, caged adult conspecific mouse). A three-way mixed ANOVA found a main effect of target [three-way mixed ANOVA, genotype (between-subjects factor), p = 0.39; sex (between-subjects factor), p = 0.50; target (within-subjects factor), p < 0.003; genotype × sex interaction, p = 0.31; genotype × target interaction, p = 0.40; sex × target interaction, p = 0.87; genotype × sex × target interaction, p = 0.46]. However, planned comparisons within each genotype found that, unlike WT mice, NL1 KO mice did not exhibit a preference for the novel social target (**p < 0.006; ns, not significant; p = 0.10, planned comparisons). n = 23 littermate pairs. D, Time spent interacting with a freely moving juvenile mouse during a test of social learning [three-way mixed ANOVA, genotype (between-subjects factor), p = 0.39; sex (between-subjects factor), p < 0.041; trial (within-subjects factor), p < 0.0007; genotype × sex interaction, p = 0.56; genotype × trial interaction, p = 0.78; sex × trial interaction, p = 0.07; genotype × sex × trial interaction, p = 0.23]. Mice were initially allowed to interact with a juvenile target mouse for 2 min (Day 1), and NL1 KO mice showed normal direct social interaction with the juvenile. Seventy-two hours later (Day 4), mice were again allowed to interact with the same juvenile target mouse, and both WT and NL1 KO mice exhibited significant social learning (recognition memory), as indicated by a decrease in the amount of time spent interacting with the juvenile (*p < 0.021 for WT; p < 0.008 for KO, planned comparisons within each genotype). n = 23 littermate pairs. E, The thickness (i.e., the height) of nests built from cotton nesting material was measured over a 90 min observation period [three-way mixed ANOVA, genotype (between-subjects factor), p < 0.044; sex (between-subjects factor), p = 0.35; time (within-subjects factor), p < 0.000013; genotype × sex interaction, p = 0.69; genotype × time interaction, p = 0.15; sex × time interaction, p = 0.18; genotype × sex × time interaction, p < 0.032]. WT mice built thicker nests than NL1 KO mice (*p < 0.021, planned comparison between genotypes at 90 min). n = 11 littermate pairs. Data represent means ± SEM.
This isolated, task-specific abnormality in social behavior is not likely attributable to altered olfactory ability because time spent interacting with a “social smell” was normal in NL1 KO mice [NL1 KO, 52.17 ± 4.03 s; WT, 53.07 ± 3.81 s, mean ± SEM; two-way ANOVA (n = 22 pairs), main effect of genotype (between-subjects factor), F(1,40) = 0.30, p = 0.59; main effect of sex (between-subjects factor), F(1,40) = 4.11, p < 0.049; genotype × sex interaction, F(1,40) = 2.01, p = 0.16]. Importantly, gross olfactory abilities were also normal in NL1 KO mice as measured by latency to find a buried treat in a neutral home cage [NL1 KO, 340.20 ± 53.30 s; WT, 387.90 ± 37.62 s, mean ± SEM; two-way ANOVA (n = 10 pairs), main effect of genotype (between-subjects factor), F(1,16) = 0.65, p = 0.43; main effect of sex (between-subjects factor), F(1,16) = 2.58, p = 0.13; genotype × sex interaction, F(1,16) = 0.13, p = 0.72].
In three other social tasks, however, no differences were observed. In a test for social versus inanimate interaction, there was no difference between WT and NL1 KO in time spent interacting with either the social target or the inanimate cage, nor was there a significant preference for the social versus inanimate target for either genotype (Fig. 2B). In a test for familiar versus novel social interaction, there was also no significant difference between NL1 KO and WT littermates in time spent interacting with either the novel or the familiar social target (Fig. 2C). Unlike WT mice, NL1 KO mice did not show a statistically significant preference for the novel social target compared with the familiar, although a similar trend was apparent (Fig. 2C) [planned comparison (contrast analysis) of novel vs familiar target: WT, F(1,42) = 8.42, p < 0.006; NL1 KO, F(1,42) = 2.89, p = 0.10]. Likewise, no differences in social interaction or social learning were observed in a task of reciprocal social interaction with a juvenile conspecific (Fig. 2D). Interestingly, although not a strictly social behavior, NL1 KO mice displayed impaired nest building skills compared with WT (Fig. 2E) [initial three-way mixed ANOVA (n = 11 pairs), main effect of genotype (between-subjects factor), F(1,18) = 4.74, p < 0.044; main effect of sex (between-subjects factor), F(1,18) = 0.93, p = 0.35; main effect of time (within-subjects factor), F(2,36) = 15.70, p < 0.000013; genotype × sex interaction, F(1,18) = 0.17, p = 0.69; genotype × time interaction, F(2.36) = 2.03, p = 0.15; sex × time interaction, F(2,36) = 1.81, p = 0.18; genotype × sex × time interaction, F(2,36) = 3.80, p < 0.032; planned comparisons (contrast analysis) comparing genotypes, 30 min, F(1,18) = 4.26, p = 0.054; 60 min, F(1,18) = 2.47, p = 0.13; 90 min, F(1,18) = 6.49, p < 0.021].
NL1 KO mice exhibit deficits in spatial memory
Because mental retardation is associated with many cases of ASDs (American Psychiatric Association, 2000) and some ASD patients with neurexin mutations exhibit low IQ (Kim et al., 2008a), we tested learning and memory in NL1 KO mice using the Morris water maze task. NL1 KO mice exhibited significant abnormalities in spatial learning and memory. Despite a normal learning curve as measured using latency to reach the platform during training (Fig. 3B), NL1 KO mice exhibited a slight learning deficit using distance traveled before reaching the hidden platform, an analysis that eliminates swim speed as a concern (Fig. 3A) [three-way mixed ANOVA (n = 23 pairs), main effect of genotype (between-subjects factor), F(1,42) = 7.52, p < 0.0089; main effect of sex (between-subjects factor), F(1,42) = 0.26, p = 0.61; main effect of day (within-subjects factor), F(10,420) = 20.95, p < 0.000001; genotype × sex interaction, F(1,42) = 0.011, p = 0.92; genotype × day interaction, F(10,420) = 0.78, p = 0.65; sex × day interaction, F(10,420) = 93, p = 0.50; genotype × sex × day interaction, F(10,420) = 0.50, p = 0.89]. In fact, NL1 KO mice showed a slight increase in average swim speed compared with WT (Fig. 3C) [three-way mixed ANOVA (n = 23 pairs), main effect of genotype (between-subjects factor), F(1,42) = 4.59, p < 0.038; main effect of sex (between-subjects factor), F(1,42) = 2.21, p = 0.14; main effect of day (within-subjects factor), F(10,420) = 5.76, p < 0.000001; genotype × sex interaction, F(1,42) = 0.0058, p = 0.94; genotype × day interaction, F(10,420) = 0.52, p = 0.88; sex × day interaction, F(10,420) = 0.50, p = 0.89; genotype × sex × day interaction, F(10,420) = 0.60, p = 0.82], likely explaining why their latency to reach the platform learning curve appeared normal. During these training trials, NL1 KO mice did not spend more time near the wall of the maze (thigmotaxis) compared with WT (Fig. 3D). On a spatial memory test 24 h after the end of Morris water maze training (probe trial), WT mice spent significantly more time in the target quadrant than all other quadrants (Fig. 3E) [planned comparisons (contrast analysis), target vs opposite, F(1,42) = 27.21, p < 0.000006; target vs adjacent left, F(1,42) = 21.15, p < 0.00004; target vs adjacent right, F(1,42) = 5.99, p < 0.02], whereas NL1 KO mice showed no significant preference for the target quadrant compared with any other quadrant [planned comparisons (contrast analysis), target vs opposite, F(1,42) = 2.10, p = 0.15; target vs adjacent left, F(1,42) = 2.06, p = 0.16; target vs adjacent right, F(1,42) = 1.05, p = 0.31]. The NL1 KO mice performed at near chance levels (Fig. 3E), indicating a spatial memory deficit. Furthermore, NL1 KO mice spent significantly less time in the target quadrant and more time in the opposite quadrant than WT (Fig. 3E) [planned comparisons (contrast analysis), target quadrant, F(1,42) = 4.67, p < 0.037; opposite quadrant, F(1,42) = 8.67, p < 0.006; adjacent left quadrant, F(1,42) = 1.89, p = 0.17; adjacent right quadrant, F(1,42) = 0.69, p = 0.41]. It is important to note that NL1 KO mice learned the visible platform task as well as controls (supplemental Fig. 2I, available at www.jneurosci.org as supplemental material), indicating that basic neurological function (swimming, vision, etc.) was intact.
Figure 3.
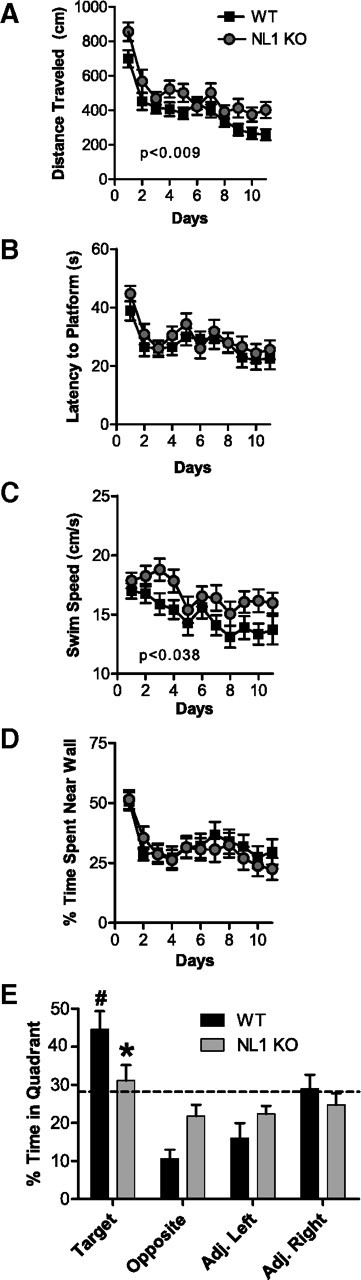
NL1 KO mice exhibit impaired spatial memory in the Morris water maze. A–D, Training trials for the Morris water maze task. For each day of training, data were averaged across the four daily trials, and, in all three-way mixed ANOVAs, “day” was treated as the repeated measure. NL1 KO mice displayed an abnormal learning curve as measured by distance to reach the submerged platform compared with WT (A) [three-way mixed ANOVA, genotype (between-subjects factor), p < 0.0089; sex (between-subjects factor), p = 0.61; day (within-subjects factor), p < 0.000001; genotype × sex interaction, p = 0.92; genotype × day interaction, p = 0.65; sex × day interaction, p = 0.50; genotype × sex × day interaction, p = 0.89]. The latency to reach the platform (B) was normal in the NL1 KO mice [three-way mixed ANOVA, genotype (between-subjects factor), p = 0.47; sex (between-subjects factor), p = 0.65; day (within-subjects factor), p < 0.000001; genotype × sex interaction, p = 0.74; genotype × day interaction, p = 0.93; sex × day interaction, p = 0.11; genotype × sex × day interaction, p = 0.50]. NL1 KO mice exhibited faster average swim speeds (C) than controls [three-way mixed ANOVA, genotype (between-subjects factor), F(1,42) = 4.59, p < 0.038; sex (between-subjects factor), p = 0.14; day (within-subjects factor), p < 0.000001; genotype × sex interaction, p = 0.94; genotype × day interaction, p = 0.88; sex × day interaction, p = 0.89; genotype × sex × day interaction, p = 0.82]. Percentage time spent near the wall of the maze (D, percentage thigmotaxis) was not different across groups [three-way mixed ANOVA, genotype (between-subjects factor), p = 0.95; sex (between-subjects factor), p = 0.82; day (within-subjects factor), p < 0.000001; genotype × sex interaction, p = 0.99; genotype × day interaction, p = 0.60; sex × day interaction, p = 0.78; genotype × sex × day interaction, p = 0.96]. There were no differences between groups in the visible platform task (supplemental Fig. 2, available at www.jneurosci.org as supplemental material). n = 23 littermate pairs. Legend in A also applies to B–D. E, Percentage of time that mice spent swimming in each quadrant of the pool during the probe trial (i.e., spatial memory test) on day 12 of the Morris water maze [three-way mixed ANOVA, sex (between-subjects factor), p = 0.22; genotype (between-subjects factor), p = 0.84; quadrant (within-subjects factor), p < 0.000001; sex × genotype interaction, p = 0.84; sex × quadrant interaction, p = 0.53; genotype × quadrant interaction, p < 0.02; sex × genotype × quadrant interaction, p = 0.96]. NL1 KO mice spent less time in the target quadrant than controls (*p < 0.037 for NL1 KO target vs WT target, planned comparisons) and, unlike control mice (#p < 0.05 compared with all other quadrants, planned comparisons) showed no preference for the target quadrant, indicating a deficit in spatial memory. n = 23 littermate pairs. Data represent means ± SEM.
NL1 KO mice exhibit deficits in hippocampal LTP
Because NL1 KO mice exhibit a decrease in hippocampus-dependent spatial memory and a decrease in the NMDA/AMPA ratio in area CA1 of the hippocampus (Chubykin et al., 2007), we predicted that NL1 KO mice would exhibit a decrease in LTP in area CA1 of the hippocampus. Indeed, theta burst stimulation (five bursts of four pulses at 100 Hz with an interburst interval of 0.2 s) resulted in a significantly reduced magnitude of LTP in area CA1 of the hippocampus in slices from NL1 KO mice compared with WT littermate controls (Fig. 4A,B) [LTP 50–60 min after TBS induction (fEPSP expressed as the fraction of control): NL1 KO (n = 6), 1.49 ± 0.09; WT (n = 6), 1.88 ± 0.13, mean ± SEM; t test, p < 0.031]. This decrease in LTP magnitude was not accompanied by any alteration in basal synaptic transmission because input–output curves (Fig. 4C,D) and paired-pulse facilitation (Fig. 4E,F) were normal. Based on the previously observed deficits in NMDAR transmission in NL1 KO mice (Chubykin et al., 2007; Kim et al., 2008b), it is reasonable to assume that the LTP phenotype is most likely caused by a deficit in LTP induction.
Figure 4.
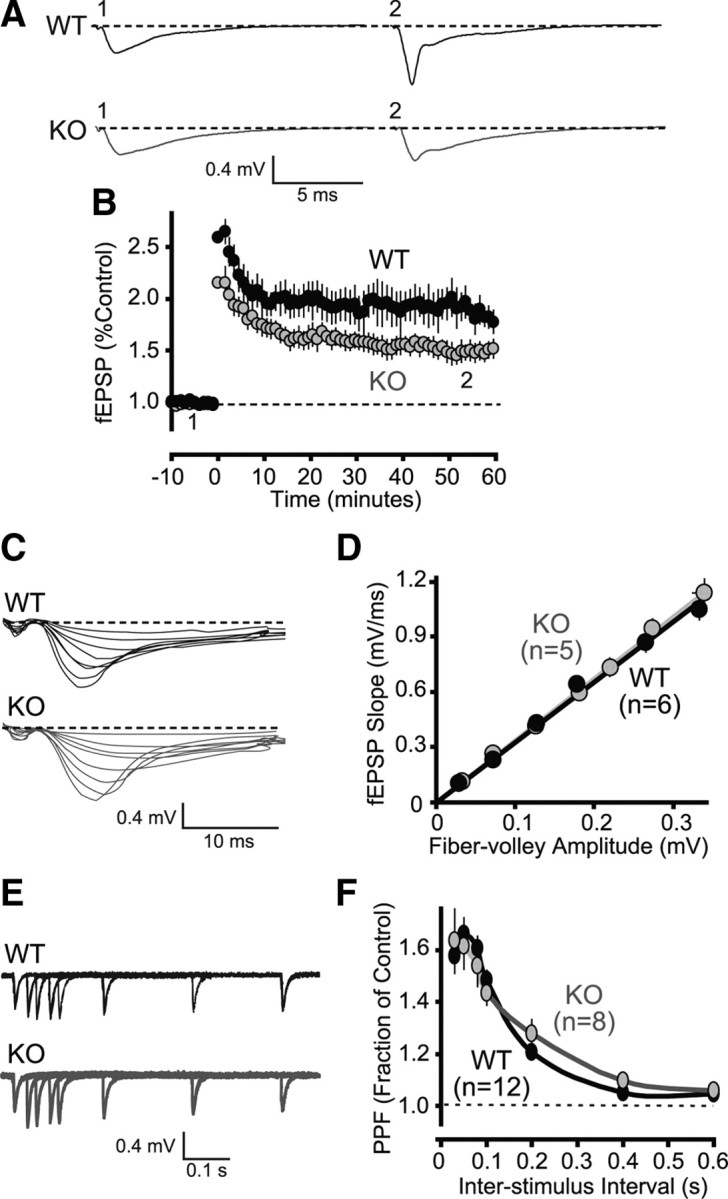
Decreased theta burst-induced LTP in area CA1 of NL1 KO mice. A, Representative traces showing fEPSPs from a WT and a KO animal before (1) and after (2) LTP induction by TBS. B, Time course of fEPSPs before and after LTP induction by TBS in WT (n = 6) and KO (n = 6) animals. The time points of the representative traces in A are noted on the graph (see 1 and 2). C, AMPA-mediated synaptic strength is unchanged in NL1 KO mice. Representative traces from a WT and a KO animal show input–output measurements with increasing stimulus intensities. Responses were measured in area CA1 of mouse hippocampus. D, Average fEPSP slope plotted against fiber-volley amplitude. Input–output measurements were performed in wild-type (n = 6) and KO (n = 5) animals. E, Overlaid representative traces from a WT and a KO animal showing paired-pulse facilitation of excitatory synaptic responses at interstimulus intervals ranging from 30 to 600 ms. F, Average paired-pulse facilitation of excitatory synaptic responses did not show any difference between WT (n = 12) and KO (n = 8) animals. Data plotted as means ± SEM.
NL1 KO mice exhibit a decrease in the NMDA/AMPA ratio in the hippocampus (Chubykin et al., 2007), which could be attributable to either a change in postsynaptic receptor function or a change in the number of NMDA- or AMPA-containing (i.e., silent or nonsilent) synapses. Therefore, we examined the effect of NL1 loss on synaptic density in vivo. We found no significant alterations in total synapse density (supplemental Fig. 3, available at www.jneurosci.org as supplemental material), excitatory synapse density (Figs. 5, 6), or inhibitory synapse density (Figs. 5, 6) in the hippocampus. Furthermore, no changes were observed in the size of immunopositive puncta with any targeted antigen (Figs. 5, 6) (supplemental Fig. 3, available at www.jneurosci.org as supplemental material). Also, as mentioned above, whole-brain immunoblots detected no significant changes in the expression levels of multiple NMDAR subunits in NL1 KO mice (Table 1). The findings that NL1 KO mice exhibit no change in the number of excitatory immunopositive puncta and no change in the expression levels of NMDAR subunits suggest that the decreased hippocampal NMDA/AMPA ratio observed in NL1 KO mice (Chubykin et al., 2007) may be attributable to altered excitatory postsynaptic receptor function rather than changes in synapse or NMDAR number. However, the current results cannot rule out more subtle effects of NL1 deletion on excitatory or inhibitory synapse number, synaptic NMDAR numbers, or specific subtypes of inhibitory or excitatory synapses.
Figure 5.
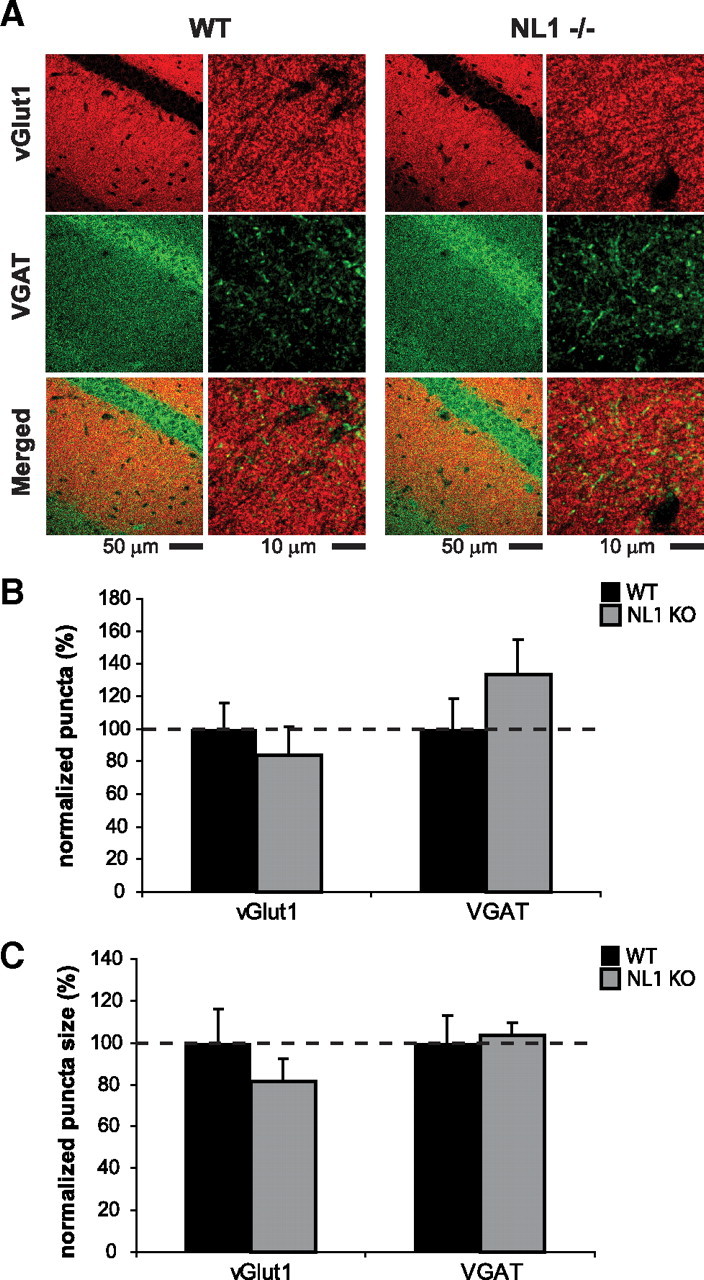
Number and size of glutamatergic and GABAergic synapses are normal in NL1-deficient CA1 region of hippocampus. A, Representative confocal images of wild-type and NL1 KO CA1 region of hippocampus double immunostained for VGLUT1 and VGAT. Number (B) and size (C) of both VGLUT1- and VGAT-positive puncta are normal in NL1 KO neurons. y-axis depicts normalized number (B) and size (C) with wild-type control. n = 3 mice per genotype.
Figure 6.
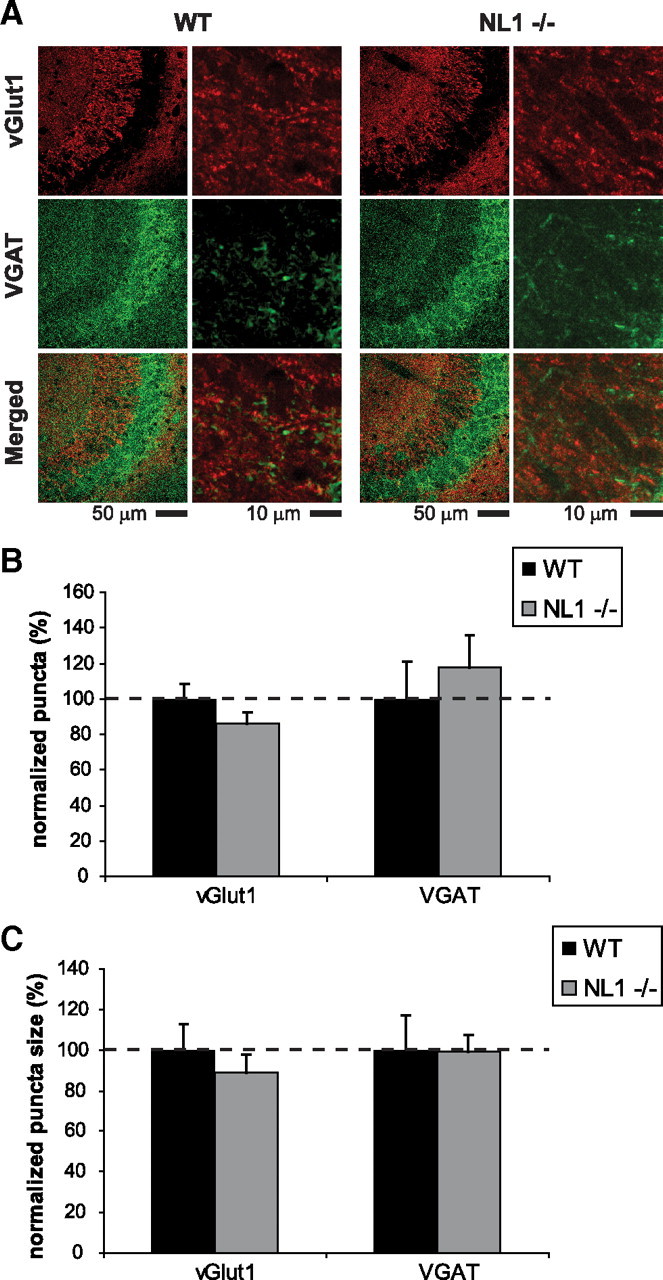
Number and size of glutamatergic and GABAergic synapses are normal in NL1-deficient CA3 region of hippocampus. A, Representative confocal images of wild-type and NL1 KO CA3 region of hippocampus double immunostained for VGLUT1 and VGAT. B, C, Number (B) and size (C) of both VGLUT1- and VGAT-positive puncta are normal in NL1 KO neurons. y-axis depicts normalized number (B) and size (C) with wild-type control. n = 3 mice per genotype.
NL1 KO mice exhibit increased repetitive grooming behavior
Because NL and neurexin 1 mutations in humans have been linked to autism spectrum disorders (Jamain et al., 2003; Chih et al., 2004; Comoletti et al., 2004; Laumonnier et al., 2004; Feng et al., 2006; Szatmari et al., 2007), we characterized grooming behavior in NL1 KO mice, a behavior that might reflect the repetitive, stereotyped behavior core symptom domain of autism (Moy et al., 2006; Crawley, 2007). NL1 KO mice spent more than double the amount of time spontaneously grooming compared with WT mice [NL1 KO (n = 22), 56.15 ± 11.32 s; WT (n = 22), 25.15 ± 7.57 s, mean ± SEM; two-way ANOVA, main effect of genotype (between-subjects factor), F(1,40) = 5.87, p < 0.020; main effect of sex (between-subjects factor), F(1,40) = 2.0, p = 0.17; genotype × sex interaction, F(1,40) = 0.63, p = 0.43]. In a marble burying task, which has been described as a task relevant to anxiety and to obsessive-compulsive/repetitive behavior (Broekkamp et al., 1986; Njung'e and Handley, 1991; Borsini et al., 2002; Deacon, 2006; Thomas et al., 2009), there was no difference between NL1 KO and WT mice [mean ± SEM number of marbles buried, NL1 KO mice, 5.5 ± 1.37; WT mice, 8.86 ± 1.54; two-way ANOVA (n = 22 pairs), main effect of genotype (between-subjects factor), F(1,40) = 2.51, p = 0.12; main effect of sex (between-subjects factor), F(1,40) = 0.01, p = 0.93; genotype × sex interaction, F(1,40) = 0.03, p = 0.86].
NL1 KO mice exhibit a reduced NMDA/AMPA ratio in the dorsal striatum
Although a decrease in the NMDA/AMPA ratio has been observed in the hippocampus of NL1 KO mice (Chubykin et al., 2007), it is unlikely that this is responsible for the increased grooming behavior observed in NL1 KO mice (see above) because the hippocampus is not known to be involved with mammalian grooming behavior. Because the dorsal striatum has been repeatedly implicated in rodent grooming behavior (Cromwell and Berridge, 1996; Aldridge et al., 2004; Welch et al., 2007), we hypothesized that NL1 KO mice might exhibit similar alterations in synaptic transmission in the dorsal striatum.
Whole-cell patch-clamp recordings of striatal medium spiny neurons were performed to determine the NMDA/AMPA ratio in corticostriatal synapses. Baseline values for access resistance (14.2 ± 0.7 MΩ, wild type; 13.6 ± 0.8 MΩ, NL1 KO), cell membrane resistance (340 ± 54 MΩ, wild type; 312 ± 58 MΩ, NL1 KO), and cell capacitance (167.2 ± 11 pF, wild type; 169.3 ± 10 pF, NL1 KO) did not differ between the groups. The NMDA/AMPA ratio was assessed by two measurements: the peak of the evoked EPSCs (eEPSCs) at −80 mV, to detect the AMPAR currents; and the current amplitude, 50 ms after spike onset and at +40 mV, to detect the NMDAR currents. AMPAR eEPSC amplitude was 471 ± 61 pA for wild type and 470 ± 62 pA for NL1 KO (t test, p = 0.995). Consistent with our hypothesis, the NMDA/AMPA ratio in the striatum of NL1 KO mice was significantly reduced by ∼30% (Fig. 7A) [NMDA/AMPA ratio, NL1 KO (n = 22), 0.77 ± 0.07; WT (n = 23), 1.00 ± 0.08, mean ± SEM; t test, p < 0.01].
Figure 7.

NL1 KO mice exhibit a decreased NMDA/AMPA ratio in the dorsal striatum accompanied by increased repetitive behavior that can be rescued pharmacologically. A, NMDAR- and AMPAR-mediated responses are indicated in wild-type traces. Current amplitude was normalized to that of the AMPAR peak. Amplitude of NMDAR currents at +40 mV was measured in a 2 ms window set at 50 ms from spike onset (dashed line on traces). B, The time spent grooming was observed 30 min after systemic administration of the NMDA partial coagonist DCS (20 mg/kg). Administration of DCS rescued the increased grooming phenotype in NL1 KO mice [*p < 0.02 compared with all other groups, post hoc Tukey's honestly significant difference test; three-way ANOVA, sex (between-subjects factor), p = 0.45; genotype (between-subjects factor), p < 0.004; treatment (between-subjects factor), p < 0.02; sex × genotype interaction, p = 0.48; sex × treatment, p = 0.54; genotype × treatment interaction, p = 0.09; sex × genotype × treatment interaction, p = 0.36]. Vehicle (Veh), n = 20 littermate pairs; DCS, n = 19 littermate pairs. Data represent means ± SEM.
Systemic d-cycloserine rescues the increased repetitive behavior in NL1 KO mice
We next examined whether a drug that is known to enhance NMDA receptor function and NMDA receptor-dependent behaviors in vivo could acutely reverse the increased grooming behavior in NL1 KO mice. Given that NL1 KO mice exhibited a decrease in the NMDA/AMPA ratio in the dorsal striatum, we hypothesized that altering NMDA receptor function pharmacologically would rescue the abnormal grooming behavior in NL1 KO mice. To test this, we systemically administered either the NMDA receptor coagonist DCS or vehicle 30 min before measuring grooming behavior. Consistent with our previous findings (see above), NL1 KO mice treated with vehicle displayed increased grooming compared with WT mice treated with vehicle (Fig. 7B) (post hoc Tukey's test, KO plus vehicle vs WT plus vehicle, p < 0.005). However, 20 mg/kg DCS given 30 min before testing rescued the increased grooming in NL1 KO mice (Fig. 7B) (post hoc Tukey's test, KO plus DCS vs WT plus DCS, p = 0.78).
Discussion
NL1 and hippocampus-dependent learning and memory
Genetic deletion of the excitatory synapse cell-adhesion molecule NL1 in mice leads to decreased long-term synaptic plasticity in area CA1 of the hippocampus and disruption of hippocampus-dependent spatial memory. In addition to a decreased rate of learning, NL1 KO mice were unable to use a spatial strategy to locate a submerged platform in the Morris water maze. These deficits were not associated with altered thigmotaxis and are not explained by differences in swim speed, coordination, locomotor activity, or vision. The most parsimonious explanation for impaired hippocampus-dependent learning and memory in NL1 KOs is the decreased LTP we observed in NL1 KO mice, a finding perhaps best linked to the decreased NMDA/AMPA current ratio observed previously in area CA1 of the hippocampus of NL1 KO mice (Chubykin et al., 2007). Our findings are consistent with a recent report suggesting that NL1 is required for NMDA receptor-mediated synaptic currents and normal expression of LTP in the amygdala (Kim et al., 2008b). A finding of mildly reduced learning, but not memory, and decreased LTP induction has also been observed in NL1 transgenic mice that overexpress NL1, although the mechanisms underlying these findings remain unclear (Dahlhaus et al., 2010). These cognitive abnormalities are consistent with the idea that NL1 or neurexin 1 may play important roles at the synapse relevant to comorbid mental retardation in autism spectrum disorders.
NL1 and social behavior
Interestingly, NL1 KO mice exhibit little to no deficits in social behavior. In only one of several tasks did NL1 KOs interact less with a caged adult target compared with wild-type littermate controls. This is observed despite normal locomotor activity, habituation, and olfactory ability for food and social odors. Furthermore, NL1 KO mice do not engage in nesting behavior as readily as their WT littermate counterparts. On many other social tasks, however, NL1 KO mice show normal interaction and approach to social targets as well as normal social recognition. Given the finding of reduced social interaction in only one social approach task and in nesting behavior, the bulk of the data do not favor a strong abnormality in social behavior in the NL1 KO mice.
NL1 and enhanced repetitive, grooming behavior
NL1 KO mice exhibit a clear, significant increase in repetitive, grooming behavior, and this phenotype is robust and reproducible. Although the clinical significance of NL1 deletion is not entirely clear, NL1 binds to presynaptic neurexins, which have been implicated in human autism (Feng et al., 2006; Szatmari et al., 2007). We find that NL1 deletion does lead to a small but significant reduction in neurexin levels in the brain, and therefore, it is possible that the enhanced repetitive behavior in NL1 KO mice could be representative of one of the symptoms of autism, namely increased repetitive, stereotyped behaviors. Indeed, neurexin 1 deficient mice also exhibit a similar increase in repetitive grooming behavior (Etherton et al., 2009). Of course, grooming behavior is not consistent across species, and increased repetitive grooming behavior has been suggested to exhibit significant face validity not only for autism spectrum disorders but also for obsessive/compulsive disorder (OCD) and trichotillomania (Welch et al., 2007; Bienvenu et al., 2009; Züchner et al., 2009).
Consistent with a link between the enhanced repetitive behavior and our findings of reduced NMDA/AMPA ratio in both the hippocampus (Chubykin et al., 2007) and the dorsal striatum (Fig. 7), we successfully rescued the increased grooming in NL1 KO mice with systemic d-cycloserine at a dose reported previously in the literature to augment NMDA receptor-dependent forms of learning and memory in the brain (Flood et al., 1992; Zlomuzica et al., 2007). Vehicle-treated NL1 KO mice showed significantly enhanced grooming, whereas DCS-treated NL1 KO mice revealed completely normal levels of grooming. No alteration of grooming behavior was observed in WT littermates with DCS treatment. These data support the hypothesis that reduced NMDA receptor-mediated synaptic transmission in NL1 KO mice (Fig. 7) mediates the enhanced grooming in these mice, suggesting a potential treatment for at least one cause of this behavioral abnormality of potential relevance to autism, OCD, or trichotillomania. Interestingly, a small pilot study of d-cycloserine treatment in autism has been published previously and indicates that DCS may be of potential benefit in patients with autism, although the social domain was more prominently affected in this inconclusive, underpowered pilot study (Posey et al., 2004).
One alternative possibility for increased grooming in our mice is an NMDAR-related altered sensation or nociception. However, three lines of evidence argue against this. First, we have detected a decrease (not an increase) in NMDAR currents in both the hippocampus and the striatum of NL1 KO mice, suggesting a low likelihood for an NMDAR-driven increase in itching or other sensation that might increase grooming in NL1 KO mice (Ferreira and Lorenzetti, 1994; Tan-No et al., 2000). Second, the grooming bouts were primarily syntactic (i.e., followed a cephalocaudal direction). It is difficult to imagine why focal sensory abnormalities would lead to a syntactic grooming pattern rather than focal scratching behavior. Finally, sensory thresholds in these mutants, as determined with the footshock and hotplate tests, were not consistently altered in a single direction, further decreasing the likelihood of altered sensory function in the increased grooming behavior of NL1 KO mice.
Role of NL1 in synaptic transmission and NMDA receptor function
The precise mechanisms through which NL1 modulates NMDA receptor-mediated synaptic transmission remain to be determined. NL1 is selectively localized to excitatory synapses (Song et al., 1999) and interacts with NMDA receptors through its interaction with postsynaptic density protein-95 (PSD-95) via a PDZ (PSD-95/Discs large/zona ocludens-1) type I domain on its C terminus (Irie et al., 1997). Several PSD-95-associated proteins are thought to be clustered before assembly of postsynaptic spines (Prange et al., 2004; Gerrow et al., 2006). NL1 and NMDARs are enriched in those clusters (Irie et al., 1997), whereas AMPARs incorporate into synapses after spine formation (Nam and Chen, 2005). It is possible that the association between NL1 and other PSD-95-associated proteins, including the NMDAR, may preclude the efficient incorporation of some of these proteins into the synapses of NL1 KO mice. NMDA receptors themselves are highly dynamic, even within the plasma membrane (Newpher and Ehlers, 2008), and phosphorylation of the NMDAR is known to alter its trafficking (Chung et al., 2004; Lin et al., 2006). Additional study is needed to determine whether this type of NMDAR modulation is altered in NL1 KO mice.
Relevance to autism, mental retardation, and other neuropsychiatric disorders
Although they exhibit increased repetitive behaviors and cognitive deficits, it is premature to suggest that NL1 KO mice represent an accurate rodent model of human autism or mental retardation. Although chromosomal rearrangements in regions that harbor the NL1 gene (Konstantareas and Homatidis, 1999; Zoghbi, 2003; Yan et al., 2004) and copy number variations of the NL1 gene (Glessner et al., 2009) have been implicated in cases of autism in humans, the evidence for a direct link between NL1 and human disease remains sparse (Talebizadeh et al., 2004; Vincent et al., 2004; Gauthier et al., 2005; Ylisaukko-oja et al., 2005). Additional study of the NL1 KO mice, however, will provide insights into the neural basis of increased repetitive behaviors of potential relevance to autism spectrum disorders, OCD, and trichotillomania. With the recent implication of neurexin 1 and shank3 in human cases of autism spectrum disorders and the known binding of neurexins and shank3 to NL1, one might expect the NL1 KO phenotype to foreshadow a subset of the neurexin 1 or shank3 KO phenotype.
Footnotes
This work was supported by grants from Autism Speaks (C.M.P.), the Simons Foundation (T.C.S.), National Institute of Mental Health Grants MH065975-05 (C.M.P.) and R37 MH52804-08 (T.C.S.), and gifts from BRAINS for Autism/Debra Caudy and Clay Heighten (Founders), the Crystal Charity Ball, and The Hartwell Foundation (C.M.P.). C.M.P. is a Hartwell Scholar.
References
- Aldridge JW, Berridge KC, Rosen AR. Basal ganglia neural mechanisms of natural movement sequences. Can J Physiol Pharmacol. 2004;82:732–739. doi: 10.1139/y04-061. [DOI] [PubMed] [Google Scholar]
- American Psychiatric Association. Diagnostic and statistical manual of mental disorders DSM-IV-TR. Washington, DC: American Psychiatric Association; 2000. [Google Scholar]
- Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, Graham D, Tsankova NM, Bolanos CA, Rios M, Monteggia LM, Self DW, Nestler EJ. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science. 2006;311:864–868. doi: 10.1126/science.1120972. [DOI] [PubMed] [Google Scholar]
- Bienvenu OJ, Wang Y, Shugart YY, Welch JM, Grados MA, Fyer AJ, Rauch SL, McCracken JT, Rasmussen SA, Murphy DL, Cullen B, Valle D, Hoehn-Saric R, Greenberg BD, Pinto A, Knowles JA, Piacentini J, Pauls DL, Liang KY, Willour VL, et al. Sapap3 and pathological grooming in humans: Results from the OCD collaborative genetics study. Am J Med Genet B Neuropsychiatr Genet. 2009;150B:710–720. doi: 10.1002/ajmg.b.30897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borsini F, Podhorna J, Marazziti D. Do animal models of anxiety predict anxiolytic-like effects of antidepressants? Psychopharmacology (Berl) 2002;163:121–141. doi: 10.1007/s00213-002-1155-6. [DOI] [PubMed] [Google Scholar]
- Boucard AA, Chubykin AA, Comoletti D, Taylor P, Südhof TC. A splice code for trans-synaptic cell adhesion mediated by binding of neuroligin 1 to alpha- and beta-neurexins. Neuron. 2005;48:229–236. doi: 10.1016/j.neuron.2005.08.026. [DOI] [PubMed] [Google Scholar]
- Broekkamp CL, Rijk HW, Joly-Gelouin D, Lloyd KL. Major tranquillizers can be distinguished from minor tranquillizers on the basis of effects on marble burying and swim-induced grooming in mice. Eur J Pharmacol. 1986;126:223–229. doi: 10.1016/0014-2999(86)90051-8. [DOI] [PubMed] [Google Scholar]
- Budreck EC, Scheiffele P. Neuroligin-3 is a neuronal adhesion protein at GABAergic and glutamatergic synapses. Eur J Neurosci. 2007;26:1738–1748. doi: 10.1111/j.1460-9568.2007.05842.x. [DOI] [PubMed] [Google Scholar]
- Chih B, Afridi SK, Clark L, Scheiffele P. Disorder-associated mutations lead to functional inactivation of neuroligins. Hum Mol Genet. 2004;13:1471–1477. doi: 10.1093/hmg/ddh158. [DOI] [PubMed] [Google Scholar]
- Chih B, Engelman H, Scheiffele P. Control of excitatory and inhibitory synapse formation by neuroligins. Science. 2005;307:1324–1328. doi: 10.1126/science.1107470. [DOI] [PubMed] [Google Scholar]
- Chubykin AA, Liu X, Comoletti D, Tsigelny I, Taylor P, Südhof TC. Dissection of synapse induction by neuroligins: effect of a neuroligin mutation associated with autism. J Biol Chem. 2005;280:22365–22374. doi: 10.1074/jbc.M410723200. [DOI] [PubMed] [Google Scholar]
- Chubykin AA, Atasoy D, Etherton MR, Brose N, Kavalali ET, Gibson JR, Südhof TC. Activity-dependent validation of excitatory versus inhibitory synapses by neuroligin-1 versus neuroligin-2. Neuron. 2007;54:919–931. doi: 10.1016/j.neuron.2007.05.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung HJ, Huang YH, Lau LF, Huganir RL. Regulation of the NMDA receptor complex and trafficking by activity-dependent phosphorylation of the NR2B subunit PDZ ligand. J Neurosci. 2004;24:10248–10259. doi: 10.1523/JNEUROSCI.0546-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Comoletti D, De Jaco A, Jennings LL, Flynn RE, Gaietta G, Tsigelny I, Ellisman MH, Taylor P. The Arg451Cys-neuroligin-3 mutation associated with autism reveals a defect in protein processing. J Neurosci. 2004;24:4889–4893. doi: 10.1523/JNEUROSCI.0468-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crawley JN. Mouse behavioral assays relevant to the symptoms of autism. Brain Pathol. 2007;17:448–459. doi: 10.1111/j.1750-3639.2007.00096.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cromwell HC, Berridge KC. Implementation of action sequences by a neostriatal site: a lesion mapping study of grooming syntax. J Neurosci. 1996;16:3444–3458. doi: 10.1523/JNEUROSCI.16-10-03444.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dahlhaus R, Hines RM, Eadie BD, Kannangara TS, Hines DJ, Brown CE, Christie BR, El-Husseini A. Overexpression of the cell adhesion protein neuroligin-1 induces learning deficits and impairs synaptic plasticity by altering the ratio of excitation to inhibition in the hippocampus. Hippocampus. 2010;20:305–322. doi: 10.1002/hipo.20630. [DOI] [PubMed] [Google Scholar]
- Deacon RM. Digging and marble burying in mice: simple methods for in vivo identification of biological impacts. Nat Protoc. 2006;1:122–124. doi: 10.1038/nprot.2006.20. [DOI] [PubMed] [Google Scholar]
- Dean C, Scholl FG, Choih J, DeMaria S, Berger J, Isacoff E, Scheiffele P. Neurexin mediates the assembly of presynaptic terminals. Nat Neurosci. 2003;6:708–716. doi: 10.1038/nn1074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding J, Peterson JD, Surmeier DJ. Corticostriatal and thalamostriatal synapses have distinctive properties. J Neurosci. 2008;28:6483–6492. doi: 10.1523/JNEUROSCI.0435-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durand CM, Betancur C, Boeckers TM, Bockmann J, Chaste P, Fauchereau F, Nygren G, Rastam M, Gillberg IC, Anckarsäter H, Sponheim E, Goubran-Botros H, Delorme R, Chabane N, Mouren-Simeoni MC, de Mas P, Bieth E, Rogé B, Héron D, Burglen L, Gillberg C, Leboyer M, Bourgeron T. Mutations in the gene encoding the synaptic scaffolding protein SHANK3 are associated with autism spectrum disorders. Nat Genet. 2007;39:25–27. doi: 10.1038/ng1933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etherton MR, Blaiss CA, Powell CM, Südhof TC. Mouse neurexin-1alpha deletion causes correlated electrophysiological and behavioral changes consistent with cognitive impairments. Proc Natl Acad Sci U S A. 2009;106:17998–18003. doi: 10.1073/pnas.0910297106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feng J, Schroer R, Yan J, Song W, Yang C, Bockholt A, Cook EH, Jr, Skinner C, Schwartz CE, Sommer SS. High frequency of neurexin 1beta signal peptide structural variants in patients with autism. Neurosci Lett. 2006;409:10–13. doi: 10.1016/j.neulet.2006.08.017. [DOI] [PubMed] [Google Scholar]
- Ferreira SH, Lorenzetti BB. Glutamate spinal retrograde sensitization of primary sensory neurons associated with nociception. Neuropharmacology. 1994;33:1479–1485. doi: 10.1016/0028-3908(94)90052-3. [DOI] [PubMed] [Google Scholar]
- Flood JF, Morley JE, Lanthorn TH. Effect on memory processing by d-cycloserine, an agonist of the NMDA/glycine receptor. Eur J Pharmacol. 1992;221:249–254. doi: 10.1016/0014-2999(92)90709-d. [DOI] [PubMed] [Google Scholar]
- Gauthier J, Bonnel A, St-Onge J, Karemera L, Laurent S, Mottron L, Fombonne E, Joober R, Rouleau GA. NLGN3/NLGN4 gene mutations are not responsible for autism in the Quebec population. Am J Med Genet B Neuropsychiatr Genet. 2005;132B:74–75. doi: 10.1002/ajmg.b.30066. [DOI] [PubMed] [Google Scholar]
- Gerrow K, Romorini S, Nabi SM, Colicos MA, Sala C, El-Husseini A. A preformed complex of postsynaptic proteins is involved in excitatory synapse development. Neuron. 2006;49:547–562. doi: 10.1016/j.neuron.2006.01.015. [DOI] [PubMed] [Google Scholar]
- Glessner JT, Wang K, Cai G, Korvatska O, Kim CE, Wood S, Zhang H, Estes A, Brune CW, Bradfield JP, Imielinski M, Frackelton EC, Reichert J, Crawford EL, Munson J, Sleiman PM, Chiavacci R, Annaiah K, Thomas K, Hou C, Glaberson W, et al. Autism genome-wide copy number variation reveals ubiquitin and neuronal genes. Nature. 2009;459:569–573. doi: 10.1038/nature07953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graf ER, Zhang X, Jin SX, Linhoff MW, Craig AM. Neurexins induce differentiation of GABA and glutamate postsynaptic specializations via neuroligins. Cell. 2004;119:1013–1026. doi: 10.1016/j.cell.2004.11.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ichtchenko K, Hata Y, Nguyen T, Ullrich B, Missler M, Moomaw C, Südhof TC. Neuroligin 1: a splice site-specific ligand for beta-neurexins. Cell. 1995;81:435–443. doi: 10.1016/0092-8674(95)90396-8. [DOI] [PubMed] [Google Scholar]
- Ichtchenko K, Nguyen T, Südhof TC. Structures, alternative splicing, and neurexin binding of multiple neuroligins. J Biol Chem. 1996;271:2676–2682. doi: 10.1074/jbc.271.5.2676. [DOI] [PubMed] [Google Scholar]
- Irie M, Hata Y, Takeuchi M, Ichtchenko K, Toyoda A, Hirao K, Takai Y, Rosahl TW, Südhof TC. Binding of neuroligins to PSD-95. Science. 1997;277:1511–1515. doi: 10.1126/science.277.5331.1511. [DOI] [PubMed] [Google Scholar]
- Jamain S, Quach H, Betancur C, Råstam M, Colineaux C, Gillberg IC, Soderstrom H, Giros B, Leboyer M, Gillberg C, Bourgeron T. Mutations of the X-linked genes encoding neuroligins NLGN3 and NLGN4 are associated with autism. Nat Genet. 2003;34:27–29. doi: 10.1038/ng1136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim HG, Kishikawa S, Higgins AW, Seong IS, Donovan DJ, Shen Y, Lally E, Weiss LA, Najm J, Kutsche K, Descartes M, Holt L, Braddock S, Troxell R, Kaplan L, Volkmar F, Klin A, Tsatsanis K, Harris DJ, Noens I, et al. Disruption of neurexin 1 associated with autism spectrum disorder. Am J Hum Genet. 2008a;82:199–207. doi: 10.1016/j.ajhg.2007.09.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim J, Jung SY, Lee YK, Park S, Choi JS, Lee CJ, Kim HS, Choi YB, Scheiffele P, Bailey CH, Kandel ER, Kim JH. Neuroligin-1 is required for normal expression of LTP and associative fear memory in the amygdala of adult animals. Proc Natl Acad Sci U S A. 2008b;105:9087–9092. doi: 10.1073/pnas.0803448105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Konstantareas MM, Homatidis S. Chromosomal abnormalities in a series of children with autistic disorder. J Autism Dev Disord. 1999;29:275–285. doi: 10.1023/a:1022155201662. [DOI] [PubMed] [Google Scholar]
- Krishnan V, Han MH, Graham DL, Berton O, Renthal W, Russo SJ, Laplant Q, Graham A, Lutter M, Lagace DC, Ghose S, Reister R, Tannous P, Green TA, Neve RL, Chakravarty S, Kumar A, Eisch AJ, Self DW, Lee FS, et al. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell. 2007;131:391–404. doi: 10.1016/j.cell.2007.09.018. [DOI] [PubMed] [Google Scholar]
- Kwon CH, Luikart BW, Powell CM, Zhou J, Matheny SA, Zhang W, Li Y, Baker SJ, Parada LF. Pten regulates neuronal arborization and social interaction in mice. Neuron. 2006;50:377–388. doi: 10.1016/j.neuron.2006.03.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laumonnier F, Bonnet-Brilhault F, Gomot M, Blanc R, David A, Moizard MP, Raynaud M, Ronce N, Lemonnier E, Calvas P, Laudier B, Chelly J, Fryns JP, Ropers HH, Hamel BC, Andres C, Barthélémy C, Moraine C, Briault S. X-linked mental retardation and autism are associated with a mutation in the NLGN4 gene, a member of the neuroligin family. Am J Hum Genet. 2004;74:552–557. doi: 10.1086/382137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levinson JN, Chéry N, Huang K, Wong TP, Gerrow K, Kang R, Prange O, Wang YT, El-Husseini A. Neuroligins mediate excitatory and inhibitory synapse formation: involvement of PSD-95 and neurexin-1beta in neuroligin-induced synaptic specificity. J Biol Chem. 2005;280:17312–17319. doi: 10.1074/jbc.M413812200. [DOI] [PubMed] [Google Scholar]
- Lijam N, Paylor R, McDonald MP, Crawley JN, Deng CX, Herrup K, Stevens KE, Maccaferri G, McBain CJ, Sussman DJ, Wynshaw-Boris A. Social interaction and sensorimotor gating abnormalities in mice lacking Dvl1. Cell. 1997;90:895–905. doi: 10.1016/s0092-8674(00)80354-2. [DOI] [PubMed] [Google Scholar]
- Lin Y, Jover-Mengual T, Wong J, Bennett MV, Zukin RS. PSD-95 and PKC converge in regulating NMDA receptor trafficking and gating. Proc Natl Acad Sci U S A. 2006;103:19902–19907. doi: 10.1073/pnas.0609924104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lutter M, Sakata I, Osborne-Lawrence S, Rovinsky SA, Anderson JG, Jung S, Birnbaum S, Yanagisawa M, Elmquist JK, Nestler EJ, Zigman JM. The orexigenic hormone ghrelin defends against depressive symptoms of chronic stress. Nat Neurosci. 2008;11:752–753. doi: 10.1038/nn.2139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moretti P, Bouwknecht JA, Teague R, Paylor R, Zoghbi HY. Abnormalities of social interactions and home-cage behavior in a mouse model of Rett syndrome. Hum Mol Genet. 2005;14:205–220. doi: 10.1093/hmg/ddi016. [DOI] [PubMed] [Google Scholar]
- Moy SS, Nadler JJ, Perez A, Barbaro RP, Johns JM, Magnuson TR, Piven J, Crawley JN. Sociability and preference for social novelty in five inbred strains: an approach to assess autistic-like behavior in mice. Genes Brain Behav. 2004;3:287–302. doi: 10.1111/j.1601-1848.2004.00076.x. [DOI] [PubMed] [Google Scholar]
- Moy SS, Nadler JJ, Magnuson TR, Crawley JN. Mouse models of autism spectrum disorders: the challenge for behavioral genetics. Am J Med Genet C Semin Med Genet. 2006;142C:40–51. doi: 10.1002/ajmg.c.30081. [DOI] [PubMed] [Google Scholar]
- Myme CI, Sugino K, Turrigiano GG, Nelson SB. The NMDA-to-AMPA ratio at synapses onto layer 2/3 pyramidal neurons is conserved across prefrontal and visual cortices. J Neurophysiol. 2003;90:771–779. doi: 10.1152/jn.00070.2003. [DOI] [PubMed] [Google Scholar]
- Nadler JJ, Moy SS, Dold G, Trang D, Simmons N, Perez A, Young NB, Barbaro RP, Piven J, Magnuson TR, Crawley JN. Automated apparatus for quantitation of social approach behaviors in mice. Genes Brain Behav. 2004;3:303–314. doi: 10.1111/j.1601-183X.2004.00071.x. [DOI] [PubMed] [Google Scholar]
- Nam CI, Chen L. Postsynaptic assembly induced by neurexin-neuroligin interaction and neurotransmitter. Proc Natl Acad Sci U S A. 2005;102:6137–6142. doi: 10.1073/pnas.0502038102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newpher TM, Ehlers MD. Glutamate receptor dynamics in dendritic microdomains. Neuron. 2008;58:472–497. doi: 10.1016/j.neuron.2008.04.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Njung'e K, Handley SL. Evaluation of marble-burying behavior as a model of anxiety. Pharmacol Biochem Behav. 1991;38:63–67. doi: 10.1016/0091-3057(91)90590-x. [DOI] [PubMed] [Google Scholar]
- Posey DJ, Kem DL, Swiezy NB, Sweeten TL, Wiegand RE, McDougle CJ. A pilot study of d-cycloserine in subjects with autistic disorder. Am J Psychiatry. 2004;161:2115–2117. doi: 10.1176/appi.ajp.161.11.2115. [DOI] [PubMed] [Google Scholar]
- Powell CM, Schoch S, Monteggia L, Barrot M, Matos MF, Feldmann N, Südhof TC, Nestler EJ. The presynaptic active zone protein RIM1alpha is critical for normal learning and memory. Neuron. 2004;42:143–153. doi: 10.1016/s0896-6273(04)00146-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prange O, Wong TP, Gerrow K, Wang YT, El-Husseini A. A balance between excitatory and inhibitory synapses is controlled by PSD-95 and neuroligin. Proc Natl Acad Sci U S A. 2004;101:13915–13920. doi: 10.1073/pnas.0405939101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheiffele P, Fan J, Choih J, Fetter R, Serafini T. Neuroligin expressed in nonneuronal cells triggers presynaptic development in contacting axons. Cell. 2000;101:657–669. doi: 10.1016/s0092-8674(00)80877-6. [DOI] [PubMed] [Google Scholar]
- Song JY, Ichtchenko K, Südhof TC, Brose N. Neuroligin 1 is a postsynaptic cell-adhesion molecule of excitatory synapses. Proc Natl Acad Sci U S A. 1999;96:1100–1105. doi: 10.1073/pnas.96.3.1100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Südhof TC. Neuroligins and neurexins link synaptic function to cognitive disease. Nature. 2008;455:903–911. doi: 10.1038/nature07456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szatmari P, Paterson AD, Zwaigenbaum L, Roberts W, Brian J, Liu XQ, Vincent JB, Skaug JL, Thompson AP, Senman L, Feuk L, Qian C, Bryson SE, Jones MB, Marshall CR, Scherer SW, Vieland VJ, Bartlett C, Mangin LV, Goedken R. Mapping autism risk loci using genetic linkage and chromosomal rearrangements. Nat Genet. 2007;39:319–328. doi: 10.1038/ng1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabuchi K, Blundell J, Etherton MR, Hammer RE, Liu X, Powell CM, Südhof TC. A neuroligin-3 mutation implicated in autism increases inhibitory synaptic transmission in mice. Science. 2007;318:71–76. doi: 10.1126/science.1146221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talebizadeh Z, Bittel DC, Veatch OJ, Butler MG, Takahashi TN, Miles JH. Do known mutations in neuroligin genes (NLGN3 and NLGN4) cause autism? J Autism Dev Disord. 2004;34:735–736. doi: 10.1007/s10803-004-5295-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan-No K, Taira A, Wako K, Niijima F, Nakagawasai O, Tadano T, Sakurada C, Sakurada T, Kisara K. Intrathecally administered spermine produces the scratching, biting and licking behaviour in mice. Pain. 2000;86:55–61. doi: 10.1016/s0304-3959(99)00312-7. [DOI] [PubMed] [Google Scholar]
- Thomas A, Burant A, Bui N, Graham D, Yuva-Paylor LA, Paylor R. Marble burying reflects a repetitive and perseverative behavior more than novelty-induced anxiety. Psychopharmacology (Berl) 2009;204:361–373. doi: 10.1007/s00213-009-1466-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsankova NM, Berton O, Renthal W, Kumar A, Neve RL, Nestler EJ. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat Neurosci. 2006;9:519–525. doi: 10.1038/nn1659. [DOI] [PubMed] [Google Scholar]
- Varoqueaux F, Jamain S, Brose N. Neuroligin 2 is exclusively localized to inhibitory synapses. Eur J Cell Biol. 2004;83:449–456. doi: 10.1078/0171-9335-00410. [DOI] [PubMed] [Google Scholar]
- Varoqueaux F, Aramuni G, Rawson RL, Mohrmann R, Missler M, Gottmann K, Zhang W, Südhof TC, Brose N. Neuroligins determine synapse maturation and function. Neuron. 2006;51:741–754. doi: 10.1016/j.neuron.2006.09.003. [DOI] [PubMed] [Google Scholar]
- Vincent JB, Kolozsvari D, Roberts WS, Bolton PF, Gurling HM, Scherer SW. Mutation screening of X-chromosomal neuroligin genes: no mutations in 196 autism probands. Am J Med Genet B Neuropsychiatr Genet. 2004;129B:82–84. doi: 10.1002/ajmg.b.30069. [DOI] [PubMed] [Google Scholar]
- Volk LJ, Pfeiffer BE, Gibson JR, Huber KM. Multiple Gq-coupled receptors converge on a common protein synthesis-dependent long-term depression that is affected in fragile X syndrome mental retardation. J Neurosci. 2007;27:11624–11634. doi: 10.1523/JNEUROSCI.2266-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welch JM, Lu J, Rodriguiz RM, Trotta NC, Peca J, Ding JD, Feliciano C, Chen M, Adams JP, Luo J, Dudek SM, Weinberg RJ, Calakos N, Wetsel WC, Feng G. Cortico-striatal synaptic defects and OCD-like behaviours in Sapap3-mutant mice. Nature. 2007;448:894–900. doi: 10.1038/nature06104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan J, Oliveira G, Coutinho A, Yang C, Feng J, Katz C, Sram J, Bockholt A, Jones IR, Craddock N, Cook EH, Jr, Vicente A, Sommer SS. Analysis of the neuroligin 3 and 4 genes in autism and other neuropsychiatric patients. Mol Psychiatry. 2005;10:329–332. doi: 10.1038/sj.mp.4001629. [DOI] [PubMed] [Google Scholar]
- Yan J, Feng J, Schroer R, Li W, Skinner C, Schwartz CE, Cook EH, Jr, Sommer SS. Analysis of the neuroligin 4Y gene in patients with autism. Psychiatr Genet. 2008a;18:204–207. doi: 10.1097/YPG.0b013e3282fb7fe6. [DOI] [PubMed] [Google Scholar]
- Yan J, Noltner K, Feng J, Li W, Schroer R, Skinner C, Zeng W, Schwartz CE, Sommer SS. Neurexin 1alpha structural variants associated with autism. Neurosci Lett. 2008b;438:368–370. doi: 10.1016/j.neulet.2008.04.074. [DOI] [PubMed] [Google Scholar]
- Yan QJ, Asafo-Adjei PK, Arnold HM, Brown RE, Bauchwitz RP. A phenotypic and molecular characterization of the fmr1-tm1Cgr fragile X mouse. Genes Brain Behav. 2004;3:337–359. doi: 10.1111/j.1601-183X.2004.00087.x. [DOI] [PubMed] [Google Scholar]
- Ylisaukko-oja T, Rehnström K, Auranen M, Vanhala R, Alen R, Kempas E, Ellonen P, Turunen JA, Makkonen I, Riikonen R, Nieminen-von Wendt T, von Wendt L, Peltonen L, Järvelä I. Analysis of four neuroligin genes as candidates for autism. Eur J Hum Genet. 2005;13:1285–1292. doi: 10.1038/sj.ejhg.5201474. [DOI] [PubMed] [Google Scholar]
- Zlomuzica A, De Souza Silva MA, Huston JP, Dere E. NMDA receptor modulation by D-cycloserine promotes episodic-like memory in mice. Psychopharmacology (Berl) 2007;193:503–509. doi: 10.1007/s00213-007-0816-x. [DOI] [PubMed] [Google Scholar]
- Zoghbi HY. Postnatal neurodevelopmental disorders: meeting at the synapse? Science. 2003;302:826–830. doi: 10.1126/science.1089071. [DOI] [PubMed] [Google Scholar]
- Züchner S, Wendland JR, Ashley-Koch AE, Collins AL, Tran-Viet KN, Quinn K, Timpano KC, Cuccaro ML, Pericak-Vance MA, Steffens DC, Krishnan KR, Feng G, Murphy DL. Multiple rare SAPAP3 missense variants in trichotillomania and OCD. Mol Psychiatry. 2009;14:6–9. doi: 10.1038/mp.2008.83. [DOI] [PMC free article] [PubMed] [Google Scholar]